
Abstract
MAIT cells are abundant, highly evolutionarily conserved innate-like lymphocytes expressing a semi-invariant T cell receptor (TCR), which recognizes microbially derived small intermediate molecules from the riboflavin biosynthetic pathway. However, in addition to their TCR-mediated functions they can also be activated in a TCR-independent manner via cytokines including IL-12, -15, -18, and type I interferon. Emerging data suggest that they are expanded and activated by a range of viral infections, and significantly that they can contribute to a protective anti-viral response. Here we describe methods used to investigate these anti-viral functions in vivo in murine models. To overcome the technical challenge that MAIT cells are rare in specific pathogen-free laboratory mice, we describe how pulmonary MAIT cells can be expanded using intranasal bacterial infection or a combination of synthetic MAIT cell antigen and TLR agonists. We also describe protocols for adoptive transfer of MAIT cells, methods for lung homogenization for plaque assays, and surface and intracellular cytokine staining to determine MAIT cell activation.
You have full access to this open access chapter, Download protocol PDF
Similar content being viewed by others


Key words
1 Introduction
MAIT cells are relatively recently described innate-like lymphocytes, with similarities to the invariant natural killer T (iNKT) and γδ T cell subsets [1,2,3,4]. They are the most abundant innate-like population in the lungs in humans [5] though relatively rare in specific pathogen-free mice [6] and show a striking evolutionary conservation between diverse species of mammals [7]. MAIT cells express a semi-invariant T cell receptor (TCR) , which recognizes microbially derived small molecule intermediates from the riboflavin biosynthetic pathway [1, 4, 8, 9]. These molecular intermediates exist only in microbes but not in mammals, and therefore constitute a signature of microbial infection. This property implicates MAIT cells in anti-bacterial host defense, and potentially also in other roles such as tissue repair [3]. However, in addition to their TCR-dependent functions, they can be activated in a TCR-independent manner via cytokines including IL-12, -15, -18, and type I interferon [10,11,12]. Emerging data suggest that they are expanded and activated by a range of human viral infections including dengue, hepatitis C, and influenza virus [11, 13]. It was not clear from observational human studies whether this would lead to enhanced immune protection, or, conversely, contribute to immunopathology. To address this question, we conducted experimental influenza A virus challenge in vivo in mice and demonstrated that MAIT cells could contribute to a protective anti-viral response [12].
Here we describe the methods used to investigate these anti-viral functions in vivo in murine models. To overcome the technical challenge that MAIT cells are rare in specific pathogen-free laboratory mice, we describe (1) how pulmonary MAIT cells can be expanded using intranasal (i.n.) bacterial infection or a combination of synthetic MAIT cell antigen and TLR agonists as well as protocols for (2) adoptive transfer of MAIT cells, (3) viral preparation and infection of mice, (4) lung homogenization, (5) surface and intracellular cytokine staining to determine MAIT cell activation, and (6) plaque assays.
2 Materials
2.1 Reagents and Buffers
- 1.
-
2.
Collagenase medium: Roswell Park Memorial Institute medium (RPMI) containing 3 mg/mL collagenase III, 5 μg/mL DNase, and 2% fetal calf serum (FCS). Aliquots can be frozen at −20 °C.
-
3.
Fluorescence activated flow cytometry (FACS ) buffer: phosphate buffered saline (PBS), 2 mM EDTA, 0.5% bovine serum albumin (BSA). From a 500 mL bottle of PBS, add 40 mL to a 50 mL falcon containing 2.5 g BSA powder, vortex hard, then filter sterilize back into PBS bottle using a syringe through a 0.22-μm filter. Do not add azide as will be toxic to the cells.
-
4.
Percoll (Density 1.13 g/mL) 40% and 70% solutions, pre-warmed to room temperature for each use.
-
5.
RPMI with pen/strep: RPMI containing 100 μg/mL streptomycin and 100 U/mL penicillin.
-
6.
Tris-based Ammonium Chloride (TAC)–HCl, pH 7.5 hypotonic red blood cell lysis buffer: 0.14 M NH4Cl, 0.017 mM Tris (pH 7.5), then adjust pH to 7.2 with HCl (2 M). The solution is filter (0.22 μm) sterilized and kept at room temperature.
-
7.
Fixation buffer: 1% formaldehyde, 2% glucose in PBS. Fully dissolved solution is kept cold (+4 °C) and dark (aluminum foil wrapped) as formaldehyde is sensitive to light.
-
8.
Media for growing MDCK cells: Dulbecco Modified Eagle Medium (DMEM) containing 2 mM l-glutamine, 1 mM MEM sodium pyruvate, 100 U/mL penicillin/streptomycin, and 10% heat-inactivated FCS.
-
9.
Serum-free (SF) DMEM: Dulbecco Modified Eagle Medium (DMEM) containing 2 mM l-glutamine, 1 mM MEM sodium pyruvate, and 100 U/mL penicillin/streptomycin.
-
10.
2× Leibovitz’s L-15 media for overlay, make 2× stock as it will be diluted 1:1 with agarose. For 1 L: Use 1 L sterile water. Remove 100 mL of the water but keep for later use. Add two 14 g packets of L-15 powdered media (kept at 40 °C). Add magnetic flea and stir for 4 h or more to ensure the powder is completely dissolved. Adjust pH to 6.8 using 1 M HCl. Then add the following to the medium.
-
(a)
8 mL of 7% w/v NaHCO3 prepared in Hanks Buffered Saline Solution (HBSS) (stored at 4 °C).
-
(b)
800 μL 1 M HEPES buffer (pH 6.8).
-
(c)
20 mL of 10,000 U/mL Pen/Strep.
-
(d)
Make up the volume to 1 L (using the 100 mL previously removed) and filter sterilize. Store at 4 °C. To reduce precipitation, aliquot into 50 mL tubes for storage.
