
Abstract
Targeted mouse mutants are a common tool used to investigate gene function. The International Knockout Mouse Consortium undertook a large-scale screen of mouse mutants, making use of the knockout-first allele design that contains the En2 splice acceptor sequence coupled to the lacZ reporter gene. Although the knockout-first allele was designed to interfere with splicing and thus disrupt gene function, the En2 sequence has been reported to be transcribed within the host gene mRNA due to a cryptic splice site within the En2 sequence which allows splicing to the next exon of the host gene. In some circumstances, this has the potential to permit translation of a mutant protein. Here, we describe our computational analysis of all the mouse protein-coding genes with established knockout-first embryonic stem cell lines, and our predictions of their transcription outcome should the En2 sequence be included. As part of the large-scale mutagenesis program, mutant mice underwent a broad phenotyping screen, and their phenotypes are available. No wide-scale effects on mouse phenotypes reported were found as a result of the predicted En2 insertion. However, the En2 insertion was found experimentally in the transcripts of 24 of 35 mutant alleles examined, including the five already described, two with evidence of readthrough. Splicing from the cryptic splice site also has the potential to disrupt expression of the lacZ reporter gene. It is recommended that mutant transcripts be checked for this insertion as well as for leaky transcription in studies involving knockout-first alleles.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The mouse genome is reported to contain approximately 22,000 protein-coding genes (http://www.ensembl.org/Mus_musculus/Info/Annotation, accessed June 2024). To understand the roles of these genes, an international collaboration, the International Knockout Mouse Consortium (IKMC), was undertaken with the goal of creating an extensive library of conditional knockouts in mouse ES cells. The data are available on the website of the International Mouse Phenotyping Consortium (IMPC; https://www.mousephenotype.org) (Groza et al. 2023; Skarnes et al. 2011). As part of the knockout mouse program, mutant mouse lines underwent the same phenotype observation pipeline, which took a broad screening approach with the goal of addressing a wide spectrum of diseases (Birling et al. 2021; White et al. 2013). These mouse mutants have proven to be a highly valuable resource for investigating multiple forms of disease, and there now are over 7,200 peer-reviewed publications citing the IMPC (https://www.mousephenotype.org/data/publications, accessed June 2024).
The knockout-first allele was designed to be a flexible approach where multiple types of alleles can be obtained from a single starting allele; the design allows for an attempt to recover gene function once it has already been disrupted (Skarnes et al. 2011; Testa et al. 2004). The starting allele consists of a large disruption cassette which is targeted to a specific intron of a protein-coding gene, chosen to be directly upstream of a “critical” exon. Ideally, a critical exon is one which is present in all transcripts of the gene, which would introduce a frameshift when deleted (i.e. its length is not divisible by 3), and would interrupt a functional domain and occur early in the gene, although for some genes not all of these requirements could be met (Skarnes et al. 2011). The large size of the cassette is predicted to prevent normal splicing, thus interfering with the transcription of the gene (Fig. 1).
The knockout-first targeted mutation allele (tm1a) is the original, starting allele. The number refers to the attempt made by the researchers; so where a second attempt is needed to create the allele, it would be called ‘tm2a’. The knockout-first allele consists of several key components, including a lacZ reporter gene, a neomycin resistance cassette, FRT sites, and loxP sites (Fig. 1). Several other alleles can be generated from the tm1a allele. The FRT sites can be used to convert the knockout-first tm1a allele to a conditional allele (tm1c), where gene function is rescued, using the flippase (FLP) enzyme to remove the large transcription-disrupting cassette (Fig. 1). The loxP sites allow recombination when exposed to the Cre recombinase enzyme which deletes the critical exon, resulting in a lacZ-tagged deletion allele (tm1b), which lacks the critical exon entirely (Fig. 1) (Skarnes et al. 2011). The tm1e allele is a targeted non-conditional allele, without conditional potential because it has lost the 3’ loxP site (Fig. 1) (https://www.mousephenotype.org/understand/start-using-the-impc/allele-design/). While the tm1e allele is still damaging, it cannot be converted into a tm1b allele, and use of Flippase would simply remove the disruption cassette, rather than creating the tm1c conditional allele.
In the context of this study, the most important aspect of the cassette is a short sequence tagged onto the start of the lacZ gene: the splice acceptor site of the engrailed 2 (En2) gene. The En2 splice acceptor site is 158 bp in length, with the following sequence: gtcccaggtcccgaaaaccaaagaagaagaaccctaacaaagaggacaagcggcctcgcacagccttcactgctgagcagctccagaggctcaaggctgagtttcagaccaacaggtacctgacagagcagcggcgccagagtctggcacaggagctc (bold marks the cryptic splice donor site discussed below; the canonical En2 acceptor site at the start of the exon sequence is underlined). This sequence was inserted in order to ensure lacZ was spliced to and transcribed effectively (Gossler et al. 1989; Skarnes et al. 2011).

