
Abstract
Deregulated or increased signalling of the epidermal growth factor receptor (EGFR) plays an integral role in the development of various cancer types, including head and neck squamous cell carcinoma (HNSCC), making it a compelling drug target. However, after initially promising results of EGFR-targeted therapies, such as the monoclonal antibody cetuximab, it became clear that both intrinsic and acquired therapeutic resistance are major roadblocks in the field of personalised cancer treatments.
In order to unravel and overcome resistance to cetuximab, at least two strategies can be adopted.
Firstly, therapeutic resistance to anti-EGFR therapy may arise from mechanisms that can compensate for reduced EGFR signalling and/or mechanisms that can modulate EGFR-dependent signalling. In this chapter, we discuss which mechanisms of cetuximab resistance are already known and which ones deserve further investigation. This enhanced knowledge will guide us to rationally design and test novel combination therapies that overcome resistance to EGFR-targeting agents in cancer treatment.
Secondly, an urgent need remains to develop novel targeted treatments for single-agent or combined therapy use. In this view, due to the particular mode of activation of the EGFR receptor, involving ligand-induced homo- and heterodimerization of the four HER receptors, an increased inhibition scope of HER receptors most likely results in a more potent blockade of the HER network, preventing premature emergence of resistance and leading to a more pronounced therapeutic benefit. We discuss two multitargeted compounds, being MEHD7945A (duligotuzumab) and afatinib, in this chapter.
Despite the huge efforts to unravel the molecular landscape of HNSCC, the main clinically validated target remains EGFR. However, immune checkpoints, like programmed cell death protein 1 (PD-1), are gaining clinical approvals as well. We underscore the importance of adopting rational drug combinations to enhance the therapeutic effect of the EGFR-inhibitor cetuximab and highlight the ongoing search for predictive biomarkers, with the ultimate goal of delivering individualized cancer therapy to HNSCC patients.
You have full access to this open access chapter, Download conference paper PDF
Similar content being viewed by others


Keywords
Introduction
Head and neck squamous cell carcinoma (HNSCC) is the sixth most common cancer type worldwide and accounts for a standardised worldwide incidence of roughly 600,000 individuals/year [1]. HNSCC remains one of the most challenging malignancies to treat. For example, the overall 5-year relative survival proportion for the Belgian 2013–2017 cohort was about 52% in males and 59% in females [2]. Unfortunately, patients with multiple metastases typically have very poor prognosis with a 5-year overall survival of only 4% [3]. Therefore, innovative therapeutic strategies are a necessity to increase the survival outcomes.
The introduction of targeted therapies that inhibit oncogenic signalling pathways is now at the forefront of personalised medicine in cancer treatment. As the epidermal growth factor receptor (EGFR) initiates important signalling pathways and is overexpressed and/or deregulated in a wide range of malignancies, this receptor is considered as an excellent drug target. Improved understanding of EGFR signalling in cancer has led to the development of two main categories of EGFR-targeting agents: the monoclonal antibodies (mAbs, such as cetuximab and panitumumab) and the tyrosine kinase inhibitors (TKIs, such as erlotinib and gefitinib) [4].
The Epidermal Growth Factor Receptor
The physiological function of EGFR is to regulate epithelial tissue development and homeostasis through cellular processes such as proliferation, maturation and apoptosis [5]. These cellular processes play an important role in the transformation of healthy cells into malignant tumour cells. The activity of the EGFR signal transduction pathway is tightly controlled in healthy cells, but deregulation of EGFR signalling plays a crucial role in carcinogenesis [6]. Accordingly, EGFR signalling has been studied intensively in order to understand its importance in cancer biology.
EGFR is a cell surface receptor that belongs to the HER or ErbB tyrosine kinases family. Besides EGFR, also known as HER1 or ErbB1, other members of the HER family include HER2 (ErbB2 or Neu), HER3 (ErbB3), and HER4 (ErbB4). The structure among these receptors is very similar: they consist of an extracellular ligand-binding domain, a single membrane-spanning region, a juxtamembrane nuclear localisation signal and a cytoplasmic tyrosine-kinase domain [7]. HER receptors are activated by a range of growth factors that belong to the EGF-family and can be divided into three groups (Fig. 3.1). The first group includes EGF, transforming growth factor-α (TGF-α) and amphiregulin, which all bind to EGFR. The second group includes betacellulin, heparin-binding EGF (HB-EGF) and epiregulin which bind to both EGFR and HER4. The third group is composed of the neuregulins (NRG1-4), which is further subdivided based on their ability to bind HER3 and HER4 (NRG1 and NRG2), or only to HER4 (NRG3 and NRG4) [8]. Until now, no known ligand exists for HER2 [7, 9].

The HER family receptors and ligands. Binding of a ligand to the extracellular domain of the receptor leads to receptor homo- and hetero-dimerisation and activation. TGF-α, transforming growth factor-α; HB-EGF, heparin-binding EGF
Binding of a ligand to the extracellular domain of these receptors leads to a conformational change that allows for receptor homo- and hetero-dimerisation and activation of intrinsic tyrosine kinase activity resulting in transphosphorylation of specific tyrosine residues within the intracellular domain. Autophosphorylation triggers a series of intracellular pathways that may result in cancer-cell proliferation, blocking apoptosis, activating invasion and metastasis, and stimulating tumour-induced neovascularisation. Figure 3.2 gives an overview of the HER receptor signalling pathways, including the Ras/Raf/mitogen-activated protein kinase (MAPK) pathway, the phosphatidylinositol 3-kinase (PI3K)/Akt pathway, the signal transducers and activators of transcription (STAT) pathway, Phospholipase Cγ, Src kinase pathways, the NcK/PAK signalling cascade and Cbl-mediated endocytosis.

HER receptor signalling pathways. Ligand binding to HER receptors is followed by receptor homo- and hetero-dimerisation and the activation of several downstream signalling pathways
During the 1980s, several studies described the overexpression of EGFR in a variety of epithelial tumours, which supported the hypothesis that dysregulated EGFR expression and signalling play an important role in the development of cancer [7, 10,11,12,13,14]. Over the last years, the oncogenic role of EGFR has been characterized in more detail and several alterations have been described [15]. Firstly, gene amplification leading to EGFR overexpression is often observed in human cancers [16, 17]. Secondly, point mutations and deletions in the EGFR gene can result in increased catalytic tyrosine kinase activity or a truncated form of the receptor, resulting in ligand-independent activity [18]. The most common tyrosine kinase EGFR mutations include the deletion of four conserved amino acids residues in exon 19 and the point mutation L858R in exon 21 [19, 20]. In addition, the EGFR variant III (EGFRvIII) is also frequently detected and constitutes a truncated form of EGFR caused by an in-frame deletion of 801 base pairs (exon 2–7) in the coding sequence of the extracellular domain [21, 22]. Next, increased ligand expression leads to constitutive stimulation of EGFR. EGF-related growth factors can be produced either by the tumour cells themselves or by surrounding stromal cells [23]. Finally, impaired receptor downregulation also results in sustained EGFR signalling [24]. All these alterations, consequently, result in increased EGFR activation and/or deregulation of EGFR signal transduction pathways. As EGFR stimulates many complex intracellular signalling pathways that are involved in proliferation, differentiation, apoptosis, angiogenesis and metastasis, activation of EGFR drives the malignant behaviour of the tumour [25].
Cetuximab as Anti-EGFR Targeting Agent
For over a decade, the EGFR monoclonal antibody cetuximab is approved by the European Medicines Agency (EMA) and Food and Drug Administration (FDA) for HNSCC treatment in three specific settings, i.e. (1) in combination with radiation therapy for locoregionally advanced HNSCC; (2) in combination with platinum-based therapy and 5-fluorouracil for first-line treatment for recurrent or metastatic (R/M) HNSCC; and (3) as a single agent for R/M HNSCC patients who failed on prior platinum-based therapy [26,27,28].
The therapeutic effect of the monoclonal antibodies, such as cetuximab and panitumumab, is exerted by binding to the extracellular domain of EGFR, thereby hindering ligands to bind and activate EGFR, preventing receptor dimerisation and promoting EGFR internalisation [15]. Importantly, as a chimeric human:mouse immunoglobulin G1 (IgG1), cetuximab can also elicit host anti-tumour immune responses. Through its IgG1 backbone, cetuximab can bind CD16 fragment crystallisable (Fc) receptors located on natural killer (NK) cells, macrophages and granulocytes, of which NK cells have been proven to be the most potent effectors [29]. Binding of the IgG1-Fc part of cetuximab to CD16 on NK cells triggers cytolytic activity called antibody-dependent cellular cytotoxicity (ADCC), which is predominantly mediated by perforin and granzymes [30]. Furthermore, cetuximab has been shown to enhance cross-priming of cytotoxic T-lymphocytes via professional antigen-presenting cells, such as dendritic cells [31], mainly through induction of immunogenic cell death of tumour cells [32]. These results confirmed the important immune-related mechanism of action of cetuximab, in addition to its receptor blocking effects.
Mechanisms of Cetuximab Resistance
Personalised medicine using targeted therapies, based on the molecular profile of the tumour, may achieve the much-needed progress in cancer treatment. After the initially promising results of EGFR-targeted therapies such as cetuximab, therapeutic resistance poses a challenging problem and limits the success of effective cancer therapies in the clinic [33]. If resistance to therapy is present at baseline, this is defined as intrinsic (primary) resistance and can be explained by resistance-conferring factors pre-existing in the bulk of tumour cells. Moreover, nearly all patients whose tumours initially respond inevitably become acquired (secondary) resistant. Acquired resistance refers to disease progression in the face of ongoing treatment that was initially effective [34].
Indeed, despite the enhanced EGFR expression in the majority of HNSCC tumours, therapeutic resistance remains a major roadblock in the search to effective HNSCC therapies and only a small subset of HNSCC patients benefit from cetuximab as a single agent (<15%) or combined with chemotherapy (36%). The addition of cetuximab to either radiotherapy in the locoregionally advanced disease setting, or to platinum and 5-fluorouracil chemotherapy (EXTREME regimen) for treatment of recurrent or metastatic HNSCC improved median overall survival from 29.3 to 49 months and from 7.4 to 10.1 months, respectively. Nevertheless, time-to-treatment failure in patients treated with the EXTREME regimen ranges only around 5 months, despite cetuximab maintenance [28, 35, 36].
Therapeutic resistance to anti-EGFR therapy may arise from mechanisms that can compensate for reduced EGFR signalling and/or mechanisms that can modulate EGFR-dependent signalling. Over the last years, a wide range of potential molecular mechanisms of resistance to EGFR-targeting agents has been described [15].
Altered Response Elicited at the Level of EGFR
Drug resistance can arise from sustained EGFR signalling that is elicited at the level of EGFR itself by ligand or receptor overexpression, amplification or mutation [15, 19, 20, 37,38,39,40,41]. Furthermore, EGFR can escape the internalisation and lysosomal degradation route and function as a transcription factor in the nucleus, inducing sustained EGFR signalling [42].
Binding of ligands to EGFR drives homodimerisation or heterodimerisation with ErbB family members, resulting in the initiation of downstream signalling pathways. Therefore, overexpression of its ligands may contribute to cetuximab resistance. A correlation with enhanced response to cetuximab therapy and overexpression of the EGFR ligands amphiregulin and epiregulin in K-Ras wild type metastatic colorectal tumours has been reported [40]. In HNSCC patients receiving cetuximab-docetaxel treatment, high amphiregulin levels were detected in 45% of the patients. A significant correlation was found between high amphiregulin levels and shortened overall survival and progression free survival compared with patients with low amphiregulin expression [41].
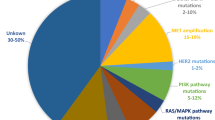
Neither the expression level of the EGFR protein, nor the amplification status of the EGFR gene could be linked to therapeutic response [43, 44]. Activating mutations have been observed in the tyrosine kinase domain or in the extracellular ligand-binding domain of EGFR [18]. The most common tyrosine kinase EGFR mutations include deletion of four conserved amino acids residues (leucine-arginine-glutamic acid-alanine) in exon 19 and a point mutation, L858R, in exon 21, which account for 90% of all EGFR tyrosine kinase mutations in non-small cell lung cancer (NSCLC) [45,46,47]. These EGFR tyrosine kinase mutations are associated with an improved clinical response to TKIs (gefitinib or erlotinib) in NSCLC patients but they are rarely found in HNSCC. Literature data suggest that the incidence of such activating mutations in HNSCC patients range from 0 to 15.7% [45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60]. In these studies, a total of 889 HNSCC samples were screened for EGFR tyrosine kinase mutations, of which 34 (3.8%) contained a mutation. Interestingly, the missense mutation T790M in exon 20, which is associated with acquired resistance to EGFR TKIs in about half of all patients with NSCLC, was found in 7.5% of all EGFR mutations in HNSCC [61]. Given that in HNSCC, the overall prevalence of mutations in the EGFR tyrosine kinase domain is only 2.8%, it is challenging to identify specific EGFR mutations related to response or resistance to anti-EGFR therapy [62].
