
Abstract
Sleep pressure builds during wakefulness, but the mechanisms underlying this homeostatic process are poorly understood. One zebrafish model suggests that sleep pressure increases as a function of global neuronal activity, such as during sleep deprivation or acute exposure to drugs that induce widespread brain activation. Given that the arousal-promoting noradrenergic system is important for maintaining heightened neuronal activity during wakefulness, we hypothesised that genetic and pharmacological reduction of noradrenergic tone during drug-induced neuronal activation would dampen subsequent rebound sleep in zebrafish larvae. During stimulant drug treatment, dampening noradrenergic tone with the α2-adrenoceptor agonist clonidine unexpectedly enhanced subsequent rebound sleep, whereas enhancing noradrenergic signalling with a cocktail of α1- and β-adrenoceptor agonists did not enhance rebound sleep. Similarly, CRISPR/Cas9-mediated elimination of the dopamine β-hydroxylase (dbh) gene, which encodes an enzyme required for noradrenalin synthesis, enhanced baseline sleep in larvae but did not prevent additional rebound sleep following acute induction of neuronal activity. Across all drug conditions, c-fos expression immediately after drug exposure correlated strongly with the amount of induced rebound sleep, but was inversely related to the strength of noradrenergic modulatory tone. These results are consistent with a model in which increases in neuronal activity, as reflected by brain-wide levels of c-fos induction, drive a sleep pressure signal that promotes rebound sleep independently of noradrenergic tone.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sleep is a widespread—possibly universal—feature of animal life (Keene and Duboue 2018), but its definitive purposes continue to elude us. There is increasing acknowledgement, however, that the functions of sleep relate primarily to the brain (Hobson 2005), perhaps encompassing the replenishment of cerebral energy stores depleted during waking (Benington and Heller 1995) and memory consolidation (Rasch and Born 2013). The timing, duration and intensity of sleep are regulated per the “two-process” model, in which an animal’s circadian rhythm dictates the time(s) of day when it will tend to sleep, while homeostatic sleep pressure accumulates during waking to drive changes in the depth and duration of sleep (Borbély and Achermann 1999). How and where homeostatic sleep pressure accumulates as a function of brain-related processes remains poorly understood. One possibility is that specific sleep-regulatory neurons signal the animal’s need for sleep. In Drosophila for example, there is evidence that the waking activity of R2 neurons of the ellipsoid body generates sleep drive (Liu et al. 2016; Donlea et al. 2018) that is then thought to be communicated to sleep-effecting dorsal fan-shaped body neurons. Alternatively, sleep drive signals could be more globally distributed; for example, in mammals, the kinase SIK3, hypothesised to be a key actor in sleep homeostasis (Funato et al. 2016), shows a broad expression profile across neuronal tissues.
Whether the activity of privileged neurons acts as a bellwether for general sleep need, or sleep drive signalling is more diffuse, it may be that homeostatic sleep need reflects the overall level of brain activity integrated over prior waking. While sleep pressure has traditionally been associated with wake duration (Borbély and Achermann 1999), not all waking behaviour involves equivalent neuronal activity (Fisher et al. 2016; Milinski et al. 2021) and within-waking arousal states can modulate the accumulation of sleep pressure (Yamagata et al. 2021; Vassalli and Franken 2017). Experiments in zebrafish have demonstrated that acutely and transiently elevating neuronal activity with arousing drugs such as caffeine is followed by increased sleep (Reichert et al. 2019). This drug-induced rebound sleep is dissociable from prior wake time and physical hyperactivity but correlates strongly with the level of preceding global neuronal activity as measured by c-fos expression and whole-brain calcium imaging. Consistent with this, the intensity of regional neuronal activity during waking in mammals is associated with the extent of local offline periods and changes in regional slow-wave activity (a measure of sleep pressure) during the following sleep period (Krueger et al. 2019), while in mice, global slow-wave activity during NREMS has been shown to reflect the integrated cortical neuronal activity levels of the preceding wake period (Thomas et al. 2020). How widespread changes in neuronal activity would ultimately trigger changes in whole animal sleep is unclear, but evidence in both mice (Ma et al. 2019) and zebrafish (Reichert et al. 2019) implicates galaninergic neurons of the anterior hypothalamus and preoptic area (POA) as an effector arm of homeostatic sleep regulation.
One vital system for maintaining brain-wide arousal and implicated in c-fos expression during waking is the noradrenergic system (Cirelli and Tononi 2000). The locus coeruleus (LC) is a small neuronal population (~ 10–20 neurons in zebrafish; Farrar et al. 2018) that is the chief source of noradrenalin in the brain (Chandler et al. 2019) and is highly conserved among vertebrates, including zebrafish (Wang et al. 2022). LC neurons ramify widely, such that noradrenalin can act throughout the brain (Du et al. 2018) and also inhibit sleep-active neurons of the POA (Liang et al. 2021; Nelson et al. 2003). Indeed, the activity of the LC is intimately coupled with the sleep/wake behavioural state of the animal, and noradrenergic signalling is required for the normal maintenance of the waking state in animals including mice and zebrafish larvae (Ouyang et al. 2004; Singh et al. 2015). During waking, the LC is tonically active; this activity falls substantially during non-REM sleep (NREMS) (Steininger et al. 2001) and virtually ceases during REM sleep (Jones 1991). Activity in the LC precedes spontaneous waking (Saper et al. 2010), and activation of the LC during sleep can cause immediate sleep-to-wake transitions (Carter et al. 2010). Additionally, phasic burst firing of the LC in response to a salient stimulus (Carter et al. 2010) helps the animal focus its attention (Jones 1991). As such, the maintenance of brain-wide noradrenergic modulatory strength or tone is thought to be crucial to sustaining wake-related arousal and neuronal activity, and is a candidate driver of sleep need (Cirelli et al. 2005).
Here, we explore the role of the noradrenergic system in modulating stimulant drug-induced sleep pressure in zebrafish larvae. Genetic and pharmacological manipulation of noradrenergic transmission surprisingly reveals that lowered noradrenergic tone enhances both stimulant-drug-induced c-fos induction and subsequent rebound sleep. This presents a new insight into the relationship of noradrenergic activity and sleep pressure generation and is consistent with a model whereby increases in neuronal activity, as reflected by c-fos expression, can generate homeostatic sleep drive independently of the noradrenergic system.
Methods and materials
All animal protocols were performed in accordance with project licence PA8D4D0E5, awarded to Jason Rihel by the UK Home Office under the UK Animals (Scientific Procedures) Act 1986. Experiments used AB/Tupfel long-fin larvae up to 8 days post fertilisation (dpf), before the onset of sexual maturation.
Sleep/wake activity assays
Embryos were reared in an incubator at 28.5 °C on a 14 h:10 h light:dark cycle, with lights on from 9am (zeitgeber time zero = ZT0). At 5 dpf, individual larvae were pipetted into each well of a 96-square well plate (Whatman). Each well contained 650 µl of fish water (0.3 g/l Instant Ocean with 40 µg/l of methylene blue). Wells were topped-up daily with fish water.
Videotracking was conducted per Reichert et al. (2019), using an automated Zebrabox system (ViewPoint Behaviour Technology) and maintaining a 14 h:10 h light:dark schedule. Ambient temperature was held at 26–28.5 °C. Constant infrared illumination allowed for videotracking throughout the day/night cycle. “Quantization mode” in the ZebraLab software was used to record larval movements (detection parameters: sensitivity 20, burst 200, freeze 3 and bin size 60 s). Custom “sleep_analysis2020” and “sleep_analysis_widget” MATLAB (MathWorks) codes were used to analyse the Zebrabox activity data (available on GitHub, https://doi.org/10.5281/zenodo.7644073). Sleep was identified as periods of inactivity lasting ≥ 1 min, as such quiescent bouts have been shown to fulfil the criteria for a behavioural definition of sleep, including an elevated arousal threshold (Prober et al. 2006).
To pharmacologically compromise noradrenergic signalling, the α2-adrenoceptor agonist clonidine was added to the fish water on the afternoon of 5 dpf. A 1 mM working solution of clonidine was prepared in 10% dimethyl sulfoxide (DMSO); 3.25 µl of this was pipetted into each 650 µl well to give a final concentration of 5 µM clonidine (after Singh et al. 2015) and 0.05% DMSO. For control wells, 3.25 µl of 10% DMSO was applied to give a final concentration of 0.05% DMSO.
To pharmacologically activate the noradrenergic system, a mixture of the α1-adrenoceptor agonist phenylephrine and the β-adrenoceptor agonist isoproterenol was added to the fish water from ZT0 + 10 min at 6 dpf. A working solution of 0.5 mM phenylephrine and 0.5 mM isoproterenol was prepared in double distilled water. 13 µl of this was pipetted into each 650 µl well to give a final concentration for each drug of 10 µM (after Yin et al. (2009), who found that either 10 µM phenylephrine or 10 µM isoproterenol alone significantly increased the zebrafish larval heart rate, and Rihel et al. (2010), who found that ~ 10 µM isoproterenol decreased larval sleep behaviour).
On 6 dpf at ZT1, the stimulant drugs caffeine or pentylenetetrazol (PTZ), or the same volume of water, were added to individual wells at 20 s intervals. Caffeine, which antagonises adenosine-receptors (Porkka-Heiskanen and Kalinchuk 2011), was applied at 2 mM final concentration. PTZ, a GABAA-receptor antagonist, was applied at 10 mM final concentration (see Table 1 for working solution concentrations). After 1 h of caffeine/PTZ treatment, at ZT2, drug wash-off began. Each larva was individually pipetted into a 13.5 cm diameter petri dish containing ~ 150 ml fish water, and then into a second 13.5 cm water dish, and then into its respective well in a fresh 96-well plate. In Figs. 1, 4, 5 and 6, the blanked-out region on each sleep trace indicates this drug wash-off period, when the larvae were removed from the video tracking apparatus. The wash-off process took about 20 s for each larva. Videotracking then resumed for two days and nights. Supplementary Fig. S1 summarises the experimental drug protocol.