Schematic of the knockout-first allele, and its different versions. Yellow boxes (e) show exons, the red box shows the critical exon (ce), green triangles show FRT sites, and brown triangles show the loxP sites. The main portion of the cassette consists of the lacZ gene (lacZ, blue rectangle), with the En2 splice acceptor sequence (En2 SA, pink) at the 5’ end, and the neomycin resistance gene (neo, teal rectangle). Both genes have T2A sites (orange, T) at their 5’ ends and the neomycin gene has a polyadenylation site (pA) at the 3’ end; the T2A sites ensure each gene product is translated independently. This is the promoterless cassette; the promoter-driven cassette has an additional loxP site and the beta-actin promoter directly before the neomycin resistance gene, which increases its expression. It also has an internal ribosome entry site at the start of the lacZ gene instead of the first T2A site, a polyadenylation sequence at the end of the lacZ gene, and does not include the second T2A site (Skarnes et al. 2011). The tm1a (knockout-first) allele has the cassette inserted into the intron before the critical exon, leaving the critical exon itself intact. The tm1b (lacZ-tagged deletion) allele can be generated using Cre recombinase (brown lines), and the tm1c conditional allele can be generated using Flippase (green lines). The tm1e (targeted, non-conditional) allele is the same as the tm1a allele but lacks the final loxP site, which is due to a crossover event in the targeted ES cell clone (Skarnes et al. 2011); it does not directly derive from the tm1a allele and cannot be converted to a conditional allele using Flippase
Mutant mice from the IKMC have been widely used for investigating gene function, and along with the reports describing this research, several studies have reported the detection of a mutant transcript which includes a 115 bp section of the En2 sequence (gtcccaggtcccgaaaaccaaagaagaagaaccctaacaaagaggacaagcggcctcgcacagccttcactgctgagcagctccagaggctcaaggctgagtttcagaccaacag; the remaining 2 bp of the cryptic splice donor site are shown in bold) (Ebrahim et al. 2016; Ghanawi et al. 2021; Hosur et al. 2020; Lachgar-Ruiz et al. 2023; Martelletti et al. 2020). Both tm1a and tm1b alleles were found to include this En2 insertion sequence. In the case of the Hnrnpr tm1a allele, the En2 insertion was included immediately before the critical exon (Ghanawi et al. 2021) (Fig. 2A), but in the case of the tm1b alleles reported thus far, the En2 insertion replaced the critical exon. In some mutants, this is predicted to result in a frameshift, the introduction of a stop codon, and nonsense-mediated decay (Whrn (Ebrahim et al. 2016); Synj2 (Martelletti et al. 2020) (Fig. 2B), and in others, a stop codon is introduced within the En2 insertion itself (Rhbdf1 (Hosur et al. 2020) (Fig. 2C). However, in some cases, it could potentially allow readthrough and the generation of a mutant protein (Ccdc50 (Lachgar-Ruiz et al. 2023) (Fig. 2D).
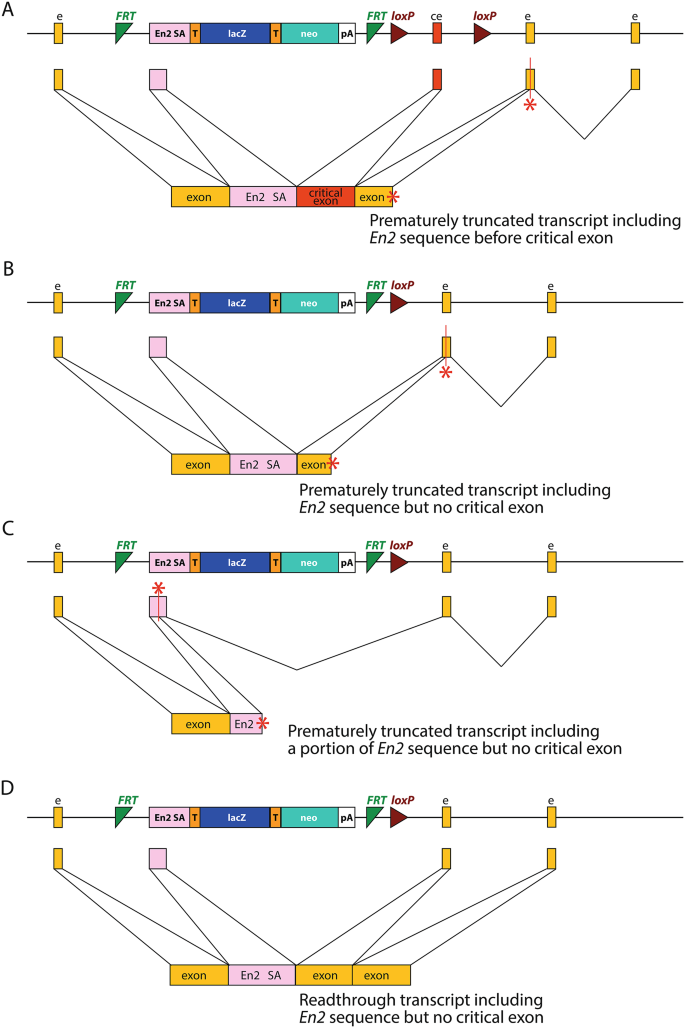
Examples of En2 splice acceptor sequence inclusion. (A) Insertion of the En2 splice acceptor before the critical exon in a tm1a allele, as seen in Hnrnpr (Ghanawi et al. 2021). In this case, the inserted sequence results in a frameshift and a transcript which truncates at a premature stop codon (red asterisk). (B) Insertion of the En2 splice acceptor in place of the critical exon in a tm1b allele, as seen in Synj2 (Martelletti et al. 2020) and Whrn (Ebrahim et al. 2016). In these cases, the inserted sequence is also predicted to result in a frameshift and a truncated protein. (C) In some cases, the insertion of the En2 splice acceptor in place of the critical exon results in a stop codon within the En2 insertion itself, as seen in Rhbdf1 (Hosur et al. 2020). (D) However, in Ccdc50 (Lachgar-Ruiz et al. 2023), the inserted sequence maintains the reading frame and results in a mutant protein with a section from the En2 gene