-
(a)
-
11.
1 mg/mL trypsin: warm up trypsin powder for 30 min at room temperature (kept at +4 °C). Weigh out 10 mg of powder and dissolve in 10 mL of PBS. Filter using 0.45 μm filter. Aliquot aseptically into 220 μL/aliquot. Store at −20 °C. 200 μL will be added to 100 mL of overlay (50 mL L-15 and 50 mL agarose) for a final concentration of 2 μg/mL trypsin/well.
-
12.
Salmonella: Salmonella enterica, serovar Typhimurium (attenuated strain BRD509) [14], stored at −20 °C in Luria-Bertani (LB) broth with 50% glycerol, to prevent freezing at this temperature.
-
13.
MR1-tetramers (5-OP-RU and 6-FP) are available from the NIH core tetramer facility, on application. Store in component parts at −80 °C until ready for use, at which point small aliquots can be tetramerized and stored at 4 °C for days to weeks. They should be reconstituted according to instructions supplied with the product. Typically a 5-μg aliquot of MR1-5-OP-RU monomer or MR1-6-FP monomer should be expanded to a total volume of 18 μL in Tris-buffered saline. About 6.8 μL of commercially available streptavidin-PE at 0.5 mg/mL should be made up to a total volume of 17 μL in Tris-buffered saline. Add 1/10 of the streptavidin-PE solution (1.7 μL) to the monomer solution every 10 min and pipette to mix, incubating at room temperature in the dark between steps. Repeat until all the streptavidin-PE solution has been added. This will give a final volume of 35 μL containing 0.143 μg/μL of tetramer. The tetramer should be titrated for use; typically 1:200–1:1000 dilutions are sufficient.
-
14.
Madin-Darby Canine Kidney (MDCK) cells.
-
15.
Live/Dead Fixable Aqua Dead Cell Stain Kit or Zombie Yellow Viability Stain Kit.
-
16.
Brefeldin A.
-
17.
Phorbol 12-myristate 13-acetate (PMA).
-
18.
Ionomycin.
-
19.
Trypsin–versene.
-
20.
1% Crystal Violet in 20% ethanol and dH2O.
-
21.
Flow cytometry compensation beads.
-
22.
Flow cytometry 6 μm blank size calibration beads.
-
23.
Fixation/permeabilization buffer and perm-wash buffer.
-
24.
LB agar plates, containing 50 μg/mL streptomycin.
-
25.
LB culture medium.
-
26.
2.4G2 (anti CD16/32) hybridoma cell culture supernatant.
-
27.
Anti-CD4 (GK1.5) and anti-CD8 (53.762) monoclonal antibodies for depletion of adoptively transferred T cell subsets.
-
28.
1% Virkon or 10% Lysol or Hypochlorite (5000 ppm).
-
29.
80%w/v EtOH.
-
30.
Hanks buffered saline solution (HBSS).
-
31.
Isoflurane.
2.2 Plastic and Other Supplies
-
1.
1 and 10 mL syringes.
-
2.
26 G needles.
-
3.
Dissection scissors.
-
4.
1 mL Eppendorf tubes.
-
5.
40 and 70 μm cell strainers.
-
6.
10 cm Petri dishes.
-
7.
10, 15, and 50 mL Falcon tubes.
-
8.
5 mL polypropylene or polycarbonate FACS tubes.
-
9.
Flat-bottom 6-well (TC6) plates.
-
10.
96-well flat-bottom plates.
-
11.
96-well U- or V-bottom plates.
2.3 Equipment
-
1.
Flow cytometer with capability for cell sorting, BD LSR Aria or equivalent.
-
2.
Spectrophotometer capable of reading at 600 nm.
-
3.
Hemocytometer and light microscope.
-
4.
Animal anesthetic circuit capable of administering volatile inhalational anesthetics.
-
5.
Shaking incubator.
-
6.
Gaseous carbon dioxide and gas exposure chamber.
-
7.
Benchtop mechanical roller for tubes.
-
8.
Tissue homogenizer for disrupting tissue into single cell suspensions.
3 Methods
Personal protective equipment (PPE) should be worn at all times (gloves, lab coat, & eye protection) (see Notes 3 and 4).
3.1 MAIT Cell Expansion in Donor Mice
MAIT cells are rare in specific pathogen-free mice [6], typically comprising about 1 × 104 recoverable pulmonary MAIT cells in an infection-naïve adult C57BL/6 mouse. Therefore, for adoptive transfer experiments, the MAIT cell population should first be expanded using intranasal infection [15] or immunization (5-OP-RU with TLR agonists) [3, 15] (see Note 5). When planning the adoptive transfer experiment, estimate that one S. Typhimurium BRD509-infected mouse will yield 1–2 × 106 sorted MAIT cells, which are enough for 10–20 recipient mice (105 MAIT cells/RAG2−/−γC−/− mouse in this case). Infect donor mice 7 days earlier than the adoptive transfer.
-
1.
Two days before infection streak out a plate of S. Typhimurium BRD509 (an attenuated vaccine strain [14]) on LB agar plates, containing 50 μg/mL streptomycin and incubate plates overnight at 37 °C.
-
2.
The day before infection, pick a single colony under flame and inoculate to 10 mL LB culture medium with 50 μg/mL streptomycin and leave static at 37 °C (double contained if working with wild type/virulent SL1344 or equivalent strains) overnight.
-
3.
On the day of infection, re-inoculate into fresh 10 mL pre-warmed LB culture medium with 0.5, 100, or 20 μL of overnight culture, under flame. This is to ensure an optimal optical density (O.D.) reading (bacteria in log phase growth) for preparing the inoculum later (see Note 6). The doubling time for Salmonella can vary between 0.5 and 1 h. Make the infection inoculum from culture with O.D.600nm reading between 0.2 and 0.6. Calculate the required CFU of bacteria estimating 1 O.D. = 5–10 × 108 CFU (this constant needs to be established for individual labs). Dilute with PBS to a final concentration of 2 × 107 CFU/mL, allowing 50 μL inoculum/mouse, i.e., 106 CFU/50 μL/mouse.