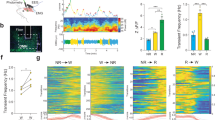
Activating α2-adrenoceptors during drug-induced arousal facilitates rebound sleep. a Sleep traces (± SEM) beginning at 5 dpf and continuing over three days and nights (time since ZT0 at 5 dpf) for larvae exposed to combinations of 5 µM clonidine/DMSO and 2 mM caffeine/water. Following drug wash-off, larvae experience rebound sleep (labelled Rebound Period). At the top, white and black bars represent day and night, respectively; the pale blue horizontal bar shows the clonidine exposure window, while the gold bar indicates the presence of stimulant. b Upper chart shows the average total sleep/h during the rebound period for each larva (black bar: mean and 95% CI). Lower chart shows the effect size (with 95% CI) of clonidine treatment on boosting rebound sleep/h among water-treated and caffeine-treated groups. c Sleep traces as in a for larvae exposed to combinations of clonidine and 10 mM PTZ. The post-drug rebound sleep period of c is summarised for each larva in d (upper chart). Lower chart shows the effect size (with 95% CI) of clonidine treatment on boosting rebound sleep/h among water-treated and PTZ-treated groups
Drug treatment for quantitative real-time polymerase chain reaction (qRT-PCR) assays
Larvae were maintained in a 28.5 °C incubator in petri dishes containing a volume of 45 ml of fish water, with up to 60 larvae in each of four dishes. Where the larvae were to be treated with clonidine or DMSO, these drugs were added to the petri dish at 5 dpf. Where the larvae were to be treated with phenylephrine and isoproterenol, these drugs were added to the petri dish 50 min prior to caffeine application. All drugs were applied to give the same final concentrations as in the sleep/wake assays. Caffeine/PTZ or water vehicle were applied at 6 dpf. After 1 h of caffeine/PTZ treatment, larvae were culled by addition of 8 ml 25× tricaine (see Table 1) to each petri dish, and groups of ~ 15–37 larvae were pipetted into 1.5 ml Eppendorf tubes. Excess fish water was removed with a fine-tipped plastic pastette and sample tubes were frozen in isopentane on dry ice. Samples were then transferred to a − 80 °C freezer.
qRT-PCR for measurement of c-fos mRNA levels
RNA isolation was performed on larval samples by homogenisation in TRIzol and treatment with chloroform. After centrifugation at 12,000g, the aqueous phase (containing RNA) was treated with 2-propanol and re-centrifuged at 12,000g. The RNA pellet was washed with 75% ethanol and resuspended in nuclease-free water. RNA quality was checked using Nanodrop. Only samples with a 260/280 nm ratio of at least 1.8 (indicating minimal protein contamination) and a 260/230 nm ratio of at least 1.9 (minimal phenol contamination) were used for analysis.
AffinityScript Reverse Transcriptase was used for reverse transcription of RNA. For each resulting sample of complementary DNA, levels of fosab (c-fos) were measured for three aliquots and of the housekeeping gene ef1α for another three aliquots, using GoTaq qPCR Master Mix, in a CFX96 Real-Time System BioRad Thermal Cycler. In zebrafish there are two paralogues to mammalian c-Fos: fosaa and fosab. The protein Fosab is the less divergent, with more highly conserved key regulatory phosphorylation sites (Kubra et al. 2022). The primers used for amplification of fosab (c-fos) and ef1α were per Reichert et al. (2019). The “quantification cycle” of c-fos from each sample was measured as the number of PCR cycles taken to reach the threshold level of fluorescence detection. This was then normalised to the quantification cycle of ef1α for the sample, giving the “delta quantification cycle” measure, “dCt”. The c-fos dCt of each sample was then normalised to the dCt measure of control sample(s), to give the “delta dCt” measure, “ddCt”. The relative c-fos expression for each sample versus control was then calculated as 2−ddCt.
F0 KO zebrafish
Filial generation zero (F0) dopamine β-hydroxylase (dbh) knockout (KO) larvae were generated using a CRISPR/Cas9 F0 KO injection method (Kroll et al. 2021). To construct each guide RNA, 1 µl of 200 µM CRISPR RNA (crRNA) was annealed with 1 µl of 200 µM trans-activating CRISPR RNA (tracrRNA), in a mixture with 1.28 µl of duplex buffer, at 95 °C for 5 min (see Table 1). 1 µl of each guide RNA was then separately assembled with 1 µl Cas9 nuclease at 37 °C for 5 min to create a ribonucleoprotein complex. Eggs were injected at the 1-cell stage, shortly after laying, with ~ 1 nl of a mixture of three different ribonucleoprotein complexes. The three guide RNAs targeted different exons in the dbh gene to give a high chance of mutagenesis. The guide RNA target sequences were as follows: sequence 1: 5′-GACGCTGGTTTGCCTATGGG-3′ (within exon 5), sequence 2: 3′-CGGGGGGGAATGGCCATCAC-5′ (within exon 6), and sequence 3: 3′-GGGACGGGGTGTCTGGACGC-5′ (within exon 3). Exons 5 and 6 were targeted because they are asymmetric (i.e., their base pair length is not a multiple of 3), increasing the likelihood of frameshift mutations in cases of exon skipping. Exon 3 was targeted because a mutation within this exon can give rise to non-functional Dbh (Singh et al. 2015).
Control eggs were injected with Cas9 assembled with non-targeting guide RNAs whose sequences were not predicted to match any genomic locus (see Table 1). Injected embryos were reared at 28.5 °C.
Deep sequencing of the dbh gene in F0 KO larvae
Illumina MiSeq was used to estimate the rate of successful mutation of dbh copies in the F0 KOs, using MiSeq Reagent Nano Kit v2 (300 Cycles) (MS-103–1001), as per Kroll et al. (2021). Of the 29 dbh F0 KO larvae used to characterise the dbh F0 KO sleep/wake phenotype (Fig. S7), ten were selected for sequencing. Two control-injected larvae were also selected. Selection was made before inspection of behavioural data. Selected larvae were culled by tricaine overdose and pipetted into individual PCR tubes, from which fish water was then removed using a fine-tipped pastette. The PCR tubes were then frozen at − 20 °C. DNA extraction was performed on the 12 individual larvae using the HotSHOT method: 50 µl of 1× base solution (see Table 1) was added to each larva before incubation for 30 min at 95 °C, then, after cooling, 50 µl of 1× neutralisation solution (see Table 1) was added to each tube. The resulting DNA samples were diluted 2.5 × with ddH2O and stored at − 20 °C for subsequent PCR.
PCR amplification was conducted for each of the three CRISPR-targeted regions for each DNA sample. Each PCR well contained: 1.20 µl DNA template, 8.86 µl nuclease-free water, 3.00 µl Phusion High-Fidelity Reaction Buffer, 0.3 µl 10 mM deoxynucleoside triphosphates (dNTPs), 0.75 µl 10 µM forward primer, 0.75 µl 10 µM reverse primer, and 0.15 µl Phusion High-Fidelity DNA Polymerase (see Table 1 for all sources). The PCR program used was 95 °C for 5 min followed by 40 cycles of: 95 °C for 30 s, 60 °C for 30 s and 72 °C for 30 s, then 72 °C for 5 min and 10 °C until collection. The following three pairs of forward and reverse primers were used, for sequences 1, 2 and 3, respectively. The MiSeq adaptor arm sequence is shown, followed by the dbh-specific sequence (underlined):
5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGACTGTCATGGAACTACAGGGCT-3′
5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGAAGGAGAGGGTTGTGGTAATGA-3′
5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGGGGCATTCGTTTATGGTACAGT-3'
5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGTGGCTTGAGTGAAGTGCAGTAT-3'
5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGGCTCAATATATCCCGTCTCCAG-3′
5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGTTATTTGTAATGTGCGAGTGGC-3′
PCR product length was verified on a selection of three PCR products (and one control containing no PCR product) for each set of primers. Gel electrophoresis was performed using UltraPure Agarose and GelRed, with a 100 bp DNA ladder and xylene cyanol loading dye. PCR product concentration was then measured for a selection of two PCR products for each set of primers using Qubit (dsDNA Broad Range Assay) and diluted as needed with ddH2O to a final DNA concentration of 15–25 ng/µl. ExoSap-IT cleanup was then performed on all samples to degrade remaining primers and nucleotides.
Sequencing data was analysed per Kroll et al. (2021). Reads from one of the scrambled-injected controls were used to normalise mutation counts, so that misalignments present in the control were not considered to be Cas9 mutations in the F0 KOs. The scrambled-injected control from column 12 of the PCR plate was used for normalisation, as the column 11 control appeared to have been contaminated with DNA from column 10.
Of the 46 dbh F0 KOs used to investigate the effect of clonidine on these larvae (Fig. 6), ten were randomly selected for sequencing. Two control-injected larvae were also randomly selected. Sequencing was performed as above (per Kroll et al. 2021), with the exception that BAM files were not filtered prior to the inputting of fastq files to ampliCan, as sense-checking using Integrative Genomics Viewer (IGV) indicated that valid reads were being excluded by the filtering process.
Statistical analysis
Statistical analyses were performed in MATLAB R2020b.