This unexpected splicing is thought to be due to a cryptic splice site partway through the En2 sequence, which allows splicing from within the sequence to the next exon of the host gene. The effect of this on transcription will depend on how the En2 sequence fits into the endogenous mRNA; the result may be a protein with the En2 splice acceptor site sequence inserted that could be at least partially functional. It is important for researchers, who may be expecting a full knockout of the gene, to know which targeted knockouts may exhibit this behaviour that can result in a different phenotype than the intended knockout allele. We have investigated the potential for these different outcomes in 14,262 knockout-first allele designs and tested for the presence of the insertion in 30 mutant lines in addition to the 5 previously reported. We have also assessed the effect of this unexpected splicing upon the expression of the lacZ reporter gene.
Methods
Phase calculations
The effect of the insertion of the En2 sequence into a transcript depends firstly on whether it is inserted before the critical exon or in place of the critical exon. Because the En2 sequence up to the cryptic splice site is 115 bp long, which is not a multiple of 3, it will introduce a frameshift if inserted before the critical exon in a knockout-first (tm1a) allele. However, if it replaces the critical exon, its effect depends on the length of the critical exon and the phase in which the En2 sequence is read. Each exon has a start and end phase, which may be 0, 1, or 2. The start phase refers to the position of the intron/exon boundary within a codon, which corresponds to the number of bases used from the previous exon to form a complete codon. For example, for an exon starting in phase 1, the first two bases of the exon will join the last base from the previous exon to form a codon (Fig. 3A). This also means the end phase of one exon is the same as the start phase of the next exon.
The transcription outcome for each of the 9 potential phase combinations are shown in Table 1. If the critical exon is read starting in phase 2, a TAA stop codon will be transcribed from within the En2 sequence, starting at base 37. However with the other two potential start phases, if the critical exon ends in the same phase as the inserted En2 sequence, there could be a readthrough, with the En2 sequence being incorporated into the transcript in place of the critical exon. If the critical exon end phase differs from that of the En2 sequence, it would result in a frameshift (Fig. 3B).

Phase calculations for the En2 insertion. A. Two representative exon sequences are shown on the top of the diagram, 10 and 11 base pairs in length. The coloured boxes show potential codons for the three different start phases, with end phases also shown. The phase number is the start phase of the current exon, as well as the end phase of the previous exon. B. Replacing exon 1 with the 115 bp En2 insertion (pink boxes, below) would permit readthrough if the exon starts in phase 0 or 1, since the end phases of the En2 insertion match the end phases of exon 1. However, for exon 2, the end phases do not match, so if the exon starts in phases 0 or 1, the result of the En2 insertion is predicted to be a frameshift. If the En2 insertion replaces an exon which starts in phase 2, a stop codon is introduced within the En2 sequence itself (red asterisk), resulting in a truncated transcript. If the previous exon ends in phase 2 with ‘TA’ (as exon 1 in the example), the first ‘G’ of the En2 insertion would make an extra stop codon at the very start of the insertion, with the same ultimate effect
Computational analysis
The online databases Ensembl (http://www.ensembl.org/index.html) and IMPC (https://www.mousephenotype.org/) were used to collect bulk data for computational analysis. All genomic data is from the Genome Reference Consortium Mouse Build 39 (GRCm39) release. Scripts were written in python to parse the files and carry out further analysis (Fig. 4); specifically, to obtain exon phases for the critical exon(s) for each targeted mutation, to classify each mutation according to the phase calculations (Table 1), and to calculate the number and severity of recorded phenotypes. The script is available on GitHub (https://github.com/prernanair/En2-Cryptic-Splice-Site).