-
4.
Infect mice i.n. with 106 CFU S. Typhimurium BRD509 in 50 μL PBS under isoflurane anesthesia (see Notes 7 and 8).
-
5.
Allow mice to recover and monitor mice for 7 days to allow the infection to take its course and MAIT cell frequencies to expand dramatically from 104 to 5 × 106 MAIT cells, or from 1% to 20–50% of all alpha-beta T cells [15] (see Note 8).
3.2 MAIT Cell Adoptive Transfer
-
1.
7 days or more after intranasal infection with S. Typhimurium, MAIT cells can be harvested (see Note 9). As MAIT cells are to be used for adoptive transfer, all procedures should be performed in a BSCII biosafety cabinet. All tools and reagents should be sterile.
-
2.
Prewarm collagenase media and shaking incubator to 37 °C.
-
3.
Mice should be euthanized (e.g., using a rising concentration of CO2 with a second method to confirm death).
-
4.
Open the diaphragm by cutting the rib cage to expose both the heart and lungs. Gently perfuse the right ventricle with 8–10 mL of ice-cold RPMI to dispense circulating blood. Perfuse using a 10-mL syringe and a 26-G needle. Efficient perfusion will result in lung inflation and a color change to pink/white.
-
5.
Remove lungs using scissors to cut through the hilum and place into a 24-well plate containing ice-cold RPMI to transfer organs to the laboratory.
-
6.
Chop lungs into fine pieces (see Note 10).
-
7.
Place lung tissue into a 1-mL Eppendorf tube containing 1–2 mL/lung of pre-warmed collagenase medium. Incubate tubes on their sides in a shaking incubator at 37 °C, at 100–180 rpm, for 90 min.
-
8.
During this time prepare Percoll gradients and antibody cocktails (see Table 1).
-
9.
After 90 min pour digested tissue through a 70-μm cell strainer and force through into a Petri dish with the plunger from a 1-mL syringe. Rinse residual sample with extra FACS buffer for maximum MAIT cell yield. Cells from multiple lungs (if required) (see Note 11) are pooled into a single 50-mL Falcon tube with a total of 50 mL of sterile FACS buffer.
-
10.
Centrifuge at 400 × g for 5 min to pellet the cells. Pour off supernatant (SN).
-
11.
Resuspend cells in 20 mL 40% Percoll. Underneath this layer use a transfer pipette to layer 20 mL 70% Percoll (see Note 12). Centrifuge this gradient at 800 × g for 20 min at room temperature with the centrifuge brake OFF. Lymphocytes and other immune cells will form a visible interphase layer between the 40% and 70% Percoll post centrifugation.
-
12.
During this centrifugation step, prepare single color controls. It is convenient to use part of a spleen forced through a 70-μm filter and resuspended in 5 mL TAC for 5 min at 37 °C, then washed once with 5 mL FACS buffer.
-
13.
Collect the interphase between 40% and 70% Percoll into a fresh 50 mL Falcon and top up with FACS buffer to a total of 50 mL. Centrifuge at 400 × g for 5 min.
-
14.
Pour off supernatant and resuspend in 5 mL FACS buffer, transferring to a 10 mL Falcon tube. Centrifuge at 400 × g for 5 min.
-
15.
Resuspend all lung cells in 750 μL FACS buffer.
-
16.
Block non-specific tetramer binding by adding 7.5 μL 2.4G2 (anti CD16/32) cell culture supernatant, containing MR1-6-FP tetramer [8, 16] (no fluorochrome, 1:100). Incubate at room temperature on a roller or bench rocker for 15 min.
-
17.
For lungs from 5 mice, add 750 μL of staining cocktail (Table 1).
-
18.
Cover in aluminum foil to protect fluorochromes from light and shake on roller for 30 min RT.
-
19.
Wash with 10 mL of FACS wash. Centrifuge at 400 × g for 5 min. Pour off supernatant.
-
20.
Wash again with 10 mL of FACS wash. Centrifuge at 400 × g for 5 min. Pour off supernatant.
-
21.
Resuspend cells in 2 mL FACS wash and filter through 40 μm filter into non-pyrogenic FACS tubes.
-
22.
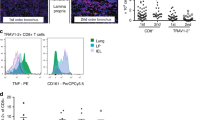
Sort live MAIT cells (defined as CD3+CD45+MR1-5-OP-RU tetramer+ cells) (Fig. 1) into 3 mL of FCS in 15 mL Falcon tube. For detailed gating strategy, refer to [17]. Wash cells and adjust cell concentration to 5 × 105 cell/mL, allowing 105 in 200 μL for injection to each mouse.
-
23.
Inject 105 cells into the tail vein of recipient mice using cells suspended in 200 μL PBS in a 1-mL syringe with a 26-G cannula after warming the mice for 5–15 min with appropriate monitoring.
-
24.
To deplete residual non-MAIT T cells (see Note 13), inject recipient mice on days 2 and days 5 or 6 with 0.1 mg each of purified anti-CD4 (GK1.5) and anti-CD8 (53.762) monoclonal antibodies i.v.
-
25.
Rest mice for a total of 2 weeks post adoptive transfer to allow MAIT cell populations to settle in the host.

Flow cytometry gating strategy for MR1-5-OP-RU-tetramer+ MAIT cells
3.3 Influenza A Virus Infection
-
1.
Thaw virus vial rapidly in a 37 °C water bath until all ice crystals have melted.
-
2.
Decontaminate the outer surface of the vial with 70% ethanol.
-
3.
