
Abstract
A combined micropalaeontological, taphonomical and sedimentological study was carried out in the non-marine Barremian of the Maestrat Basin (E Iberian Chain) to elucidate the palaeoecology of Barremian charophytes. Understanding the palaeoecology of fossil charophytes is of prime importance for the accurate application of charophyte biozonations, since most species used as biostratigraphic markers and/or biozone index species are restricted to facies formed in a particular environment. Five charophyte biocoenoses were distinguished. (1) Charophyte association from freshwater alkaline lakes receiving low clastic input, mainly composed of Atopochara trivolvis var. triquetra, Clavator harrisii, Ascidiella stellata, and A. triquetra. The thalli known as Munieria grambastii, corresponding to Clavator utricles, were extremely abundant in the shallowest facies of the lake margins, and Hemiclavator-rich populations were also locally dominant. (2) An association characteristic of freshwater alkaline lakes with high clastic input was mainly composed of Echinochara lazarii, Globator maillardii var. trochiliscoides, A. trivolvis var. triquetra, C. harrisii, Hemiclavator neimongolensis var. neimongolensis, and locally also Clavator calcitrapus. (3) Monospecific populations of Porochara maestratica thrived in brackish settings with little clastic influence, whereas (4) monospecific E. lazarii populations thrived in clastic-influenced brackish settings. (5) A charophyte association grew in lakes that developed in mudflat to floodplain environments and was mainly constituted by E. lazarii, A. trivolvis var. triquetra, and C. harrisii var. harrisii and var. reyi. Accordingly, the palaeoenvironmental conditions inferred indicate that most of the charophyte species were controlled by (i) bathymetry, (ii) clastic content in the sedimentary environment and (iii) salinity. The stratigraphic succession studied shows an upwards increase in clastic content from the base to the top, which is associated with the more humid period recorded along the margins of the Tethys and the Boreal realm during the early Barremian–early late Barremian. In consequence, those species restricted to clastic-poor settings are constrained to the lower part of the section, whereas those taxa that were able to thrive in clastic-rich settings persisted throughout the entire section. This observation establishes a link between the changes observed in the charophyte floras recognized and the environmental changes that occurred during the Barremian in western Europe.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Charophyte remains, including thalli and fructifications (utricles and gyrogonites), are among the most abundant components of Early Cretaceous non-marine marls, clays and limestones of the western Tethys (e.g., Martín-Closas and Diéguez 1998; Climent-Domènech et al. 2009; Trabelsi et al. 2016). Three charophyte families coexisted during the Early Cretaceous, the Porocharaceae, Characeae, and Clavatoraceae, with the last being the most common and abundant in the Tethyan Domain (e.g., Martín-Closas and Grambast-Fessard 1986; Schudack 1993; Martín-Closas 2000; Trabelsi et al. 2016; Pérez-Cano et al. 2020). In view of their high specific diversity and evolutionary rates, clavatoracean fructifications (utricles) found in marls and clays have been extensively used as biostratigraphic markers in the continental record (e.g., Grambast 1974; Riveline et al. 1996; Martín-Closas et al. 2009; Pérez-Cano et al. 2022).
Despite extensive taxonomic knowledge of clavatoracean morphology and its widespread biostratigraphic application in the Early Cretaceous record, the palaeoenvironmental distribution of most species is still poorly known or understood. Previous palaeoecological studies of Early Cretaceous charophytes in general and clavatoraceans in particular have usually been limited to a few genera (Martín-Closas and Grambast-Fessard 1986; Schudack 1993) or a few species (e.g., Vicente and Martín-Closas 2013). Palaeoecological studies carried out in the Campanian–Danian of the Pyrenees (Vicente et al. 2015, 2016) and the Eocene–Oligocene of the Ebro Basin (Sanjuan and Martín-Closas 2012) have demonstrated that charophyte palaeoecology is of major importance in charophyte biostratigraphy, as species that are useful for biostratigraphic characterization of the non-marine record may be restricted to particular facies. The aforementioned studies also show that species with a more restricted biostratigraphic distribution usually evidence more stringent palaeoecological requirements, being absent in stratigraphic successions that do not record the particular facies in which the species preferably thrived.
The present study was carried out in a continuous early Barremian to early late Barremian section from the Maestrat Basin (E Iberian Chain) with the aim of determining the palaeoecology of Barremian charophytes. To this end, a combined sedimentological and taphonomical study was performed of charophyte-rich marls and clays, together with a study of charophyte-rich microfacies from limestones. A better understanding of the palaeoecology of Barremian charophyte species, mainly those used as index species and/or as biostratigraphic indicators, will improve the precision and accuracy of Barremian charophyte biozonation. Moreover, a taphonomic and palaeoecological characterization of the Barremian charophyte associations will provide a tool to perform more detailed palaeoenvironmental reconstructions of coeval non-marine settings.
Geological setting
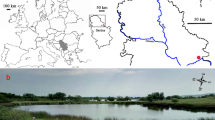
The Maestrat Basin is located in the eastern Iberian Chain (Fig. 1A). During the Late Jurassic and Early Cretaceous, basin evolution was mainly controlled by a rifting process, which recorded three discrete rift/post-rift stages (Salas et al. in Martín-Chivelet et al. 2019). These are (1) Kimmeridgian–Berriasian tectonic rifting, mainly related to the opening of the North Atlantic sea floor, (2) late Berriasian−late Hauterivian post-rift, in which the creation of depositional space was mainly controlled by thermal subsidence, and (3) Barremian–early Albian rifting, which was mainly associated with the opening of the Bay of Biscay (Tugend et al. 2015). Listric faults and local uplift shoulders produced a clear division of the Maestrat Basin into several smaller sub-basins (Fig. 1B; Salas and Guimerà 1996; Salas et al. 2001; Salas et al. in Martín-Chivelet et al. 2019). The present-day structure and topographic relief are the result of tectonic inversion of the Mesozoic structures, which occurred during the Alpine Orogeny, lasting from the late Eocene to the early Miocene (e.g., Guimerà 2018; Nebot and Guimerà 2018).

modified from Salas et al. in Martín-Chivelet et al. 2019), with indication of the thickness of Upper Jurassic to Lower Cretaceous deposits. CB: Cedramán Sub-Basin; GB: Galve Sub-Basin; LB: Las Parras Sub-Basin; MB: Morella Sub-Basin; OB: Oliete Sub-Basin; OrB: Orpesa Sub-Basin; PB: Penyagolosa Sub-Basin; PeB: El Perelló Sub-Basin; SB: Salzedella Sub-Basin. Scale bar equals 50 km. C Geological map of the Herbers area and location of the section (modified from Canérot and Leyva 1976). Scale bar equals 500 m
A Map of the Iberian Peninsula and the location of the Maestrat Basin. Scale bar equals 200 km. B Sub-basins of the Maestrat Basin (
The Herbers-Mas de Petxí section studied here crops out very close to the Herbers Fault, on the northern margin of the Morella Sub-Basin (Fig. 1B, C). The Barremian record in this area is very continuous and thick (ca. 800 m), and can be subdivided into five formations named the Cantaperdius, Artoles, Morella, Cervera del Maestrat and Xert (Fig. 2), which comprise almost all the Barremian deposition in the Maestrat Basin (Bover-Arnal et al. 2016). During the Barremian, the Maestrat Basin was a gulf that opened onto the Tethys Ocean with different depositional settings and palaeonvironmental conditions (Salas 1987). The base of the Barremian is characterized by the occurrence of lateritic deposits (e.g., Combes 1969; Salas 1987; Yuste et al. 2017) overlying the Hauterivian freshwater limestones of the Herbers Formation or older units (Fig. 2). Above the laterite, the Cantaperdius Formation is mainly built up of lacustrine and palustrine carbonates and reaches a thickness of ~ 400 m on the northern margin of the Morella Sub-Basin. This lithostratigraphic unit passes laterally and vertically to the shallow-marine marls and limestones of the Artoles Formation (Fig. 2). In the central part of the basin, the Artoles Formation is ~ 700 m thick, whereas at the margins its thickness decreases to ~ 200 m (Salas 1987; Salas et al. 2001).

The upper part of the Artoles Formation changes laterally and vertically to the Morella Formation (Fig. 2), which consists of red and grey clays and cross-bedded sandstones and contains abundant dinosaur remains (e.g., Gasulla et al. 2015). These deposits have been attributed to a tidal plain or mudflat environment (e.g., Canérot et al. 1982; Gàmez et al. 2003) and in turn, they pass laterally and vertically to the coastal and shallow-marine deposits of the Cervera del Maestrat Formation (Fig. 2; Salas 1987; Bover-Arnal et al. 2016). Above, the latest Barremian marine carbonates rich in orbitolinids of the Xert Formation occur (Canérot et al. 1982; Bover-Arnal et al. 2016).
The lithostratigraphic units rich in charophyte remains studied here are the Cantaperdius, Artoles, and Morella formations (Fig. 2). The age of this succession is early Barremian–early late Barremian, based on charophyte biostratigraphy and strontium-isotope stratigraphy (Martín-Closas 1989; Martín-Closas and Salas 1994; Pérez-Cano et al. 2022).
Materials and methods
The present study was carried out at Herbers-Mas de Petxí section (base 40º 42′ 39ʺ N, 0º 0′ 36ʺ W; top 40º 42′ 34ʺ N, 0º 0′ 30ʺ E), which is a 690 m thick complete section of Barremian non-marine facies (Supplementary material 1). It crops out on the north-eastern margin of the Maestrat Basin (Fig. 1C). One hundred and twelve limestone beds were sampled from the Cantaperdius, Artoles, and Morella formations for microfacies analysis. For each sample, two 30 µm thick thin sections were made, one parallel and the other perpendicular to the stratification. This methodology enables a better observation of elongate components, such as charophyte thalli. The thin sections were studied under a Motic BA 310 petrographic microscope and photographed using Motic Images Plus 2.0 software.
Marls from the same formations were systematically sampled to study the micropalaeontological assemblages, especially charophytes (Fig. 3). For each interval, around 3 kg of marls were collected. The samples were submerged in a solution of water, sodium carbonate (Na2CO3), and hydrogen peroxide (H2O2) for several days to disaggregate the marls, deflocculate clay minerals, and eliminate organic matter. Later, the samples were wet-sieved using three different mesh sizes of 1000, 500 and 200 μm. Once the samples were dried, microfossils (charophyte fructifications and thalli, ostracods and other associated microfossils) were handpicked under a Wild M5A binocular microscope using magnifications of × 12 and × 25 at the Departament de Dinàmica de la Terra i de l’Oceà (Universitat de Barcelona). Selected fossil remains were photographed with a Quanta 200 SEM at the Centres Científics i Tecnològics de la Universitat de Barcelona (CCiTUB). The figured specimens and thin sections with figured specimens have been deposited in the Museu Geològic del Seminari Conciliar de Barcelona (MGSCB) with reference numbers MGSCB 85570–85677 and 87440–87477, whereas the remaining samples are housed in the Departament de Dinàmica de la Terra i de l’Oceà, Facultat de Ciències de la Terra, Universitat de Barcelona.