Transcription outcome prediction flowchart. Input files are shown in the coloured shapes, and automated processes are shown in squares. Files that were the output of the script are shown in parallelograms. The numbers (bold red text) show the number of designs that are present at each stage of analysis. The start and end phase combinations of the critical exon(s) are also shown
Ethics statement
Mouse studies were carried out in accordance with UK Home Office regulations and the UK Animals (Scientific Procedures) Act of 1986 under UK Home Office licences, and the study was approved by the Wellcome Sanger Institute or the King’s College London Animal Welfare and Ethical Review Bodies. Mice were culled using methods approved under these licences to minimise any possibility of suffering. Mice were group-housed in individually ventilated cages at a standard temperature and humidity and in specific-pathogen-free conditions, with lighting on a 12 h on/12 hours off cycle, and in accordance with the EU Directive 2010/63/EU for animal experiments. Both males and females were used. Samples were collected within a 1.5-hour window from 6 h after lights on.
Sample collection
Samples from thirty mouse (Mus musculus, NCBI Taxon ID 10090) lines representing 27 genes were used to test our predictions (Table 2). Brain tissue was snap-frozen in liquid nitrogen, and inner ear tissue was preserved in RNAlater (Invitrogen, cat.no AM7024) before being frozen.
RNA extraction, cDNA creation and RT-PCR
Tissue samples were used from previous published and unpublished studies (e.g. (Chen et al. 2024; Ingham et al. 2020; Lachgar-Ruiz et al. 2023). For all samples, RNA was either extracted from brain using TRIzol (Invitrogen, cat no 15596018) or from inner ear tissue using a QIAgen RNEasy kit (cat no 74104) or a Lexogen SPLIT kit (cat no 008.48), all as per the manufacturer’s instructions. cDNA was made using either Superscript II Reverse Transcriptase (Invitrogen, cat no 18064014), Superscript VILO Reverse Transcriptase (Invitrogen, cat no 11766050) or Primerdesign RT nanoscript 2 (cat no RT-premix2-48). The samples used represent a range of ages (Table 2).
Primers were designed to amplify from exons before and after the critical exon using Primer3 (Untergasser et al. 2012). Primer sequences are in Online Resource 1. A touchdown PCR protocol was used to amplify cDNA, as follows: 94 °C for 2 min; 94 °C for 30 s; 64 °C for 45 s (decreasing by 0.5 °C per cycle); 72 °C for 45 s; Carry out steps 2–4 16 times in total; 94 °C for 30 s then 55°C for 45 s then 72 °C for 45 s, repeated 21 times in total; 72 °C for 7 min.
PCR samples underwent an enzymatic cleanup using Illustra ExoProStar (Cytiva Life Sciences, cat no GEUS77705) and were sequenced by Source Bioscience (Nottingham, UK). Sequencing reads were aligned and analysed using Gap4 (Bonfield et al. 1995).
Results
Categorization into transcription predictions
14,262 allele designs had phase information and were analysed and sorted into categories based on the phase combinations described in Table 1, assuming that the En2 sequence replaces the critical exon. 2,481 designs were categorized separately into the ‘Other’ category as they had either negative start or end phases, indicating the exon/intron boundary is in the non-coding region of the pre-mRNA (Table 3). These were not analysed further, with the exception of those exons starting in phase 2 and ending in a negative phase, where the inclusion of the En2 sequence would result in a slightly truncated protein; these were included with the En2-induced stop codon counts.
Phenotypic differences
Data on the phenotypes of mice carrying these alleles were obtained from the IMPC (https://www.mousephenotype.org, (Groza et al. 2023) FTP site (http://ftp.ebi.ac.uk/pub/databases/impc/all-data-releases/latest/results/procedureCompletenessAndPhenotypeHits.csv.gz and http://ftp.ebi.ac.uk/pub/databases/impc/all-data-releases/latest/results/laczExpression.csv.gz, accessed June 2024) as a list of mutant lines and the outcome of tests carried out, including associated Mammalian Phenotype (MP) terms (https://www.informatics.jax.org/vocab/mp_ontology/). Each MP term associated with a mutant mouse line represents a significant difference between control and mutant mice. Significance in the IMPC data is assessed using the OpenStats software package, which was designed for high-throughput phenotypic data (Haselimashhadi et al. 2020). We counted the number of associated MP terms for knockout-first and targeted non-conditional alleles, and for all forms of reporter-tagged deletion allele, and considered the percentage of genes which produced no abnormal phenotype, the percentage of lines with a lethality phenotype, and the average number of abnormal phenotypes per mouse line. None of these showed any notable differences between the different transcription outcome categories (Table 4; Fig. 5).