Perform serial dilutions in sterile PBS to achieve the desired inoculum. For example:
-
(a)
If titer of PR8 stock = 1.0 × 109 plaque-forming units (PFU)/mL, require 25 PFU/inoculum (see Note 14).
-
(b)
Volume of inoculum required for intranasal infection = 50 μL/inoculum.
-
(c)
25 PFU/50 μL = 500 PFU/mL, dilution required from virus stock DF = 2,000,000.
-
Dilution 1: 1/100: 10 μL of virus stock +990 μL PBS.
-
Dilution 2: 1/100: 10 μL of dilution 1 + 990 μL PBS.
-
Dilution 3: 1/100: 10 μL of dilution 2 + 990 μL PBS.
-
Dilution 4: ½: (depends on volume required, e.g., 20 inoculations = 1000 μL) 500 μL of dilution 3 and 500 μL of PBS.
-
-
(a)
-
4.
Mix virus with vortex before administering the inoculum.
-
5.
Infect mice i.n. with 50 μL under isoflurane anesthesia (see Note 7).
-
6.
Allow mice to recover and monitor mice for until recovery (typically 10 days) (see Note 15).
3.4 Lung Homogenization
-
1.
Collect the lungs into 2 mL of RPMI supplemented with penicillin/streptomycin.
-
2.
For homogenization, place the lung and the 2 mL of media into 10 mL falcon tubes with lids (see Note 16).
-
3.
Prepare 10 or 15 mL Falcon tubes with 2 × 5 mL 80%w/v EtOH for cleaning the homogenization probe initially and 1 tube containing HBSS. For each group of samples, prepare further 1 × 5 mL EtOH and 1 × 5 mL HBSS, and for the final probe clean set up 2 × 5 mL of EtOH.
-
4.
Homogenize the sample using a homogenizer, mounted on a retort stand with the probe set to medium for 30 s per sample. Keep samples on ice (see Note 17).
-
5.
Centrifuge the samples at 1000 × g for 7 min.
-
6.
Using a 1 mL pipette, carefully draw up approximately 1 mL (a little bit more is good) of supernatant, avoiding the pellet and fatty residue on top. Divide this volume into two 1.5 mL Eppendorf tubes. Store at −80 °C for subsequent plaque assays.
3.5 MAIT Cell Intracellular Cytokine Staining
To analyze MAIT cell frequencies and function during viral infection.
-
1.
Prewarm collagenase media and shaking incubator to 37 °C.
-
2.
Mice should be euthanized (e.g., using a rising concentration of CO2 with a second method to confirm death).
-
3.
Open the diaphragm by cutting the rib cage to expose both the heart and lungs. Gently perfuse the right ventricle with 8–10 mL of ice-cold RPMI to remove circulating blood. Perfuse using a 10-mL syringe and a 26-G needle. Proper perfusion will result in lung inflation and a color change to pink/white.
-
4.
Remove lungs (see Note 18) using scissors to cut through the hilum and place into a 24-well plate containing ice-cold RPMI to transfer organs to the laboratory.
-
5.
Chop lungs into very small pieces (see Note 9).
-
6.
Place lung tissue into a 1-mL Eppendorf tube containing 0.5 mL/lung of pre-warmed collagenase/DNase medium. This should also contain, 0.5 μL of Brefeldin A (1:1000) (final concentration 3.0 μg/mL).
-
7.
Incubate tubes on their sides in a shaking incubator at 37 °C, at 100–180 rpm, for 90 min.
-
8.
After 90 min pour digested tissue through a 70-μm cell strainer and force through into Petri dish with the plunger from a 1-mL syringe. Rinse residual cells into a total of 10 mL FACS wash in 10 mL falcon tubes at RT. Centrifuge at 400 × g for 5 min at RT.
-
9.
Resuspend in 2 mL per lung (see Note 19) of pre-warmed TAC lysis buffer at 37 °C. Vortex well, then place in a pre-warmed water bath at 37 °C. After 5 min neutralize by adding an equal volume of FACS buffer. Centrifuge at 400 × g for 5 min, RT.
-
10.
Numbers of lung cells can be estimated using a hemocytometer or spectrophotometer (see Notes 20 and 21).
-
11.
Transfer 100 μL containing 0.5–1 million cells to a 96-well U- or V-bottom plate format or into FACS tubes for staining, passing them through a 40-μm mesh (see Note 22).
-
(a)
1 × 100 μL into a plate for surface stain (steps 16–22).
-
(b)
2 × 100 μL (unstimulated and stimulated) to a second plate (see Note 23), and include a no Brefeldin control (steps 12, 13, and 21–30).
In vitro stimulation phase:
-
(a)
-
12.
Keep the cells for the surface stain on ice, while setting up PMA /Ionomycin stimulation to induce production of cytokines of interest.
-
(a)
PMA final concentration: 20 ng/mL.
-
(b)
Ionomycin 1 μg/mL.
-
(c)
1000× stock Brefeldin A (final concentration 3.0 μg/mL).
-
(a)
-
13.
Incubate for 3 h at 37 °C with 5% CO2.
Surface staining:
-
14.
During stimulation phase perform surface staining for extracellular markers (see Note 24).
-
15.
If performing Zombie Yellow vital staining wash cells with 1–2 mL PBS. Centrifuge at 400 × g, 5 min (or if using plate format wash twice with 200 μL FACS buffer centrifuging for 2 min at 400 × g). Resuspend in 20 μL PBS + 0.4 μL Zombie Yellow for 15 min.
-
16.
Add 20 μL of 2.4G2 (anti-CD16/32) containing 0.2 μL of MR1-6-FP tetramer (no fluorochrome conjugate) to block non-specific binding. Incubate for 15 min dark, room temp.
-
17.