Levigated remains of charophytes from the Herbers-Mas de Petxí section. A, B Porochara maestratica (sample H8). A Lateral view (specimen no. 85579 MGSCB); B Apical view (specimen no. 85580 MGSCB); C aff. Mesochara harrisii. (lateral view; sample H39; specimen no. 85585 MGSCB); D Echinochara lazarii (outer series of utricle; sample H88; specimen no. 85601 MGSCB); E Globator maillardii var. trochiliscoides (lateral view; sample H27; specimen no. 85602 MGSCB); F Globator maillardii var. biutricularis (lateral view; sample H44; specimen no. 85605); G Atopochara trivolvis var. triquetra (lateral view; sample H51; specimen no. 85608 MGSCB); H Atopochara trivolvis var. trivolvis (lateral view; sample H94; specimen no. 85611 MGSCB); I Clavator grovesii var. jiuquanensis (lateral view; sample H95; specimen no. 85614 MGSCB); J Clavator harrisii var. dongjingensis (lateral view; sample H38; specimen no. 85617 MGSCB); K Clavator harrisii var. harrisii (lateral view; sample H38; specimen no. 85620 MGSCB); L Clavator harrisii var. reyi (lateral view; sample H76; specimen no. 85622 MGSCB); M Clavator calcitrapus var. jiangluoensis (apical view; sample H38; specimen no. 85623 MGSCB); N Clavator calcitrapus var. calcitrapus (apical view; sample H51; specimen no. 85626 MGSCB); O Ascidiella stellata var. stellata (lateral view; sample H6; specimen no. 85677); P Ascidiella stellata var. lata (lateral view; sample H44; specimen no. 85632 MGSCB); Q Ascidiella triquetra (apical view; sample H27; specimen no. 85637 MGSCB); R Ascidiella cruciata (apical view; sample H94; specimen no. 85639 MGSCB); S Hemiclavator adnatus (lateral view; sample H12; specimen no. 85642 MGSCB); T Hemiclavator neimongolensis var. posticecaptus (lateral view; sample H12; specimen no. 85645 MGSCB); U Hemiclavator neimongolensis var. neimongolensis (adaxial view; sample H39; specimen no. 85647 MGSCB); V Pseudoglobator paucibracteatus (lateral view; sample H96; specimen no 85659 MGSCB); W Charaxis spicatus (sample H124; specimen no. 87440 MGSCB); X Munieria grambastii (sample H5; specimen no. 85656 MGSCB); Y Clavatoraxis sp. (sample H8; specimen no. 85658 MGSCB); Z Favargerella aquavivae (sample H7 specimen no. 85659 MGSCB). Scale bars equal 250 µm
Palaeoecological inferences were drawn following a taphonomic analysis of the charophyte remains and associated microfossils combined with a sedimentological study. This methodology has been demonstrated as useful to infer the palaeoecology of charophyte species (e.g., Sanjuan and Martín-Closas 2012; Vicente et al. 2016). The taphonomic analysis made it possible to distinguish between autochthonous/parautochthonous and allochthonous fossils in the palaeontological association. Two criteria were applied in the taphonomic analysis: (1) fossil abrasion and fragmentation and, in the case of charophytes, (2) the association of fructifications and thalli attributed to the same species, which has been considered as evidence of absence of selection. The relative abundance of fructifications was obtained after analyzing ca. 100 g of sample, ca. 50 g from the > 500 µm–1 mm mesh size sample and ca. 50 g of the > 200–500 µm mesh size sample. Supplementary materials 2 and 3 show the total number of charophyte fructifications collected from each sample and the relative abundance of each taxon in each sample, respectively.
Results
Based on the lithology, sedimentary features and microfossils, mainly charophytes, 19 different facies (taphofacies and/or biofacies) were identified in four major lithostratigraphic intervals in the Herbers-Mas de Petxí section (Figs. 3, 4A–D; Tables 1, 2). These facies are located above a laterite characteristic of the base of the Barremian in the Maestrat Basin (Fig. 5A) and are arranged into different parasequences (Ps) (Ps1–Ps8). The repetition and alternation of these eight parasequences builds up the studied section. The definition of ‘parasequence’ used here follows Spence and Tucker (2007) who proposed applying this term to all types of meter-scale sedimentary packages characterized by a succession of facies.




A Stratigraphic log of the lower part of the Herbers-Mas de Petxí section (0–280 m), including the facies recognized. Facies codes are explained in the text. See legend in (D). B Stratigraphic log of the middle part of the Herbers-Mas de Petxí section (280–450 m), showing the facies recognized. Facies codes are explained in the text. See legend in (D). C Stratigraphic log of the upper part of the Herbers-Mas de Petxí section (450–690 m), showing the facies recognized. Facies codes are explained in the text. See legend in (D). D Legend for Figs. 4A–C and 8

Field photographs of the Herbers-Mas de Petxí section. A Basal part of the Cantaperdius Formation, showing the basal laterite and the first lacustrine limestones above; B Characteristic marl–limestone succession of a limestone-dominated interval of the Cantaperdius Formation; C Characteristic marl–limestone succession of a marl-dominated interval of the Cantaperdius Formation; D Outcrop of the Artoles Formation showing the characteristic marl–limestone inter-bedding; E Large-scale cross-bedding exhibited by facies bgL (Echinochara-rich marls and limestones); F Outcrop of the Morella Formation made up of Clavatoracean-rich clays and limestones showing an alternation between grey and red clays. Encircled hammer for scale (E and F).
Clavatoracean-rich limestones and marls from the Cantaperdius Formation
Description of facies and their charophyte assemblages
1) Clavatoracean-rich marls (cM) contain a micropalaeontological assemblage mainly composed of charophyte thalli and fructifications (utricles and gyrogonites), and ostracods (Table 2).
Neither the charophyte assemblages nor the preservation of charophyte remains within the marls is homogeneous throughout the section, and vertical changes are apparent in the taxonomic composition of charophyte assemblages (Tables 2, 3; Supplementary materials 2–3). In the lower part of the Cantaperdius Formation (11–274 m; Fig. 4A; Table 3), this facies is characterized by the occurrence of thin marl beds with charophyte assemblages composed of abundant, well-preserved utricles of Atopochara trivolvis var. triquetra, Clavator harrisii var. dongjingensis and var. harrisii, Ascidiella stellata var. stellata and var. lata and A. triquetra, and gyrogonites of aff. Mesochara harrisii (Fig. 3). The utricles are associated with abundant, long portions of thalli belonging to Favargerella aquavivae, Clavatoraxis sp., and Munieria grambastii (Fig. 3; Table 3). The Ascidiella utricles are sometimes found attached to the F. aquavivae thalli, while C. harrisii var. dongjingensis and var. harrisii are found attached to Clavatoraxis sp. thalli, indicating autochthony. Porochara maestratica is locally abundant (Table 3), and it is interpreted as parautochthonous in this facies. Globator maillardii var. trochiliscoides is usually rare in the lowermost part of the Cantaperdius Formation, where it locally occurs superficially eroded and thus, is interpreted as parautochthonous. The larger populations of G. maillardii var. trochiliscoides are found in the thicker marl beds occurring between 100 and 187 m of the stratigraphic section (Fig. 4A; Table 3), associated with a few utricles of Echinochara lazarii and portions of Charaxis spicatus (Table 3). The latter two taxa belong to the same charophyte plant (Pérez-Cano et al. 2020). G. maillardii var. trochiliscoides and E. lazarii are interpreted as being autochthonous in these thicker marl beds. Abundant, well-preserved Hemiclavator adnatus and Hemiclavator neimongolensis var. posticecaptus are limited to marls between meters 129 and 130 of the section (Table 3; Supplementary materials 2–3). These utricles are found attached to portions of Clavatoraxis sp., suggesting autochthony.
The charophyte assemblage of marls in the upper part of the Cantaperdius Formation (274–366 m; Fig. 4B; Table 4) consists of abundant, well-preserved utricles of E. lazarii, G. maillardii var. trochiliscoides and var. biutricularis, A. trivolvis var. triquetra, C. harrisii var. harrisii, and var. reyi, and H. neimongolensis var. neimongolensis. In addition, Clavator calcitrapus var. jiangluoensis and var. calcitrapus (Fig. 3) are also common but less abundant (Table 4; Supplementary materials 2–3). E. lazarii utricles are attached to their corresponding thalli, C. spicatus (Fig. 3), suggesting autochthony. The same interpretation is proposed for the utricles of C. harrisii var. harrisii and H. neimongolensis var. neimongolensis, which occur attached to thalli of Clavatoraxis sp. Thalli portions of M. grambastii are abundant and display up to 4 anatomically attached internodes, and are also interpreted as autochthonous remains. Other species such as Ascidiella stellata var. lata, A. triquetra and thalli portions of F. aquavivae are rare (Table 4; Supplementary materials 2–3), suggesting some type of biostratinomic or palaeoecological selection.
This facies is interpreted as having been formed by deposition of suspended mud through decantation processes in low-energy, relatively deeper lacustrine settings. The abundance and diversity of charophyte species and the taphonomic evidence suggests that rich clavatoracean meadows developed in these, mostly freshwater, lakes. However, the occurrence of rare and broken and/or superficially eroded dasycladaleans and benthic foraminifera (see Table 2) is interpreted as occasional marine influence, probably during higher tides or storms.
2) Limestones rich in clavatoracean utricles and thalli (cL) are characterized by the occurrence of long portions of charophyte thalli belonging to Clavatoraxis sp. (Figs. 6A, 7A–C; Supplementary material 4.1, Fig. A–E) and Favargerella sp. (Fig. 7D–F; Supplementary material 4.1, Fig. F–H). These thalli are usually associated with well-preserved atopocharoid utricles (Supplementary material 4.2, Fig. A–C), such as Atopochara sp. (Fig. 7G, H), and clavatoroid utricles (Fig. 6A; Supplementary material 4.2, Fig. D–O). Some of these clavatoroid utricles were assigned to Ascidiella sp. based on their characteristic bract cell distribution in cross-section, with 2 bracts and the phylloid arranged 120º around the gyrogonite (Fig. 7I). Thalli and utricles are commonly anatomically attached (Fig. 7J, K; Supplementary material 4.2, Fig. M–O), suggesting autochthony. Thalli portions and clavatoroid utricles are locally coated by up to 300 µm thick homogeneous cyanobacterial crusts. This coating is interpreted as having been formed by calcification of the periphyton (Martín-Closas 1999). Rare remains of thalli provisionally attributed to Palaeonitella aff. trifurcata (Martín-Closas et al. 2021) within cortoids are occasionally found in these limestones (Supplementary materials 4.1, Fig. I–L).