Proportion of tested lines showing lethal, nonlethal and no phenotypes grouped by allele type and transcription outcome prediction. Genes with multiple designs in different categories have been excluded
We also checked for the absence of any lacZ expression, but found only 36 genes where there was no expression recorded in any of 27 or more tissues tested out of a total of 2909 genes.
Testing for the presence of the En2 insertion in the transcripts of targeted genes
There were thirty-five mutant lines for which data were available, thirty from this study and five previously reported. Twenty-eight were knockout-first (tm1a, tm2a) or targeted non-conditional (tm1e) alleles, and seven were lacZ-tagged deletion (tm1b) alleles. Of the 28 tm1a, tm1e and tm2a alleles, 18 carried the 115 bp En2 sequence before the critical exon and one (Col4a3tm1a) carried the En2 sequence in place of the critical exon (Table 5). Of the seven tm1b alleles, five carried the En2 sequence in place of the critical exon (Table 5). Two of the 35 alleles analysed experimentally showed readthrough or readthrough potential, where the En2 sequence was seen in place of the critical exon.
Discussion
After testing, and including the alleles already reported (Ebrahim et al. 2016; Ghanawi et al. 2021; Hosur et al. 2020; Lachgar-Ruiz et al. 2023; Martelletti et al. 2020), the En2 sequence was found to be present in mRNA from 24 of 35 lines tested overall; 19 of 28 knockout-first (tm1a, tm2a) and targeted non-conditional (tm1e) alleles and five of seven lacZ-tagged deletion (tm1b) alleles (Table 5). There were seven cases where a subset of individual animals exhibited the En2 insertion, suggesting that its inclusion depends on more than just the host gene and chosen site for the cassette insertion. This may be a source of variation between mutant animals carrying the same allele. It should be noted that for these experiments, the tissue used was from either the brain or the inner ear, and different results may be obtained from other tissues due to differences in tissue-specific expression and splicing.
For most of the knockout-first and targeted non-conditional alleles where the En2 insertion was observed, it was inserted before the critical exon, thus inducing a frameshift, so the ultimate purpose of the allele, transcriptional disruption of the host gene, would still be achieved. However, we observed one example (Col4a3) where the En2 insertion replaced the critical exon, in which case the ultimate outcome depends on the start and end phases of the critical exon, as it does for the lacZ-tagged deletion (tm1b) alleles. For those alleles where the En2 sequence replaces the critical exon and where readthrough is predicted to occur, a malformed protein may result. This malformed protein may be a functional null, but may also be a hypomorph, or even exhibit gain-of-function effects, and thus the phenotypes observed may not be the result of a loss of protein function. This is a separate phenomenon to “leaky” transcription, which occurs when some full-length mRNA is transcribed with correct splicing despite the allele being designed to prevent it. Leaky transcription has been reported from several knockout-first alleles (Ingham et al. 2020; Martelletti et al. 2020; White et al. 2013) and is a known potential issue with the targeted mutation allele design.
No quantitative analysis of mRNA level was done in this study, so the overall effects of the En2 insertion on transcription levels have not been determined. However, a number of tm1a, tm2a, tm1e and tm1b alleles (such as Sgms1tm1a, Sgms1tm1b, Evi5tm1a, Amz2tm1e, and Ptprdtm2a) have been reported to result in downregulation in homozygous mice (Chen et al. 2024; Ingham et al. 2020). All these alleles had some evidence of the En2 insertion in mutant mice (Table 5).
Despite the apparent prevalence of the En2 insertion in these mouse mutants, we did not observe any major differences in the occurrence of preweaning lethality, or the number of phenotypes per mouse line in the three transcription outcome categories from the IMPC data (Table 4; Fig. 5). This suggests that even when the En2 insertion permits readthrough of the mutant transcript, the majority of these alleles will still result in a less functional protein. However, that does not rule out the possibility of an ameliorating effect on a minority of readthrough alleles. For example, Frmd5tm1a mice exhibit preweaning lethality with incomplete penetrance, but there is no lethality phenotype recorded for the Frmd5tm1b allele. The En2 insertion in Frmd5 is predicted to result in a readthrough (Online Resource 2), which may explain the unexpected loss of the lethality phenotype in the tm1b allele. However, there are other reasons this can occur, including reinitiation from a downstream start codon (as seen in Rhbdf1tm1b (Hosur et al. 2020), which emphasises the importance of verifying the transcriptional and translational outcomes of each mutant allele used in a study. It should also be noted that approximately 70% of the genes in each transcription outcome category had no available phenotype data, and that mutant phenotypes may be missed if the relevant organ systems or life stages are not tested. The IMPC is a broad phenotyping screen, but it does not cover everything.