For sleep/wake assays where two variables were manipulated (e.g. stimulant treatment and noradrenergic status), rebound sleep was compared between paired groups using DABEST estimation statistics (Ho et al. 2019). DABEST, or “data analysis with boostrap-coupled estimation”, is more robust than parametric methods for datasets with non-normal distributions. It calculates the effect size of a variable as the difference between group means and uses bootstrapping to construct a 95% confidence interval (95% CI) for this effect size, to illustrate its uncertainty (Ho et al. 2019).
For sleep/wake assays where one variable was manipulated (e.g., noradrenergic status), one-way ANOVA was used if the dataset satisfied the Bartlett test for normality and homogeneity of variance. Otherwise, the nonparametric Kruskal–Wallis test was used.
Differences in qRT-PCR measurements of c-fos expression were statistically analysed across conditions using the Wilcoxon two-sample test, at the level of the dCt metric (Yuan et al. 2006). This nonparametric test was appropriate given the small sample sizes, making no assumption of data normality.
Linear regression analysis was performed to assess the relationship between c-fos expression and rebound sleep across drug conditions, with calculation of the R2 goodness-of-fit measure.
The Kolmogorov–Smirnov two-sample test was used to assess the difference between the frequency distributions of sleep/wake bout lengths of dbh F0 KOs and controls.
Results
Pre-treatment of larvae with clonidine facilitates drug-induced rebound sleep
To assess the effects of suppressing noradrenergic transmission during neuronal hyperactivation on subsequent homeostatic rebound sleep, we induced rebound sleep in larval zebrafish with acute stimulant exposure while also pharmacologically targeting α2-adrenoceptors (Fig. 1). α2-adrenoceptors are G-protein-coupled-receptors that principally bind Gi-proteins to inhibit adenylyl cyclase activity (Perez 2020; Jasper et al. 1998). As such, activation of α2-adrenoceptors tends to inhibit neuronal activity, including autoinhibiting the LC, causing sedation (Nelson et al. 2003). Indeed, clonidine has been shown to enhance sleep in zebrafish (Rihel et al. 2010). Accordingly, following clonidine administration at 5 days post fertilisation (dpf), and prior to exposure to stimulant drugs, sleep levels were increased (Fig. 1a, c). After a ~ 20 h exposure to clonidine, larvae were then treated with either caffeine (Fig. 1a, b) or PTZ (Fig. 1c, d) for 1 h to acutely increase neuronal activity and generate rebound sleep upon wash-off. As expected, treatment with either caffeine (Fig. 1a) or PTZ (Fig. 1c) alone caused sleep levels to be greatly increased during the rebound period from the end of the drug wash-off to lights off at ZT14. This rebound sleep is thought to reflect the greater sleep need caused by enhanced neuronal activity during stimulant exposure (Reichert et al. 2019).
In both experiments, prior clonidine treatment also had a boosting effect on subsequent sleep in the rebound period. In the caffeine protocol (Fig. 1b), although clonidine had no effect on sleep in water-treated control larvae (+ 1.6 min/h; 95% CI lower and upper bound [− 4.1; + 7.3] min/h), clonidine enhanced rebound sleep after caffeine exposure by + 7.2 min/h [+ 0.6; + 13.7]. Similarly, clonidine enhanced sleep following PTZ exposure by a comparable amount to that in the caffeine experiment (+ 5.8 min/h, [− 1.2; + 12.7], Fig. 1d). Considered together, clonidine has an overall boosting effect on rebound sleep across groups. One explanation for this could be that clonidine washed out of the larval brain less quickly than caffeine/PTZ, continuing to agonise α2-adrenoceptors somewhat into the rebound period. However, inspection of clonidine-treated larvae that were not given a stimulant drug (blue traces in Fig. 1a, c) reveals that their daytime sleep levels were only heightened versus controls (gray traces) when clonidine was present in the fish water. Directly after wash-off, sleep of clonidine-only treated animals was similar to control levels, suggesting successful rapid wash-off. To confirm the rebound sleep effects of clonidine in caffeine-treated larvae, the experiment was simplified and repeated with only two experimental conditions: 96 larvae were treated at 5 dpf with either clonidine or DMSO vehicle and then exposed to caffeine for 1 h on the following morning at 6 dpf (Fig. S2). Larvae treated with clonidine showed significantly higher levels of rebound sleep following caffeine wash-off than DMSO-treated larvae (p = 8.8 × 10–8, F(1, 94) = 33.67, one-way ANOVA). These results not only demonstrate that noradrenergic arousal is not required for neuronal activity-dependent rebound sleep but also suggest that reduced noradrenergic tone may in fact enhance rebound sleep.
c-fos induction by neuronal activity-promoting drugs is greater following pre-treatment with clonidine
In zebrafish, both PTZ- and caffeine-induced rebound sleep are positively correlated with the neuronal activity driven during stimulant exposure (Reichert et al. 2019). Clonidine is a sedative and was predicted to dampen neuronal activity during stimulant exposure, yet it enhanced rebound sleep. Therefore, we next investigated the effects of clonidine on stimulant-induced neuronal activity by assessing expression of the immediate early gene c-fos. Brain-wide c-fos expression is enhanced upon waking and after stimulation (Cirelli and Tononi 2000) and is a widely-used indicator of neuronal activity, including in zebrafish (Baraban et al. 2005; Reichert et al. 2019). In control experiments, caffeine-treated larvae showed on average 71-fold higher c-fos expression than water-treated larvae (Fig. 2a, S3a). This is consistent with the observations of Reichert et al. (2019), who found that drugs such as caffeine that elicit rebound sleep induce widespread neuronal c-fos mRNA expression. However, contrary to expectations, when larvae were co-treated with caffeine and clonidine, c-fos expression was elevated even further, being 47% higher than in larvae treated only with caffeine (Fig. 2b, S3b). Our experimental technique does not reveal whether the clonidine-induced further elevation of c-fos occurred uniformly or in particular neuronal subsets, but there was a strong correlation (R2 = 0.985) between the relative c-fos expression induced by combinations of clonidine and caffeine and the associated rebound sleep (Fig. 2c), consistent with previous findings in zebrafish that rebound sleep duration correlates with c-fos levels induced during drug exposure (Reichert et al. 2019).

c-fos expression is higher in larvae following combined treatment with clonidine and caffeine than following caffeine alone. a qRT-PCR on groups of ~ 20 larvae (n = 4 and n = 5 biological replicates per condition) reveals that larvae treated with caffeine had a significant, 71-fold increase in c-fos expression compared to water-treated larvae (*p < 0.05, two tailed Wilcoxon rank sum test performed on the “dCt” metric, see Fig. S3a). b c-fos expression of larvae soaked in clonidine before and during caffeine exposure was significantly higher by 47% than in larvae exposed to caffeine alone (n = 6 biological replicates per condition, **p < 0.01, see Fig. S3b). c The relative c-fos expression induced by different combinations of vehicle, clonidine and caffeine is positively, linearly correlated (R2 = 0.985) with the total rebound sleep induced by these drugs. qRT-PCR was performed on groups of 37 larvae (see Fig. S3c). Note that c plots together the results of two separate experiments; in both experiments there were four groups of larvae each treated with one of the four combinations of clonidine, caffeine, DMSO and water, but in one experiment c-fos expression was measured after drug treatment, and in the other rebound sleep was measured (sleep data is per Fig. 1a and b). Each square in a–c is the mean of three technical replicates
To test whether clonidine’s enhancement of caffeine-induced c-fos expression was drug-specific, we also measured c-fos expression in larvae following treatment with clonidine and PTZ. As observed for caffeine, treatment with clonidine and PTZ further enhanced c-fos expression compared to PTZ treatment alone (Fig. 3a, S3). As in the clonidine/caffeine experiments, there was a strong correlation (R2 = 0.993) between the relative c-fos expression levels in the different clonidine/PTZ treatment conditions and their associated amount of rebound sleep (Fig. 3b). Thus, depressing the noradrenergic system by activating α2-adrenoceptors actually enhances the expression of c-fos, and the level of c-fos induction predicts the duration of subsequent rebound sleep.

Rebound sleep levels correlate with c-fos expression across different clonidine/PTZ treatment combinations. a qRT-PCR on groups of ~ 17 larvae (n = 3 biological replicates per condition) reveals that larvae treated with both clonidine and PTZ had a trend towards higher c-fos expression than those treated with PTZ alone (see also Fig. S4a). b The mean c-fos expression induced by each drug combination is strongly positively correlated (R2 = 0.993) with the amount of rebound sleep induced by each drug condition (see Fig. 1c and d). Each square in a is the mean of three technical replicates
Stimulation of α1- and β-adrenoceptors with isoproterenol and phenylephrine does not boost neuronal activity-induced rebound sleep
Since the inhibition of noradrenergic signalling with clonidine enhanced stimulant-induced c-fos expression and rebound sleep, we next tested the effects of activating noradrenergic transmission by agonising both α1- and β-adrenoceptors while inducing rebound sleep with caffeine exposure. Phenylephrine is an agonist of the principally Gq-coupled α1-adrenoceptors (Perez 2020) and thus tends to enhance neuronal excitability. Isoproterenol is an agonist of β-adrenoceptors, which couple to Gs-proteins to enhance neuronal activity via the stimulation of adenylyl cyclase (Perez 2020), and has been shown to reduce sleep in zebrafish (Rihel et al. 2010). Larvae (6 dpf) were pre-treated at ZT0 with a cocktail of phenylephrine and isoproterenol to activate both α1- and β-adrenoceptors, followed by a 1 h caffeine exposure at ZT1 and then wash-off of all drugs (Fig. 4a). Although caffeine induced robust rebound sleep (Fig. 4a), the addition of isoproterenol and phenylephrine did not enhance sleep (Fig. 4a and b). In fact, isoproterenol and phenylephrine reduced rebound sleep in control larvae (− 4.1 min/h [− 8.1; − 0.3]) but had no measurable effect on rebound sleep in caffeine-treated larvae (− 0.6 min/h [− 6.8; + 5.3]), Fig. 4b.