Microfacies of Clavatoracean-rich limestones and marls from the Cantaperdius Formation. A Facies cL showing a wackestone–packstone texture, which contains a utricle (u), and Clavatoraxis sp. (c) (sample H10c; thin section no. 87446 MGSCB); B Facies mL shows a packstone texture and mainly displays transversal sections of Munieria (m) (sample H47b; thin section no. 85671 MGSCB); C Facies pL showing the association of Porochara sp. gyrogonites (p) with Charaxis spicatus (cs) and dasycladaleans (d) (sample H35b; thin section no. 87454 MGSCB); D Facies pccL showing coated portions of Favargerella (f) and Clavatoraxis (c) (sample H7c; thin section no. 87445 MGSCB); E Facies pccL showing porocharacean gyrogonites (p) associated with coated portions of Clavatoraxis sp. (c) and utricles (u) (sample H20b; thin section no. 87450 MGSCB); F Facies pccL showing dasycladaleans (d) associated with Clavatoraxis sp. (c) (sample H20c; thin section no. 87451 MGSCB); G Irregular oncoid of facies oL (sample H5d; thin section no. 87442 MGSCB). Scale bar A–G equals 1000 µm

Charophyte components of Clavatoracean-rich limestones from the Cantaperdius Formation. A Tangential section of Clavatoraxis sp. thallus (sample H10c; thin section no. 87447 MGSCB); B Longitudinal section of a Clavatoraxis sp. thallus (sample H7b; thin section no. 85665 MGSCB); C Transversal section through a Clavatoraxis sp. thallus (sample H10c; thin section no. 87446 MGSCB); D Tangential section of a Favargerella sp. branchlet (sample H10e; thin section no. 87449 MGSCB); E Longitudinal section of a Favargerella sp. branchlet (sample H10c; thin section no. 87447 MGSCB); F Transversal section of a Favargerella sp. node (sample H10e; thin section no. 87448 MGSCB); G Longitudinal section of a utricle belonging to Atopochara sp. (sample H6d; thin section no. 87443 MGSCB); H Transversal section of a utricle belonging to Atopochara sp. (sample H7B; thin section no. 85665 MGSCB); I Transversal section of an Ascidiella sp. utricle showing sections of bracts (b) and phylloids (p) (sample H20C; thin section no. 85670 MGSCB); J Longitudinal section of a clavatoroid utricle attached to a Clavatoraxis sp. branchlet (sample H20C; thin section no. 85670 MGSCB); K Clavatoroid utricle (u) attached to a Favargerella sp. branchlet (sample H7C; thin section no. 87444 MGSCB); L Clavatoroid utricle in facies mL (sample H47b; thin section no. 87455 MGSCB); M Longitudinal section of Munieria grambastii (sample H5b; thin section no. 87441 MGSCB); N Longitudinal section of a porocharacean gyrogonite (sample H35b; thin section no. 87453 MGSCB); O Oblique section through the basal part of a porocharacean gyrogonite. The arrow indicates the basal plate (sample H35b; thin section no. 87454 MGSCB); P Transversal section of a Charaxis sp. thallus (sample H35b; thin section no. 87454 MGSCB); Q Transversal section of a Clavatoraxis sp. thallus coated with a microbial crust (sample H20c; thin section no. 87452 MGSCB); R Clavatoroid utricle coated with microbial Thickets (arrowhead) (sample H7c; thin section no. 87445 MGSCB). Scale bar A–R equals 250 µm
This facies is interpreted as having been formed in shallow lacustrine belts with clavatoracean meadows dominated by Atopochara sp., and clavatoroids, including Ascidiella sp. The utricles of these species are associated with their corresponding vegetative remains, i.e., Clavatoraxis sp. (for Atopochara sp. and many clavatoroids) and Favargerella sp. (for Ascidiella sp.).
3) Munieria grambastii-rich limestones (mL) are characterized by the dominant occurrence of long, well-articulated portions of Munieria grambastii internodes (Fig. 7M), rarely associated with clavatoroid utricles (Fig. 7L). A few portions of Favargerella sp. and Clavatoraxis sp. thalli coated with periphyton crusts are sometimes found in addition to M. grambastii.
Facies mL is interpreted as having been formed by the deposition of in-situ biocalcified remains of M. grambastii, which would have formed almost monospecific meadows. The occurrence of intraclasts and rootmarks (see Table 2) suggests that these meadows occurred near the lake shores.
4) Limestones with porocharaceans (pL). This facies is not common in the studied section. The charophyte assemblage is composed of abundant well-preserved porocharacean gyrogonites (Fig. 7N–O) showing the characteristic multipartite basal plate, which have been attributed to Porochara sp. and are associated with their possible thalli, Charaxis sp. (Fig. 7P). A few well-preserved utricles of Echinochara sp. and portions of their corresponding thalli, Charaxis spicatus (Fig. 6C), also occur. This co-occurrence of thalli and utricles suggests that these charophytes are autochthonous in the depositional setting. Cypridean and non-cypridean ostracods, benthic foraminifera and dasycladalean portions are also present (Fig. 6C; Table 2).
Facies pL has been interpreted as formed in brackish lakes, as attested by the occurrence of dominant assemblages of genus Porochara (e.g., Martín-Closas and Grambast 1986; Mojon 1989; Schudack 1993; Climent-Domènech et al. 2009). The occurrence of dasycladaleans and benthic foraminifera indicates that these lakes occurred in a coastal area.
5) Limestones with porocharaceans and coated clavatoraceans (pccL) are associated with facies cL in the same bed, with pccL overlying cL (Fig. 4A). The fossil assemblage in this facies mainly consists of abundant well-preserved porocharacean gyrogonites associated with clavatoracean utricles (Fig. 6E; Table 2). Charophyte thalli are represented by long portions of Clavatoraxis sp. and Favargerella sp., including several articulated internodes (Fig. 6D, E; Supplementary Material 4.1, Fig. B, D, F), and less-abundant Charaxis sp. (Fig. 6F).
Porocharacean gyrogonites are usually well preserved and associated with portions of Charaxis sp. thalli. These remains are interpreted as being parts of the same plant, and are thus considered autochthonous in the depositional setting. The preservation of clavatoracean remains is similar to that described in facies cL, also suggesting autochthony. However, clavatoracean remains in facies pccL are always coated by cyanobacterial crusts (Figs. 6D, 7Q–R). Crusts around thalli (Favargerella sp. and Clavatoraxis sp.) are regular in thickness and display calcified entangled filaments (Fig. 7Q), whereas clavatoroid utricles are encrusted by irregular thickets of radially arranged cyanobacterial filaments (Fig. 7R). In contrast, these cyanobacterial coatings are absent on porocharacean remains.
Monospecific porocharacean assemblages are commonly associated with brackish conditions in the Lower Cretaceous record (see facies pL) and this may also be the case here. The coexistence of clavatoroid charophyte remains encrusted by cyanobacterial coats with non-coated porocharacean remains suggests some type of ecological separation between the two groups. The growth of cyanobacterial crusts around charophytes has been associated in present and fossil lakes with rapidly changing environmental conditions, leading to the demise of the original charophyte meadows and their substitution by cyanobacteria (e.g., eutrophication in the case documented by Martín-Closas 1999). In facies pccL, the absence of cyanobacterial crusts around porocharacean remains—these charophytes being tolerant to brackish water—suggests that the clavatoroids may correspond to an early condition of the lake, when freshwater conditions prevailed, before it suddenly shifted to more brackish conditions, when porocharacean thrived. Between the two phases cyanobacteria grew on dying clavatoroidean plants. Evidence of episodical marine influence is supported by benthic foraminifera and dasycladaleans (Fig. 6F, Table 2).
6) Oncoid-rich limestones (oL) are not common facies in the studied section. These limestones are characterized by an abundance of > 1 cm large, irregular oncoids (Fig. 6G), and few short portions of charophyte thalli (Clavatoraxis sp., Favargerella sp., and M. grambastii). The oncoids are formed of laminae, whose thickness varies laterally from 10 to 500 µm, and show filaments with cyanobacterial affinity.
The growth of similar oncoid structures commonly occurs in low-energy conditions (e.g., Schäfer and Stapf 1978; Lanés and Palma 1998) in lacustrine slope areas (e.g., Tucker and Wright 1990; Platt and Wright 1991). The fragments of charophyte thalli are, thus, interpreted to have been laterally transported from shallower neighboring areas, where the main charophyte meadows grew.
Parasequences and palaeoecology of charophyte assemblages
Two distinct types of parasequence have been distinguished within the Clavatoracean-rich limestones and marls from the Cantaperdius Formation:
Parasequence 1 (Ps1). This parasequence is formed of a succession of the following facies, from base to top: clavatoracean-rich marls (cM), oncoid-rich limestones (oL), limestones rich in clavatoracean utricles and thalli (cL), and Munieria grambastii-rich limestones (mL) (Fig. 8). The top of the parasequence is characterized by the occurrence of rootmarks and intraclasts attributed to palustrine features (c.f. Alonso-Zarza and Wright 2010). The clavatoracean-rich marls (cM) and oncoid-rich limestones (oL) are locally absent from this type of parasequence. In such cases, the succession of facies is formed solely by limestone facies cL and mL (Fig. 4A). The succession of facies is interpreted as a shallowing-upwards parasequence, formed in shallow, alkaline lakes and ponds (e.g., Murphy and Wilkinson 1980; Strasser 1988; Platt and Wright 1991; Meléndez et al. 2009). This parasequence is dominated by limestone facies in the stratigraphic intervals between 11–100 m and 187–274 m (Figs. 4A, 5B). Meanwhile, marlier parasequences dominated by facies cM occur in the intervals 100–187 m and 274–366 m of the stratigraphic section (Figs. 4A, B, 5C), and indicate higher terrigenous inputs within the lacustrine system.

Parasequences described in the Barremian of the Herbers-Mas de Petxí section
From a palaeoecological point of view, the clavatoracean-rich marls (cM) and oncoid-rich limestones (oL) represent deeper and relatively less-vegetated parts of the lake, whereas the limestones rich in clavatoracean utricles and thalli (cL) would correspond to sublittoral areas of the lake. In such facies, charophyte meadows reached the highest diversity and were formed by Atopochara and clavatoroids, including Ascidiella. Finally, the lake margin would have been populated by poorly diverse charophyte assemblages dominated by the thallus M. grambastii, probably belonging in most cases to Clavator plants.
Parasequence 2 (Ps2). This parasequence is formed of a succession of clavatoracean-rich marls (cM), limestones rich in clavatoracean utricles and thalli (cL), limestones with porocharaceans and coated clavatoraceans (pccL), and limestones with porocharaceans (pL) at the top (Fig. 8). Parasequence Ps2 is locally found interbedded with Ps1 (Fig. 4A).