We did observe differences between the knockout-first and targeted non-conditional alleles (tm1a, tm1e) and the lacZ-tagged deletion alleles (tm1b). The percentage of lines with lethality is higher for lacZ-tagged deletion alleles, and they also have a higher average number of phenotypes per line, while a lower percentage of lines carrying lacZ-tagged deletion alleles show no phenotype. This suggests that leaky transcription may be rescuing phenotypes in the knockout-first and targeted non-conditional lines. The overall count of phenotypes per line is similar to that observed by Dickinson et al. (Dickinson et al. 2016).
The function of the En2 sequence in the cassette is to enable splicing to and transcription of the lacZ reporter gene in the time and location in which the host gene would normally be expressed. If splicing takes place between the En2 cryptic splice site and the next splice acceptor site in the host gene, transcription of the lacZ gene could potentially be disrupted. Such splicing was detected in 24 of 35 alleles tested in this study and others (Table 5), and is independent of the outcome category (readthrough, frameshift or introduced stop codon). However, the presence of the En2-included mutant transcript does not mean that the lacZ fusion transcript is not also present, and for 19 of the 35 alleles tested, lacZ expression has been reported either in peer-reviewed publications or by the IMPC (Table 5). We further investigated this using the reported lacZ expression data from the IMPC, and found 36 genes out of 2909 where none of the 27 or more tissues tested exhibited any lacZ expression. However, these genes may be specifically expressed at a life stage or in a tissue which wasn’t investigated, which means it isn’t possible to say from this analysis whether loss of lacZ expression is a problem for this small subset of genes.
This study highlights the importance of confirming the nature of a mutation before conducting experiments that rely on it. Suitable methods to do this include sequencing cDNA made using RNA extracted from the relevant tissue, and Western blots using antibodies against different regions of the protein, if they are available, in order to confirm protein size. With all mutant mouse ES cells created by the IKMC having undergone the same design pipeline, they are all susceptible to the same potential for inclusion of the 115 bp En2 sequence. It is recommended that when researchers work with knockout-first alleles and alleles derived from them, they assess the transcription outcome prior to interpreting their experiments. A knockdown instead of a knockout of transcription is a more common outcome to consider, but readthrough leading to an abnormal mRNA and potentially an abnormal protein, as described here, is an additional consideration.
Data availability
All data supporting the findings of this study are available within the paper and its Supplementary Information. The python script used for the analysis described in this paper is available at https://github.com/prernanair/En2-Cryptic-Splice-Site.
References
Birling MC, Yoshiki A, Adams DJ, Ayabe S, Beaudet AL, Bottomley J, Bradley A, Brown SDM, Burger A, Bushell W, Chiani F, Chin HG, Christou S, Codner GF, DeMayo FJ, Dickinson ME, Doe B, Donahue LR, Fray MD, Gambadoro A, Gao X, Gertsenstein M, Gomez-Segura A, Goodwin LO, Heaney JD, Herault Y, de Angelis MH, Jiang ST, Justice MJ, Kasparek P, King RE, Kuhn R, Lee H, Lee YJ, Liu Z, Lloyd KCK, Lorenzo I, Mallon AM, McKerlie C, Meehan TF, Fuentes VM, Newman S, Nutter LMJ, Oh GT, Pavlovic G, Ramirez-Solis R, Rosen B, Ryder EJ, Santos LA, Schick J, Seavitt JR, Sedlacek R, Seisenberger C, Seong JK, Skarnes WC, Sorg T, Steel KP, Tamura M, Tocchini-Valentini GP, Wang CL, Wardle-Jones H, Wattenhofer-Donze M, Wells S, Wiles MV, Willis BJ, Wood JA, Wurst W, Xu Y, Teboul C, Murray L SA (2021) A resource of targeted mutant mouse lines for 5,061 genes. Nat Genet 53:416–419. https://doi.org/10.1038/s41588-021-00825-y
Bonfield JK, Smith K, Staden R (1995) A new DNA sequence assembly program. Nucleic Acids Res 23:4992–4999. https://doi.org/10.1093/nar/23.24.4992