Activating noradrenergic transmission with isoproterenol and phenylephrine marginally depresses caffeine-induced c-fos expression. a Sleep traces for larvae exposed to combinations of 10 µM isoproterenol + 10 µM phenylephrine (“Iso + Phenyl”) and/or caffeine. At top left, the pale blue horizontal bar shows the isoproterenol + phenylephrine exposure window while the gold bar indicates the presence of caffeine. The post-drug rebound sleep period of a is summarised for each larva in b (upper chart). Lower chart shows the effect size (with 95% CI) of Iso + Phenyl treatment among water-treated and caffeine-treated groups. c qRT-PCR on groups of ~ 18 larvae reveals that each group of larvae pre-treated with isoproterenol + phenylephrine and then caffeine (n = 3 biological replicates) had lower relative c-fos expression than the groups of larvae treated with water and then caffeine (n = 3 biological replicates); see also Fig. S5a. Each square is the mean of three technical replicates. d The average relative c-fos expression induced by each condition is strongly positively correlated (R2 = 0.889) with the total rebound sleep that was induced by the same drug condition (from a and b)
We then tested the effects of isoproterenol and phenylephrine treatment during caffeine exposure on the induction of c-fos expression. In contrast to the enhancement of c-fos expression observed when noradrenergic tone was dampened with clonidine, caffeine-induced c-fos expression was lower in groups pre-treated with isoproterenol and phenylephrine than in water-treated controls (Fig. 4c). We repeated this c-fos measurement with six additional groups of larvae treated with isoproterenol and phenylephrine and six groups treated with water and confirmed that caffeine-induced c-fos expression was on average collectively lower among groups pre-treated with isoproterenol and phenylephrine, but the effect only trended toward significance (p = 0.077, Fig. S5b). However, as in the clonidine experiments, there was a strong positive correlation (R2 = 0.889) between the relative c-fos expression induced by the different drug treatments and the duration of rebound sleep (Fig. 4d), again suggesting a relationship between the magnitude of c-fos induction during stimulant treatment and the sleep pressure generated.
dbh F0 KO larvae phenocopy the high sleep levels of dbh −/− mutants
To complement our pharmacological manipulations of the noradrenergic system and ensure that the effects we had observed were not drug-specific (e.g., off-target effects), we used a genetic knock-out approach to disrupt the dopamine β-hydroxylase (dbh) gene, which is necessary for noradrenalin synthesis. To eliminate dbh function, we injected zebrafish eggs with Cas9 nuclease assembled with guide RNAs that targeted three loci within the dbh gene (see “Methods and materials”; Kroll et al. 2021). The resulting dbh F0 KO larvae were used for experiments at 5–8 dpf.
To verify that dbh function was successfully disrupted in most if not all cells of the F0 KO larvae, we performed deep-sequencing on larval samples and ascertained the frameshift and mutation rates for each of the three targeted loci within the dbh gene. For 10 sequenced F0 KOs (taken at the end of the experiment, see Fig. S7), the proportion of reads that harboured either mutations or frameshift mutations exceeded 50% at each locus in most larvae. One larva, F0 KO 9, was an exception with no mutated reads at any locus, likely due to experimenter error (e.g., an uninjected egg that was trapped in the transfer pipette) (Fig. S6a). Considering all three targeted loci together, 9/10 of the F0 KO larvae had at least 50% frameshifted copies of dbh, and 7/10 had above 80% (Fig. S6b). This high rate of success, which does not take into account the likelihood that non-frameshifting mutations are also deleterious, indicates that most F0 KO larvae were largely, if not completely, functionally null for dbh in most or all cells.
Previous studies have shown that dbh knockout zebrafish (dbh−/−) have elevated baseline sleep, especially during the day (Singh et al. 2015). This reflects the inability of dbh−/− mutants to synthesise the arousal-promoting neurotransmitters noradrenalin and adrenalin (which is synthesised from noradrenalin). We hypothesised that if our dbh F0 KOs were loss-of-function, they would similarly show enhanced sleep, particularly during the day when the arousal systems of diurnal species are most active. Tracking dbh F0 KOs from 5 dpf over several day/night cycles revealed that they had significantly elevated sleep levels, especially during the day, with dbh F0 KOs sleeping on average 50% of the time at 6 dpf (Fig. S7a-b), versus 15% for controls. dbh F0 KOs were unable to sustain wakefulness for long periods, showing significantly shorter wake bouts and a trend towards longer sleep bouts than controls (Fig. S7c-d).
To ascertain more carefully how closely dbh F0 KOs recapitulated the sleep phenotype of published dbh−/− null mutants, we compared the sleep parameters of dbh F0 KOs to those of stable dbh−/− knockout animals as reported in Singh et al. (2015) (underlying data courtesy of David Prober). On average, 6 dpf dbh F0 KOs showed + 233% higher total daytime sleep compared to control larvae, similar to the + 225% elevation of daytime sleep in dbh−/− null mutants (Fig. S8a). Similar results were found in night-time sleep, with dbh F0 KO larvae having an average + 49% increase in total night-time sleep (compared to + 58% in dbh−/− null mutants) (Fig. S8b). As in dbh−/− null mutants, the day and night increases in sleep were due to both an increase in the number and length of sleep bouts. In the day, dbh F0 KO larvae had an increase in sleep bout number (+ 107%, compared to + 201% in dbh−/− mutants) and sleep bout length (+ 63%, compared to + 17% in dbh−/− mutants) (Fig. S8c, S8e). This discrepancy in daytime effect sizes could reflect the different lighting and temperature conditions in which the larvae were raised (in two different labs on separate continents) as well as the potentially incomplete knockout of dbh in F0 KOs. At night, dbh F0 KO larvae and dbh−/− mutants showed broadly similar elevations of sleep bout number (+ 17% and + 27% respectively) and sleep bout length (+ 26% and + 30%) (Fig. S8d, S8f), demonstrating a high degree of similarity in sleep phenotypes between dbh F0 KOs and dbh−/− mutants at night.
Taken together, the sequencing data combined with the similarity between dbh F0 KO and stable dbh−/− knockout animals’ sleep phenotypes suggests that dbh F0 KOs lack Dbh function and are therefore, like dbh−/− mutants (Singh et al. 2015), depleted of noradrenalin.
dbh F0 KOs show enhanced caffeine-induced c-fos expression and robust rebound sleep
Having verified that our CRISPR/Cas9 technique was generating effective dbh knockouts, we used dbh F0 KOs in an assay of caffeine-induced rebound sleep to test the effect of genetic noradrenergic impairment. An important distinction in this experiment versus our pharmacological noradrenergic manipulations is that the genetic noradrenergic impairment is persistent, whereas pharmacological activation of adrenoceptors should cease after drug wash-off. As such, here we observed the ongoing effects of noradrenergic impairment on rebound sleep, rather than the after-effects. Based on the effects of pharmacological manipulation of adrenoceptors, we predicted that rebound sleep would occur robustly in dbh F0 KOs. Indeed, after caffeine wash-off, dbh F0 KOs showed an average increase of + 16.4 min/h (+ 58%) of rebound sleep versus water-treated dbh F0 KOs, indicating that drug-induced rebound sleep can still occur without noradrenalin (Fig. 5a and b). Furthermore, the effect size of caffeine on rebound sleep was quite similar for wild-type (+ 20.9 min/h [+ 16.2; + 25.7]) and dbh F0 KO (+ 16.4 min/h [+ 8.8; + 24.2]) larvae (Fig. 5b), again suggesting that stimulant-induced rebound sleep can occur independently of dbh.

Caffeine triggers c-fos expression more strongly in dbh F0 KOs. a Sleep traces for dbh F0 KO and control-injected larvae exposed to either caffeine or water. The post-drug rebound sleep period of a is summarised for each larva in b (upper chart). Caffeine treatment had a strong boosting effect on rebound sleep in both control-injected and dbh F0 KO larvae (lower chart). The caffeine effect size is not significantly different between the two genotypes. c qRT-PCR on groups of ~ 15 larvae revealed that each group of dbh F0 KO larvae treated with caffeine (n = 3 biological replicates) showed greater relative c-fos expression than the groups of control larvae treated with caffeine (n = 3 biological replicates); see also Fig. S9a. Each square is the mean of triplicate technical replicates. d There is a weak positive correlation between c-fos expression and subsequent rebound sleep levels (R2 = 0.62), but water-treated dbh F0 KO larvae do not conform to this correlation, showing high sleep levels despite low c-fos expression
To assess how the loss of dbh impacted neuronal activity during the stimulant treatment, c-fos expression immediately following caffeine treatment was measured in both dbh F0 KOs and controls. As for larvae with pharmacologically compromised noradrenergic systems (via activation of α2-adrenoceptors with clonidine), c-fos expression was elevated in dbh F0 KOs treated with caffeine versus caffeine-treated wild-type controls (Fig. 5c). However, unlike in the clonidine experiments (Figs. 2c, 3b), there was only a weak correlation between c-fos expression and sleep across all dbh conditions (Fig. 5d). This difference from the pharmacological experiments is likely due to the high sleep levels during the rebound phase of dbh F0 KOs that were exposed only to water, despite the low induction of c-fos expression in these animals. This indicates that, unsurprisingly, high c-fos expression during the stimulant window is not a prerequisite for the high levels of baseline sleep seen in dbh F0 KOs. Nonetheless, exposure to caffeine does induce c-fos expression and subsequent rebound sleep in animals that lack noradrenalin.