Next to the above-discussed mutations, the EGFR variant III (EGFRvIII) is a truncated form of EGFR. The causing mutation consists of an in-frame deletion of 801 base pairs (exon 2–7) in the coding sequence of the extracellular domain, resulting in ligand-independent tyrosine kinase activity [21, 63]. The mutant EGFRvIII form is associated with increased proliferation, tumour growth, cell motility and invasion in vitro and resistance to anti-EGFR therapy [21, 64]. The mutation frequency of EGFRvIII in HNSCC ranges from 0 till up to 48% [41, 49, 51, 64,65,66,67]. It has been suggested that the EGFRvIII might be more available in the recurrent/metastatic disease setting and might be responsible for the lack of response to EGFR-targeted therapies [67].
Occasionally, a part of the EGFR receptor escapes the internalisation and lysosomal degradation route and translocates to the nucleus [42, 68]. In oral squamous cancers, nuclear EGFR was observed in 24.3% of patients [69]. This nuclear EGFR functions either as a transcription factor of cyclin D1, inducible Nitric Oxide Synthase (iNOS), B-Myb and cyclo-oxygenase-2 (COX-2), or as a tyrosine kinase phosphorylating and stabilizing proliferating cell nuclear antigen (PCNA), resulting in an activation of the nitric oxide pathway and increased G1/S progression of the cell cycle [70,71,72,73,74]. Consequently, the proliferative potential of the cancer cells is thereby enhanced. The presence of nuclear EGFR is not only associated with poor prognosis, but also with treatment resistance [69, 75, 76]. Besides its potential involvement in resistance mechanisms, nuclear EGFR is also associated with local recurrence [75].
Molecular Alterations in Effectors Downstream of EGFR
Resistance to EGFR inhibitors can also be the result of molecular alterations in effectors downstream of EGFR [15]. In particular, the RAS, PI3K, Akt, STAT and Src proteins have been suggested to contribute to drug resistance [77,78,79,80,81,82,83,84,85,86,87,88]. We previously showed that proteins related to the Ras-MAPK pathway are involved in mechanisms of resistance towards cetuximab in HNSCC [89]. This is confirmed in other studies, highlighting the significance of persistent activation or reactivation of the Ras-MAPK pathway in EGFR targeting drug resistance [90,91,92].
Firstly, K-Ras is a protein located downstream of EGFR in the Ras-MAPK pathway. Somatic point mutations in K-Ras occur in a variety of human malignancies, most frequently in pancreatic cancer, NSCLC and colon cancers [93, 94]. A mutation in codon 12 or 13 in this gene leads to constitutive activation of the protein, regardless of upstream activating signals. In colorectal tumours, these mutations confer resistance to therapy with the EGFR targeting monoclonal antibodies cetuximab and/or panitumumab [77, 78, 95, 96]. Approximately 30–40% of colorectal tumours harbour a K-Ras mutation [97, 98]. In contrast, in HNSCC, these K-Ras mutations are infrequent; in different reports the frequency of K-Ras mutations in HNSCC is ranging from 0 to 9.1% [45, 51, 99].
Another family member of the Ras proto-oncogenes is H-Ras. Mutations in H-Ras have been reported in literature and vary between 0 and 22% [100,101,102,103,104]. A very recent study demonstrated that KRAS/HRAS mutations were associated with poor progression-free survival among HNSCC patients treated with cetuximab in the first-line recurrent setting, but not among patients treated with cetuximab in combination with radiotherapy [105], thus suggesting that not only K-Ras but also H-Ras mutations might play a role in cetuximab resistance.
Secondly, further downstream of K-Ras in the MAPK signalling pathway, a member of the dual-specificity phosphatase (DUSP) family is located. DUSP proteins are involved in a negative feedback mechanism of the MAPK signalling pathway by dephosphorylation of the threonine-glutamic acid-tyrosine motif on MAP kinases [106]. Therefore, DUSP proteins can be seen as tumour suppressor proteins, and loss of their expression may promote constitutive activation of ERK and uncontrolled cell growth. Moreover, inhibition of the MAPK pathway can be compensated by suppression of the DUSP enzymes [92]. Both the cytoplasmic DUSP5 and the nuclear DUSP6 can dephosphorylate ERK1/2, thereby blocking the MAPK signal transduction cascade [107].
Low DUSP6 expression has proven to be clinically significant as it was observed in 40% of patients with oesophageal squamous cell carcinomas and 75% of nasopharyngeal patients [108]. This might occur through hypermethylation of CpG islands in intron 1 or loss of heterozygosity of the DUSP6 locus [109, 110]. As DUSP6 is a critical negative regulator of Erk1/2 [111], we previously evaluated the level of Erk1/2 phosphorylation and demonstrated that significantly more Erk1/2 phosphorylation was present in cetuximab resistant HNSCC cells after cetuximab treatment compared with cetuximab sensitive HNSCC cells. Furthermore, apigenin, an Erk1/2 inhibitor, dose-dependently inhibited survival of cetuximab resistant cells and a significant decrease in cell survival was observed when these cells were treated with a combination of apigenin and cetuximab [89]. Additionally, sustained or reactivated Erk, caused by downregulation of DUSP6, has been observed in lung cancer cells with acquired erlotinib resistance [91]. This highlights the significance of our findings and indicates that the exact function of the DUSP family proteins in relation to cetuximab resistance in HNSCC should be further elucidated.
Thirdly, besides activation of the Ras/Raf/MAPK signalling pathway, EGFR can also mediate activation of the PI3K/Akt pathway. This pathway is involved in various biological processes essential for normal cellular functionality, including survival, proliferation, differentiation, angiogenesis, protein synthesis and glucose metabolism. Besides these physiological functions, the pathway is also associated with a number of oncogenic processes and is one of the most frequently dysregulated pathways in cancer, including HNSCC [112, 113]. As such, aberrant signalling can lead to the stimulation of cell growth, inhibition of cell death and the promotion of invasion and migration [114,115,116], which is all in favour of cancer cells. Furthermore, increasing evidence indicates that the PI3K/Akt pathway frequently remains activated, despite anti-EGFR treatment and therefore plays an important role in resistance to EGFR-targeting therapies [117,118,119,120].
Fourthly, Src kinases are upstream as well as downstream activators of EGFR and other receptor tyrosine kinases. Upon EGFR stimulation, Src kinases are activated and associate with EGFR. As such, they can affect cellular proliferation and survival by activation of STAT family of transcription factors, especially STAT3 and STAT5 [121, 122]. In vitro studies showed reduced activity of Src kinases following EGFR inhibition [123]. Elevated Src levels and/or kinase activity have been shown in HNSCC and other malignancies [122, 124]. Therefore, activation of Src kinases by EGFR upstream or downstream signalling might result in resistance to anti-EGFR therapy.
As mentioned earlier, nuclear translocation of EGFR is a possible mechanism of resistance to therapy and this has been observed in patients treated with cetuximab and radiotherapy. Phosphorylation of EGFR on tyrosine 845 by the Src kinases enhances EGFR-mediated mitogenesis by binding and phosphorylating the STAT5b transcription factor and this has been described as the underlying mechanism responsible for nuclear translocation of EGFR [88, 125]. Indeed, dasatinib, a Src inhibitor, blocks EGFR translocation to the nucleus in HNSCC cell lines and, therefore, might be a potential way to evade resistance to anti-EGFR therapy [126]. In addition, in oral squamous cell carcinoma, it was shown that the combination of cetuximab and a Src inhibitor may provide more effective therapy than either inhibitor alone [127]. Collectively, these results indicate that Src inhibitors may be useful in overcoming anti-EGFR resistance by decreasing activated STAT3 and STAT5.
Finally, when considering resistance to anti-EGFR therapy, the signal transducer and activator of transcription (STAT) family proteins are also important downstream EGFR effectors. This family plays an important role in transmitting survival signals and anti-apoptotic signals that are initiated through activation of EGFR; especially activation of STAT3 and STAT5 has been linked to phosphorylation of EGFR [122, 128, 129]. Therefore, dysregulation of the STAT signalling pathway has been proposed to be implicated in malignant transformation.
Activation of STAT3 leads to the activation of several survival proteins, including Bcl-xl, Bcl-2 and survivin [84]. In HNSCC, STAT3 activation can be mediated by JAK and Src signalling, and partially by EGFR signalling [126, 130]. As such, STAT3 can be inhibited via EGFR blocking and this has been demonstrated in vitro and in vivo [131]. It has been shown that the anti-proliferative effects of cetuximab, as well as cetuximab-induced apoptosis, are more pronounced in STAT3 knockdown cells compared to control cells [84]. Recently, increased STAT3 expression was found in two acquired cetuximab-resistant HNSCC cell lines, compared to their parental lines. Moreover, STAT3 knockdown promoted increased cytotoxicity both in the presence and absence of cetuximab in the resistant lines [132], suggesting that STAT3 may be a common target in cetuximab resistance.
Cross-talk with Other Receptor Tyrosine Kinases
Selective stress of EGFR-targeting agents can lead to activation of alternative signalling pathways to compensate for the reduced EGFR signalling, thereby promoting cell survival [15]. Examples of alternative receptor pathways include other HER receptor family members, insulin growth factor type 1 receptor (IGF-1R) and MET [133,134,135,136,137,138,139].
Firstly, as mentioned earlier, EGFR is a family member of the ErbB receptor family, and activation of other members of this family might result in resistance to anti-EGFR therapy. In the literature, activation of HER2 signalling has been associated with cetuximab resistance, as its signalling occurs through many of the same downstream effectors of EGFR. Using an in vitro model of acquired cetuximab resistance, a marked increase in the phosphorylation status of the C-terminal fragment of HER2, 611-CTF, was observed. Combination therapy of afatinib (an irreversible EGFR/HER2/HER4 inhibitor) with cetuximab resulted in a dramatic reduction in cetuximab resistant tumour volumes compared to either agent alone in monotherapy [133]. Therefore, it was suggested that dual inhibition of EGFR and HER2 could be an effective approach to enhance the efficacy of cetuximab, in order to prevent and/or overcome cetuximab resistance. Likewise, a study by Yonesaka et al. has shown that cetuximab resistance could be induced by activation of ErbB2 signalling. The underlying mechanism involved amplification of ErbB2 or upregulation of heregulin, both leading to persistent ERK1/2 activation. Moreover, restoring cetuximab sensitivity was accomplished by inhibition of ErbB2 or by disruption of ErbB2/ErbB3 heterodimerisation in vitro as well as in vivo [134]. More studies are warranted in order to determine the frequency of HER2 mutations in HNSCC and their role in the response to TKIs.
Secondly, activation of the insulin growth factor type 1 receptor (IGF-1R) leads to downstream activation of the Ras/Raf/MAPK and PI3K/Akt pathway and enhances survivin expression, all contributing to cell proliferation, altered cell adhesion, enhanced motility properties and impaired apoptosis [140, 141]. Analysis of the HNSCC subsets of the Cancer Genome Atlas has identified 4% amplification and mutation of IGF-1R gene in human papillomavirus (HPV) negative HNSCC patients [142]. Furthermore, activation of IGF-1R has been reported to induce resistance to EGFR TKIs [143]. It was shown that heterodimerization of EGFR with IGF-1R was increased in cetuximab resistant HNSCC cancer cells [144]. This heterodimerization of EGFR with IGF-1R lead to increased activity of EGFR and might be an important platform for cetuximab-mediated signalling in head and neck tumours that have become resistant to anti-EGFR therapy. As such, dual targeting of EGFR and IGF-1R could be a promising therapeutic strategy.
Thirdly, the MET proto-oncogene encodes a transmembrane receptor tyrosine kinase MET, also known as c-MET or hepatocyte growth factor receptor (HGFR). The MET pathway can be deregulated in two different ways: on the one hand by mutation and/or amplification of MET, and on the other hand by increased ligand expression and/or activity, both resulting in persistent activation of the PI3K/Akt signalling pathway [138]. Circa 80% of primary HNSCC tumours express the ligand hepatocyte growth factor (HGF), MET, or both, thus activating important downstream signals, which overlap with EGFR signalling [65, 145]. Moreover, MET mutations or amplifications have been observed in 13.5% and 13% of HNSCC tumours, respectively [146]. As high MET expression could be observed in 58% of patients with recurrent/metastatic HNSCC [65], the role of MET in resistance to anti-EGFR therapy has been investigated in a number of studies. Chau et al. did not detect any association between response to erlotinib and time to progression or overall survival, in recurrent/metastatic HNSCC patients with high MET expression [65]. Experiments in vitro and in vivo showed that MET confers resistance to cetuximab via activation of the MAPK pathway. In addition to the direct role of MET in reactivation of the MAPK pathway, MET stimulation also abrogated the well-known cetuximab-induced compensatory feedback loop of HER2/HER3 expression [147]. In a HNSCC xenograft model, a delay in tumour growth was observed after administration of crizotinib, a MET TKI [145]. Collectively, these data suggest that high MET expression might play a role in cetuximab resistance.