Buniello A, Ingham NJ, Lewis MA, Huma AC, Martinez-Vega R, Varela-Nieto I, Vizcay-Barrena G, Fleck RA, Houston O, Bardhan T, Johnson SL, White JK, Yuan H, Marcotti W, Steel KP (2016) Wbp2 is required for normal glutamatergic synapses in the cochlea and is crucial for hearing. EMBO Mol Med 8:191–207. https://doi.org/10.15252/emmm.201505523
Chen J, Ingham N, Kelly J, Jadeja S, Goulding D, Pass J, Mahajan VB, Tsang SH, Nijnik A, Jackson IJ, White JK, Forge A, Jagger D, Steel KP (2014) Spinster homolog 2 (spns2) deficiency causes early onset progressive hearing loss. PLoS Genet 10:e1004688. https://doi.org/10.1371/journal.pgen.1004688
Chen J, Lewis MA, Wai A, Yin L, Dawson SJ, Ingham NJ, Steel KP (2024) A new mutation of Sgms1 causes gradual hearing loss associated with a reduced endocochlear potential. Hear Res 451:109091. https://doi.org/10.1016/j.heares.2024.109091
Dickinson ME, Flenniken AM, Ji X, Teboul L, Wong MD, White JK, Meehan TF, Weninger WJ, Westerberg H, Adissu H, Baker CN, Bower L, Brown JM, Caddle LB, Chiani F, Clary D, Cleak J, Daly MJ, Denegre JM, Doe B, Dolan ME, Edie SM, Fuchs H, Gailus-Durner V, Galli A, Gambadoro A, Gallegos J, Guo S, Horner NR, Hsu CW, Johnson SJ, Kalaga S, Keith LC, Lanoue L, Lawson TN, Lek M, Mark M, Marschall S, Mason J, McElwee ML, Newbigging S, Nutter LM, Peterson KA, Ramirez-Solis R, Rowland DJ, Ryder E, Samocha KE, Seavitt JR, Selloum M, Szoke-Kovacs Z, Tamura M, Trainor AG, Tudose I, Wakana S, Warren J, Wendling O, West DB, Wong L, Yoshiki A, International Mouse Phenotyping C, Jackson L, Infrastructure Nationale Phenomin ICdlS, Charles, River L, Harwell MRC, Toronto Centre for P, Wellcome Trust Sanger I, Center RB, MacArthur DG, Tocchini-Valentini GP, Gao X, Flicek P, Bradley A, Skarnes WC, Justice MJ, Parkinson HE, Moore M, Wells S, Braun RE, Svenson KL, de Angelis MH, Herault Y, Mohun T, Mallon AM, Henkelman RM, Brown SD, Adams DJ, Lloyd KC, McKerlie C, Beaudet AL, Bucan M, Murray SA (2016) High-throughput discovery of novel developmental phenotypes. Nature 537, 508–514 https://doi.org/10.1038/nature19356
Ebrahim S, Ingham NJ, Lewis MA, Rogers MJC, Cui R, Kachar B, Pass JC, Steel KP (2016) Alternative splice forms influence functions of Whirlin in Mechanosensory Hair Cell Stereocilia. Cell Rep 15:935–943. https://doi.org/10.1016/j.celrep.2016.03.081
Ghanawi H, Hennlein L, Zare A, Bader J, Salehi S, Hornburg D, Ji C, Sivadasan R, Drepper C, Meissner F, Mann M, Jablonka S, Briese M, Sendtner M (2021) Loss of full-length hnRNP R isoform impairs DNA damage response in motoneurons by inhibiting Yb1 recruitment to chromatin. Nucleic Acids Res 49:12284–12305. https://doi.org/10.1093/nar/gkab1120
Gossler A, Joyner AL, Rossant J, Skarnes WC (1989) Mouse embryonic stem cells and reporter constructs to detect developmentally regulated genes. Science 244:463–465. https://doi.org/10.1126/science.2497519
Groza T, Gomez FL, Mashhadi HH, Munoz-Fuentes V, Gunes O, Wilson R, Cacheiro P, Frost A, Keskivali-Bond P, Vardal B, McCoy A, Cheng TK, Santos L, Wells S, Smedley D, Mallon AM, Parkinson H (2023) The International mouse phenotyping Consortium: comprehensive knockout phenotyping underpinning the study of human disease. Nucleic Acids Res 51:D1038–D1045. https://doi.org/10.1093/nar/gkac972
Haselimashhadi H, Mason JC, Mallon AM, Smedley D, Meehan TF, Parkinson H (2020) OpenStats: a robust and scalable software package for reproducible analysis of high-throughput phenotypic data. PLoS ONE 15:e0242933. https://doi.org/10.1371/journal.pone.0242933
Hosur V, Low BE, Li D, Stafford GA, Kohar V, Shultz LD, Wiles MV (2020) Genes adapt to outsmart gene-targeting strategies in mutant mouse strains by skipping exons to reinitiate transcription and translation. Genome Biol 21:168. https://doi.org/10.1186/s13059-020-02086-0
Ingham NJ, Rook V, Di Domenico F, James E, Lewis MA, Girotto G, Buniello A, Steel KP (2020) Functional analysis of candidate genes from genome-wide association studies of hearing. Hear Res 387:107879. https://doi.org/10.1016/j.heares.2019.107879