Clonidine’s sedative effects are not mediated solely by α2-autoreceptor suppression of noradrenalin release
One model for how the α2-adrenoceptor agonist dexmedetomidine initiates sedation is by primarily activating auto-inhibitory α2-adrenoceptors found presynaptically on LC neurons, thereby suppressing release of noradrenalin (Nelson et al. 2003). However, other work indicates that α2-adrenoceptors can act as heteroreceptors, sitting presynaptically on non-noradrenergic neurons to inhibit release of glutamate (Harris et al. 2018; Shields et al. 2009). Additionally, α2-adrenoceptors can sit post-synaptically and even be excitatory (Harris et al. 2018; Jasper et al. 1998). Indeed, Hu et al. (2012) found that Dbh−/− mice are hypersensitive to dexmedetomidine, indicating that the sedative effects of this α2-adrenoceptor agonist do not rely solely on the inhibition of noradrenergic release. We reasoned that if clonidine causes sedation primarily via suppression of noradrenalin release, then the sedative effects of clonidine should be blunted in dbh F0 KO larvae. Alternatively, if clonidine enhances sleep independently of its inhibition of noradrenergic release, the sleep-inducing effect of clonidine should occur additively, on top of the elevated baseline sleep phenotype seen in dbh F0 KOs.
Applying clonidine to 5 dpf larvae caused daytime sleep levels to rise substantially in both dbh F0 KO and control-injected larvae (Fig. 6a and b). Although clonidine had a stronger boosting effect on baseline sleep in control-injected larvae (+ 26.1 min/h [+ 17.1; + 34.6]), clonidine also enhanced sleep in dbh F0 KO animals (+ 9.8 min/h [+ 4.9; + 15.8]). Thus, clonidine’s sedative effects are not solely due to the suppression of noradrenalin release, as additional sedation was induced in dbh knockout animals that lack noradrenalin. However, the effect of clonidine was significantly blunted in dbh F0 KO larvae compared to controls (Fig. 6b), which is consistent with clonidine’s sedative effects being at least partially mediated by suppression of the noradrenergic system. That said, baseline daytime sleep levels are already very elevated in dbh F0 KOs, capping the sedative effect that could be achieved by the addition of clonidine, and so limiting interpretation.

Clonidine enhances sleep, and rebound sleep, in dbh F0 KO larvae. a Sleep traces for dbh F0 KO and control-injected larvae exposed to clonidine/DMSO and caffeine. b At 5 dpf from clonidine treatment until lights-out, clonidine had a positive effect on sleep levels in both control-injected and dbh F0 KO larvae, as illustrated by positive effect sizes and 95% CIs. The effect size of clonidine was greater among control-injected larvae. The post-caffeine rebound sleep period is summarised for each larva in c. Among dbh F0 KOs, clonidine had a positive effect on rebound sleep. The clonidine effect size was not significantly different between the two genotypes. Deep sequencing was used to verify the successful loss-of-function targeting of dbh in 10 randomly selected dbh F0 KO larvae: all animals had > 93% (mean, 96%) of their amplified dbh copies frameshifted (see Fig. S10)
Following the pre-treatment of larvae with clonidine, we also induced homeostatic rebound sleep with acute exposure to caffeine, to test the effects of clonidine on subsequent rebound sleep in dbh F0 KOs. In dbh F0 KO larvae, clonidine had a positive effect on rebound sleep (+ 4.4 min/h [+ 0.4; + 8.8]), per Fig. 6c. Indeed, clonidine’s rebound sleep-enhancing effects in dbh F0 KOs appeared similar to its effects in wild-type larvae (+ 3.2 min/h [− 3.7; + 10.1]), indicating that clonidine's enhancement of rebound sleep may not arise from the after-effects of its α2-autoreceptor-mediated suppression of noradrenergic release.
Discussion
Noradrenergic tone is highest during waking and promotes neuronal activity and behavioural arousal in vertebrate species including rodents and zebrafish (Carter et al. 2010; Wang et al. 2022). We therefore tested the effects of altering noradrenergic signalling in zebrafish on stimulant-drug-induced rebound sleep, which is hypothesised to be dependent on heightened neuronal activity (Reichert et al. 2019). Unexpectedly, pharmacological inhibition of noradrenergic signalling enhanced stimulant-induced homeostatic rebound sleep, while stimulant-induced c-fos expression was strongest in noradrenergic-compromised larvae. This enhancement of immediate early gene expression may thus underlie the increase in rebound sleep, for example by strengthening a sleep pressure signal, either on a brain-wide basis or in a key sleep-regulatory cell population. Alternatively, diminished noradrenergic arousal may de-potentiate widespread neuronal transmission, causing lingering quiescence into the rebound phase.
Noradrenergic tone inversely modulates stimulant-induced c-fos expression
We found that stimulation of noradrenergic α1- and β-adrenoceptors with a cocktail of phenylephrine and isoproterenol slightly reduced c-fos induction by caffeine in zebrafish larvae and did not enhance subsequent rebound sleep. On the other hand, treatment of larvae with the α2-adrenoceptor agonist clonidine, a sedative, boosted c-fos induction by caffeine and enhanced rebound sleep. Likewise, dbh F0 KOs, which lack noradrenalin, showed elevated stimulant-induced c-fos expression and robust rebound sleep. These effects on the induction of c-fos are consistent with studies that identify c-fos expression as a measure of increases, as opposed to absolute levels, of neuronal activity. Indeed, c-Fos can show a refractory period after seizure induction, during which further seizures do not bring on c-Fos expression (Barros et al. 2015), and immediate early genes are not continually expressed in neurons that are chronically active (Hudson 2018). Rather, c-fos expression occurs in response to a change in stimulation, after which there may be self-inhibition of the c-fos promoter (Hudson 2018). Such self-inhibitory regulation of c-fos expression could explain why c-fos induction is stronger when a stimulus is applied to an animal after a period of sensory deprivation (Cirelli and Tononi 2000). During waking, because noradrenalin enhances the excitability of thalamic relay projections in mammals (Jones 1991; Szymusiak and McGinty 2008), a noradrenergic-compromised animal may be less aware of stimuli, akin to being sensorily deprived. As such, absolute levels of neuronal activity may not be higher in noradrenergic-compromised larvae that in control larvae following stimulant treatment, but the induction of c-fos may be stronger due to a greater magnitude of increase in neuronal activity. This prediction could be tested using larval zebrafish whole-brain neuronal imaging with genetically encoded calcium indicators to measure the ongoing neuronal activity during and after stimulant treatment. Another factor that may contribute to enhanced c-fos expression in noradrenergic-compromised larvae is that we performed our stimulant drug assay during the day, when dbh F0 KO larvae are much more likely to be asleep than wild-type controls. Thus, daytime drug administration will have caused a higher proportion of noradrenergic-compromised animals to undergo sleep-to-wake transitions, potentially bringing on a larger c-fos induction. Regardless of the precise mechanistic underpinnings, in our experiments, both genetic and pharmacological inhibition of noradrenergic signalling led to enhanced stimulant-induced c-fos expression.
Magnification of immediate early gene induction may enhance a sleep pressure signal
Consistent with the findings of Reichert et al. (2019) that levels of pharmacologically-induced rebound sleep correlate with brain-wide c-fos levels, we found a strong correlation between c-fos expression and sleep across noradrenergic/stimulant drug treatment combinations. One explanation for this could be that c-Fos protein, a transcription factor, drives expression of a homeostatic sleep pressure signal (Cirelli et al. 1995). Greater c-fos expression in noradrenergic-compromised, stimulant-treated larvae would then drive a stronger sleep pressure signal, enhancing rebound sleep. To test whether elevated c-fos expression plays a role in driving heightened rebound sleep, behaviour could be assayed in transgenic zebrafish larvae with inducible extra copies of the c-fos gene, which under this model would heighten rebound sleep following stimulant treatment. Conversely, animals with knock-down of c-fos would be expected to show blunted rebound sleep. If c-fos manipulations do indeed alter rebound sleep, additional experiments that restrict the overexpression or knockdown to particular subsets of neurons could be used to dissect whether distinct sleep-regulatory neuronal populations have particular roles in mediating sleep homeostasis. In addition, expression levels of many other immediate early genes including Bdnf and Egr1 have been shown to correlate with homeostatic sleep pressure in mice (Vassalli and Franken 2017) and are acutely and strongly induced by arousing drugs in zebrafish (Sabine Reichert, unpublished observation). Furthermore, the protein product of another immediate early gene, Npas4, was recently shown to help repair neuronal activity-induced DNA double strand breaks (Pollina et al. 2023), and in zebrafish, the build-up of neuronal DNA damage during waking has been shown to increase sleep pressure (Zada et al. 2021). Although it is unknown whether induction of these and other immediate early genes changes in response to manipulation of the noradrenergic system, their possible roles in regulating drug-induced rebound sleep in zebrafish larvae should be explored.
Alternatively, the correlation of the level of c-fos induction with subsequent rebound sleep may reflect altered activity of CREB, which mediates c-fos transcription in response to various stimuli (Ahn et al. 1998). Recent work in mice has demonstrated that CREB, in conjunction with the histone deacetylase HDAC4, acts downstream of the kinase SIK3 to regulate sleep (Kim et al. 2022; Zhou et al. 2022). Heightened c-fos induction during waking may cause changes in CREB’s interaction with HDAC4 and altered transcription of their targets as a function of sleep need. Such a model could be tested by modulating SIK3, HDAC4, or other components of this pathway in zebrafish and observing how drug-induced rebound sleep is affected.