Alterations in Proteins Outside the EGFR Pathway
Not only alterations in proteins involved in EGFR signalling but also proteins such as cyclin D1 and p53, linked to more general characteristics of cancer (such as proliferation, apoptosis, invasion and metastasis) can confer resistance to EGFR inhibitors [148,149,150,151].
The Aurora kinases A and B are highly conserved serine/threonine kinases that play an essential and distinct role in mitosis [152, 153]. Overexpression of both kinases is frequently present in many types of malignant tumours, and in the case of HNSCC, overexpression of Aurora kinase A is found in up to 90% of tumours [153,154,155]. Overexpression of Aurora kinase A is correlated with tumour progression, a metastatic phenotype and shortened survival, and is therefore regarded as a negative prognostic marker [152, 154, 155]. High expression levels of Aurora kinase B are found in glioblastoma, ovarian carcinoma and hepatocellular carcinoma and are associated with poor prognosis [156].
The EGFR pathway can elicit overexpression of Aurora kinase A at two different levels, i.e. (i) EGF increases the translational efficiency of Aurora kinase A; and (ii) translocation of EGFR to the nucleus results in binding to the Aurora kinase A promoter and thereby increasing its transcription. Both ultimately result in chromosome instability and tumourigenesis [73, 157].
Next to its role as a prognostic factor, studies indicated evidence for a role of Aurora kinase A in the response to therapy. Overexpression of Aurora kinase A triggered the activation of two important molecules involved in the regulation of drug resistance, Akt and NF-κB [158]. Interestingly, knockdown of Aurora kinase A in HeLa cells resulted in sensitisation to cisplatin, and Aurora kinase A overexpression could overcome cell death induced by paclitaxel [158]. Furthermore, treatment of HNSCC cells with cetuximab and a pan-Aurora kinase inhibitor R763 resulted in a rapid and efficient decrease in the level of the Aurora kinase substrate S10HH3. These results could not be confirmed by using a specific Aurora kinase A inhibitor and, therefore, it was concluded that the effects of the pan-Aurora kinase inhibitor were most likely mediated by its blockage of Aurora kinase B activity [152]. Similarly, we previously showed that cell growth of cetuximab resistant cells could be inhibited by blocking Aurora kinase B [89]. Collectively, these results indicate that the Aurora kinases may be an interesting target for HNSCC tumours resistant to anti-EGFR therapy.
The G1/S-specific cyclin D1 forms a complex with CDK4 and CDK6 and functions as a regulatory subunit of CDK4 and CDK6, the activity of which is required for cell cycle G1/S transition. As previously mentioned, nuclear EGFR functions as a transcription factor for cyclin D1. Moreover, constitutive activation of STAT3 is required for EGFR-mediated cell growth and results in elevated levels of STAT3 target genes, including cyclin D1 [129, 159].
HNSCCs that are unrelated to the human papillomavirus (HPV), are often driven by p16INK4A inactivation and cyclin D1 overexpression that cause hyperactivation of cyclin-dependent kinase 4/6 (CDK4/6), which drives the cell cycle and tumour growth. Deregulated cyclin D1 expression also causes resistance to EGFR inhibitors. These somatic genomic alterations pointed to inhibition ofCDK4/6 as a potential targeted therapeutic strategy in HPV-unrelated HNSCC. The CDK4/6 inhibitor palbociclib arrests cell cycle progression by selective CDK4/6 inhibition and might also reverse intrinsic resistance to cetuximab by countering the actions of deregulated cyclin D1. The antiproliferative and antitumour effects of selective CDK4/6 inhibition have indeed been demonstrated in HNSCC cell lines and xenografts. In HPV-unrelated HNSCC cell lines, the combination of palbociclib and an EGFR inhibitor synergistically reduced cell viability andERK1/2 phosphorylation. Importantly, a recent multicentre, phase 2 trial, showed that the combination of palbociclib and cetuximab exhibited substantial antitumour activity in platinum-resistant and in cetuximab-resistant HPV-unrelated HNSCC [160]. As such, further investigation of selective CDK4/6 inhibition as a therapeutic strategy in HPV-unrelated HNSCC is certainly warranted.
The tumour suppressor protein p53 has a critical role in controlling cell cycle progression and, consequently, loss of its function is linked to the carcinogenic process. In response to a variety of cellular stimuli, p53 can induce cell cycle arrest, apoptosis or senescence.
A study investigating the difference between cetuximab resistant and their sensitive parental lung cancer cells, identified p53 as the most downregulated and pERK1/2 as the most upregulated cellular signalling protein. Downregulation of p53 was also observed in erlotinib resistant cells. Furthermore, silencing of p53 in cetuximab sensitive cells resulted in reduced sensitivity to the drug, whereas restoring p53 function in resistant cells resulted in enhanced cetuximab sensitivity [149]. In vivo experiments, using a stable cetuximab resistant clone with tetracycline-inducible p53 showed that repair of p53 restored cetuximab sensitivity in tumour xenografts resistant to cetuximab [149]. In addition, cetuximab was able to inhibit cell growth in p53 wild type cells, but not in p53 mutated cells [151]. In general, there is insufficient experimental evidence to unequivocally state that loss of functional p53 can be predictive of resistance to anti-EGFR therapy.
Epithelial-to-Mesenchymal Transition
We and others have proposed epithelial-to-mesenchymal transition (EMT) as a mechanism of resistance towards EGFR targeting therapeutics [161,162,163,164,165,166,167,168,169]. EMT is characterized by loss of epithelial cell characteristics and acquisition of mesenchymal phenotypic traits, causing tumour cells to detach from neighbouring cells and to migrate into adjacent tissue [170,171,172]. However, it has been reported that EGFR inhibition can promote an infiltrative front composed of mesenchymal-like cells, which made up a small subpopulation of the tumour before therapy [173]. Increased expression of IL8 and HB-EGF have been linked with EMT [37, 174,175,176,177,178,179,180] and we showed that both genes were upregulated in our cetuximab resistant HNSCC cells and these cells shows traits of EMT, including higher migratory and invasive capacity. Moreover, our microarray profile revealed upregulation of several epithelial markers in cetuximab sensitive HNSCC cells, whereas cetuximab resistant cells were characterized by upregulation of protease urokinase (PLAU), transgelin (TAGLN), ADAM19 and thrombospondin (TSP-1), all of which have functions associated with features of EMT [181,182,183,184,185]. Similarly, it has been reported that HNSCC cells with a mesenchymal-like morphology and elevated migratory potential were found to be less sensitive to irradiation and cetuximab [186]. Overall, these findings clearly indicate that cetuximab resistant cells show enhanced characteristics of EMT.
Hypoxia and Angiogenesis
Regions within solid tumours often experience mild to severe oxygen deprivation (hypoxia) and it has been well documented that poor oxygenation is a pathophysiological property of the majority of human solid tumours, including HNSCC [187]. Importantly, oxygen deficiency has a major impact on clinical responses to cancer treatment, and it was shown that hypoxic tumour regions often contain viable cells that are intrinsically more resistant to treatment with radiotherapy and/or chemotherapy [188, 189]. Both preclinical and clinical studies support an important link between hypoxia and upregulation of EGFR in cancers that do not display genetic alterations of the receptor [190]. Subsequent EGFR signalling stimulates hypoxia-inducible factor (HIF) signalling and thus augments induction of proteins that promote cellular survival in a hostile microenvironment. As the HIF transcription factors play a pivotal role in the cellular adaptation to hypoxic stress, EGFR-induced HIF signalling thus augments the induction of proteins that promote cellular survival in a hostile microenvironment. As a consequence, the presence of tumour hypoxia may contribute to resistance to EGFR inhibitors [191]. HNSCC patients with high levels of hypoxia-associated factors indeed were more likely to relapse, following induction therapy that included cetuximab [192], suggesting that the role of tumour hypoxia in therapeutic resistance might be particularly relevant for regimens containing EGFR-targeting monoclonal antibodies [192].
Lee et al. reported only minimal distribution of cetuximab to hypoxic tumour regions [193]. As monoclonal antibodies are large molecules, which are consumed by binding to receptors on the cell surface, this might indeed lead to poor penetration within solid tumours. However, cetuximab has a long half-life in the circulation, so that a more uniform distribution in tissues might be established, even if penetration of tissue is relatively slow. Indeed, in contradiction to the observation by Lee et al., Santiago et al. reported that cetuximab was homogeneously distributed within FaDu HNSCC xenografts, with no difference between hypoxic and non-hypoxic tumour cells [194]. These findings were in line with clinical data on the distribution of anti-EGFR antibodies in HNSCC [195] and indicate that cetuximab accesses not only (oxygenated) cells in proximity to the tumour blood vessels, but indiscriminately reaches all tumour cells.
Only few papers illustrate hypoxia-induced treatment resistance and most studies on EGFR-targeting antibodies supported a markedly increased anti-tumour potency of cetuximab in vivo (over that observed in vitro), suggesting that factors of the tumour microenvironment might influence the in vivo response.
The first reports on this topic addressed the association between the EGFR pathway and tumour angiogenesis. Together with the demonstrated antiproliferative and pro-apoptotic effects, the anti-angiogenic activity of cetuximab is now believed to contribute to its overall anti-tumour activity in vivo. With regard to this anti-angiogenic effect, numerous studies have shown that treatment of human cancer cells in vitro and in vivo with cetuximab reduced the production of VEGF [196,197,198]. Luwor et al. found that cetuximab reduced the levels of HIF-1α, leading to transcriptional inhibition of VEGF expression [199]. Immunohistochemical analysis of HNSCC tumour xenografts after systemic administration of cetuximab demonstrated inhibition of the in vivo expression of tumour angiogenesis markers, including VEGF and Factor VIII [200].
Apart from the observed anti-angiogenic effects, it has also been speculated that hypoxia enhances the sensitivity to the cytotoxic effect of EGFR-targeted monoclonal antibodies [201]. For example, cetuximab was more cytotoxic against hypoxic than well-oxygenated A431 epidermoid cancer cells grown in vitro and it reduced the overexpression of hypoxia markers (HIF-1α, CA9, VEGF) [198]. Likewise, we observed that both EGFR-inhibitors cetuximab and erlotinib maintained their growth inhibitory effect under hypoxia in vitro in three cetuximab-sensitive HNSCC cell lines [201]. Whether this was a direct interaction between hypoxia- and EGFR-mediated signalling pathways or indirectly via reoxygenation as a consequence of cell loss due to the cytotoxic effect of cetuximab [202] was not elucidated in these studies, but both mechanisms might be involved.
Therefore, several studies have focused on the molecular mechanisms behind the cross-talk between hypoxia and EGFR inhibition and on the role of HIF-1α in this process [191]. Importantly, it was observed that cetuximab could clearly downregulate HIF-1α levels in cancer cell lines that were sensitive to EGFR inhibition and it was shown that inhibition of HIF-1α was required, although it might not be sufficient, to mediate the response of cancer cells to EGFR-targeted monoclonal antibodies [199, 203,204,205]. In contrast, overexpression of HIF-1α in cancer cells that were originally sensitive to treatment with cetuximab conferred substantial resistance to this anti-EGFR therapy [204]. It was also reported that cetuximab sensitised HNSCC cells to radiation in part through inhibition of the radiation-induced upregulation of HIF-1α [206]. Overall, further in-depth studies are needed to fully understand these observations.
As inhibition of proteasomal degradation did not alter the rate of HIF-1α reduction by cetuximab treatment, it was suggested that cetuximab mainly acts by inhibiting HIF-1α protein synthesis [199, 207]. In hypoxic gastric cancer cells, it was shown that cetuximab reduced HIF-1α expression via inhibition of both MAPK and PI3K/AKT signalling downstream of EGFR [208]. However, most other studies suggested that the exact mechanism of reducing HIF-1α synthesis by cetuximab involved only inhibition of the PI3K/AKT pathway. The inhibition was shown to be prevented in cancer cells transfected with constitutively active PI3K or constitutively active AKT, but not in cells with a constitutively active MEK [204].
Overall, despite their individual key roles in promoting cancer progression and treatment resistance, our knowledge about the impact of intratumoural hypoxia on the activity of the EGFR signalling pathway in cancer and vice versa remains rather limited. As such, further studies are warranted to define the precise mechanistic and therapeutic implications of the hypoxic response relative to the EGFR signalling pathway in cancer.
Strategies to Overcome Cetuximab Resistance
Despite the reported intrinsic and acquired resistance to EGFR-targeting agents, interest in targeting EGFR for the treatment of HNSCC remains high, with new strategies, such as inhibitor combinations and novel irreversible or multi-targeting inhibitors, currently being evaluated.
Irreversible and Multiple HER Receptor Inhibition to Overcome Resistance
The ongoing challenge of therapy resistance has prompted a new approach to treat cancer patients, notably multiple inhibition of HER receptors simultaneously or irreversible inhibition. As mentioned above, the HER family of receptor tyrosine kinases comprises four members, i.e. EGFR (HER1, ErbB1), HER2 (ErbB2), HER3 (ErbB3) and HER4 (ErbB4). The particular mode of activation of the HER network involving ligand-induced homo- and hetero-dimerisation of the four HER receptors has prompted a new approach to inhibit this complex network and prevent premature emergence of resistance [15, 209]. The simultaneous inhibition of both partners in a HER dimer, using covalent binders that confer irreversible inhibition, represents one of these new paradigms. In this light, we will discuss two multitargeted compounds, being MEHD7945A (duligotuzumab) and afatinib.