Add surface cocktail (Table 2) using a cocktail made up in 10 μL FACS buffer. Pipette carefully to mix. Stain for 20–30 min at room temperature.
-
18.
For single color controls use splenocytes or compensation beads.
-
19.
Wash cells twice with 2 mL FACS buffer, centrifuging at 400 × g for 5 min (or if using plate format wash three times with 200 μL FACS buffer centrifuging for 2 min at 400 × g).
-
20.
Resuspend cells in 100 μL FACS buffer (see Note 25). To enable estimation of absolute cell numbers, add a known number of calibration beads.
-
(a)
Vortex calibration beads hard. Dilute (1:10) counting beads in PBS before using. To each sample 25 μL of these diluted beads was added, and an additional 10 μL of beads were saved to be counted with a hemocytometer, giving a count of X in a large square, i.e., X × 104 beads/mL (which is X × 10 beads/μL, or X × 10 × 25 beads/sample). Typically add a total of 25,000 beads per sample.
-
(b)
When samples have been acquired on flow cytometer, these calibration beads can be detected using their FSC/SSC profile and the absolute number of cells of interest can be estimated using the following approach. Total number of MAIT cells per sample = Number of MAIT cells counted on flow cytometer × Number of beads added/Number of beads counted/proportion of total lung cell suspension actually used for staining.
Intracellular staining:
-
(a)
-
21.
After 3-h stimulation, continue processing the cells for intracellular staining. Resuspend into FACS tube with +1 mL PBS. Centrifuge at 400 × g, 5 min. (Alternatively, if in 96-well format resuspend in 100 μL PBS, centrifuge at 400 × g for 2 min and repeat.)
-
22.
Resuspend in 20 μL PBS with 0.4 μL Zombie Yellow for 15 min.
-
23.
Add 20 μL of 2.4G2 (anti CD16/32) SN containing 0.2 μL of unlabeled MR1-6-FP tetramer to block non-specific tetramer staining. Incubate for 15 min dark, room temperature.
-
24.
Add surface cocktail (Table 3) using a cocktail made up in 10 μL FACS buffer. Pipette carefully to mix. Stain for 20–30 min at room temperature.
-
25.
For single color controls use splenocytes or compensation beads. These may be available from being made up earlier in the protocol.
-
26.
Wash cells once with 1 mL FACS buffer, centrifuging at 400 × g for 5 min (or if using plate format wash twice with 200 μL FACS buffer centrifuging for 2 min at 400 × g).
-
27.
Resuspend in 200 μL of commercially available Fixation/Permeabilization solution and incubate for 30 min on ice.
-
28.
Wash with 2 mL Perm Wash (diluted 1:9 with FACS buffer). Centrifuge at 400 × g, 5 min (or if using plate format wash twice with 200 μL Perm Wash centrifuging for 2 min at 400 × g).
-
29.
Resuspend in 50 μL of Perm Wash containing intracellular cocktail (Table 4) and pipette carefully to mix. Incubate for 45 min or leave to stain overnight.
-
30.
Wash cells with 2 mL Perm Wash (or if using plate format wash twice with 200 μL Perm Wash centrifuging for 2 min at 400 × g). Resuspend cells in 100 μL FACS wash. If cells are in plate format use a multichannel pipette to transfer them to 1.2 mL “bullet” cluster tubes for acquisition, or use a plate reader attachment with the cytometer.
-
31.
Analyze cells on flow cytometer.
3.6 Viral Plaque Assay
Viral plaque assays are used to determine influenza viral titers. A diluted solution of egg-adapted Influenza A viruses/lung -infected tissue homogenates are applied to a six-well tissue culture dish containing a monolayer of Madin-Darby canine kidney (MDCK) cells. The infected MDCK cells grow under a semisolid overlay medium (agar) containing trypsin. A plaque is produced when a virus particle infects a cell, replicates, and then kills the cell. This process can be repeated several times as surrounding cells can be infected by newly replicated virus and killed. When visualized by eye, plaques appear as white spots. The assay is measured in PFU/mL.
Passaging MDCK cells:
-
1.
Warm up MDCK cell media, trypsin–versene, and PBS at 37 °C.
-
2.
Check the confluency of MDCK cells, aspirate the medium, add 10 mL of PBS, aspirate the medium, and repeat wash.
-
3.
Discard PBS, add 2–3 mL of trypsin–versene (stored −20 °C) to MDCK monolayers, and incubate at 37 °C for 5 min. After 5 min tap the flasks, and incubate for longer if required (maximum 15 min).
-
4.
In the meantime, add 15 mL of MDCK cell media to fresh T75 flasks.
-
5.
Add MDCK cell media to a total volume of 10 mL to the trypsinized cells, and transfer cells to a 10-mL tube.
-
6.
Count cells using a hemocytometer.
-
7.
Set up multiple T75 flasks with different cell densities to determine the growth pattern of MDCK cells. Generally ~3–5 × 105 for 3-day split.
-
8.
Incubate at 37 °C, 5% CO2.
Amplification of MDCK cells for plaque assay:
-
9.
Warm up MDCK cell media, trypsin–versene, and PBS at 37 °C.
-
10.
Check the confluency of MDCK cells, aspirate the medium, add 10 mL PBS, aspirate the medium, and repeat wash.
-
11.
Discard PBS, add 2–3 mL of trypsin–versene (stored −20 °C) to MDCK monolayers, and incubate at 37 °C for 5 min. After 5 min tap the flasks, and incubate for longer if required (maximum 15 min).
-
12.
In the meantime, add 40 mL of MDCK cell media to fresh T175 flasks. Set up one T175 flask of MDCK cells per ~4 plates for plaque assay. Each 6-well plate assays 3 viral dilutions (as dilutions are done in duplicate).
-
13.
Add MDCK cell media to a total volume of 10 mL to trypsinized cells, and transfer cells to a 10-mL tube.