Heightened noradrenergic tone is not required for stimulant-induced c-fos expression or sleep rebound
How drug-induced neuronal activation leads to heightened rebound sleep is unclear; however, the neuropeptide galanin plays a critical role in the response to sleep pressure signals in zebrafish, functioning as an output arm of a sleep homeostat (Reichert et al. 2019). In mammals, a “flip-flop” model of sleep regulation posits that mutual inhibition between wake-promoting neurons such as those of the LC and sleep-promoting GABAergic/galaninergic neurons of the POA enables rapid and absolute transitions between sleep and wake (Saper et al. 2010). dbh F0 KOs lack noradrenalin, so noradrenergic tone is already supressed regardless of the drug treatment they receive. We found that control larvae showed a slightly greater increase in rebound sleep after caffeine treatment (+ 20.9 min/h) than dbh F0 KOs (+ 16.4 min/h), especially just after wash-off (Fig. 5a and b). This suggests that suppression of noradrenergic release is one mechanism involved in driving rebound sleep, consistent with a flip-flop model. In this interpretation, noradrenergic output cannot be further supressed in the dbh F0 KOs, explaining their reduced increase in sleep early in the rebound period compared to the control larvae. However, across the entire rebound period, both dbh F0 KOs and controls had similarly strong sleep rebound responses to caffeine, suggesting that release of noradrenalin from the LC during stimulant drug exposure is not necessary for rebound sleep to subsequently ensue. Indeed, the fact that administering caffeine to dbh F0 KOs enhances their rebound sleep at all, which was similarly observed in clonidine-treated larvae, indicates that noradrenergic tone during waking is not required for the generation of robust neuronal activity-induced rebound sleep.
Figure 7 illustrates a simple model that assimilates our findings with those of Reichert et al. (2019): stimulant drugs drive increases in neuronal activity, as demonstrated by heightened c-fos expression, which drive a sleep pressure signal that is ultimately put into effect by release of galanin from the POA. This process can occur independently of noradrenalin-driven arousal. Given that noradrenergic signalling is a vital downstream effector for the arousing effects of hypocretin (Carter et al. 2012; Singh et al. 2015), the hypocretin system may also be dispensable for neuronal activity-induced rebound sleep, at least insofar as hypocretin-induced arousal relies on noradrenalin. This could be tested by performing stimulant-induced rebound sleep assays on hypocretin receptor knockout larvae.

Noradrenergic activity is not required for stimulant-induced c-fos expression and rebound sleep. During waking, the LC releases noradrenalin to brain-wide targets, promoting arousal (Carter et al. 2010) and inhibiting sleep-promoting GABAergic/galaninergic neurons of the POA (Liang et al. 2021). Despite the role of the LC in maintaining arousal and heightened neuronal activity during waking, our results suggest that stimulant-induced neuronal activity and rebound sleep can occur in the absence of prior noradrenergic tone. Building on the work of Reichert et al. (2019), we propose a model in which stimulant-induced increases in neuronal activity subsequently promote activation of GABAergic/galaninergic sleep-promoting neurons of the POA, which drive sleep, independently of noradrenergic activity. Arrowheads denote activating projections; the bar head denotes an inhibitory projection
A period of reduced noradrenergic activity could directly facilitate subsequent sleep
While an effect of magnified increases in neuronal activity on sleep pressure signalling is one plausible explanation of our results, another possibility is that the animal’s arousal state during waking directly affects subsequent sleep. When noradrenalin activates α1-adrenoceptors at excitatory glutamatergic synapses, this enhances synaptic transmission and can cause long-term potentiation (Perez 2020). Thus, reduced noradrenergic activity could relatively de-potentiate glutamatergic transmission in the wide-ranging brain regions to which the LC projects, limiting subsequent arousal. Cheng et al. (2020) suggest that in rats, sleep-promoting POA neurons receive excitatory glutamatergic afferents that promote sleep. Possible sources of these afferents include glutamatergic sleep-active neurons of the ventrolateral medulla, which reportedly directly excite POA GABAergic neurons in mice (Teng et al. 2022), and NREMS-promoting neurotensin-expressing glutamatergic neurons of the ventrolateral periaqueductal gray, which have been shown to project to the mouse POA (Zhong et al. 2019). Reduced noradrenergic activation of inhibitory α2-adrenoceptors on sleep-promoting POA neurons (Liang et al. 2021) might facilitate potentiation of these glutamatergic afferents (DeBock et al. 2003), thereby promoting sleep.
One seemingly paradoxical implication of direct inhibition of the sleep-promoting POA by noradrenalin is that α2-adrenoceptor agonists such as clonidine will also directly inhibit these sleep-promoting neurons. Indeed, McCarren et al. (2014) found that microinjection of the α2-adrenoceptor agonist dexmedetomidine into isoflurane-anaesthetised mouse ventrolateral POA increased behavioural arousal in vivo and reduced depolarisation in vitro. However, there is also evidence that noradrenergic inhibition of sleep-promoting neurons occurs indirectly, via activation of local GABAergic interneurons (Chamberlin et al. 2003; De Luca et al. 2022). α2-adrenoceptor agonists may therefore cause sedation when applied systemically because reduced noradrenergic activation of GABAergic interneurons that project to POA sleep-promoting neurons might outweigh the effects of the direct inhibition of the POA. This net disinhibition of sleep-promoting neurons would add to the general brain-wide sedating effects of α2-autoreceptor-mediated prevention of release of noradrenalin, along with the possible inhibitory heteroreceptor and postsynaptic effects of α2-adrenoceptor agonists.
The idea that noradrenergic activity during waking might affect subsequent sleep makes intuitive sense. To maximise survival, animals must optimally coordinate sleep and wake, balancing conflicting needs (Eban-Rothschild et al. 2018). A period of heightened noradrenergic tone might reflect an environmental change or threat, making sleep riskier than usual. A sleep-inhibiting after-effect of heightened noradrenergic arousal might therefore be adaptive. In Drosophila, Seidner et al. (2015) found that activating octopaminergic circuitry—the invertebrate counterpart of the noradrenergic system (Roeder 1999)—during sleep deprivation suppressed subsequent rebound sleep. One possible interpretation of this result is that sleep need continued to build during sleep deprivation, but counter-balancing after-effects of octopaminergic potentiation suppressed rebound sleep. Similarly, Suzuki et al. (2013) observed that mice kept awake by their spontaneous exploration of novel environments, which would engage the LC, showed greater sleep latencies afterwards than animals sleep deprived by gentle handling. Findings in other species are therefore at least consistent with the idea that changes in waking levels of noradrenergic/octopaminergic arousal can inversely impact subsequent sleep. To test the idea that noradrenergic after-effects on sleep occur due to plastic changes in synaptic transmission, experiments could be performed that measure electrophysiological changes in GABAergic/galaninergic POA neurons following opto- or chemo-genetic manipulation of the LC.
Nonetheless, our observation that clonidine boosts both baseline sleep and caffeine-induced rebound sleep in dbh F0 KOs is not consistent with the idea that clonidine enhances rebound sleep solely via the after-effects of its suppression of noradrenergic transmission. Rather, clonidine’s action on α2-adrenoceptors that sit on glutamatergic axon terminals, reducing the release of glutamate, and/or clonidine’s postsynaptic action as a neuronal inhibitor may also contribute to the rebound sleep enhancement that we observed. In any case, the interpretation that heightened immediate early gene expression explains the heightened rebound sleep in noradrenergic-compromised larvae does not preclude direct effects of prior noradrenergic tone on subsequent sleep; the two ideas are not mutually exclusive.
Conclusion
Our results are consistent with previous findings in zebrafish that stimulant-induced rebound sleep increases as a function of preceding neuronal activity, as measured by c-fos expression. Additionally, we find that rebound sleep and c-fos expression are not dependent on heightened prior noradrenergic tone. In fact, reducing noradrenergic tone appears to enhance subsequent rebound sleep, perhaps by magnifying the increase in neuronal activity caused by the stimulant drug, as reflected by brain-wide levels of c-fos induction, and so augmenting a sleep pressure signal.
Data availability
Data are available on request from the authors.