For MEHD7945A (duligotuzumab), a monoclonal antibody with dual EGFR/HER3 specificity, we demonstrated that this compound has only a limited potential to establish a clear concentration-dependent cytotoxic effect in intrinsically and acquired cetuximab resistant HNSCC cell lines [210]. An additive but not synergistic interaction between MEHD7945A and cisplatin was observed. As the cytotoxic effect of MEHD7945A was not dependent on the expression of EGFR and HER3 in HNSCC cell lines, other mechanisms besides HER3 expression and signalling seem to play a pivotal role in resistance to cetuximab. This finding was supported by clinical data from the MEHGAN study, a randomized phase II study comparing MEHD7945A with cetuximab in platinum-pretreated but cetuximab-naïve HNSCC patients. This study demonstrated no benefit for MEHD7945A over cetuximab in neither all randomized patients, nor in patients whose tumours expressed high levels of HER3 or neuregulin (NRG1), a ligand of HER3. In addition, MEHD7945A also demonstrated disappointing results in a clinical study with RAS wild type CRC patients [211]. In this study, MEHD7945A plus FOLFIRI (leucovorin, 5-fluorouracil and irinotecan) did not appear to improve the outcomes of RAS wild type CRC patients compared with cetuximab plus FOLFIRI. Similarly, no association was found between progression free survival or objective response rate and HER3 or NRG1 expression. Due to the lack of survival benefit reported in several clinical trials, no additional clinical studies have recently been initiated with MEHD7945A (ClinicalTrials.gov), indicating the need to further investigate the potential of other multiple HER receptor inhibitors, such as for example afatinib.
In contrast to the first-generation EGFR inhibitors, afatinib is an irreversible HER family blocker that inhibits the enzymatic activity of EGFR, HER2 and HER4 [212,213,214,215]. As HER3 is kinase-inactive and requires obligate heterodimerization with other HER-family receptors, afatinib also inhibits HER3-mediated signal transduction. The increased inhibition scope of HER receptors by afatinib most likely leads to a more robust blockade of the HER network [216]. Previous preclinical research demonstrated effective cytotoxic activity of afatinib in HNSCC cell lines and xenograft models [217]. Consequently, treatment with afatinib might result in a distinct and more pronounced therapeutic benefit.
In this light, we demonstrated that afatinib was able to establish cytotoxicity in cetuximab sensitive, intrinsically and acquired resistant HNSCC cell lines, independent of the HPV status [218]. Neither cetuximab resistance nor HPV status had a significant impact on the efficacy of afatinib. Nevertheless, we noticed that intrinsically and acquired cetuximab resistant HNSCC cell lines tended to show higher IC50 values compared to their isogenic cetuximab sensitive counterparts, thus suggesting the possibility of cross-resistance between cetuximab and afatinib.
In HNSCC patients, the randomized phase II study of afatinib monotherapy versus cetuximab in R/M HNSCC patients reported that afatinib showed antitumor activity comparable to cetuximab with lack of cross-resistance [219]. In contrast, however, subgroup analysis of the phase III LUX-Head and Neck 1 trial with R/M HNSCC patients progressing on or after platinum-based therapy, suggested in 2017 that afatinib is more effective in patients whose tumours are cetuximab naïve [220, 221]. Nevertheless, a phase Ib study in patients with non-small cell lung cancer and HNSCC demonstrated promising results when afatinib was given in combination with standard-dose cetuximab [222].
The above-mentioned subgroup analysis of the LUX-Head and Neck 1 trial also suggested, based on prespecified biomarker assessment, increased benefit in patients whose tumours were, HPV-negative, had EGFR amplification, low HER3 expression and high PTEN expression [221]. However, our preclinical data suggested that the efficacy of afatinib was not significantly influenced by the HPV status of the cell line. Furthermore, in 2018, Machiels et al. reported that none of these biomarkers were significantly predictive of response for afatinib in a window of opportunity study of the European Organization for Research and Treatment of Cancer (EORTC) study in treatment-naïve HNSCC patients selected for primary curative surgery [223]. Possible explanations for these differences include the low number of patients resulting in low statistical power and the different clinical settings (curative versus palliative). Although these data were exploratory, Machiels et al. reported that the hypoxic gene signature and TP53 status needed to be further investigated as a predictive biomarker of afatinib activity. Our preclinical data support this finding, as the cytotoxic effect of afatinib was increased under hypoxic conditions in HNSCC cell lines. Consequently, further preclinical and clinical research are required to draw final conclusions upon the possible predictive role of cetuximab sensitivity, HPV status, hypoxia and TP53 status for the treatment with afatinib
Overall, the extended inhibition scope of HER receptors by afatinib leads to a more robust blockade of the HER network than MEHD7945A.Nevertheless, optimisation of combination treatment regimens with afatinib and conventional as well as other targeted therapies is necessary. Furthermore, identifying predictive biomarkers in order to select the patients that benefit most from these particular combination strategies is of crucial importance.
Identification of Drug Resistance Mechanisms and Predictive Biomarkers
In addition to optimising therapy strategies, optimal patient selection for anti-EGFR-based therapy remains a major challenge. As such, efforts at identifying predictive biomarkers to select HNSCC patients most likely to benefit from EGFR-targeted therapy have yet to succeed [15, 224, 225]. Unravelling the molecular pathways underlying resistance to EGFR inhibitors could have important implications, not only regarding patient selection, but also regarding the identification of novel drug targets for the treatment of HNSCC patients. In the paragraphs above, we discussed which mechanisms of cetuximab resistance are already known and which ones deserve further investigation. This enhanced knowledge will guide us to rationally design and test novel combination therapies that overcome resistance to EGFR-targeting agents in cancer treatment.
Conclusion and Future Perspectives
In conclusion, we hypothesize that the anti-tumour effects of cetuximab will be synergistic with agents targeting oncogenic bypass pathways responsible for therapeutic resistance towards cetuximab in HNSCC. Of particular interest and complexity are regimens combining immunotherapy with EGFR-targeted therapy. Indeed, the integration of immunotherapeutic approaches is now considered as a new perspective for the treatment of HNSCC patients. In this regard, the anti-PD-1 immune checkpoint inhibitor pembrolizumab has recently been approved by the American FDA and the European EMA for first-line treatment of R/M HNSCC [226]. As discussed previously, the working mechanism of cetuximab has largely been attributed to the direct effects of EGFR inhibition, but cetuximab also demonstrates additional immune-based mechanisms of activity through stimulation of antibody-dependent cellular cytotoxicity and enhancement of cytotoxic T-lymphocyte cross priming by dendritic cells [227,228,229]. As such, the immune system of the patient is involved in the anti-tumour effect of cetuximab and combinations with immunotherapeutic approaches also look highly promising for the treatment of HNSCC. We are hopeful that, with these novel combination strategies, cetuximab resistance can be prevented and a more pronounced therapeutic benefit can be achieved, ultimately resulting in improved survival and quality of life for HNSCC patients.
References
Leemans CR, Braakhuis BJ, Brakenhoff RH. The molecular biology of head and neck cancer. Nat Rev Cancer. 2011;11(1):9–22.
Registry BC. Stichting Kankerregister. Available from: https://kankerregister.org
Beckham TH, Leeman JE, Xie P, Li X, Goldman DA, Zhang Z, et al. Long-term survival in patients with metastatic head and neck squamous cell carcinoma treated with metastasis-directed therapy. Br J Cancer. 2019;121(11):897–903.
Zhang H, Berezov A, Wang Q, Zhang G, Drebin J, Murali R, et al. ErbB receptors: from oncogenes to targeted cancer therapies. J Clin Invest. 2007;117(8):2051–8.
Sigismund S, Avanzato D, Lanzetti L. Emerging functions of the EGFR in cancer. Mol Oncol. 2018;12(1):3–20.
Gschwind A, Fischer OM, Ullrich A. The discovery of receptor tyrosine kinases: targets for cancer therapy. Nat Rev Cancer. 2004;4(5):361–70.
Wheeler DL, Dunn EF, Harari PM. Understanding resistance to EGFR inhibitors-impact on future treatment strategies. Nat Rev Clin Oncol. 2010;7(9):493–507.
Hynes NE, Lane HA. ERBB receptors and cancer: the complexity of targeted inhibitors. Nat Rev Cancer. 2005;5(5):341–54.
Klapper LN, Glathe S, Vaisman N, Hynes NE, Andrews GC, Sela M, et al. The ErbB-2/HER2 oncoprotein of human carcinomas may function solely as a shared coreceptor for multiple stroma-derived growth factors. Proc Natl Acad Sci USA. 1999;96(9):4995–5000.
Ullrich A, Coussens L, Hayflick JS, Dull TJ, Gray A, Tam AW, et al. Human epidermal growth factor receptor cDNA sequence and aberrant expression of the amplified gene in A431 epidermoid carcinoma cells. Nature. 1984;309(5967):418–25.
Libermann TA, Razon N, Bartal AD, Yarden Y, Schlessinger J, Soreq H. Expression of epidermal growth factor receptors in human brain tumors. Cancer Res. 1984;44(2):753–60.
Libermann TA, Nusbaum HR, Razon N, Kris R, Lax I, Soreq H, et al. Amplification, enhanced expression and possible rearrangement of EGF receptor gene in primary human brain tumours of glial origin. Nature. 1985;313(5998):144–7.
Veale D, Ashcroft T, Marsh C, Gibson GJ, Harris AL. Epidermal growth factor receptors in non-small cell lung cancer. Br J Cancer. 1987;55(5):513–6.
Ushiro H, Cohen S. Identification of phosphotyrosine as a product of epidermal growth factor-activated protein kinase in A-431 cell membranes. J Biol Chem. 1980;255(18):8363–5.
Boeckx C, Baay M, Wouters A, Specenier P, Vermorken JB, Peeters M, et al. Anti-epidermal growth factor receptor therapy in head and neck squamous cell carcinoma: focus on potential molecular mechanisms of drug resistance. Oncologist. 2013;18(7):850–64.
Ohgaki H, Dessen P, Jourde B, Horstmann S, Nishikawa T, Di Patre PL, et al. Genetic pathways to glioblastoma: a population-based study. Cancer Res. 2004;64(19):6892–9.
Sunpaweravong P, Sunpaweravong S, Puttawibul P, Mitarnun W, Zeng C, Baron AE, et al. Epidermal growth factor receptor and cyclin D1 are independently amplified and overexpressed in esophageal squamous cell carcinoma. J Cancer Res Clin Oncol. 2005;131(2):111–9.
Laurent-Puig P, Lievre A, Blons H. Mutations and response to epidermal growth factor receptor inhibitors. Clin Cancer Res. 2009;15(4):1133–9.
Shigematsu H, Gazdar AF. Somatic mutations of epidermal growth factor receptor signaling pathway in lung cancers. Int J Cancer. 2006;118(2):257–62.
Lynch TJ, Bell DW, Sordella R, Gurubhagavatula S, Okimoto RA, Brannigan BW, et al. Activating mutations in the epidermal growth factor receptor underlying responsiveness of non-small-cell lung cancer to gefitinib. New Engl J Med. 2004;350(21):2129–39.
Wheeler SE, Suzuki S, Thomas SM, Sen M, Leeman-Neill RJ, Chiosea SI, et al. Epidermal growth factor receptor variant III mediates head and neck cancer cell invasion via STAT3 activation. Oncogene. 2010;29(37):5135–45.
Uribe P, Gonzalez S. Epidermal growth factor receptor (EGFR) and squamous cell carcinoma of the skin: molecular bases for EGFR-targeted therapy. Pathol Res Practice. 2011;207(6):337–42.
Salomon DS, Brandt R, Ciardiello F, Normanno N. Epidermal growth factor-related peptides and their receptors in human malignancies. Critic Rev Oncol Hematol. 1995;19(3):183–232.
Peschard P, Park M. Escape from Cbl-mediated downregulation: a recurrent theme for oncogenic deregulation of receptor tyrosine kinases. Cancer Cell. 2003;3(6):519–23.
Normanno N, De Luca A, Bianco C, Strizzi L, Mancino M, Maiello MR, et al. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene. 2006;366(1):2–16.
Bonner JA, Harari PM, Giralt J, Azarnia N, Shin DM, Cohen RB, et al. Radiotherapy plus cetuximab for squamous-cell carcinoma of the head and neck. N Engl J Med. 2006;354(6):567–78.
Bonner JA, Harari PM, Giralt J, Cohen RB, Jones CU, Sur RK, et al. Radiotherapy plus cetuximab for locoregionally advanced head and neck cancer: 5-year survival data from a phase 3 randomised trial, and relation between cetuximab-induced rash and survival. Lancet Oncol. 2010;11(1):21–8.
Vermorken JB, Mesia R, Rivera F, Remenar E, Kawecki A, Rottey S, et al. Platinum-based chemotherapy plus cetuximab in head and neck cancer. N Engl J Med. 2008;359(11):1116–27.