-
14.
Count cells using a hemocytometer.
-
15.
Add ~7–8 × 105 cells per T175 flask for 3-day culture.
Seeding flat-bottomed 6-well tissue culture (TC6) plates for plaque assay:
-
16.
Warm up MDCK cell media, trypsin–versene, and PBS at 37 °C.
-
17.
Check the confluency of MDCK cells, aspirate the medium, add 10 mL of PBS, aspirate the medium, and repeat the wash.
-
18.
Discard PBS, add 5–8 mL of trypsin–versene (−20 °C) to MDCK monolayers in T175 flask, incubate at 37 °C for 5 min. After 5 min, tap bottles, incubate for longer if required (maximum 15 min).
-
19.
Add 17–20 mL of MDCK cell media to each flask (total 25 mL including 5–8 mL trypsin) to inhibit trypsin–versene.
-
20.
Pool cells into one flask.
-
21.
Count the cells, adjust the concentration to 3.3 × 105 cells/mL.
-
22.
Add 3 mL of 3.3 × 105 cells/mL to each well of TC6 plates (~1 × 106/well), swirl plates gently to distribute cells evenly. Include a negative control plate.
-
23.
Incubate cells at 37 °C, 5% CO2 overnight. Aim for monolayers to be confluent in 6-well plates for assay.
Plaque assay
-
24.
Warm up SF-DMEM at 37 °C.
-
25.
Prepare dilutions of samples to be titrated. This can be done in a 96-well flat-bottom plate.
-
26.
Cells will be infected with 150 μL of each dilution in duplicate, so a minimum of 300 μL of each dilution is required. A 96-well flat-bottom plate can hold ~350 μL/well. 35 μL of samples would be added to 315 μL of media for a final volume of 350 (35 in 350 = 1:10 dilution). If a sample is to be plaqued neat, add 350 μL of sample to the first well.
-
27.
Add 315 μL of SF-DMEM in each well of 96-well plate. A multichannel can be used and add 157 and 158 μL (total 315) to each well.
-
28.
Add 35 μL of sample to the first well with media (tenfold dilution) and continue serial tenfold dilutions by transferring 35 μL across wells, changing tips between dilutions.
-
29.
For titration of viral stocks use dilutions from 10−4 to 10−6. 10−1 can be used as a positive control. Half-log dilutions can also be performed. For titration of mouse lung homogenates, generally:
-
(a)
Days 1–5: 10−1 to 10−3.
-
(b)
Days 6–10: neat to 10−2 (see Note 26).
-
(a)
-
30.
Wash MDCK cells with 1–2 mL of PBS/well.
-
31.
Infect cells with 150 μL/well of the appropriate dilution, swirl gently to cover all cells and incubate at 37 °C, 5% CO2 for 60 min, shake gently every 15 min.
-
32.
In the meantime, prepare overlay media, best to start doing so in the beginning of the 1 h incubation of MDCK cells.
-
33.
Weigh out 1.8 g of agarose into 200 mL glass bottle, add 100 mL of sterile water (1.8% agarose), and melt in microwave. Store in a 55 °C water bath.
-
34.
Aliquot 50 mL of 2 × L-15 medium into 50 mL tubes (need 2 × 50 mL aliquots of 2 × L-15 medium and 1 bottle (100 mL) of 1.8% agarose) and store in 37 °C water bath.
-
35.
Thaw trypsin–versene at 40 °C. Thaw a 200 μL aliquot/50 mL of 2 × L-15.
-
36.
After 1 h incubation of MDCK cells with sample: Add 200 μL trypsin to each 50 mL tube of 2 × L-15.
-
37.
Make overlay media by adding 100 mL of 2 × L-15 + trypsin solution to 100 mL of 1.8% agarose and mix well (see Note 27).
-
38.
Add 3 mL of overlay medium/well and leave at RT until it sets.
-
39.
Incubate upside-down at 37 °C, 5% CO2 for 3 days. Plaques maybe visible by the end of day 2 and the plates can be incubated till day 4 if plaques are too small on day 3.
-
40.
Count the plaques. This can be done by holding the plates against the light. Alternatively, remove agarose overlay and stain with crystal violet. To stain, cover the cells with a minimal amount of crystal violet solution for ~15 min. Rock plates if necessary to ensure even coverage. Gently wash off the crystal violet stain with water. Once fixed, stained, and dried, store plaques indefinitely for future analysis.
-
41.
Calculate viral titer in PFU/mL: Average count of duplicate well × Dilution factor × (1000/150) = PFU/mL (multiply lung homogenate counts by 2 to give total viral load, as lungs were taken and homogenized in 2 mL RPMI).
4 Notes
-
1.
Allow a little extra for pipetting wastage when making up antibody cocktails. Keep on ice and protect from light (e.g., with aluminum foil). Make up cocktails in FACS buffer, but for the intracellular stains these should be made up in Perm Wash buffer containing 0.1% Saponin.
-
2.
Congenic markers could be reversed or other markers are used as appropriate to the mouse strains being used and to the specific experimental set-up.
-
3.
Biological Hazards—S. Typhimurium BRD509 is a risk group 2 pathogen. Influenza A virus-PR8-strain (H1N1) is a lab adapted strain of IAV virus. Work should be risk assessed and we recommend controls that include but are not restricted to the following: Lab coat, safety glasses, and gloves should be worn when performing this protocol. Gloves should be removed or sterilized before exiting the biohazard hood. Solutions of Lysol (200 ppm) or hypochlorite (5000 ppm) should be accessible in case of a spill.
-
4.
Decontaminate all pipette tips in 1% Virkon when working in the biohazard cabinet. After use, the biohazard hood should be decontaminated by wiping down with 70% ethanol and by UV sterilization for 15 min before any further use. All waste and its container must be disposed as hazardous waste.