References
Ahn S, Olive M, Aggarwal S, Krylov D, Ginty DD, Vinson C (1998) A dominant-negative inhibitor of CREB reveals that it is a general mediator of stimulus-dependent transcription of c-fos. Mol Cell Biol 18(2):967–977. https://doi.org/10.1128/MCB.18.2.967
Baraban SC, Taylor MR, Castro PA, Baier H (2005) Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience 131:759–768. https://doi.org/10.1016/j.neuroscience.2004.11.031
Barros VN, Mundim M, Testa Galindo L, Bittencourt S, Porcionatto M, Mello LE (2015) The pattern of c-Fos expression and its refractory period in the brain of rats and monkeys. Front Cell Neurosci 9:72. https://doi.org/10.3389/fncel.2015.00072
Benington JH, Heller HC (1995) Restoration of brain energy metabolism as the function of sleep. Prog Neurobiol 45:347–360. https://doi.org/10.1016/0301-0082(94)00057-O
Borbély AA, Achermann P (1999) Sleep homeostasis and models of sleep regulation. J Biol Rhythms 14(6):559–568. https://doi.org/10.1177/074873099129000894
Carter ME, Yizhar O, Chikahisa S, Nguyen H, Adamantidis A, Nishino S, Deisseroth K, de Lecea L (2010) Tuning arousal with optogenetic modulation of locus coeruleus neurons. Nat Neurosci 13(12):1526–1535. https://doi.org/10.1038/nn.2682
Carter ME, Brill J, Bonnavion P, Huguenard JR, Huerta R, de Lecea L (2012) Mechanism for Hypocretin-mediated sleep-to-wake transitions. PNAS 109(39):E2635–E2644. https://doi.org/10.1073/pnas.1202526109
Chamberlin NL, Arrigoni E, Chou TC, Scammell TE, Greene RW, Saper CB (2003) Effects of adenosine on GABAergic synaptic inputs to identified ventrolateral preoptic neurons. Neuroscience 119:913–918. https://doi.org/10.1016/S0306-4522(03)00246-X
Chandler DJ, Jensen P, McCall JG, Pickering AE, Schwarz LA, Totah NK (2019) Redefining noradrenergic neuromodulation of behavior: impacts of a modular locus coeruleus architecture. J Neurosci 39(42):8239–8249. https://doi.org/10.1523/JNEUROSCI.1164-19.2019
Cheng J, Wu F, Zhang M, Ding D, Fan S, Chen G, Zhang J, Wang L (2020) The interaction between the ventrolateral preoptic nucleus and the tuberomammillary nucleus in regulating the sleep-wakefulness cycle. Front Neurosci 14:615854. https://doi.org/10.3389/fnins.2020.615854
Cirelli C, Tononi G (2000) On the functional significance of c-fos induction during the sleep-waking cycle. Sleep 23(4):453–469
Cirelli C, Pompeiano M, Arrighi P, Tononi G (1995) Sleep-waking changes after c-fos antisense injections in the medial preoptic area. NeuroReport 6:801–805. https://doi.org/10.1097/00001756-199503270-00024
Cirelli C, Huber R, Gopalakrishnan A, Southard TL, Tononi G (2005) Locus ceruleus control of slow-wave homeostasis. J Neurosci 25(18):4503–4511. https://doi.org/10.1523/JNEUROSCI.4845-04.2005
De Luca R, Nardone S, Grace KP, Venner A, Cristofolini M, Bandaru SS, Sohn LT, Kong D, Mochizuki T, Viberti B, Zhu L, Zito A, Scammell TE, Saper CB, Lowell BB, Fuller PM, Arrigoni E (2022) Orexin neurons inhibit sleep to promote arousal. Nat Commun 13:1–15. https://doi.org/10.1038/s41467-022-31591-y
DeBock F, Kurz J, Azad SC, Parsons CG, Hapfelmeier G, Zieglgansberger W, Rammes G (2003) α2-Adrenoreceptor activation inhibits LTP and LTD in the basolateral amygdala: involvement of Gi/o-protein-mediated modulation of Ca2+-channels and inwardly rectifying K+-channels in LTD. Eur J Neurosci 17:1411–1424. https://doi.org/10.1046/j.1460-9568.2003.02544.x
Donlea JM, Pimentel D, Talbot CB, Kempf A, Omoto JJ, Hartenstein V, Miesenboeck G (2018) Recurrent circuitry for balancing sleep need and sleep. Neuron 97:378–389. https://doi.org/10.1016/j.neuron.2017.12.016
Du W, Zhang R, Li J, Yuan C, Yu T, Du J (2018) The locus coeruleus modulates intravenous general anesthesia of zebrafish via a cooperative mechanism. Cell Rep 24:3146–3155. https://doi.org/10.1016/j.celrep.2018.08.046
Eban-Rothschild A, Applebaum L, de Lecea L (2018) Neuronal mechanisms for sleep/wake regulation and modulatory drive. Neuropsychopharmacology 43:937–952. https://doi.org/10.1038/npp.2017.294
Farrar MJ, Kolkman KE, Fetcho JR (2018) Features of the structure, development, and activity of the zebrafish noradrenergic system explored in new CRISPR transgenic lines. J Comp Neurol 526:2493–2508. https://doi.org/10.1002/cne.24508
Fisher SP, Cui N, McKillop LE, Gemignani J, Bannerman DM, Oliver PL, Peirson SN, Vyazovskiy VV (2016) Stereotypic wheel running decreases cortical activity in mice. Nat Commun 7:13138. https://doi.org/10.1038/ncomms13138
Funato H, Miyoshi C, Fujiyama T, Kanda T, Sato M, Wang Z, Ma J, Nakane S, Tomita J, Ikkyu A, Kakizaki M, Hotta-Hirashima N, Kanno S, Komiya H, Asano F, Honda T, Kim SJ, Harano K, Muramoto H, Yonezawa T, Mizuno S, Miyazaki S, Connor L, Kumar V, Miura I, Suzuki T, Watanabe A, Abe M, Sugiyama F, Takahashi S, Sakimura K, Hayashi Y, Liu Q, Kume K, Wakana S, Takahashi JS, Yanagisawa M (2016) Forward-genetics analysis of sleep in randomly mutagenized mice. Nature 539:378–383. https://doi.org/10.1038/nature20142
Harris NA, Isaac AT, Gunther A, Merkel K, Melchior J, Xu M, Eguakun E, Perez R, Nabit BP, Flavin S, Gilsbach R, Shonesy B, Hein L, Abel T, Baumann A, Matthews R, Centanni SW, Winder DG (2018) Dorsal BNST α2-adrenergic receptors produce HCN-dependent excitatory actions that initiate anxiogenic behaviors. J Neurosci 38:8922–8942. https://doi.org/10.1523/JNEUROSCI.0963-18.2018
Ho J, Tumkaya T, Aryal S, Choi H, Claridge-Chang A (2019) Moving beyond P values: data analysis with estimation graphics. Nat Methods 16:565–566. https://doi.org/10.1038/s41592-019-0470-3
Hobson JA (2005) Sleep is of the brain, by the brain, and for the brain. Nature 437:1254–1256. https://doi.org/10.1038/nature04283
Hu FY, Hanna GM, Han W, Mardini F, Thomas SA, Wyner AJ, Kelz MB (2012) Hypnotic hypersensitivity to volatile anesthetics and dexmedetomidine in dopamine β-hydroxylase knockout mice. Anaesthesiology 117:1006–1017. https://doi.org/10.1097/ALN.0b013e3182700ab9
Hudson AE (2018) Genetic reporters of neuronal activity: c-Fos and G-CaMP6. Methods Enzymol 603:197–220. https://doi.org/10.1016/bs.mie.2018.01.023
Jasper JR, Lesnick JD, Chang LK, Yamanishi SS, Chang TK, Hsu SAO, Daunt DA, Bonhaus DW, Eglen RM (1998) Ligand efficacy and potency at recombinant α2 adrenergic receptors. Biochem Pharmacol 55:1035–1043. https://doi.org/10.1016/S0006-2952(97)00631-X
Jones BE (1991) The role of noradrenergic locus coeruleus neurons and neighboring cholinergic neurons of the pontomesencephalic tegmenturn in sleep-wake states. In: Pompeiano O, Barnes CD (eds) Progress in brain research, vol 88. Neurobiology of the locus coeruleus. Elsevier, Amsterdam, pp 533–543
Keene AC, Duboue ER (2018) The origins and evolution of sleep. J of Exp Biol 221:jeb159533. https://doi.org/10.1242/jeb.159533
Kim SJ, Hotta-Hirashima N, Asano F, Kitazono T, Iwasaki K, Nakata S, Komiya H, Asama N, Matsuoka T, Fujiyama T, Ikkyu A, Kakizaki M, Kanno S, Choi J, Kumar D, Tsukamoto T, Elhosainy A, Mizuno S, Miyazaki S, Tsuneoka Y, Sugiyama F, Takahashi S, Hayashi Y, Muratani M, Liu Q, Miyoshi C, Yanagisawa M, Funato H (2022) Kinase signalling in excitatory neurons regulates sleep quantity and depth. Nature 612:512–518. https://doi.org/10.1038/s41586-022-05450-1
Kroll F, Powell GT, Ghosh M, Gestri G, Antinucci P, Hearn TH, Tunbak H, Lim S, Dennis HW, Fernandez JM, Whitmore D, Dreosti E, Wilson SW, Hoffman EJ, Rihel J (2021) A simple and effective F0 knockout method for rapid screening of behaviour and other complex phenotypes. Elife 10:1–34. https://doi.org/10.7554/eLife.59683
Krueger JM, Nguyen JT, Dykstra-Aiello CJ, Taishi P (2019) Local sleep. Sleep Med Rev 43:14–21. https://doi.org/10.1016/j.smrv.2018.10.001
Kubra K, Gaddu GK, Liongue C, Heidary S, Ward AC, Dhillon AS, Basheer F (2022) Phylogenetic and expression analysis of Fos transcription factors in zebrafish. Int J Mol Sci 23:10098. https://doi.org/10.3390/ijms231710098
Labun K, Guo X, Chavez A, Church G, Gagnon J, Valen E (2019) Accurate analysis of genuine CRISPR editing events with ampliCan. Genome Res 29(5):843–847. https://doi.org/10.1101/gr.244293.118
Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R (2009) The sequence aligment/map format and SAMtools. Bioinformatics 25(16):2078–2079. https://doi.org/10.1093/bioinformatics/btp352
Liang Y, Shi W, Xiang A, Hu D, Wang L, Zhang L (2021) The NAergic locus coeruleus-ventrolateral preoptic area neural circuit mediates rapid arousal from sleep. Curr Biol 31:3729–3742. https://doi.org/10.1016/j.cub.2021.06.031