Trivedi S, Srivastava RM, Concha-Benavente F, Ferrone S, Garcia-Bates TM, Li J, et al. Anti-EGFR targeted monoclonal antibody isotype influences antitumor cellular immunity in head and neck cancer patients. Clin Cancer Res. 2016;22(21):5229–37.
Monteverde M, Milano G, Strola G, Maffi M, Lattanzio L, Vivenza D, et al. The relevance of ADCC for EGFR targeting: a review of the literature and a clinically-applicable method of assessment in patients. Crit Rev Oncol Hematol. 2015;95(2):179–90.
Kansy BA, Lin Y, Ding F, Gibson SP, Jie H-B, Ferris RL. Anti-EGFR mAb cetuximab therapy increases T cell receptor (TCR) diversity in the peripheral blood and focuses TCR richness in the tumor microenvironment. J Immuno Ther Cancer. 2015;3(2):P73.
Pozzi C, Cuomo A, Spadoni I, Magni E, Silvola A, Conte A, et al. The EGFR-specific antibody cetuximab combined with chemotherapy triggers immunogenic cell death. Nat Med. 2016;22(6):624–31.
Cohen RB. Current challenges and clinical investigations of epidermal growth factor receptor (EGFR)- and ErbB family-targeted agents in the treatment of head and neck squamous cell carcinoma (HNSCC). Cancer Treat Rev. 2013.
Leto SM, Trusolino L. Primary and acquired resistance to EGFR-targeted therapies in colorectal cancer: impact on future treatment strategies. J Mol Med. 2014;92(7):709–22.
Lee YS, Johnson DE, Grandis JR. An update: emerging drugs to treat squamous cell carcinomas of the head and neck. Expert Opin Emerg Drugs. 2018;23(4):283–99.
Vermorken JB, Herbst RS, Leon X, Amellal N, Baselga J. Overview of the efficacy of cetuximab in recurrent and/or metastatic squamous cell carcinoma of the head and neck in patients who previously failed platinum-based therapies. Cancer. 2008;112(12):2710–9.
Hatakeyama H, Cheng H, Wirth P, Counsell A, Marcrom SR, Wood CB, et al. Regulation of heparin-binding EGF-like growth factor by miR-212 and acquired cetuximab-resistance in head and neck squamous cell carcinoma. PLoS One. 2010;5(9):e12702.
Jijon HB, Buret A, Hirota CL, Hollenberg MD, Beck PL. The EGF receptor and HER2 participate in TNF-alpha-dependent MAPK activation and IL-8 secretion in intestinal epithelial cells. Mediators Inflamm. 2012;2012:207398.
Bedi A, Chang X, Noonan K, Pham V, Bedi R, Fertig EJ, et al. Inhibition of TGF-beta enhances the in vivo antitumor efficacy of EGF receptor-targeted therapy. Mol Cancer Ther. 2012;11(11):2429–39.
Khambata-Ford S, Garrett CR, Meropol NJ, Basik M, Harbison CT, Wu S, et al. Expression of epiregulin and amphiregulin and K-ras mutation status predict disease control in metastatic colorectal cancer patients treated with cetuximab. J Clin Oncol. 2007;25(22):3230–7.
Tinhofer I, Klinghammer K, Weichert W, Knodler M, Stenzinger A, Gauler T, et al. Expression of amphiregulin and EGFRvIII affect outcome of patients with squamous cell carcinoma of the head and neck receiving cetuximab-docetaxel treatment. Clin Cancer Res. 2011;17(15):5197–204.
Dittmann K, Mayer C, Rodemann HP. Nuclear EGFR as novel therapeutic target: insights into nuclear translocation and function. Strahlenther Onkol. 2010;186(1):1–6.
Licitra L, Mesia R, Rivera F, Remenar E, Hitt R, Erfan J, et al. Evaluation of EGFR gene copy number as a predictive biomarker for the efficacy of cetuximab in combination with chemotherapy in the first-line treatment of recurrent and/or metastatic squamous cell carcinoma of the head and neck: EXTREME study. Ann Oncol. 2011;22(5):1078–87.
Rivera F, Garcia-Castano A, Vega N, Vega-Villegas ME, Gutierrez-Sanz L. Cetuximab in metastatic or recurrent head and neck cancer: the EXTREME trial. Expert Rev Anticancer Ther. 2009;9(10):1421–8.
Van Damme N, Deron P, Van Roy N, Demetter P, Bols A, Van Dorpe J, et al. Epidermal growth factor receptor and K-RAS status in two cohorts of squamous cell carcinomas. BMC Cancer. 2010;10:189.
Willmore-Payne C, Holden JA, Layfield LJ. Detection of EGFR- and HER2-activating mutations in squamous cell carcinoma involving the head and neck. Mod Pathol. 2006;19(5):634–40.
Loeffler-Ragg J, Witsch-Baumgartner M, Tzankov A, Hilbe W, Schwentner I, Sprinzl GM, et al. Low incidence of mutations in EGFR kinase domain in Caucasian patients with head and neck squamous cell carcinoma. Eur J Cancer. 2006;42(1):109–11.
Murray S, Bobos M, Angouridakis N, Nikolaou A, Linardou H, Razis E, et al. Screening for EGFR mutations in patients with head and neck cancer treated with gefitinib on a compassionate-use program: a hellenic cooperative oncology group study. J Oncol. 2010;2010:709678.
Hama T, Yuza Y, Saito Y, Ou J, Kondo S, Okabe M, et al. Prognostic significance of epidermal growth factor receptor phosphorylation and mutation in head and neck squamous cell carcinoma. Oncologist. 2009;14(9):900–8.
Hama T, Yuza Y, Suda T, Saito Y, Norizoe C, Kato T, et al. Functional mutation analysis of EGFR family genes and corresponding lymph node metastases in head and neck squamous cell carcinoma. Clin Exp Metastasis. 2011.
Szabo B, Nelhubel GA, Karpati A, Kenessey I, Jori B, Szekely C, et al. Clinical significance of genetic alterations and expression of epidermal growth factor receptor (EGFR) in head and neck squamous cell carcinomas. Oral Oncol. 2011;47(6):487–96.
Cohen EE, Lingen MW, Martin LE, Harris PL, Brannigan BW, Haserlat SM, et al. Response of some head and neck cancers to epidermal growth factor receptor tyrosine kinase inhibitors may be linked to mutation of ERBB2 rather than EGFR. Clin Cancer Res. 2005;11(22):8105–8.
Chung CH, Ely K, McGavran L, Varella-Garcia M, Parker J, Parker N, et al. Increased epidermal growth factor receptor gene copy number is associated with poor prognosis in head and neck squamous cell carcinomas. J Clin Oncol. 2006;24(25):4170–6.
Lee JW, Soung YH, Kim SY, Nam HK, Park WS, Nam SW, et al. Somatic mutations of EGFR gene in squamous cell carcinoma of the head and neck. Clin Cancer Res. 2005;11(8):2879–82.
Lemos-Gonzalez Y, Paez de la Cadena M, Rodriguez-Berrocal FJ, Rodriguez-Pineiro AM, Pallas E, Valverde D. Absence of activating mutations in the EGFR kinase domain in Spanish head and neck cancer patients. Tumour Biol. 2007;28(5):273–9.
Huang SF, Chuang WY, Chen IH, Liao CT, Wang HM, Hsieh LL. EGFR protein overexpression and mutation in areca quid-associated oral cavity squamous cell carcinoma in Taiwan. Head Neck. 2009;31(8):1068–77.
Schwentner I, Witsch-Baumgartner M, Sprinzl GM, Krugmann J, Tzankov A, Jank S, et al. Identification of the rare EGFR mutation p.G796S as somatic and germline mutation in white patients with squamous cell carcinoma of the head and neck. Head Neck. 2008;30(8):1040–4.
Sheikh Ali MA, Gunduz M, Nagatsuka H, Gunduz E, Cengiz B, Fukushima K, et al. Expression and mutation analysis of epidermal growth factor receptor in head and neck squamous cell carcinoma. Cancer Sci. 2008;99(8):1589–94.
Hsieh CH, Chang JW, Hsieh JJ, Hsu T, Huang SF, Liao CT, et al. Epidermal growth factor receptor mutations in patients with oral cavity cancer in a betel nut chewing-prevalent area. Head Neck. 2011;33(12):1758–64.
Na II, Kang HJ, Cho SY, Koh JS, Lee JK, Lee BC, et al. EGFR mutations and human papillomavirus in squamous cell carcinoma of tongue and tonsil. Eur J Cancer. 2007;43(3):520–6.
Perisanidis C. Prevalence of EGFR tyrosine kinase domain mutations in head and neck squamous cell carcinoma: cohort study and systematic review. In Vivo. 2017;31(1):23–34.
Cassell A, Grandis JR. Investigational EGFR-targeted therapy in head and neck squamous cell carcinoma. Expert Opin Investig Drugs. 2010;19(6):709–22.
Uribe P, Gonzalez S. Epidermal growth factor receptor (EGFR) and squamous cell carcinoma of the skin: molecular bases for EGFR-targeted therapy. Pathol Res Pract. 2011;207(6):337–42.
Sok JC, Coppelli FM, Thomas SM, Lango MN, Xi S, Hunt JL, et al. Mutant epidermal growth factor receptor (EGFRvIII) contributes to head and neck cancer growth and resistance to EGFR targeting. Clin Cancer Res. 2006;12(17):5064–73.
Chau NG, Perez-Ordonez B, Zhang K, Pham NA, Ho J, Zhang T, et al. The association between EGFR variant III, HPV, p16, c-MET, EGFR gene copy number and response to EGFR inhibitors in patients with recurrent or metastatic squamous cell carcinoma of the head and neck. Head Neck Oncol. 2011;3:11.
Pectasides E, Fountzilas G, Kountourakis P, Gouveris P, Sasaki C, Duffey D, et al. Evalutaion of the incidence and prognostic value of mutant epidermal growth factor receptor (EGFRvIII) protein expression in head and neck squamous cell carcinomas (HNSCC) using AQUA. J Clin Oncol. 2010;28:15s.
McIntyre JB, Bose P, Klimowicz AC, Brockton NT, Petrillo S, Matthews W, et al. Specific and sensitive hydrolysis probe-based real-time PCR detection of epidermal growth factor receptor variant III in oral squamous cell carcinoma. PLoS One. 2012;7(2):e31723.
Liccardi G, Hartley JA, Hochhauser D. EGFR nuclear translocation modulates DNA repair following cisplatin and ionizing radiation treatment. Cancer Res. 2011;71(3):1103–14.
Lo HW, Xia W, Wei Y, Ali-Seyed M, Huang SF, Hung MC. Novel prognostic value of nuclear epidermal growth factor receptor in breast cancer. Cancer Res. 2005;65(1):338–48.
Wang SC, Nakajima Y, Yu YL, Xia W, Chen CT, Yang CC, et al. Tyrosine phosphorylation controls PCNA function through protein stability. Nat Cell Biol. 2006;8(12):1359–68.
Lin SY, Makino K, Xia W, Matin A, Wen Y, Kwong KY, et al. Nuclear localization of EGF receptor and its potential new role as a transcription factor. Nat Cell Biol. 2001;3(9):802–8.
Hanada N, Lo HW, Day CP, Pan Y, Nakajima Y, Hung MC. Co-regulation of B-Myb expression by E2F1 and EGF receptor. Mol Carcinog. 2006;45(1):10–7.
Hung LY, Tseng JT, Lee YC, Xia W, Wang YN, Wu ML, et al. Nuclear epidermal growth factor receptor (EGFR) interacts with signal transducer and activator of transcription 5 (STAT5) in activating Aurora-A gene expression. Nucleic Acids Res. 2008;36(13):4337–51.
Lo HW, Hsu SC, Ali-Seyed M, Gunduz M, Xia W, Wei Y, et al. Nuclear interaction of EGFR and STAT3 in the activation of the iNOS/NO pathway. Cancer Cell. 2005;7(6):575–89.
Psyrri A, Yu Z, Weinberger PM, Sasaki C, Haffty B, Camp R, et al. Quantitative determination of nuclear and cytoplasmic epidermal growth factor receptor expression in oropharyngeal squamous cell cancer by using automated quantitative analysis. Clin Cancer Res. 2005;11(16):5856–62.
Hoshino M, Fukui H, Ono Y, Sekikawa A, Ichikawa K, Tomita S, et al. Nuclear expression of phosphorylated EGFR is associated with poor prognosis of patients with esophageal squamous cell carcinoma. Pathobiol. 2007;74(1):15–21.
Lievre A, Bachet JB, Le Corre D, Boige V, Landi B, Emile JF, et al. KRAS mutation status is predictive of response to cetuximab therapy in colorectal cancer. Cancer Res. 2006;66(8):3992–5.
Amado RG, Wolf M, Peeters M, Van Cutsem E, Siena S, Freeman DJ, et al. Wild-type KRAS is required for panitumumab efficacy in patients with metastatic colorectal cancer. J Clin Oncol. 2008;26(10):1626–34.