From a palaeoecological viewpoint, the lower part of the parasequence is similar to Ps1. However, the upper part differs substantially since it is characterized by facies rich in Porochara, which suggests that brackish conditions prevailed. In sum, this parasequence records a vertical increase in marine influence. In some cases, the change from a freshwater lake to a brackish environment is captured in facies pccL, in which waning clavatoracean communities are encrusted by cyanobacteria, whereas porocharacean remains are devoid of periphyton, representing the new, succeeding community.
Porocharacean-rich limestones and marls from the Artoles Formation
Description of facies and their charophyte assemblages
1) Marls rich in porocharaceans and benthic foraminifera (pfM) mainly contain abundant Porochara maestratica gyrogonites (Fig. 3A, B), ostracod carapaces and benthic foraminifera (Table 2). The preservation of the gyrogonites is good to medium (slightly eroded), with rare occurrences of broken specimens. Charophyte thalli were not found. These features suggest moderate transport from shallower neighboring lacustrine belts. Similar assemblages, solely of gyrogonites, were regarded as parautochthonous (e.g., Vicente et al. 2015). Other, less-abundant charophyte species, i.e., Echinochara lazarii, Atopochara trivolvis var. triquetra, Globator maillardii var. trochiliscoides, Ascidiella stellata var. lata, and A. triquetra (Fig. 3; Tables 3, 4; Supplementary materials 2–3), are superficially eroded or broken and are interpreted as having been deposited after longer transport.
This facies is interpreted as having been developed under low hydrodynamic conditions in a marginal coastal area. The presence of fine siliciclastics within the depositional setting is indicative of continental input. Euryhaline conditions are suggested by the coexistence of brackish (i.e., Porochara maestratica) and marine fossils (benthic foraminifera and dasycladales; Table 2).
2) Limestones with porocharaceans and benthic foraminifera (pfL) are characterized by the occurrence of abundant porocharacean gyrogonites, attributed to the genus Porochara and benthic foraminifera (Fig. 9A–D). Locally, peloids may be abundant (Fig. 9B, C). Most of the porocharacean gyrogonites are well-preserved (Fig. 9D–F) but a few are broken (Fig. 9F, G). Since no thalli were found in association, the charophyte assemblage was interpreted as parautochthonous.

Microfacies and components of Porocharacean-rich limestones and marls from the Artoles Formation. A Wackestone facies pfL (sample H59; thin section no. 87458 MGSCB); B, C Packstone facies pfL showing abundant benthic biseriate foraminifera (f) and miliolids (m), porocharacean gyrogonites (p), and peloids (pl) (sample H123; thin section no. 87460 MGSCB); D Detail of (A) showing a porocharacean gyrogonite and a biseriate foraminifer (sample H59; thin section no. 87458 MGSCB); E Well-preserved porocharacean gyrogonite partially filled by micrite entering from the apical pore (sample H59; thin section no. 87456 MGSCB); F Well-preserved porocharacean gyrogonites associated with small fragments of similar gyrogonites (arrow) (sample H59; thin section no. 87457 MGSCB); G Broken porocharacean gyrogonite (sample H59; thin section no. 87457 MGSCB). Scale bar A–C equals 500 µm; Scale bar D–G equals 200 µm
Facies pfL has been interpreted as formed in coastal brackish marshes subjected to wave action. Facies with similar taxonomic and taphonomic features have been documented by Climent-Domènech et al. (2009) and Bover-Arnal and Strasser (2013).
3) Limestones with molluscs and benthic foraminifera (mfL) mainly contain well-preserved, agglutinated foraminifera, miliolids, gastropods and bivalves, but rare oyster shells are broken (Fig. 4B). This facies has been interpreted as being deposited in low-energy areas in a coastal marine setting where small benthic foraminifera and molluscs thrived (see, e.g., Colombié and Strasser 2005 for comparison with a similar setting).
Parasequences and palaeoecology of charophyte assemblages
The repeated superposition of the facies described above leads to two parasequence types (Fig. 8):
Parasequence 3 (Ps3). This parasequence is formed of two facies, with marls rich in porocharacean and benthic foraminifera (facies pfM) at the base and limestones with porocharaceans and benthic foraminifera (facies pfL) at the top (Fig. 8). The facies succession reflects an upwards increase in energy and marine influence, passing from a brackish setting with porocharaceans to marginal-marine conditions relatively rich in benthic foraminifera. The absence of palustrine features, such as rootlet marks, at the top of the parasequence indicates permanent subaquatic conditions (e.g., Osleger 1991).
Parasequence 4 (Ps4). This parasequence consists of a succession formed by limestones with molluscs and benthic foraminifera (facies mfL) at the base, followed by limestones with porocharacean gyrogonites and benthic foraminifera (facies pfL) at the top (Fig. 8). The facies succession reflects a transition from low-energy, shallow-marine settings, where matrix-supported textures and exclusively marine components prevail, to higher-energy brackish to euryhaline environments, where grain-supported textures were generated and porocharacean charophytes occurred. Similar parasequences have been associated with shallowing-upwards marginal-marine successions (e.g., Bádenas et al. 2004; Védrine and Strasser 2009), which in the case studied would include a shift towards brackish conditions.
Echinochara-rich marls and limestones from the Artoles Formation
Description of facies and their charophyte assemblages
1) Echinochara lazarii-rich grey marls (elGM) are characterized by the occurrence of abundant utricles of E. lazarii (Fig. 3D), ostracods and well-preserved agglutinated foraminifera. E. lazarii is excellently preserved, showing the two layers of the utricle structure, and fertile whorls in anatomical connection. In addition, calcified portions of the corresponding thalli, Charaxis spicatus, are found in anatomical connection with these utricles (Fig. 3W). All these features indicate that this species is autochthonous in the depositional setting. Other clavatoraceans such as G. maillardii var. biutricularis, A. trivolvis var. triquetra, Ascidiella cruciata and Pseudoglobator paucibracteatus are rare, and superficially eroded or even broken (Table 5; Supplementary materials 2–3), suggesting parautochthony or allochthony. The ostracod carapaces surveyed usually present anatomically attached valves, which indicates autochthony.
Facies elGM is interpreted as having been deposited in low-energy marginal-marine environments with significant fine siliciclastic input and populated by charophyte meadows of Echinochara lazarii. The co-occurrence of these charophytes and marine organisms (Table 2) suggests brackish to euryhaline conditions.
2) Echinochara lazarii-rich red clays (elRC) contain a micropalaeontological assemblage largely equivalent to that of elGM (Table 2). However, with the exception of E. lazarii and C. spicatus charophytes are generally more fragmented and/or superficially eroded than in the previous facies. Following the aforementioned taphonomic features, only the utricles and thalli of E. lazarii are interpreted as autochthonous.
Facies elRC is interpreted as having been deposited in brackish to euryhaline marginal-marine settings with a higher siliciclastic input than elGM, and populated by E. lazarii meadows. Subsequently these settings were exposed to oxidization under atmospheric conditions (e.g., Kraus and Aslan 1993).
3) Marls with molluscs and echinoids (meM) consist of up to 14 m thick intervals that contain abundant macrofossils (Fig. 4C) such as oysters, internal casts of infaunal bivalves and irregular echinoids (Heteraster sp.). All fossils are well preserved, suggesting autochthony.
Marls with a similar fossil assemblage have been associated with deposition in low-energy, shallow-marine areas with abundant terrigenous inputs (e.g., Védrine and Strasser 2009) as it must be the case here.
4) Limestones with rare clavatoracean thalli (tL) contain small portions of charophyte thalli belonging to Favargerella sp., Clavatoraxis sp. and Charaxis sp, and rare clavatoracean utricles and porocharacean gyrogonites (Table 2). The limestone beds of facies tL are locally capped by a ferruginous crust.
This facies is related to deposition in shallow euryhaline ponds and lakes. Low abundance and erosion features suggest that most charophyte remains are parautochthonous in this facies, whereas the association of freshwater, brackish and marine taxa suggest that euryhaline conditions prevailed.
5) Limestones rich in ostracods and charophytes (ocL) may contain intraclasts and are mainly composed of abundant well-preserved ostracod carapaces (generally non-cypridean and less-abundant cypridean species; Fig. 10B, C), and charophyte remains. Less abundant are gastropods, foraminifera, and dasycladaleans. The charophyte assemblage mainly consists of Charaxis spicatus thalli (Fig. 10D), rare and broken Clavatoraxis sp. thalli, utricles of Echinochara sp., and clavatoroid utricles (Fig. 10E, F).

Microfacies and components of Echinochara-rich marls and limestones from the Artoles Formation. A Facies tL (sample H61B; thin section no 87459 MGSCB), scale bar equals 1000 µm; B Facies ocL containing abundant carapaces of cypridean and non-cypridean ostracodes (cy), and charophyte thalli (sample H127; thin section no. 87461), scale bar equals 1000 µm; C Non-cypridean ostracod carapace, facies ocL (sample H127; thin section no. 87461 MGSCB), scale bar equals 250 µm; D Node with adjacent internodes of Charaxis spicatus, facies ocL (sample H127; thin section no. 87461), scale bar equals 250 µm; E Clavatoroid utricle, facies ocL (sample H125; thin section no. 87462 MGSCB), scale bar equals 250 µm; F Clavatoroid utricle, facies ocL (sample H125; thin section no. 87462 MGSCB) scale bar equals 250 µm; G Facies bgL showing abundant bivalve shells (sample H134; thin section no. 87465 MGSCB), scale bar equals 1000 µm; H Complete gastropod, facies bgL (sample H130; thin section no 87463), scale bar equals 250 µm; I Tangential section of a dasycladalean thallus, facies bgL (sample H71; thin section no. 87468 MGSCB), scale bar equals 250 µm; J Intraclast containing charophyte thalli (sample H133; thin section no. 87464), scale bar equals 1000 µm; K Intraclasts with small portions of charophyte thalli, arrow indicates component enlarged in (L) (sample H135; thin section no. 87466 MGSCB), scale bar equals 250 µm; L Transversal section of a charophyte thallus, (sample H135; thin section no. 87466 MGSCB), scale bar equals 250 µm; M Facies fcyL, showing the irregular base of the laminae (storm events) including bioclasts such as bivalve shells (b), and miliolids (m) (sample H68; thin section no. 87467 MGSCB), scale equals 1000 µm; N Detail of the laminae showing micritized benthic foraminifera (f) and fragments of echinoid (e) (sample H68; thin section no. 87467 MGSCB), scale bar equals 250 µm
Facies ocL is interpreted as having been deposited in coastal, shallow lakes and ponds, where E. lazarii thrived. The co-occurrence of charophytes, cypridean and non-cypridean ostracods and marine organisms (Table 2) indicates both a freshwater and marine influence. Intraclasts have been related with the wave reworking of the already compacted mud in shoreline lacustrine areas (see, e.g., Freytet and Plaziat 1982).