-
5.
Pulmonary MAIT cells can be expanded using any source of 5-OP-RU and an appropriate TLR agonist [15, 17]. A systematic assessment of effective TLR agonists has shown strong MAIT cell expansion 7 days after intranasal inoculation with 76 pmol 5-OP-RU on days 1, 2, and 4 in combination with a single dose of agonist on day 1 to TLR3 (high molecular weight poly I:C), TLR4 (lipopolysaccharide from E. coli ), TLR2/6 (FSL-1 (Pam2CGDPKHPKSF)), or TLR9 (CpG ODN1826), but not with agonists of TLR1/2 (Pam3CSK4), TLR2, TLR5, TLR7 [3]. Each inoculum should be instilled in 50 μL PBS. However, the requirement for accurate repeated inoculations can introduce significant variability in MAIT cell expansion. A simple, less costly on reagents and time, and equally effective, if not more so, is a single intranasal inoculation with S. Typhimurium BRD509 in 50 μL PBS.
-
6.
Growth of bacteria is estimated by measuring the culture in a spectrophotometer at 600 nm. To do so fill a cuvette with fresh LB media, place in spectrophotometer, and use this to blank. Then take 500 μL of bacteria-containing broth and measure optical density. To calculate the inoculum dose, use the estimate that an O.D.600nm of 1 = 5 × 108 CFU/mL.
-
7.
Accurate intranasal inoculation depends critically on the depth of anesthesia. Administer isoflurane and observe breathing pattern until respiratory rate has decreased to approximately 100 breaths/min and is deep and relaxed. If insufficient depth is achieved mice will sneeze. If depth of anesthesia is too great (further slowing of respiratory rate and very deep breaths), then mice tend to spontaneously breath-hold and again, volume inhaled will be unreliable. Place 50 μL of inoculum onto the left nasal opening (if user is right-handed) using a P200 pipette, gradually ejecting the 50 μL over a few breath cycles until all has been inspired.
-
8.
Intranasal S. Typhimurium is well tolerated in immunocompetent strains such as C57BL/6 and BALB/c with less than 5% of animals showing minor signs of illness (ruffled hair) within 1–2 days after infection. These animals fully recover after days 3–5. The lethal dose of S. Typhimurium BRD509 is >2 × 107 CFU/mouse (wild-type C57BL/6 adult). Caution should be used in immunocompromised strains in which pilot experiments should be performed to confirm optimal safe inocula.
-
9.
This MAIT cell expansion is long-lived [15], so donor mice can be prepared several weeks in advance.
-
10.
The lungs can conveniently be chopped up using the back of an upturned Petri dish. Using fine forceps lift lungs from the RPMI in which they have been transferred, gently blot off excess liquid with tissue paper and place on the Petri dish. Use a large curved scalpel blade to repeatedly chop through the lungs at multiple angles for at least 60 s each until a very fine and homogeneous texture is achieved.
-
11.
Typically this method will yield 1.5 × 106 pulmonary MAIT cells per mouse, so multiple mice may be required as donors, depending on the requirements of the experiment.
-
12.
This will be sufficient for lungs from 8 mice.
-
13.
If transferring cells into a Rag2−/−γC−/− mouse then low frequencies of “contaminating” conventional CD4+ or CD8+ T cells tend to expand more rapidly than the MAIT cells and produce artifacts (not obvious for other T-cell-deficient mice, e.g., TCRα−/− or RAG2−/−). As many MAIT cells are double-negative, it is possible to prevent this effect by repeated injections with T-cell-depleting anti-CD4 and anti-CD8 antibodies [17].
-
14.
The PR8 strain of influenza virus is highly virulent in mice and only low inoculate are tolerated. The exact inoculum required for each experimental system will need to be carefully determined depending on the exact strain and batch of PR8 and the strain of mice, and local welfare and monitoring requirements. In our hands C57BL/6 mice receiving 100 PFU of A/PR/8/34 AF18 WCN experienced severe pneumonia in mice, characterized by parenchymal necrosis and infiltrates of macrophages, lymphocytes, and neutrophils, with 10–25% mortality due to welfare concerns or weight loss >20%.
-
15.
Virally infected mice experience a transient viral illness with transient. Viral titers peak at day 3. Weight loss peaks at day 5–7 post infection, and there would be a significant weight gain expected by day 8 and resolving by day 10 post infection. Typically mice should be monitored and/or weighed daily for signs of ill health such as ruffled fur, hunched-up appearance, gait abnormalities, lethargy and loss of body condition for 10 days after challenge or till all the symptoms disappear and body weight returns to pre-challenge level. Monitoring can then return to twice weekly.
-
16.
For many homogenization probes a wide tube is needed, such as the sterile, capped, round-bottom polypropylene tubes which are available.
-
17.
The homogenizer generates a lot of heat at the probe tip. Samples should be kept on ice before and after homogenization, and the probe should be intermittently rested to cool down in ice-cold EtOH between groups of 5 or 10 samples. Between samples or groups of samples clean the probe by running briefly in EtOH and then rinsing briefly in HBSS. Often connective tissue will clog the probe and this can be removed with large forceps. After use the probe tip should be sterilized.
-
18.
Only approximately 2/7 of one lung is needed for intracellular cytokine staining, so the other lung, or other sections of lung, can be saved for viral titer estimation, histology, or other assays if required.
-
19.
To clarify terminology there are two lungs in each animal, so “one lung” refers to all the 2 or 3 lobes in a single hemithorax. Due to the presence of the heart on the left side, the left lung is smaller with only 2 lobes.
-
20.
Using a spectrophotometer saves time for large numbers of samples. To do this resuspend cell pellet in 1–2 mL PBS (or adjust according to pellet size/counts). Select O.D.600nm. Blank cuvette with 1 mL FACS wash/PBS. Measure O.D.600nm with 200 μL samples + 800 μL PBS (5×). Calculate the number of cells: this is a simple linear relationship between O.D. and the number of cells, which can be derived by measuring a few cell counts in parallel on both the hemocytometer and the spectrophotometer.