De Roock W, Claes B, Bernasconi D, De Schutter J, Biesmans B, Fountzilas G, et al. Effects of KRAS, BRAF, NRAS, and PIK3CA mutations on the efficacy of cetuximab plus chemotherapy in chemotherapy-refractory metastatic colorectal cancer: a retrospective consortium analysis. Lancet Oncol. 2010;11(8):753–62.
Rebucci M, Peixoto P, Dewitte A, Wattez N, De Nuncques MA, Rezvoy N, et al. Mechanisms underlying resistance to cetuximab in the HNSCC cell line: role of AKT inhibition in bypassing this resistance. Int J Oncol. 2011;38(1):189–200.
Yamatodani T, Ekblad L, Kjellen E, Johnsson A, Mineta H, Wennerberg J. Epidermal growth factor receptor status and persistent activation of Akt and p44/42 MAPK pathways correlate with the effect of cetuximab in head and neck and colon cancer cell lines. J Cancer Res Clin Oncol. 2009;135(3):395–402.
Mriouah J, Boura C, Pinel S, Chretien AS, Fifre A, Merlin JL, et al. Cellular response to cetuximab in PTEN-silenced head and neck squamous cell carcinoma cell line. Int J Oncol. 2010;37(6):1555–63.
Kondo N, Tsukuda M, Taguchi T, Nakazaki K, Sakakibara A, Takahashi H, et al. Gene status of head and neck squamous cell carcinoma cell lines and cetuximab-mediated biological activities. Cancer Sci. 2011;102(9):1717–23.
Bonner JA, Yang ES, Trummell HQ, Nowsheen S, Willey CD, Raisch KP. Inhibition of STAT-3 results in greater cetuximab sensitivity in head and neck squamous cell carcinoma. Radiother Oncol. 2011;99(3):339–43.
Koppikar P, Lui VW, Man D, Xi S, Chai RL, Nelson E, et al. Constitutive activation of signal transducer and activator of transcription 5 contributes to tumor growth, epithelial-mesenchymal transition, and resistance to epidermal growth factor receptor targeting. Clin Cancer Res. 2008;14(23):7682–90.
Leeman-Neill RJ, Wheeler SE, Singh SV, Thomas SM, Seethala RR, Neill DB, et al. Guggulsterone enhances head and neck cancer therapies via inhibition of signal transducer and activator of transcription-3. Carcinogenesis. 2009;30(11):1848–56.
Wheeler DL, Iida M, Kruser TJ, Nechrebecki MM, Dunn EF, Armstrong EA, et al. Epidermal growth factor receptor cooperates with Src family kinases in acquired resistance to cetuximab. Cancer Biol Ther. 2009;8(8):696–703.
Kloth MT, Laughlin KK, Biscardi JS, Boerner JL, Parsons SJ, Silva CM. STAT5b, a mediator of synergism between c-Src and the epidermal growth factor receptor. J Biol Chem. 2003;278(3):1671–9.
Boeckx C, Op de Beeck K, Wouters A, Deschoolmeester V, Limame R, Zwaenepoel K, et al. Overcoming cetuximab resistance in HNSCC: The role of AURKB and DUSP proteins. Cancer Lett. 2014;354(2):365–77.
Oliveras-Ferraros C, Vazquez-Martin A, Queralt B, Adrados M, Ortiz R, Cufi S, et al. Interferon/STAT1 and neuregulin signaling pathways are exploratory biomarkers of cetuximab (Erbitux(R)) efficacy in KRAS wild-type squamous carcinomas: a pathway-based analysis of whole human-genome microarray data from cetuximab-adapted tumor cell-line models. Int J Oncol. 2011;39(6):1455–79.
Ercan D, Xu C, Yanagita M, Monast CS, Pratilas CA, Montero J, et al. Reactivation of ERK signaling causes resistance to EGFR kinase inhibitors. Cancer Discov. 2012.
Gazel A, Nijhawan RI, Walsh R, Blumenberg M. Transcriptional profiling defines the roles of ERK and p38 kinases in epidermal keratinocytes. J Cell Physiol. 2008;215(2):292–308.
Davies H, Bignell GR, Cox C, Stephens P, Edkins S, Clegg S, et al. Mutations of the BRAF gene in human cancer. Nature. 2002;417(6892):949–54.
Bos JL. ras oncogenes in human cancer: a review. Cancer Res. 1989;49(17):4682–9.
Lievre A, Bachet JB, Boige V, Cayre A, Le Corre D, Buc E, et al. KRAS mutations as an independent prognostic factor in patients with advanced colorectal cancer treated with cetuximab. J Clin Oncol. 2008;26(3):374–9.
De Roock W, Piessevaux H, De Schutter J, Janssens M, De Hertogh G, Personeni N, et al. KRAS wild-type state predicts survival and is associated to early radiological response in metastatic colorectal cancer treated with cetuximab. Ann Oncol. 2008;19(3):508–15.
Deschoolmeester V, Boeckx C, Baay M, Weyler J, Wuyts W, Van Marck E, et al. KRAS mutation detection and prognostic potential in sporadic colorectal cancer using high-resolution melting analysis. Br J Cancer. 2010;103(10):1627–36.
Andreyev HJ, Norman AR, Cunningham D, Oates JR, Clarke PA. Kirsten ras mutations in patients with colorectal cancer: the multicenter “RASCAL” study. J Natl Cancer Inst. 1998;90(9):675–84.
Weber A, Langhanki L, Sommerer F, Markwarth A, Wittekind C, Tannapfel A. Mutations of the BRAF gene in squamous cell carcinoma of the head and neck. Oncogene. 2003;22(30):4757–9.
Sheng ZM, Barrois M, Klijanienko J, Micheau C, Richard JM, Riou G. Analysis of the c-Ha-ras-1 gene for deletion, mutation, amplification and expression in lymph node metastases of human head and neck carcinomas. Br J Cancer. 1990;62(3):398–404.
Anderson JA, Irish JC, McLachlin CM, Ngan BY. H-ras oncogene mutation and human papillomavirus infection in oral carcinomas. Arch Otolaryngol Head Neck Surg. 1994;120(7):755–60.
Yarbrough WG, Shores C, Witsell DL, Weissler MC, Fidler ME, Gilmer TM. ras mutations and expression in head and neck squamous cell carcinomas. Laryngoscope. 1994;104(11 Pt 1):1337–47.
Rampias T, Giagini A, Florou K, Gouveris P, Vaja E, Haralambakis N, et al. H-RAS and PIK3CA mutations and response to cetuximab in head and neck squamous cell carcinoma (HNSCC). J Clin Oncol. 2011;29.
Rampias T, Giagini A, Matsuzaki H, Bartzi V, Siolos S, Vaja E, et al. Genetic alterations in HRAS gene in relation to outcoume and response to cetuximab in head and neck squamous cell carcinoma. J Clin Oncol. 2012;498:30.
Leblanc O, Vacher S, Lecerf C, Jeannot E, Klijanienko J, Berger F, et al. Biomarkers of cetuximab resistance in patients with head and neck squamous cell carcinoma. Cancer Biol Med. 2020;17(1):208–17.
Keyse SM. Protein phosphatases and the regulation of mitogen-activated protein kinase signalling. Curr Opin Cell Biol. 2000;12(2):186–92.
Arkell RS, Dickinson RJ, Squires M, Hayat S, Keyse SM, Cook SJ. DUSP6/MKP-3 inactivates ERK1/2 but fails to bind and inactivate ERK5. Cell Signal. 2008;20(5):836–43.
Wong VC, Chen H, Ko JM, Chan KW, Chan YP, Law S, et al. Tumor suppressor dual-specificity phosphatase 6 (DUSP6) impairs cell invasion and epithelial-mesenchymal transition (EMT)-associated phenotype. Int J Cancer. 2012;130(1):83–95.
Xu S, Furukawa T, Kanai N, Sunamura M, Horii A. Abrogation of DUSP6 by hypermethylation in human pancreatic cancer. J Hum Genet. 2005;50(4):159–67.
Okudela K, Yazawa T, Woo T, Sakaeda M, Ishii J, Mitsui H, et al. Down-regulation of DUSP6 expression in lung cancer: its mechanism and potential role in carcinogenesis. Am J Pathol. 2009;175(2):867–81.
Bermudez O, Pages G, Gimond C. The dual-specificity MAP kinase phosphatases: critical roles in development and cancer. Am J Physiol Cell Physiol. 2010;299(2):C189–202.
Liu P, Cheng H, Roberts TM, Zhao JJ. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Reviews Drug Discov. 2009;8(8):627–44.
Simpson DR, Mell LK, Cohen EEW. Targeting the PI3K/AKT/mTOR pathway in squamous cell carcinoma of the head and neck. Oral Oncol. 2015;51(4):291–8.
Castellano E, Downward J. Role of RAS in the regulation of PI 3-kinase. Curr Top Microbiol Immunol. 2010;346:143–69.
Vivanco I, Sawyers CL. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev Cancer. 2002;2(7):489–501.
Shaw RJ, Cantley LC. Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature. 2006;441(7092):424–30.
Bowles DW, Ma WW, Senzer N, Brahmer JR, Adjei AA, Davies M, et al. A multicenter phase 1 study of PX-866 in combination with docetaxel in patients with advanced solid tumours. Br J Cancer. 2013;109(5):1085–92.
Guix M, Faber AC, Wang SE, Olivares MG, Song Y, Qu S, et al. Acquired resistance to EGFR tyrosine kinase inhibitors in cancer cells is mediated by loss of IGF-binding proteins. J Clin Invest. 2008;118(7):2609–19.
Sequist LV, Waltman BA, Dias-Santagata D, Digumarthy S, Turke AB, Fidias P, et al. Genotypic and histological evolution of lung cancers acquiring resistance to EGFR inhibitors. Sci Transl Med. 2011;3(75):75ra26.
Donev IS, Wang W, Yamada T, Li Q, Takeuchi S, Matsumoto K, et al. Transient PI3K inhibition induces apoptosis and overcomes HGF-mediated resistance to EGFR-TKIs in EGFR mutant lung cancer. Clin Cancer Res. 2011;17(8):2260–9.
Sen B, Saigal B, Parikh N, Gallick G, Johnson FM. Sustained Src inhibition results in signal transducer and activator of transcription 3 (STAT3) activation and cancer cell survival via altered Janus-activated kinase-STAT3 binding. Cancer Res. 2009;69(5):1958–65.
Xi S, Zhang Q, Dyer KF, Lerner EC, Smithgall TE, Gooding WE, et al. Src kinases mediate STAT growth pathways in squamous cell carcinoma of the head and neck. J Biol Chem. 2003;278(34):31574–83.
Yang Z, Bagheri-Yarmand R, Wang RA, Adam L, Papadimitrakopoulou VV, Clayman GL, et al. The epidermal growth factor receptor tyrosine kinase inhibitor ZD1839 (Iressa) suppresses c-Src and Pak1 pathways and invasiveness of human cancer cells. Clin Cancer Res. 2004;10(2):658–67.
van Oijen MG, Rijksen G, ten Broek FW, Slootweg PJ. Overexpression of c-Src in areas of hyperproliferation in head and neck cancer, premalignant lesions and benign mucosal disorders. J Oral Pathol Med. 1998;27(4):147–52.
Dittmann K, Mayer C, Kehlbach R, Rodemann HP. Radiation-induced caveolin-1 associated EGFR internalization is linked with nuclear EGFR transport and activation of DNA-PK. Mol Cancer. 2008;7:69.
Li C, Iida M, Dunn EF, Wheeler DL. Dasatinib blocks cetuximab- and radiation-induced nuclear translocation of the epidermal growth factor receptor in head and neck squamous cell carcinoma. Radiother Oncol. 2010;97(2):330–7.
Nozaki M, Yasui H, Ohnishi Y. Ligand-independent EGFR activation by anchorage-stimulated Src promotes cancer cell proliferation and cetuximab resistance via ErbB3 phosphorylation. Cancers. 2019;11(10).
Lee TL, Yeh J, Van Waes C, Chen Z. Epigenetic modification of SOCS-1 differentially regulates STAT3 activation in response to interleukin-6 receptor and epidermal growth factor receptor signaling through JAK and/or MEK in head and neck squamous cell carcinomas. Mol Cancer Ther. 2006;5(1):8–19.
Kijima T, Niwa H, Steinman RA, Drenning SD, Gooding WE, Wentzel AL, et al. STAT3 activation abrogates growth factor dependence and contributes to head and neck squamous cell carcinoma tumor growth in vivo. Cell Growth Differ. 2002;13(8):355–62.
Onishi A, Chen Q, Humtsoe JO, Kramer RH. STAT3 signaling is induced by intercellular adhesion in squamous cell carcinoma cells. Exp Cell Res. 2008;314(2):377–86.
Hambek M, Baghi M, Strebhardt K, May A, Adunka O, Gstottner W, et al. STAT 3 activation in head and neck squamous cell carcinomas is controlled by the EGFR. Anticancer Res. 2004;24(6):3881–6.