6) Finely laminated limestones with benthic foraminifera and cyanobacterial filaments (fcyL) are formed by an alternation of bioclastic laminae and thicker micrite horizons (Table 2). Skeletal components such as benthic foraminifera and fragments of bivalves and echinoids occur in the bioclastic laminae (Fig. 10M, N). The micritic horizons show relicts of calcified microbial filaments (10–20 µm across), rare miliolids, and agglutinated bi- or tri- seriate foraminifera.
Facies fcyL is interpreted as microbial mats formed in an intertidal to supratidal setting (see, e.g., Shinn et al. 1969; Pérez-Valera and Pérez-López 2008; Bover-Arnal and Strasser 2013). Thin bioclastic horizons have similar sedimentary and palaeontological features to the storm-event deposits described within microbial mats (see, e.g., Bover-Arnal and Strasser 2013) and it must be origin of the laminae observed herein as well.
7) Limestones rich in bivalves and gastropods (bgL) are lenticular, cross-bedded limestone beds that contain abundant molluscs (oysters, other bivalves and gastropods; Fig. 10G, H) associated with other marine fossils (Fig. 10I; Table 2). Fossils are usually broken, suggesting that they were transported in the depositional setting. Intraclasts mainly occur at the base of the beds, forming a lag deposit (Fig. 4C). Some of the intraclasts contain portions of charophyte thalli (Fig. 10J–L).
The sedimentary features (Table 2), and the presence of marine fossils indicate that this facies corresponds to channel-fill deposits formed in a marine coastal area. Similar deposits have been associated with intertidal to shallow subtidal areas (e.g., Colombié and Strasser 2005; Védrine and Strasser 2009; Bover-Arnal and Strasser 2013) and it must be the case in this deposit as well.
Parasequences and palaeoecology of charophyte assemblages
The previous facies can be arranged into three distinct parasequences.
Parasequence 5 (Ps5). This was identified at the base of the Artoles Formation (Fig. 4B). It is characterized by the succession of Echinochara lazarii-rich grey marls (facies elGM) and limestones with rare clavatoracean thalli (facies tL) at top (Fig. 8). The top of this parasequence is commonly marked by a ferruginous crust. The parasequence is interpreted as a shallowing-upwards succession, changing from a relatively deeper setting to shallower ponds and lakes, both under brackish to euryhaline conditions. The absence of emergence features such as mudcracks and the presence of ferruginous crusts at the top suggest the persistence of subaquatic conditions throughout the parasequence (e.g., Osleger 1991). From the viewpoint of charophyte palaeoecology, the facies succession is of significance given that it documents the development of charophyte meadows dominated by E. lazarii in brackish settings.
Parasequence 6 (Ps6). This parasequence is characteristic of the upper part of the Artoles Formation including an intercalation of the Morella Formation (meters 450–610 of the section studied; Fig. 4C). It is usually composed of a succession of Echinochara lazarii-rich grey marls (elGM) and E. lazarii-rich red clays (elRC). However, the facies elGM is locally overlain by limestones rich in ostracods and charophytes (ocL).
The succession of lacustrine grey marls to marginal lacustrine limestones and finally to red clays is associated with a shallowing-upwards sequence formed in ponds developed in floodplains and mudflats. The shallowing-upwards trend culminated in sub-aerial exposure and sediment oxidation (e.g., Freytet and Plaziat 1982; Breda and Preto 2011). This parasequence also indicates that E. lazarii was able to thrive in temporary ponds exposed to periodic sub-aerial conditions (facies elRC), regardless of the amount of clastic input received.
Parasequence 7 (Ps7). This parasequence is intercalated with the previous one between meters 450 and 610 of the stratigraphic section (Fig. 4C). It is mainly formed of the succession of marls with molluscs and echinoids (facies meM) followed by channelized limestones rich in bivalves and gastropods (facies bgL) (Fig. 8), which are locally topped by finely laminated limestones with benthic foraminifera and cyanobacterial filaments (facies fcyL). This parasequence is interpreted as having been formed in a tidal flat to shallow-marine area under normal-marine salinity conditions (facies meM) with tidal channels (facies bgL). Abandoned channels would have locally evolved into restricted, poorly oxigenated ponds, where microbial mats developed (facies fcyL).
Clavatoracean-rich clays from the Morella Formation
Description of facies and their charophyte assemblages
1) Clavatoracean-rich grey clays (cGC) mainly contain abundant and diverse clavatoraceans, ostracods and agglutinated foraminifera (Table 2). The charophyte assemblage is mainly composed of well-preserved utricles of E. lazarii with abundant well-preserved utricles of A. trivolvis var. triquetra and C. harrisii var. reyi, and var. harrisii (Table 5; Supplementary materials 2–3), suggesting autochthony. Small populations (usually < 20 utricles; Table 5; Supplementary materials 2–3) of well-preserved or slightly abraded utricles of Globator maillardii var. trochiliscoides and var. biutricularis (Fig. 3E–F), Clavator grovesii var. jiuquanensis (Fig. 3I), A. cruciata (Fig. 3R), and P. paucibracteatus (Fig. 3V) and gyrogonites of aff. Mesochara harrisii (Fig. 3C) are considered parautochthonous within the assemblage. H. neimongolensis var. neimongolensis is extremely rare in facies cGC and the utricles are highly eroded indicating that they represent allochthonous components. Abundant, well-preserved benthic foraminifera, and few echinoid fragments also occur.
This facies is interpreted as having been deposited in permanent lakes and ponds located near the marine shoreline, with abundant terrigenous influence, where charophyte meadows dominated by E. lazarii, A. trivolvis var. triquetra, and C. harrisii var. harrisii and var. reyi occurred. The presence of abundant, well-preserved agglutinated foraminifera together with charophytes suggests euryhaline conditions with alternating periods of freshwater, brackish and/or marine influence.
2) Red clays with clavatoraceans (cRC) mainly contain charophyte remains, ostracods and some benthic foraminifera. The charophyte assemblage is mainly composed of abundant, well-preserved utricles of C. harrisii var. reyi (Fig. 3L), which may be associated with a few, locally abraded utricles of E. lazarii, A. trivolvis var. triquetra, H. neimongolensis var. neimongolensis, C. grovesii var. jiuquanensis, and P. paucibracteatus, and gyrogonites of aff. M. harrisii (Fig. 3; Table 5; Supplementary materials 2–3). Thalli are rare, and mainly belong to small portions of Clavatoraxis sp. (Fig. 3Y). As regards utricles, C. harrisii var. reyi is interpreted as autochthonous, whereas the other species are considered as parautochthonous components, i.e., transported from neighboring areas, where they probably grew.
Red clays are interpreted as tidal flat to fluvial floodplain deposits oxidized under sub-aerial conditions (see, e.g., Kraus and Aslan 1993; Abdul-Aziz et al. 2003; Vicente et al. 2016). The occurrence of abundant charophyte remains associated with benthic foraminifera indicates sedimentation in temporary coastal lakes and ponds exposed to euryhaline conditions. The presence of rootmarks and mottled features suggests the development of hydromorphic soils within a floodplain setting (e.g., Kraus and Aslan 1993).
3) Limestones with clavatoracean thalli and utricles (ctuL) contain abundant charophyte remains, thalli portions and utricles (Supplementary material 5). The thalli are usually encrusted by an up to 400 µm thick cyanobacterial coat (Supplementary material 5, Fig. G–H). Long portions of the thallus C. spicatus (Fig. 11C, E) and the corresponding E. lazarii utricles are common (Fig. 11D, E; Supplementary material 5, Fig. F–H). Both plant organs (C. spicatus and E. lazarii) are locally found in anatomical connection (Fig. 11E), indicating that the species bearing them was autochthonous in this setting. In some cases, E. lazarii-C. spicatus may form monospecific assemblages (Fig. 11B). Other well-preserved charophyte thalli are Clavatoraxis robustus (Fig. 11G–I) and Favargerella sp. The thallus Clavatoraxis sp. can also form monospecific assemblages in these limestones (Fig. 11F). Clavatoroid utricles are very abundant but their taxonomic determination from thin sections was not generally possible (Fig. 11J–L; Supplementary material 5, Fig. A–C, E). An exception is Ascidiella sp. that was recognized on the basis of its 3 bract cells and the phylloid arranged around the gyrogonite in transversal sections (Fig. 11M, Supplementary material 5, Fig. C). Atopocharoid utricles belonging to Atopochara sp. (Fig. 11N) are common. These charophytes are associated with ostracods, and locally also with benthic foraminifera and dasycladales.

Microfacies and components of Clavatoracean-rich clays and limestones from the Morella Formation. A Facies ctuL showing the co-occurrence of Clavatoraxis sp. (cl), Charaxis sp. (ch) and a Globator sp. utricle (u) (sample H77-79; thin section no. 87469 MGSCB); B Another view of facies ctuL dominated by Charaxis spicatus (sample H90b; thin section no. 87477 MGSCB); C Transversal section of C. spicatus internode (sample H90; thin section no. 87473 MGSCB); D Complete fertile whorl of Echinochara (sample H90; thin section no. 87475 MGSCB); E Charaxis spicatus anatomically connected to Echinochara (sample H90b; thin section no. 85676 MGSCB); F Facies ctuL, showing predominant Clavatoraxis sp. thalli (sample H87; thin section no. 87472); G Transversal section of Clavatoraxis sp (sample H90; thin section no. 87473); H Longitudinal section of Clavatoraxis sp. and a clavatoroid utricle (sample H87; thin section no. 87471 MGSCB); I Transversal section of Clavatoraxis sp. with poorly calcified cells (sample H90; thin section no. 87475 MGSCB); J Clavatoroid utricle showing the apical neck (arrow) (sample H90; thin section no. 87476 MGSCB); K Clavatoroid utricle with a thick nodular layer (sample H87; thin section no. 87471 MGSCB); L Transversal section of a clavatoroid utricle (sample H90; thin section no. 87474 MGSCB); M Ascidiella sp. utricle (sample H77-79; thin section no. 87469 MGSCB); N Atopochara sp. utricle (sample H78; thin section no. 87470 MGSCB); Scale bar A, B, F equals 1000 µm; C–E, G–N equals 250 µm
The deposits in this facies are interpreted as being the result of sedimentation in alkaline lakes (e.g., Freytet and Plaziat 1982; Bádenas et al. 2018), where diverse clavatoroid and atopocharoid charophyte meadows thrived, locally dominated by E. lazarii.
4) Limestones with bivalves and gastropods (bgL). This facies has similar features to those in the Echinochara-rich marls and limestones of the Artoles Formation.