Liu S, Liu Q, Tabuchi M, Wu MN (2016) Sleep drive is encoded by neural plastic changes in a dedicated circuit. Cell 165(6):1347–1360. https://doi.org/10.1016/j.cell.2016.04.013
Ma Y, Miracca G, Yu X, Harding EC, Miao A, Yustos R, Vyssotski AL, Franks NP, Wisden W (2019) Galanin neurons unite sleep homeostasis and α2-adrenergic sedation. Curr Biol 29:3315–3322. https://doi.org/10.1016/j.cub.2019.07.087
McCarren HS, Chalifoux MR, Han B, Moore JT, Cheng Meng Q, Baron-Hionis N, Sedigh-Sarvestani M, Contreras D, Beck SG, Kelz MB (2014) α2-Adrenergic stimulation of the ventrolateral preoptic nucleus destabilizes the anesthetic state. J Neurosci 34(49):16385–16396. https://doi.org/10.1523/JNEUROSCI
Milinski L, Fisher SP, Cui N, McKillop LE, Blanco-Duque C, Ang G, Yamagata T, Bannerman DM, Vyazovskiy VV (2021) Waking experience modulates sleep need in mice. BMC Biol 19:65. https://doi.org/10.1186/s12915-021-00982-w
Nelson LE, Lu J, Guo T, Saper CB, Franks NP, Maze M (2003) The α2-adrenoceptor agonist dexmedetomidine converges on an endogenous sleep-promoting pathway to exert its sedative effects. Anesthesiology 98:428–436. https://doi.org/10.1097/00000542-200302000-00024
Ouyang M, Hellman K, Abel T, Thomas SA (2004) adrenergic signaling plays a critical role in the maintenance of waking and in the regulation of REM sleep. J Neurophysiol 92:2071–2082. https://doi.org/10.1152/jn.00226.2004
Perez DM (2020) α1-Adrenergic receptors in neurotransmission, synaptic plasticity, and cognition. Front Pharmacol 11:581098. https://doi.org/10.3389/fphar.2020.581098
Pollina EA, Gilliam DT, Landau AT, Lin C, Pajarillo N, Davis CP, Harmin DA, Yap E-L, Vogel IR, Griffith EC, Nagy MA, Ling E, Duffy EE, Sabatini BL, Weitz CJ, Greenberg ME (2023) A NPAS4–NuA4 complex couples synaptic activity to DNA repair. Nature 614:732–741. https://doi.org/10.1038/s41586-023-05711-7
Porkka-Heiskanen T, Kalinchuk AV (2011) Adenosine, energy metabolism and sleep homeostasis. Sleep Med Rev 15:123–135. https://doi.org/10.1016/j.smrv.2010.06.005
Prober DA, Rihel J, Onah AA, Sung R-J, Schier AF (2006) Hypocretin/orexin overexpression induces an insomnia-like phenotype in zebrafish. J Neurosci 26(51):13400–13410. https://doi.org/10.1523/JNEUROSCI.4332-06.2006
Quinlan AR, Hall IM (2010) BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26(6):841–842. https://doi.org/10.1093/bioinformatics/btq033
Rasch B, Born J (2013) About sleep’s role in memory. Physiol Rev 93:681–766. https://doi.org/10.1152/physrev.00032.2012
Reichert S, Pavon Arocas O, Rihel J (2019) The neuropeptide galanin is required for homeostatic rebound sleep following increased neuronal activity. Neuron 104:370–384. https://doi.org/10.1016/j.neuron.2019.08.010
Rihel J, Prober DA, Arvanites A, Lam K, Zimmerman S, Jang S, Haggarty SJ, Kokel D, Rubin LL, Peterson RT, Schier A (2010) Zebrafish behavioural profiling links drugs to biological targets and rest/wake regulation. Science 327:348–351. https://doi.org/10.1126/science.1183090
Roeder T (1999) Octopamine in Invertebrates. Prog Neurobiol 59:533–561. https://doi.org/10.1016/S0301-0082(99)00016-7
Saper CB, Fuller PM, Pederson NP, Lu J, Scammell TE (2010) Sleep state switching. Neuron 68:1023–1042. https://doi.org/10.1016/j.neuron.2010.11.032
Seidner G, Robinson JE, Wu M, Worden K, Masek P, Roberts SW, Keene AC, Joiner WJ (2015) Identification of neurons with a privileged role in sleep homeostasis in Drosophila melanogaster. Curr Biol 25:2928–2938. https://doi.org/10.1016/j.cub.2015.10.006
Shields AD, Wang Q, Winder DG (2009) α2A-Adrenergic receptors heterosynaptically regulate glutamatergic transmission in the bed nucleus of the stria terminalis. Neuroscience 163:339–351. https://doi.org/10.1016/j.neuroscience.2009.06.022
Singh C, Oikonomou G, Prober D (2015) Norepinephrine is required to promote wakefulness and for hypocretin-induced arousal in zebrafish. Elife 4:e07000. https://doi.org/10.7554/eLife.07000
Steininger TL, Gong H, McGinty D, Szymusiak R (2001) Subregional organization of preoptic area/anterior hypothalamic projections to arousal-related monoaminergic cell groups. J Comp Neurol 429:638–653
Suzuki A, Sinton CM, Greene RW, Yanagisawa M (2013) Behavioral and biochemical dissociation of arousal and homeostatic sleep need influenced by prior wakeful experience in mice. PNAS 110(25):10288–10293. https://doi.org/10.1073/pnas.1308295110
Szymusiak R, McGinty D (2008) Hypothalamic regulation of sleep and arousal. Ann N Y Acad Sci 1129(1):275–286. https://doi.org/10.1196/annals.1417.027
Teng S, Zhen F, Wang L, Canovas Schalchli J, Simko J, Chen X, Jin H, Makinson CD, Peng Y (2022) Control of non-REM sleep by ventrolateral medulla glutamatergic neurons projecting to the preoptic area. Nat Commun 13:4748. https://doi.org/10.1038/s41467-022-32461-3
Thomas CW, Guillaumin MCC, McKillop LE, Achermann P, Vyazovskiy VV (2020) Global sleep homeostasis reflects temporally and spatially integrated local cortical neuronal activity. Elife 9:e54148. https://doi.org/10.7554/eLife.54148
Vassalli A, Franken P (2017) Hypocretin (orexin) is critical in sustaining theta/gamma-rich waking behaviors that drive sleep need. PNAS 114(27):E5464–E5473. https://doi.org/10.1073/pnas.1700983114
Wang S, Wang Z, Mu Y (2022) Locus coeruleus in non-mammalian vertebrates. Brain Sci 12:134. https://doi.org/10.3390/brainsci12020134
Yamagata T, Kahn MC, Prius-Mengual J, Meijer E, Sabanovic M, Guillaumin MCC, van der Vinne V, Huang Y, McKillop LE, Jagannath A, Peirson SN, Mann EO, Foster RG, Vyazovskiy VV (2021) The hypothalamic link between arousal and sleep homeostasis in mice. PNAS 118(51):e2101580118. https://doi.org/10.1073/pnas.2101580118
Yin N, Jin X, He J, Yin Z (2009) Effects of adrenergic agents on the expression of zebrafish (Danio rerio) vitellogenin Ao1. Toxicol Appl Pharmacol 238:20–26. https://doi.org/10.1016/j.taap.2009.04.004
Yuan JS, Reed A, Chen F, Stewart CN (2006) Statistical analysis of real-time PCR data. BMC Bioinform 7:85. https://doi.org/10.1186/1471-2105-7-85
Zada D, Sela Y, Matosevich N, Monsonego A, Lerer-Goldshtein T, Nir Y, Appelbaum L (2021) Parp1 promotes sleep, which enhances DNA repair in neurons. Mol Cell 81:4979–4993. https://doi.org/10.1016/j.molcel.2021.10.026
Zhong P, Zhang Z, Barger Z, Ma C, Liu D, Ding X, Dan Y (2019) Control of non-REM sleep by midbrain neurotensinergic neurons. Neuron 104:795–809. https://doi.org/10.1016/j.neuron.2019.08.026
Zhou R, Wang G, Li Q, Meng F, Liu C, Gan R, Ju D, Liao M, Xu J, Di S, Gao X, Zhou S, Wu K, Sun Q, Guo Y, Wu C, Chen Z, Chen L, Shi B, Wang H, Wang X, Li H, Cai T, Li B, Wang F, Funato H, Yanagisawa M, Erquan Zhang E, Liu Q (2022) A signalling pathway for transcriptional regulation of sleep amount in mice. Nature 612:519–527. https://doi.org/10.1038/s41586-022-05510-6
Acknowledgements
David Prober kindly provided us with the underlying dbh-/- null mutant sleep data (Singh et al. 2015). We also thank the UCL Fish Facility for fish husbandry, Sumi Lim and Amanda Tan for qRT-PCR guidance, Francois Kroll for instruction in both designing targets for F0 knockout and in MiSeq analysis, and all Rihel lab members for their ad hoc help and thoughts.
Funding
This work was supported by an EMBO Fellowship awarded to DGL (ALTF 1097-2016), a European Research Council Starting Grant (282027) to JR, and a Wellcome Trust Investigator Award (217150/Z/19/Z) to JR.
Author information
Authors and Affiliations
Contributions
All authors contributed to the conception and design of the study and interpretation of the results. EB performed the experiments, statistically analysed the data, prepared the figures and wrote the first draft of the manuscript. JR and DGL discussed and critically edited draft versions of the manuscript, and all authors read and approved the final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethical approval
Experiments and zebrafish husbandry followed UCL fish facility protocols per project licence PA8D4D0E5, awarded to Jason Rihel by the UK Home Office under the UK Animals (Scientific Procedures) Act 1986.
Additional information
Communicated by V. Vyazovskiy.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Benoit, E., Lyons, D.G. & Rihel, J. Noradrenergic tone is not required for neuronal activity-induced rebound sleep in zebrafish. J Comp Physiol B 194, 279–298 (2024). https://doi.org/10.1007/s00360-023-01504-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00360-023-01504-6