Willey CD, Anderson JC, Trummell HQ, Naji F, de Wijn R, Yang ES, et al. Differential escape mechanisms in cetuximab-resistant head and neck cancer cells. Biochem Biophys Res Commun. 2019;517(1):36–42.
Quesnelle KM, Grandis JR. Dual kinase inhibition of EGFR and HER2 overcomes resistance to cetuximab in a novel in vivo model of acquired cetuximab resistance. Clin Cancer Res. 2011;17(18):5935–44.
Yonesaka K, Zejnullahu K, Okamoto I, Satoh T, Cappuzzo F, Souglakos J, et al. Activation of ERBB2 signaling causes resistance to the EGFR-directed therapeutic antibody cetuximab. Sci Transl Med. 2011;3(99):99ra86.
Barnes CJ, Ohshiro K, Rayala SK, El-Naggar AK, Kumar R. Insulin-like growth factor receptor as a therapeutic target in head and neck cancer. Clin Cancer Res. 2007;13(14):4291–9.
Zuo Q, Shi M, Li L, Chen J, Luo R. Development of cetuximab-resistant human nasopharyngeal carcinoma cell lines and mechanisms of drug resistance. Biomed Pharmacother. 2010;64(8):550–8.
Tandon R, Kapoor S, Vali S, Senthil V, Nithya D, Venkataramanan R, et al. Dual epidermal growth factor receptor (EGFR)/insulin-like growth factor-1 receptor (IGF-1R) inhibitor: a novel approach for overcoming resistance in anticancer treatment. Eur J Pharmacol. 2011;667(1-3):56–65.
Sierra JR, Tsao MS. c-MET as a potential therapeutic target and biomarker in cancer. Ther Adv Med Oncol. 2011;3(1 Suppl):S21–35.
Heindl S, Eggenstein E, Keller S, Kneissl J, Keller G, Mutze K, et al. Relevance of MET activation and genetic alterations of KRAS and E-cadherin for cetuximab sensitivity of gastric cancer cell lines. J Cancer Res Clin Oncol. 2012;138(5):843–58.
Pollak M. Insulin and insulin-like growth factor signalling in neoplasia. Nat Rev Cancer. 2008;8(12):915–28.
van der Veeken J, Oliveira S, Schiffelers RM, Storm G, van Bergen En Henegouwen PM, Roovers RC. Crosstalk between epidermal growth factor receptor- and insulin-like growth factor-1 receptor signaling: implications for cancer therapy. Curr Cancer Drug Targets. 2009;9(6):748–60.
Cancer Genome Atlas N. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature. 2015;517(7536):576–82.
Jameson MJ, Beckler AD, Taniguchi LE, Allak A, Vanwagner LB, Lee NG, et al. Activation of the insulin-like growth factor-1 receptor induces resistance to epidermal growth factor receptor antagonism in head and neck squamous carcinoma cells. Mol Cancer Ther. 2011;10(11):2124–34.
Iyer G, Price J, Bourgeois S, Armstrong E, Huang S, Harari PM. Insulin-like growth factor 1 receptor mediated tyrosine 845 phosphorylation of epidermal growth factor receptor in the presence of monoclonal antibody cetuximab. BMC Cancer. 2016;16(1):773.
Knowles LM, Stabile LP, Egloff AM, Rothstein ME, Thomas SM, Gubish CT, et al. HGF and c-Met participate in paracrine tumorigenic pathways in head and neck squamous cell cancer. Clin Cancer Res. 2009;15(11):3740–50.
Seiwert TY, Jagadeeswaran R, Faoro L, Janamanchi V, Nallasura V, El Dinali M, et al. The MET receptor tyrosine kinase is a potential novel therapeutic target for head and neck squamous cell carcinoma. Cancer Res. 2009;69(7):3021–31.
Novoplansky O, Fury M, Prasad M, Yegodayev K, Zorea J, Cohen L, et al. MET activation confers resistance to cetuximab, and prevents HER2 and HER3 upregulation in head and neck cancer. Int J Cancer. 2019;145(3):748–62.
Kalish LH, Kwong RA, Cole IE, Gallagher RM, Sutherland RL, Musgrove EA. Deregulated cyclin D1 expression is associated with decreased efficacy of the selective epidermal growth factor receptor tyrosine kinase inhibitor gefitinib in head and neck squamous cell carcinoma cell lines. Clin Cancer Res. 2004;10(22):7764–74.
Huang S, Benavente S, Armstrong EA, Li C, Wheeler DL, Harari PM. p53 modulates acquired resistance to EGFR inhibitors and radiation. Cancer Res. 2011;71(22):7071–9.
Ogino S, Meyerhardt JA, Cantor M, Brahmandam M, Clark JW, Namgyal C, et al. Molecular alterations in tumors and response to combination chemotherapy with gefitinib for advanced colorectal cancer. Clin Cancer Res. 2005;11(18):6650–6.
Huether A, Hopfner M, Baradari V, Schuppan D, Scherubl H. EGFR blockade by cetuximab alone or as combination therapy for growth control of hepatocellular cancer. Biochem Pharmacol. 2005;70(11):1568–78.
Hoellein A, Pickhard A, von Keitz F, Schoeffmann S, Piontek G, Rudelius M, et al. Aurora kinase inhibition overcomes cetuximab resistance in squamous cell cancer of the head and neck. Oncotarget. 2011;2(8):599–609.
Kelly KR, Ecsedy J, Mahalingam D, Nawrocki ST, Padmanabhan S, Giles FJ, et al. Targeting aurora kinases in cancer treatment. Curr Drug Targets. 2011;12(14):2067–78.
McLaughlin J, Markovtsov V, Li H, Wong S, Gelman M, Zhu Y, et al. Preclinical characterization of Aurora kinase inhibitor R763/AS703569 identified through an image-based phenotypic screen. J Cancer Res Clin Oncol. 2010;136(1):99–113.
Reiter R, Gais P, Jutting U, Steuer-Vogt MK, Pickhard A, Bink K, et al. Aurora kinase A messenger RNA overexpression is correlated with tumor progression and shortened survival in head and neck squamous cell carcinoma. Clin Cancer Res. 2006;12(17):5136–41.
Lens SM, Voest EE, Medema RH. Shared and separate functions of polo-like kinases and aurora kinases in cancer. Nat Rev Cancer. 2010;10(12):825–41.
Lai CH, Tseng JT, Lee YC, Chen YJ, Lee JC, Lin BW, et al. Translational up-regulation of Aurora-A in EGFR-overexpressed cancer. J Cell Mol Med. 2010;14(6B):1520–31.
Wu CC, Yu CT, Chang GC, Lai JM, Hsu SL. Aurora-A promotes gefitinib resistance via a NF-kappaB signaling pathway in p53 knockdown lung cancer cells. Biochem Biophys Res Commun. 2011;405(2):168–72.
Arany I, Chen SH, Megyesi JK, Adler-Storthz K, Chen Z, Rajaraman S, et al. Differentiation-dependent expression of signal transducers and activators of transcription (STATs) might modify responses to growth factors in the cancers of the head and neck. Cancer Lett. 2003;199(1):83–9.
Adkins D, Ley J, Neupane P, Worden F, Sacco AG, Palka K, et al. Palbociclib and cetuximab in platinum-resistant and in cetuximab-resistant human papillomavirus-unrelated head and neck cancer: a multicentre, multigroup, phase 2 trial. Lancet Oncol. 2019;20(9):1295–305.
Byers LA, Diao L, Wang J, Saintigny P, Girard L, Peyton M, et al. An epithelial-mesenchymal transition gene signature predicts resistance to EGFR and PI3K inhibitors and identifies Axl as a therapeutic target for overcoming EGFR inhibitor resistance. Clin Cancer Res. 2013;19(1):279–90.
Haddad Y, Choi W, McConkey DJ. Delta-crystallin enhancer binding factor 1 controls the epithelial to mesenchymal transition phenotype and resistance to the epidermal growth factor receptor inhibitor erlotinib in human head and neck squamous cell carcinoma lines. Clin Cancer Res. 2009;15(2):532–42.
Skvortsova I, Skvortsov S, Raju U, Stasyk T, Riesterer O, Schottdorf EM, et al. Epithelial-to-mesenchymal transition and c-myc expression are the determinants of cetuximab-induced enhancement of squamous cell carcinoma radioresponse. Radiother Oncol. 2010;96(1):108–15.
Frederick BA, Helfrich BA, Coldren CD, Zheng D, Chan D, Bunn PA Jr, et al. Epithelial to mesenchymal transition predicts gefitinib resistance in cell lines of head and neck squamous cell carcinoma and non-small cell lung carcinoma. Mol Cancer Ther. 2007;6(6):1683–91.
Thomson S, Buck E, Petti F, Griffin G, Brown E, Ramnarine N, et al. Epithelial to mesenchymal transition is a determinant of sensitivity of non-small-cell lung carcinoma cell lines and xenografts to epidermal growth factor receptor inhibition. Cancer Res. 2005;65(20):9455–62.
Yauch RL, Januario T, Eberhard DA, Cavet G, Zhu W, Fu L, et al. Epithelial versus mesenchymal phenotype determines in vitro sensitivity and predicts clinical activity of erlotinib in lung cancer patients. Clin Cancer Res. 2005;11(24 Pt 1):8686–98.
Buck E, Eyzaguirre A, Barr S, Thompson S, Sennello R, Young D, et al. Loss of homotypic cell adhesion by epithelial-mesenchymal transition or mutation limits sensitivity to epidermal growth factor receptor inhibition. Mol Cancer Ther. 2007;6(2):532–41.
Holz C, Niehr F, Boyko M, Hristozova T, Distel L, Budach V, et al. Epithelial-mesenchymal-transition induced by EGFR activation interferes with cell migration and response to irradiation and cetuximab in head and neck cancer cells. Radiother Oncol. 2011;101(1):158–64.
Boeckx C, Blockx L, de Beeck KO, Limame R, Camp GV, Peeters M, et al. Establishment and characterization of cetuximab resistant head and neck squamous cell carcinoma cell lines: focus on the contribution of the AP-1 transcription factor. Am J Cancer Res. 2015;5(6):1921–38.
Cowling VH, Cole MD. E-cadherin repression contributes to c-Myc-induced epithelial cell transformation. Oncogene. 2007;26(24):3582–6.
Thiery JP. Epithelial-mesenchymal transitions in development and pathologies. Curr Opin Cell Biol. 2003;15(6):740–6.
Guarino M. Epithelial-mesenchymal transition and tumour invasion. Int J Biochem Cell Biol. 2007;39(12):2153–60.
Basu D, Bewley AF, Sperry SM, Montone KT, Gimotty PA, Rasanen K, et al. EGFR inhibition promotes an aggressive invasion pattern mediated by mesenchymal-like tumor cells within squamous cell carcinomas. Mol Cancer Ther. 2013;12(10):2176–86.
Smith JP, Pozzi A, Dhawan P, Singh AB, Harris RC. Soluble HB-EGF induces epithelial-to-mesenchymal transition in inner medullary collecting duct cells by upregulating Snail-2. Am J Physiol Renal Physiol. 2009;296(5):F957–65.
Yagi H, Yotsumoto F, Miyamoto S. Heparin-binding epidermal growth factor-like growth factor promotes transcoelomic metastasis in ovarian cancer through epithelial-mesenchymal transition. Mol Cancer Ther. 2008;7(10):3441–51.
Wang F, Sloss C, Zhang X, Lee SW, Cusack JC. Membrane-bound heparin-binding epidermal growth factor like growth factor regulates E-cadherin expression in pancreatic carcinoma cells. Cancer Res. 2007;67(18):8486–93.
Li XJ, Peng LX, Shao JY, Lu WH, Zhang JX, Chen S, et al. As an independent unfavorable prognostic factor, IL-8 promotes metastasis of nasopharyngeal carcinoma through induction of epithelial-mesenchymal transition and activation of AKT signaling. Carcinogenesis. 2012;33(7):1302–9.
Fernando RI, Castillo MD, Litzinger M, Hamilton DH, Palena C. IL-8 signaling plays a critical role in the epithelial-mesenchymal transition of human carcinoma cells. Cancer Res. 2011;71(15):5296–306.
Hwang WL, Yang MH, Tsai ML, Lan HY, Su SH, Chang SC, et al. SNAIL regulates interleukin-8 expression, stem cell-like activity, and tumorigenicity of human colorectal carcinoma cells. Gastroenterol. 2011;141(1):279–91, 91 e1-5.
Shimura T, Yoshida M, Fukuda S, Ebi M, Hirata Y, Mizoshita T, et al. Nuclear translocation of the cytoplasmic domain of HB-EGF induces gastric cancer invasion. BMC Cancer. 2012;12:205.
Lin Y, Buckhaults PJ, Lee JR, Xiong H, Farrell C, Podolsky RH, et al. Association of the actin-binding protein transgelin with lymph node metastasis in human colorectal cancer. Neoplasia. 2009;11(9):864–73.
Jo M, Eastman BM, Webb DL, Stoletov K, Klemke R, Gonias SL. Cell signaling by urokinase-type plasminogen activator receptor induces stem cell-like properties in breast cancer cells. Cancer Res. 2010;70(21):8948–58.