Parasequences and palaeoecology of charophyte assemblages
Parasequence 8 (Ps8). The repetition of this basic accretional unit mainly gives rise to the base of the Morella Formation (Fig. 4C). It is formed by the succession of (1) limestones with clavatoracean thalli and utricles (ctuL), (2) clavatoracean-rich grey clays (cGC) and (3) red clays with clavatoraceans (facies cRC) (Fig. 8). It is interpreted as a shallowing-upward succession recording a depositional change from sedimentation in an alkaline lake to ponds with significant clastic input, finally leading to temporary ponds subjected to sub-aerial exposure (e.g., Freytet and Plaziat 1982; Abdul-Aziz et al. 2003; Abels et al. 2009; Breda and Preto 2011; Bádenas et al. 2018). Euryhaline conditions prevailed throughout the parasequence. Regarding the charophyte palaeoecology, the shallowing-upwards process recorded a charophyte species shift from a diverse assemblage including E. lazarii, Atopochara trivolvis var. triquetra and var. trivolvis, and C. harrisii var. harrisii and var. reyi, which developed in both alkaline and clastic-influenced lakes, to a poorly diverse assemblage containing only abundant C. harrisii, developed in floodplain ponds.
A variation of this parasequence occurs at the boundary between the Morella Formation and the overlying marine facies of the Cervera Formation (Fig. 4C), where only the two end types occur intercalated with tidal channels formed by limestones rich in bivalves and gastropods (facies bgL).
Discussion
Palaeoenvironmental evolution
The Herbers-Mas de Petxí section contains a thick and continuous sedimentary record, which is representative of the vertical and lateral evolution of the Barremian depositional environments established on the north-western margin of the Morella Sub-Basin in the northern Maestrat Basin. Based on the facies and parasequences recognized, and on the taphonomy and palaeoecology of their charophyte assemblages (Fig. 4A–C), three different stages of palaeoenvironmental evolution can be interpreted (Figs. 12, 13, 14).

Palaeoenvironmental reconstruction of the first depositional stage recorded in the Herbers-Mas de Petxí section, corresponding to carbonate-dominated wetland environments of the Cantaperdius Formation. A Palaeoecological model including the parasequences and charophyte distribution interpreted and discussed in the text for the Cantaperdius Formation. B Google Earth photograph of wetlands in the Everglades National Park (Florida, United States of America), which are considered a modern analogue of (A)
(1) The first stage is mainly represented by the early Barremian clavatoracean-rich limestones and marls of the Cantaperdius Formation. It is characterized by the inter-bedding of parasequences Ps1 and Ps2, which are laterally equivalent to one another, forming a patchwork of freshwater to brackish lakes and ponds (Fig. 12A). Parasequence Ps1 was the most common type of succession that developed in these alkaline palaeoenvironments, whereas Ps2 represents freshwater lakes that received marine influence and were thus located closer to the coast. Similar freshwater alkaline environments receiving intermittent or sporadic marine influence have been reported in other ancient coastal wetlands (e.g., Armenteros and Edwards 2012; Suárez-González et al. 2015) and in extant coastal wetlands, such as the Everglades National Park (Florida) (e.g., Platt and Wright 1992; Marshall et al. 2009; Fig. 12B).
Charophyte assemblages in these wetlands are distributed according to depth, alkalinity, salinity and clastic influence (Fig. 12A). Charophyte floras in freshwater alkaline lakes with low clastic inputs were composed of Atopochara trivolvis var. triquetra, Clavator harrisii var. harrisii, Ascidiella stellata var. stellata and var. lata, and A. triquetra. Locally, Hemiclavator-dominated populations (H. adnatus and H. neimongolensis var. posticecaptus) occur. These charophyte assemblages would have grown in relatively deep areas of the lake, passing to almost monospecific assemblages of Munieria grambastii at the lake margins (Fig. 12A). These types of lake are better represented in the limestone-dominated intervals of the lower part of the Cantaperdius Formation (between meters 0 and 100 of the section in Fig. 4A). Meanwhile, under higher clastic inputs (mainly in marly intervals between meters 100 and 187 of the section; Fig. 4A), previous charophyte assemblages were rich in Globator maillardii var. trochiliscoides. Otherwise, brackish floras from this interval were mainly composed of Porochara maestratica and occasionally, E. lazarii.
(2) The second palaeoenvironmental stage is late early Barremian in age and corresponds to the upper part of the Cantaperdius Formation and the lower part of the Artoles Formation (Fig. 4B). It is formed by three depositional belts ranging from lacustrine to shallow-marine facies (Fig. 13A). The most proximal belt is formed by freshwater lakes and ponds of Ps1, showing a dominance of clavatoracean-rich assemblages with E. lazarii, A. trivolvis var. triquetra, G. maillardii var. trochiliscoides C. harrisii var. harrisii, C. calcitrapus var. jiangluoensis and var. calcitrapus, and Hemiclavator neimongolensis var. neimongolensis, changing to Munieria-rich meadows at the lake margins. The second depositional belt was formed by supratidal brackish lakes and ponds (Ps3 and Ps5) interspersed with tidal channels, represented by Ps6. Echinochara lazarii inhabited the deeper areas of the lakes, whereas other clavatoraceans predominated at the lake margins (Ps5). The third depositional belt included coastal brackish lakes, exclusively colonized by porocharacean-rich meadows (Ps3) that changed seawards to benthic foraminifera and molluscs in shallow-marine conditions (Ps6). Based on the facies descriptions, their lateral relationships and the fossil organisms identified, the more heavily clastic-influenced areas of the Everglades National Park would constitute a modern analogue of the depositional environments recorded in the upper part of the Cantaperdius Formation and the lower part of the Artoles Formation (Marshall et al. 2009) (Fig. 13B).

Palaeoenvironmental reconstruction of the second depositional stage recognized in the Herbers-Mas de Petxí section, corresponding to wetland environments with higher clastic influence of the upper part of the Cantaperdius Formation and lower part of the Artoles Formation. A Palaeoecological model including the parasequences and charophyte distribution interpreted and discussed in the text; B Google Earth photograph of more terrigenous-influenced wetlands in the Everglades National Park (Florida, United States of America), which are considered a modern analogue of A
In comparison with the previous stage, the wider development of brackish facies in this second depositional stage was related to the early phase of the marine transgression of late early Barremian age recorded in the Maestrat Basin (e.g., Bover-Arnal et al. 2016). This transgressive pulse is best characterized in the more depocentral areas in the Maestrat Basin such as in the Salzedella Sub-Basin (Fig. 1B), where the Artoles Formation is fully marine (Salas et al. 2001). In the section studied, the increase in clastic content in the sedimentary system during this second depositional stage is interpreted to indicate enhanced rainfall and clastic run-off (see e.g., Pasquier and Strasser 1997; Strasser and Védrine 2009) during the late early Barremian in the northern Maestrat Basin.
(3) The third depositional stage was reconstructed based on the lateral and upwards change from the Echinochara-rich marls and limestones of the upper Artoles Formation to the clavatoracean-rich clays and marls of the Morella Formation (Fig. 4C), which recorded a regression of early late Barremian age. Three palaeoenvironmental belts were distinguished (Fig. 14A). The most distal belt is represented by shallow-marine subtidal areas with abundant infaunal bivalves and irregular echinoids, and tidal channels (Ps7). The latter may become abandoned and develop into restricted and microbially dominated ponds. This belt graded landwards to a second depositional belt formed by brackish supratidal lakes and ponds. These were rich in Echinochara lazarii and passed laterally either to lakeshore areas or to exposed, oxidized flood plain areas (Ps6; Fig. 14A). Finally, the most proximal depositional belt corresponds to floodplains with freshwater alkaline lakes populated by Echinochara sp., Atopochara sp., Ascidiella sp., and other clavatoroidean charophytes, and clastic ponds where Echinochara lazarii, Atopochara trivolvis, and Clavator harrisii thrived. These lakes contained more diverse charophyte assemblages than the brackish belt (Ps6). In some areas, small tidal channels drained the floodplain. Modern analogue environments with similar facies and sedimentary features are found in non-marine areas around Buckingham Bay in Northern Australia (Fig. 14B) (Short 2020).

Palaeoenvironmental reconstruction of the third depositional stage of recorded in the Herbers-Mas de Petxí section, corresponding to wetland environments of the upper Artoles and Morella formations. A Palaeoecological model including the parasequences and charophyte distribution interpreted and discussed in the text; B Google Earth photograph of a clastic-dominated tidal flat in Buckingham Bay (N Australia), which is regarded as a modern analogue of (A)
In this last depositional stage, the sedimentary system shows an increase in clay content (Fig. 14A), suggesting greater terrigenous run-off than in the previous stage (Fig. 13A). Evaporitic and dolomitic beds, which are very frequent in arid or strongly seasonal tidal flats (e.g., Breda and Preto 2011; Bádenas et al. 2018), are absent, indicating that humid conditions prevailed in the Maestrat Basin during the early late Barremian. In a more general context, humid conditions have been reported in the Tethyan and Boreal domains during the mid Barremian (= late early Barremian and early late Barremian) (e.g., Ruffell and Batten 1990; Godet et al. 2008; Mutterlose et al. 2014).
Charophyte palaeoecology
The palaeonvironmental reconstruction proposed in the previous section allows a precise characterization of the palaeoecological constraints of the most representative charophyte species from the Barremian of Iberia. These species were mainly distributed according to lake bathymetry, salinity (ranging from freshwater to brackish or euryhaline), and clastic inputs into the lakes.
Charophytes from freshwater lakes with different degrees of clastic influence
Freshwater settings contain the richest and most diverse charophyte assemblages, with most species living in shallow water, whereas only one species usually prevailed in the shallowest margins (Munieria grambastii, probably representing the thalli of some Clavator species). Only two species, Atopochara trivolvis var. triquetra and Clavator harrisii, were capable of growing in these Barremian lakes regardless of the amount of clastic input received. Another species, Echinochara lazarii, was limited during the early Barremian to clastic-influenced lakes, whereas later, by the late Barremian, it had colonized freshwater settings with low clastic input, such as the alkaline lakes recorded in the Morella Formation, indicating that its palaeoecological requirements expanded over time. In this regard, Charaxis spicatus, which represents the thallus of E. lazarii, was first described in freshwater lacustrine limestones from Las Hoyas in the south-western part of the Iberian Chain (Martín-Closas and Diéguez 1998) where it occurs attached to non-calcified gyrogonites, latter attributed to E. lazarii (Pérez-Cano et al. 2020).
Charophytes that preferred freshwater lakes with low clastic inputs, include two species, Ascidiella stellata and A. triquetra, which are more abundant in the lower part of the Cantaperdius Formation (Figs. 4A and 12A). The abundance of these latter species wanes as clastic content increases in the depositional systems. The palaeoenvironmental distribution of A. cruciata shows a greater tolerance to clastic influence than its ancestor, A. triquetra, since it has been found in facies of temporary alkaline lakes with low clastic influence in floodplains of the Morella Formation (Fig. 14A) and in palustrine areas with higher clastic influence in floodplains of the La Huérguina Formation in the south-western part of the Iberian Chain (Vicente and Martín-Closas 2013; Buscalioni et al. 2018).