Kochaj RM, Martelletti E, Ingham NJ, Buniello A, Sousa BC, Wakelam MJO, Lopez-Clavijo AF, Steel KP (2022) The Effect of a Pex3 mutation on hearing and lipid content of the inner ear. Cells 1110.3390/cells11203206
Lachgar-Ruiz M, Morin M, Martelletti E, Ingham NJ, Preite L, Lewis MA, Serrao de Castro LS, Steel KP, Moreno-Pelayo MA (2023) Insights into the pathophysiology of DFNA44 hearing loss associated with CCDC50 frameshift variants. Dis Model Mech 1610.1242/dmm.049757
Martelletti E, Ingham NJ, Houston O, Pass JC, Chen J, Marcotti W, Steel KP (2020) Synaptojanin2 mutation causes progressive high-frequency hearing loss in mice. Front Cell Neurosci 14:561857. https://doi.org/10.3389/fncel.2020.561857
Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V, Mujica AO, Thomas M, Harrow J, Cox T, Jackson D, Severin J, Biggs P, Fu J, Nefedov M, de Jong PJ, Stewart AF, Bradley A (2011) A conditional knockout resource for the genome-wide study of mouse gene function. Nature 474:337–342. https://doi.org/10.1038/nature10163
Testa G, van der Schaft J, Glaser S, Anastassiadis K, Zhang Y, Hermann T, Stremmel W, Stewart AF (2004) A reliable lacZ expression reporter cassette for multipurpose, knockout-first alleles. Genesis 38:151–158. https://doi.org/10.1002/gene.20012
Untergasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, Remm M, Rozen SG (2012) Primer3–new capabilities and interfaces. Nucleic Acids Res 40:e115. https://doi.org/10.1093/nar/gks596
White JK, Gerdin AK, Karp NA, Ryder E, Buljan M, Bussell JN, Salisbury J, Clare S, Ingham NJ, Podrini C, Houghton R, Estabel J, Bottomley JR, Melvin DG, Sunter D, Adams NC, Sanger Institute Mouse, Genetics P, Tannahill D, Logan DW, Macarthur DG, Flint J, Mahajan VB, Tsang SH, Smyth I, Watt FM, Skarnes WC, Dougan G, Adams DJ, Ramirez-Solis R, Bradley A, Steel KP (2013) Genome-wide generation and systematic phenotyping of knockout mice reveals new roles for many genes. Cell 154:452–464. https://doi.org/10.1016/j.cell.2013.06.022
Acknowledgements
We thank Karim Boustani, Annalisa Buniello, Jing Chen, Hoi Ling Abigail Chung, Julia Crunden, Francesca Di Domenico, Neil Ingham, Elysia James, Darcey Kirwin, Rafael Kochaj, Maria Lachgar-Ruiz, Daniel Pentland, Lorenzo Preite, Victoria Rook, Sonja Tang and Nina Treder for access to the tissue samples used, the Wellcome Sanger Institute Mouse Genetics Project for generating and providing the mutant mice, and Professor Miguel Angel Moreno Pelayo for useful discussions.
Funding
This research was funded by Wellcome (221769/Z/20/Z; 098051, WT089622MA), the Biotechnology and Biological Sciences Council (BBSRC; BB/M02069X/1), The Medical Research Council (MRC; MR/N012119/1) and the Royal National Institute for Deaf People (RNID; G88). For the purpose of Open Access, the authors have applied a CC BY public copyright licence to any Author Accepted Manuscript (AAM) version arising from this submission.
Author information
Authors and Affiliations
Contributions
P.N. and M.A.L. carried out the bioinformatic analyses and sequencing. M.A.L. and K.P.S. collected additional samples for sequencing. P.N. and M.A.L. wrote the first draft of the paper and all authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
Mouse studies were carried out in accordance with UK Home Office regulations and the UK Animals (Scientific Procedures) Act of 1986 under UK Home Office licences, and the study was approved by the Wellcome Sanger Institute or the King’s College London Animal Welfare and Ethical Review Bodies. Mice were culled using methods approved under these licences to minimise any possibility of suffering. Mice were group-housed in individually ventilated cages at a standard temperature and humidity and in specific-pathogen-free conditions, with lighting on a 12 h on/12 hours off cycle, and in accordance with the EU Directive 2010/63/EU for animal experiments.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nair, P., Steel, K.P. & Lewis, M.A. Investigating the effects of a cryptic splice site in the En2 splice acceptor sequence used in the IKMC knockout-first alleles. Mamm Genome (2024). https://doi.org/10.1007/s00335-024-10071-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00335-024-10071-2