Noh H, Hong S, Huang S. Role of urokinase receptor in tumor progression and development. Theranostics. 2013;3(7):487–95.
Seals DF, Courtneidge SA. The ADAMs family of metalloproteases: multidomain proteins with multiple functions. Genes Dev. 2003;17(1):7–30.
Qian X, Tuszynski GP. Expression of thrombospondin-1 in cancer: a role in tumor progression. Proc Soc Exp Biol Med. 1996;212(3):199–207.
Holz C, Niehr F, Boyko M, Hristozova T, Distel L, Budach V, et al. Epithelial-mesenchymal-transition induced by EGFR activation interferes with cell migration and response to irradiation and cetuximab in head and neck cancer cells. Radiother Oncol. 2011.
Vaupel P, Mayer A. Hypoxia in cancer: significance and impact on clinical outcome. Cancer Metastasis Rev. 2007;26(2):225–39.
Bertout JA, Patel SA, Simon MC. The impact of O2 availability on human cancer. Nat Rev Cancer. 2008;8(12):967–75.
Wouters A, Pauwels B, Lardon F, Vermorken JB. Review: implications of in vitro research on the effect of radiotherapy and chemotherapy under hypoxic conditions. Oncologist. 2007;12(6):690–712.
Wouters A, Boeckx C, Vermorken JB, den Weyngaert DV, Peeters M, Lardon F. The intriguing interplay between therapies targeting the epidermal growth factor receptor, the hypoxic microenvironment and hypoxia-inducible factors. Curr Pharm Des. 2012 [Epub ahead of print].
Wouters A, Boeckx C, Vermorken JB, Van den Weyngaert D, Peeters M, Lardon F. The intriguing interplay between therapies targeting the epidermal growth factor receptor, the hypoxic microenvironment and hypoxia-inducible factors. Curr Pharm Des. 2013;19(5):907–17.
Byers LA, Holsinger FC, Kies MS, William WN, El-Naggar AK, Lee JJ, et al. Serum signature of hypoxia-regulated factors is associated with progression after induction therapy in head and neck squamous cell cancer. Mol Cancer Ther. 2010;9(6):1755–63.
Lee CM, Tannock IF. The distribution of the therapeutic monoclonal antibodies cetuximab and trastuzumab within solid tumors. BMC Cancer. 2010;10:255.
Santiago A, Eicheler W, Bussink J, Rijken P, Yaromina A, Beuthien-Baumann B, et al. Effect of cetuximab and fractionated irradiation on tumour micro-environment. Radiother Oncol. 2010.
Bier H, Reiffen KA, Haas I, Stasiecki P. Dose-dependent access of murine anti-epidermal growth factor receptor monoclonal antibody to tumor cells in patients with advanced laryngeal and hypopharyngeal carcinoma. Eur Archiv Oto-rhino-laryngol. 1995;252(7):433–9.
Petit AM, Rak J, Hung MC, Rockwell P, Goldstein N, Fendly B, et al. Neutralizing antibodies against epidermal growth factor and ErbB-2/neu receptor tyrosine kinases down-regulate vascular endothelial growth factor production by tumor cells in vitro and in vivo: angiogenic implications for signal transduction therapy of solid tumors. Am J Pathol. 1997;151(6):1523–30.
Perrotte P, Matsumoto T, Inoue K, Kuniyasu H, Eve BY, Hicklin DJ, et al. Anti-epidermal growth factor receptor antibody C225 inhibits angiogenesis in human transitional cell carcinoma growing orthotopically in nude mice. Clin Cancer Res. 1999;5(2):257–65.
Riesterer O, Mason KA, Raju U, Yang Q, Wang L, Hittelman WN, et al. Enhanced response to C225 of A431 tumor xenografts growing in irradiated tumor bed. Radiother Oncol. 2009;92(3):383–7.
Luwor RB, Lu Y, Li X, Mendelsohn J, Fan Z. The antiepidermal growth factor receptor monoclonal antibody cetuximab/C225 reduces hypoxia-inducible factor-1 alpha, leading to transcriptional inhibition of vascular endothelial growth factor expression. Oncogene. 2005;24(27):4433–41.
Huang SM, Harari PM. Modulation of radiation response after epidermal growth factor receptor blockade in squamous cell carcinomas: inhibition of damage repair, cell cycle kinetics, and tumor angiogenesis. Clin Cancer Res. 2000;6(6):2166–74.
Boeckx C, Van den Bossche J, De Pauw I, Peeters M, Lardon F, Baay M, et al. The hypoxic tumor microenvironment and drug resistance against EGFR inhibitors: preclinical study in cetuximab-sensitive head and neck squamous cell carcinoma cell lines. BMC Res Notes. 2015;8:203.
Krause M, Ostermann G, Petersen C, Yaromina A, Hessel F, Harstrick A, et al. Decreased repopulation as well as increased reoxygenation contribute to the improvement in local control after targeting of the EGFR by C225 during fractionated irradiation. Radiother Oncol. 2005;76(2):162–7.
Lu Y, Liang K, Li X, Fan Z. Responses of cancer cells with wild-type or tyrosine kinase domain-mutated epidermal growth factor receptor (EGFR) to EGFR-targeted therapy are linked to downregulation of hypoxia-inducible factor-1alpha. Mol Cancer. 2007;6:63.
Li X, Lu Y, Liang K, Pan T, Mendelsohn J, Fan Z. Requirement of hypoxia-inducible factor-1alpha down-regulation in mediating the antitumor activity of the anti-epidermal growth factor receptor monoclonal antibody cetuximab. Mol Cancer Ther. 2008;7(5):1207–17.
Li X, Fan Z. The epidermal growth factor receptor antibody cetuximab induces autophagy in cancer cells by downregulating HIF-1alpha and Bcl-2 and activating the beclin 1/hVps34 complex. Cancer Res. 2010;70(14):5942–52.
Lu H, Liang K, Lu Y, Fan Z. The anti-EGFR antibody cetuximab sensitizes human head and neck squamous cell carcinoma cells to radiation in part through inhibiting radiation-induced upregulation of HIF-1alpha. Cancer Lett. 2012.
Lu Y, Li X, Lu H, Fan Z. 1, 9-Pyrazoloanthrones downregulate HIF-1alpha and sensitize cancer cells to cetuximab-mediated anti-EGFR therapy. PLoS One. 2010;5(12):e15823.
Luo HY, Wei W, Shi YX, Chen XQ, Li YH, Wang F, et al. Cetuximab enhances the effect of oxaliplatin on hypoxic gastric cancer cell lines. Oncol Rep. 2010;23(6):1735–45.
Shepard HM, Brdlik CM, Schreiber H. Signal integration: a framework for understanding the efficacy of therapeutics targeting the human EGFR family. J Clin Invest. 2008;118(11):3574–81.
De Pauw I, Wouters A, Van den Bossche J, Deschoolmeester V, Baysal H, Pauwels P, et al. Dual targeting of epidermal growth factor receptor and HER3 by MEHD7945A as monotherapy or in combination with cisplatin partially overcomes cetuximab resistance in head and neck squamous cell carcinoma cell lines. Cancer Biother Radiopharm. 2017;32(7):229–38.
Hill AG, Findlay MP, Burge ME, Jackson C, Alfonso PG, Samuel L, et al. Phase II study of the dual EGFR/HER3 inhibitor duligotuzumab (MEHD7945A) versus cetuximab in combination with FOLFIRI in second-line RAS wild-type metastatic colorectal cancer. Clin Cancer Res. 2018;24(10):2276–84.
Li D, Ambrogio L, Shimamura T, Kubo S, Takahashi M, Chirieac LR, et al. BIBW2992, an irreversible EGFR/HER2 inhibitor highly effective in preclinical lung cancer models. Oncogene. 2008;27(34):4702–11.
Minkovsky N, Berezov A. BIBW-2992, a dual receptor tyrosine kinase inhibitor for the treatment of solid tumors. Curr Opin Investigat Drugs (London, England : 2000). 2008;9(12):1336–46.
Solca F, Dahl G, Zoephel A, Bader G, Sanderson M, Klein C, et al. Target binding properties and cellular activity of afatinib (BIBW 2992), an irreversible ErbB family blocker. J Pharmacol Exp Ther. 2012;343(2):342–50.
De Pauw I, Wouters A, Van den Bossche J, Peeters M, Pauwels P, Deschoolmeester V, et al. Preclinical and clinical studies on afatinib in monotherapy and in combination regimens: potential impact in colorectal cancer. Pharmacol Ther. 2016;166:71–83.
Ioannou N, Seddon AM, Dalgleish A, Mackintosh D, Modjtahedi H. Treatment with a combination of the ErbB (HER) family blocker afatinib and the IGF-IR inhibitor, NVP-AEW541 induces synergistic growth inhibition of human pancreatic cancer cells. BMC Cancer. 2013;13:41.
Young NR, Soneru C, Liu J, Grushko TA, Hardeman A, Olopade OI, et al. Afatinib efficacy against squamous cell carcinoma of the head and neck cell lines in vitro and in vivo. Targeted Oncol. 2015;10(4):501–8.
De Pauw I, Lardon F, Van den Bossche J, Baysal H, Fransen E, Deschoolmeester V, et al. Simultaneous targeting of EGFR, HER2, and HER4 by afatinib overcomes intrinsic and acquired cetuximab resistance in head and neck squamous cell carcinoma cell lines. Mol Oncol. 2018;12(6):830–54.
Seiwert TY, Fayette J, Cupissol D, Del Campo JM, Clement PM, Hitt R, et al. A randomized, phase II study of afatinib versus cetuximab in metastatic or recurrent squamous cell carcinoma of the head and neck. Ann Oncol. 2014;25(9):1813–20.
Machiels JP, Haddad RI, Fayette J, Licitra LF, Tahara M, Vermorken JB, et al. Afatinib versus methotrexate as second-line treatment in patients with recurrent or metastatic squamous-cell carcinoma of the head and neck progressing on or after platinum-based therapy (LUX-Head & Neck 1): an open-label, randomised phase 3 trial. Lancet Oncol. 2015;16(5):583–94.
Cohen EEW, Licitra LF, Burtness B, Fayette J, Gauler T, Clement PM, et al. Biomarkers predict enhanced clinical outcomes with afatinib versus methotrexate in patients with second-line recurrent and/or metastatic head and neck cancer. Ann Oncol. 2017;28(10):2526–32.
Gazzah A, Boni V, Soria JC, Calles A, Even C, Doger B, et al. A phase 1b study of afatinib in combination with standard-dose cetuximab in patients with advanced solid tumours. Eur J Cancer. 2018;104:1–8.
Machiels JP, Bossi P, Menis J, Lia M, Fortpied C, Liu Y, et al. Activity and safety of afatinib in a window preoperative EORTC study in patients with squamous cell carcinoma of the head and neck (SCCHN). Ann Oncol. 2018;29(4):985–91.
Yang J, Li S, Wang B, Wu Y, Chen Z, Lv M, et al. Potential biomarkers for anti-EGFR therapy in metastatic colorectal cancer. Tumour Biol. 2016;37(9):11645–55.
Sacco AG, Cohen EE. Current treatment options for recurrent or metastatic head and neck squamous cell carcinoma. J Clin Oncol. 2015;33(29):3305–13.
Burtness B, Harrington KJ, Greil R, Soulieres D, Tahara M, de Castro G, et al. Pembrolizumab alone or with chemotherapy versus cetuximab with chemotherapy for recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-048): a randomised, open-label, phase 3 study. Lancet. 2019;394(10212):1915–28.
Kimura H, Sakai K, Arao T, Shimoyama T, Tamura T, Nishio K. Antibody-dependent cellular cytotoxicity of cetuximab against tumor cells with wild-type or mutant epidermal growth factor receptor. Cancer Sci. 2007;98(8):1275–80.
Yang X, Zhang X, Mortenson ED, Radkevich-Brown O, Wang Y, Fu YX. Cetuximab-mediated tumor regression depends on innate and adaptive immune responses. Mol Ther. 2013;21(1):91–100.
Ferris RL, Lenz HJ, Trotta AM, Garcia-Foncillas J, Schulten J, Audhuy F, et al. Rationale for combination of therapeutic antibodies targeting tumor cells and immune checkpoint receptors: harnessing innate and adaptive immunity through IgG1 isotype immune effector stimulation. Cancer Treatment Rev. 2018;63:48–60.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2021 The Author(s)
About this paper

Cite this paper
De Pauw, I., Boeckx, C., Wouters, A. (2021). Mechanisms of Cetuximab Resistance and How to Overcome It. In: Vermorken, J.B., Budach, V., Leemans, C.R., Machiels, JP., Nicolai, P., O’Sullivan, B. (eds) Critical Issues in Head and Neck Oncology. Springer, Cham. https://doi.org/10.1007/978-3-030-63234-2_3
Download citation
DOI: https://doi.org/10.1007/978-3-030-63234-2_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-63233-5
Online ISBN: 978-3-030-63234-2
eBook Packages: MedicineMedicine (R0)