Globator maillardii var. trochiliscoides and var. biutricularis, Clavator harrisii var. reyi, and C. calcitrapus var. jiangluoensis and var. calcitrapus were largely limited to clastic-influenced settings, either lakes or floodplains. This palaeoecological preference has already been reported in other sedimentary basins in Eurasia (e.g., Martín-Closas 2000, 2015; Vicente and Martín-Closas 2013).
In contrast to the previous species, Hemiclavator neimongolensis was able to change its palaeoecological requirements during the Barremian. In the early Barremian, the older anagenetic stage of this species, H. neimongolensis var. posticecaptus, dominated in particular charophyte assemblages in alkaline lakes with low clastic influence (Fig. 12A), whereas the younger variety, H. neimongolensis var. neimongolensis, shifted its habitat to coastal lakes with high clastic inputs. In the latter, it formed part of diverse charophyte assemblages that include Echinochara lazarii, Atopochara trivolvis var. triquetra, Clavator harrisii var. harrisii, and C. calcitrapus (Fig. 13A). In Chinese basins, H. neimongolensis var. neimongolensis has also been found in clastic-influenced lakes in a fluvial context (pers. comm. Q.F. Wang in Martín-Closas 2015). The expanding palaeoenvironmental distribution of this latter species may account for its wide biogeographic distribution across all Eurasia in the late early Barremian and late Barremian (see Martín-Closas 2015).
Charophytes from brackish lakes with different degrees of clastic influence.
The Early Cretaceous brackish charophyte assemblages have long been known to have been dominated by species of Porochara sp. (family Porocharaceae), commonly forming monospecific assemblages (e.g., Martín-Closas and Grambast-Fessard 1986; Mojon 1989; Climent-Domènech et al. 2009). In a comparison with ostracods, Schudack (1993) suggested that the clavatoracean genera Echinochara, Atopochara, Globator and Clavator may also have grown in brackish conditions. In particular, he concluded that the genera Porochara and Echinochara showed similar tolerances to high salinity (from oligohaline to brachyhaline). The data presented here show that these genera differed palaeoecologically in their tolerance to the input of clastic sediments in such brackish settings. Thus, autochthonous populations of Porochara maestratica were restricted to limestone-dominated intervals, whereas Echinochara lazarii meadows were abundant in marl-dominated intervals, suggesting that this latter species preferred settings with higher clastic influence.
The palaeoecological analysis has also shown that, as already suggested by Schudack (1993), Atopochara trivolvis var. triquetra, and var. trivolvis, and Clavator harrisii var. harrisii, and var. reyi also grew in brackish settings, but they never formed large or monospecific populations there, in contrast to E. lazarii. These differences may indicate different degrees of salinity, with the assemblages composed of E. lazarii, A. trivolvis, and C. harrisii corresponding to a lower degree of salinity and the monospecific assemblages of E. lazarii representing a higher salinity degree.
Did climatic cyclicity control the evolution of charophyte communities in the Barremian?
In the Herbers-Mas de Petxí section, at least three different types of sedimentary cycle were recognized. These are superimposed on one another and present different frequencies.
(1) The highest frequency sedimentary cycles correspond to parasequences (Ps1–Ps8). Such small-scale cycles are usually associated with Milankovitch-driven climatic variations (e.g., Strasser et al. 1999; Bádenas et al. 2004). However, the occurrence of channel deposits (Ps7, which is mainly observed in the upper part of the stratigraphic section; Fig. 4B, C) suggests that autocyclic processes, such as lateral channel migration or a sudden increase in tectonic subsidence, played a role in this type of cycle (e.g., Kindler 1992; Drummond and Wilkinson 1993).
(2) The occurrence of limestone- and clastic-dominated (either marl or clay) intervals is the second type of sedimentary cyclicity observed in the stratigraphic succession (Figs. 4A–C). Limestone- and marl-dominated intervals are formed by the stacking of a variable number of parasequences. This sedimentary cyclicity is interpreted as being related to rainfall variations in the hinterland areas, with marls or clay-dominated intervals being formed during the wettest periods (e.g., Pasquier and Strasser 1997; Védrine and Strasser 2009; Strasser and Védrine 2009). Species such as Globator maillardii var. trochiliscoides and var. biutricularis, Clavator calcitrapus, and Echinochara lazarii (the latter either in freshwater or brackish settings) mainly occurred during high rainfall periods, whereas other species such as Ascidiella stellata, A. triquetra, and P. maestratica (the last only in brackish settings) primarily occurred during the driest periods. These different palaeoecological preferences indicate that rainfall variations may have exerted a strong control over the distribution of charophyte floras.
The thickness of clastic intervals steadily increases from the base to the top of the stratigraphic section (depositional stages 2 and 3; Figs. 13A and 14A), in parallel with the establishment of more humid conditions in the Tethyan and Boreal realms during the early late Barremian (e.g., Ruffell and Batten 1990; Godet et al. 2008; Mutterlose et al. 2014). Thus, particular species restricted to environments with low clastic input, such as Ascidiella triquetra, were extinct in the late early Barremian. In contrast, species that thrived in clastic settings and those able to change or expand their palaeoecological requirements to clastic-influenced settings persisted at least until the early late Barremian. These results show that the long-term climatic changes recorded throughout the Barremian also exerted strong control over the evolution of clavatoraceans.
(3) The third type of sedimentary cyclicity, and the one that presents the lowest frequency, corresponds to the longer-term transgressive–regressive trends recorded in the stratigraphic section. Two Barremian transgressive episodes were recognized. The first transgressive cycle is of late early Barremian age and corresponds to the progradation of the marine Artoles Formation over the non-marine Cantaperdius Formation (Figs. 2 and 4B), whereas the second transgressive event is represented by the progradation of the marine Cervera Formation (not studied here) over the non-marine Morella Formation in the early late Barremian (Figs. 2 and 4C). This type of cyclicity is usually associated with large-scale eustatic changes, which are well-documented in the Barremian (e.g., Hardenbol et al. 1998; Haq 2014). In fact, these two longer-term transgressions recorded in the Morella Sub-basin had regional significance in the Maestrat Basin (Bover-Arnal et al. 2016) and seem to be correlatable with the global transgressive events that occurred between sea-level events KBa3 and KBa5 of Haq (2014). Accordingly, variations in the relative sea level favored the expansion or contraction of freshwater and brackish biotopes, and, as a consequence, they governed the development of halphobic or halophile charophyte species.
Conclusions
A palaeoecological study of the Herbers-Mas de Petxí section has provided new information about the palaeoenvironmental distribution of Barremian charophyte species in coastal wetlands. This study shows that the occurrence of most charophyte species was mainly controlled by the following interrelated factors: (1) amount of clastic supply, (2) waterbody salinity and (3) water depth. In this regard, the species used as index species of a biozone were restricted to particular environments, as in the case of Globator maillardii var. trochiliscoides, which was restricted to clastic-influenced freshwater lakes. Moreover, species used as biostratigraphic markers for the early Barremian were restricted to freshwater lakes with low clastic influence, as in the case of Ascidiella stellata var. lata and Ascidiella triquetra, or to clastic-influenced freshwater lakes, as in the case of Clavator calcitrapus. In contrast, in the late Barremian record, this distinction between the species restricted to freshwater lakes with low clastic influence and those restricted to freshwater lakes with high clastic influence becomes less clear, since the former were rare.
In the late Barremian, salinity seems to be a significant palaeoenvironmental parameter controlling the distribution of charophyte species. Only three species, Atopochara trivolvis var. triquetra, Clavator harrisii var. harrisii, and E. lazarii (the latter only during the late Barremian) were able to thrive in clastic-influenced and non-clastic freshwater settings alike. These species are also the only clavatoraceans that were able to thrive in brackish settings. However, the occurrence of monospecific populations of E. lazarii in late Barremian brackish limestones and marls indicates that this species was better able to thrive in environments with higher salinity than A. trivolvis or C. harrisii. Future research may enhance these results by comparison with other palaeobiological indicators, such as ostracods, or with geochemical data, such as stable isotopes and clay mineralogy.
Three different types of sedimentary cycle with distinct ranks were distinguished in the stratigraphic section studied. In decreasing cyclicity frequency, these are (1) the parasequences, with a metric thickness, (2) the clastic-rich and clastic-poor intervals with decametric thickness, and (3) the transgressive–regressive cycles, of hectometric thickness. The inter-bedding of limestone- and clastic-dominated intervals together with the marine/non-marine influence governed by transgressive–regressive cycles, exerted a strong control over the distribution of charophytes in the Barremian record of the Maestrat Basin. This suggests that large-scale climatic parameters determining environmental changes such as humidity, clastic run-off or changes in the relative sea level were the main drivers of charophyte diversity, distribution, and evolution during the Barremian.
References
Abdul-Aziz H, Sanz-Rubio E, Calvo JP, Hilgen FJ, Kirjgsman W (2003) Palaeoenvironmental reconstruction of a middle Miocene alluvial fan to cyclic shallow lacustrine depositional system in the Calatayud Basin (NE Spain). Sedimentology 50:211–236. https://doi.org/10.1046/j.1365-3091.2003.00544.x
Abels HA, Abdul-Aziz H, Vendra D, Hilgen FJ (2009) Orbital climate forcing in the mudflat to marginal lacustrine deposits in the Miocene Teruel Basin (Northeast Spain). J Sediment Res 70:831–847. https://doi.org/10.2110/jsr.2009.081
Alonso-Zarza AM, Wright VP (2010) Palustrine carbonates. In: Alonso-Zarza AM, Tanner LH (eds) Carbonates in continental settings: facies, environments and processes. Developments in sedimentology, vol 61. Elsevier, Amsterdam, pp 103–132. https://doi.org/10.1016/S0070-4571(09)06102-0
Armenteros I, Edwards N (2012) Palaeogeographic, palaeoclimatic, palaeohydrological and chemical/biochemical controls on accumulation of late Eocene coastal lacustrine-palustrine limestones, Southern England. Sediment Geol 281:101–118. https://doi.org/10.1016/j.sedgeo.2012.08.006
Bádenas B, Salas R, Aurell M (2004) Three orders of regional sea-level changes control facies and stacking patterns of shallow platform carbonates in the Maestrat Basin (Tithonian-Berriasian, NE Spain). Int J Earth Sci 93:144–162. https://doi.org/10.1007/s00531-003-0367-2
Bádenas B, Aurell M, Gasca JM (2018) Facies model of a mixed clastic-carbonate, wave-dominated open-coast tidal flat (Tithonian–Berriasian, north-east Spain). Sedimentol 65:1631–1666. https://doi.org/10.1111/sed.12441
Bover-Arnal T, Strasser A (2013) Relative sea-level change, climate, and sequence boundaries insights from the Kimmeridgian to Berriasian platform carbonates of Mount Salève (E France). Int J Earth Sci 102:493–515. https://doi.org/10.1007/s00531-012-0804-1
Bover-Arnal T, Moreno-Bedmar JA, Frijia G, Pascual-Cebrián E, Salas R (2016) Chronostratigraphy of the Barremian-Early Albian of the Maestrat Basin (E Iberian Peninsula): integrating strontium-isotope stratigraphy and ammonoid biostratigraphy. Newsl Stratigr 49(1):41–48. https://doi.org/10.1127/nos/2016/0072
Breda A, Preto N (2011) Anatomy of an Upper Triassic continental to marginal-marine system: the mixed siliciclastic-carbonate Travenanzes Formation (Dolomites, Northern Italy). Sedimentology 58:1613–1647. https://doi.org/10.1111/j.1365-3091.2011.01227.x
Buscalioni A, Martín-Closas C, Delvene G, Munt MC, Barral A, Tinner-Grellet G, Gomez B, Daviero-Gomez B, Chamenro B (2018) Taphonomy and palaeoecology in the upper Barremian of the SW Iberian Chain (Spain): a model to compare taxonomy and diversity of biotas from different coeval basins. Palaeogeogr Palaeoclim Palaeoecol 490:305–324. https://doi.org/10.1016/j.palaeo.2017.11.009
Canérot J, Leyva F (1976) Mapa Geológico de España, E.1:50.000 nº 520 (Peñarroya de Tanstavins). Segunda serie (MAGNA). IGME, Madrid
Canérot J, Cugny P, Pardo G, Salas R, Villena J (1982) Ibérica central-Maestrazgo. In: García A (ed) El Cretácico de España. Universidad Complutense de Madrid, Madrid, pp 273–344
Climent-Domènech H, Martín-Closas C, Salas R (2009) Charophyte-rich microfacies in the Barremian of the Eastern Iberian Chain (Spain). Facies 55:387–400. https://doi.org/10.1007/s10347-008-0173-1
Colombié C, Strasser A (2005) Facies, cycles, and controls on the evolution of a keep-up carbonate platform (Kimmeridgian, Swiss Jura). Sedimentology 52:1207–1227. https://doi.org/10.1007/s10347-008-0173-1
Combes PJ (1969) Recherches sur la genèse des bauxites dans le nord-est de l'Espagne, le Languedoc et l'Ariège (France). Mémoires du Centre d'Études et de Rech. Géol. et Hydrogéol. 3–4, 335 pp.