-
21.
An alternative is to resuspend the entire pellet in 700 μL of FACS buffer and take 200 μL into 96-well plate: this should contain approximately 1–1.5 × 106 cells, appropriate for staining.
-
22.
To avoid using multiple filters, it is possible to buy large sheets of 40 μm mesh. A single rectangle can be cut which covers a whole plate. Using this, multiple cells can be pipette simultaneously with a multichannel pipette.
-
23.
In round-bottom plates cells may clump so consider using flat-bottom plate for the stimulation step, especially if doing further steps in FACS tubes rather than staining in plate format.
-
24.
While surface markers can be measured on the intracellularly stained cells, the most accurate measurement of MAIT cell frequencies will be obtained from immediate surface staining prior to stimulation, due to activation-induced downregulation of the TCR.
-
25.
If cells are not to be acquired immediately, then they can instead be resuspended in 100 μL of fixation buffer and stored at 4 °C until required.
-
26.
This may differ depending on virus and mouse strains.
-
27.
The overlay media will start setting so proceed to the following steps quickly. Overlay media can be made in batches to assist with that.
References
Eckle SB, Corbett AJ, Keller AN et al (2015) Recognition of vitamin B precursors and byproducts by mucosal associated invariant T cells. J Biol Chem 290:30204–30211
Godfrey DI, Uldrich AP, Mccluskey J et al (2015) The burgeoning family of unconventional T cells. Nat Immunol 16:1114–1123
Hinks TSC, Marchi, E, Jabeen, M et al (2019) Activation and in vivo evolution of the MAIT cell transcriptome in mice and humans reveals tissue repair functionality. Cell Reports 28(12):3249–3262.e5
Kjer-Nielsen L, Patel O, Corbett AJ et al (2012) MR1 presents microbial vitamin B metabolites to MAIT cells. Nature 491:717–723
Hinks TS, Zhou X, Staples KJ et al (2015) Innate and adaptive T cells in asthmatic patients: relationship to severity and disease mechanisms. J Allergy Clin Immunol 136:323–333
Rahimpour A, Koay HF, Enders A et al (2015) Identification of phenotypically and functionally heterogeneous mouse mucosal-associated invariant T cells using MR1 tetramers. J Exp Med 212:1095–1108
Tsukamoto K, Deakin JE, Graves JA et al (2013) Exceptionally high conservation of the MHC class I-related gene, MR1, among mammals. Immunogenetics 65:115–124
Corbett AJ, Eckle SB, Birkinshaw RW et al (2014) T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature 509:361–365
Patel O, Kjer-Nielsen L, Le Nours J et al (2013) Recognition of vitamin B metabolites by mucosal-associated invariant T cells. Nat Commun 4:2142
Ussher JE, Bilton M, Attwod E et al (2014) CD161++ CD8+ T cells, including the MAIT cell subset, are specifically activated by IL-12+IL-18 in a TCR-independent manner. Eur J Immunol 44:195–203
Van Wilgenburg B, Scherwitzl I, Hutchinson EC et al (2016) MAIT cells are activated during human viral infections. Nat Commun 7:11653
Wilgenburg BV, Loh L, Chen Z et al (2018) MAIT cells contribute to protection against lethal influenza infection in vivo. Nat Commun 9:4706
Loh L, Wang Z, Sant S et al (2016) Human mucosal-associated invariant T cells contribute to antiviral influenza immunity via IL-18-dependent activation. Proc Natl Acad Sci U S A 113:10133–10138
Hoiseth SK, Stocker BA (1981) Aromatic-dependent Salmonella typhimurium are non-virulent and effective as live vaccines. Nature 291:238–239
Chen Z, Wang H, D’souza C et al (2017) Mucosal-associated invariant T-cell activation and accumulation after in vivo infection depends on microbial riboflavin synthesis and co-stimulatory signals. Mucosal Immunol 10:58–68
Reantragoon R, Corbett AJ, Sakala IG et al (2013) Antigen-loaded MR1 tetramers define T cell receptor heterogeneity in mucosal-associated invariant T cells. J Exp Med 210:2305–2320
Wang H, D’souza C, Lim XY et al (2018) MAIT cells protect against pulmonary Legionella longbeachae infection. Nat Commun 9:3350
Acknowledgments
This work was funded by grants to T.S.C.H. from the Wellcome Trust (104553/z/14/z, 211050/Z/18/z) and Project Grants 1062889 and 1120467 and Program Grant 1113293 from the National Health and Medical Research Council of Australia. B.W. was supported by the Royal Society (IE160540). A.J.C. is supported by a Future Fellowship from the Australian Research Council, FT1600100083. The research leading to these results has received funding from the People Programme (Marie Curie Actions) of the European Union’s Seventh Framework Programme (FP7/2007-2013) under REA grant agreement number 608765. The content represents only the authors’ views and not those of the European Commission. HW was supported by a Melbourne International Engagement Award (University of Melbourne).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2020 The Author(s)
About this protocol

Cite this protocol
Hinks, T.S.C. et al. (2020). Study of MAIT Cell Activation in Viral Infections In Vivo. In: Kaipe, H., Magalhaes, I. (eds) MAIT Cells. Methods in Molecular Biology, vol 2098. Humana, New York, NY. https://doi.org/10.1007/978-1-0716-0207-2_17
Download citation
DOI: https://doi.org/10.1007/978-1-0716-0207-2_17
Published:
Publisher Name: Humana, New York, NY
Print ISBN: 978-1-0716-0206-5
Online ISBN: 978-1-0716-0207-2
eBook Packages: Springer Protocols