Drummond CN, Wilkinson BH (1993) Carbonate cycle stacking patterns and hierarchies of orbitally forced eustatic sea level change. J Sediment Petrol 63:369–377. https://doi.org/10.1306/D4267B04-2B26-11D7-8648000102C1865D
Freytet P, Plaziat JC (1982) Continental Carbonate Sedimentation and Pedogenesis–Late Cretaceous and Early Tertiary of Southern France, E. Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller), Stuttgart
Gàmez D, Paciotti P, Colombo F, Salas R (2003) La Formación Arcillas de Morella (Aptiense inferior), Cadena Ibérica oriental (España) caracterización sedimentológica. Geogaceta 34:191–194
Gasulla JM, Escaso F, Narváez I, Ortega F, Sanz JL (2015) A new sail-backed Styracosternan (Dinosauria: Ornithopoda) from the Early Cretaceous of Morella, Spain. PLoS ONE 10(12):e0144167. https://doi.org/10.1371/journal.pone.0144167
Godet A, Bodin S, Adatte T, Föllmi KB (2008) Platform-induced clay mineral fractionation along a norther Tethyan basin-platform transect: implications for the interpretation of Early Cretaceous climate change (Late Hauterivian-Early Aptian). Cret Res 29:830–847. https://doi.org/10.1016/j.cretres.2008.05.028
Grambast L (1974) Phylogeny of the Charophyta. Taxon 23:463–481. https://doi.org/10.2307/1218769
Guimerà J (2018) Structure of an intraplate fold-and-thrust belt: the Iberian Chain. A synthesis. Geol Acta 16:427–438. https://doi.org/10.1344/GeologicaActa2018.16.4.6
Haq BU (2014) Cretaceous eustasy revisited. Glob Planet Change 113:44–58. https://doi.org/10.1016/j.gloplacha.2013.12.007
Hardenbol J, Thierry J, Farley MB, Jacquin T, de Gracianski PC, Vail PR (1998) Mesozoic and Cenozoic sequence chronostratigraphic framework of European basins. In: de Gracianski PC, Hardenbol J, Jacquin T, Vail PR (eds) Mesozoic and Cenozoic Sequence Stratigraphy of European Basins, SEPM Special Publications 60, 39 pp
Kindler P (1992) Coastal response to the Holocene transgression in the Bahamas: episodic sedimentation versus continuous sea-level rise. Sediment Geol 80:319–329. https://doi.org/10.1016/0037-0738(92)90049-W
Kraus MJ, Aslan A (1993) Eocene hydromorphic paleosols: significance for interpreting ancient floodplain processes. J Sediment Petrol 63(3):453–463. https://doi.org/10.1306/D4267B22-2B26-11D7-8648000102C1865D
Lanés S, Palma R (1998) Environmental implications of oncoids and associated sediments from the Romoredo Formation (Lower Jurassic) Mendoza, Argentina. Palaeogeogr Palaeoclim Palaeoecol 140:357–366. https://doi.org/10.1016/S0031-0182(98)00037-6
Marshall FE, Wingard GL, Pitts P (2009) A simulation of historic hydrology and salinity in Everglades National Park: coupling paleoecologic assemblage data with regression models. Estuaries Coasts 32:37–53. https://doi.org/10.1007/s12237-008-9120-1
Martín-Chivelet J, López-Gómez J, Aguado R, Arias C, Arribas J, Arribas ME, Aurell M, Bádenas B, Benito MI, Bover-Arnal T, Casas-Sainz A, Castro JM, Coruña F, de Gea GA, Fornós JJ, Fregenal-Martínez M, García-Senz J, Garófano D, Gelabert B, Giménez J, González-Acebrón J, Guimerà J, Liesa CL, Mas R, Meléndez N, Molina JM, Muñoz JA, Navarrete R, Nebot M, Nieto LM, Omodeo-Salé S, Pedrera A, Peropadre C, Quijada IE, Quijano ML, Reolid M, Robador A, Rodríguez-López JP, Rodríguez-Perea A, Rosales I, Ruiz-Ortiz PA, Sàbat F, Salas R, Soria AR, Suárez-González P, Vilas L (2019) The late jurassic-early cretaceous rifting. In: Quesada C, Oliveira JT (eds) The geology of Iberia: a geodynamic approach. Volume 5: the alpine cycle. Springer, Heidelberg, pp 169–249. https://doi.org/10.1007/978-3-030-11295-0_5
Martín-Closas C (1989) Els caròfits del Cretaci inferior de les conques perifèriques del Bloc de l'Ebre. PhD thesis. Universitat de Barcelona, Barcelona, pp 608
Martín-Closas C (1999) Epiphytic overgrowth of charophyte thalli by stromatolite-like structures and fungi in the Lower Cretaceous of the Iberian Ranges. Aust J Bot 47:305–313. https://doi.org/10.1071/BT97098
Martín-Closas C (2000) Els caròfits del Juràssic superior i Cretaci inferior de la Península Ibèrica. Arxius de les Seccions de Ciències, vol 125. Institut d'Estudis Catalans, Barcelona
Martín-Closas C (2015) Cosmopolitanism in Northern Hemisphere Cretaceous Charophyta (Clavatoroidae). Palaeogeogr Palaeoclim Palaeoecol 438:9–23. https://doi.org/10.1016/j.palaeo.2015.07.029
Martín-Closas C, Diéguez C (1998) Charophytes from the Lower Cretaceous of the Iberian Ranges (Spain). Palaeontol 41:1133–1152
Martín-Closas C, Grambast-Fessard N (1986) Les charophytes du Crétacé inférieur de la région du Maestrat (Chaîne Ibérique, Catalanides, Espagne). Paléobiol Cont 15:1–66
Martín-Closas C, Salas R (1994) Lower Cretaceous Charophytes. Biostratigraphy and evolution in the Maestrat Basin (Eastern Iberian Ranges). Excursion Guidebook VIII Meeting of the European group of Charophyte Specialists. Diagonal, Barcelona
Martín-Closas C, Clavel B, Charollais J, Conrad MA (2009) Charophytes from the Barremian–lower Aptian of the Northern Subalpine Chains and Jura Mountains, France: correlation with associated marine assemblages. Cret Res 30:49–62. https://doi.org/10.1016/j.cretres.2008.05.001
Martín-Closas C, Segura-Altés R, Pérez-Cano J, Bover-Arnal T, Sanjuan J (2021) Palaeonitella trifurcata n. sp., a cortoid building charophyte from the Lower Cretaceous of Catalonia. Rev Palaeobot Palynol 295:104523. https://doi.org/10.1016/j.revpalbo.2021.104523
Meléndez MN, Liesa CL, Soria AR, Meléndez A (2009) Lacustrine system evolution during early rifting: El Castellar Formation (Galve Sub-basin, Central Iberian Chain). Sed Geol 222:64–77. https://doi.org/10.1016/j.sedgeo.2009.05.019
Mojon PO (1989) Polymorphisme écophénotypique et paléoecologie des porocharacées (Charophytes) du Cretacé Basal (Berriasien) du Jura Franco-Suisse. Rev Paléobiol 8(2):505–524
Murphy DH, Wilkinson BH (1980) Carbonate deposition and facies distribution in a central Michigan marl lake. Sedimentology 27:123–135. https://doi.org/10.1111/j.1365-3091.1980.tb01164.x
Mutterlose J, Bodin S, Fähnrich L (2014) Strontium-isotope stratigraphy of the Early Cretaceous (Valanginian–Barremian): implications for Boreal-Tethys correlation and paleoclimate. Cret Res 50:252–263. https://doi.org/10.1016/j.cretres.2014.03.027
Nebot M, Guimerà J (2018) Kinematic evolution of a fold-and-thrust belt developed during basin inversion: the Mesozoic Maestrat basin, E Iberian Chain. Geol Mag 155:630–640. https://doi.org/10.1017/S001675681600090X
Osleger D (1991) Subtidal carbonate cycles: implications for allocyclic vs. autocyclic control. Geology 19:917–920. https://doi.org/10.1130/0091-7613(1991)019%3c0917:SCCIFA%3e2.3.CO;2
Pasquier JB, Strasser A (1997) Platform-to-basin correlation by high-resolution sequence stratigraphy and cyclostratigraphy (Berriasian, Switzerland and France). Sedimentology 44:1071–1092. https://doi.org/10.1046/j.1365-3091.1997.d01-62.x
Pérez-Cano J, Bover-Arnal T, Martín-Closas C (2020) Barremian charophytes from the Maestrat Basin (Iberian Chain). Cret Res 115:104544. https://doi.org/10.1016/j.cretres.2020.104544
Pérez-Cano J, Bover-Arnal T, Martín-Closas C (2022) Barremian–early Aptian charophyte biostratigraphy revisited. Newsl Stratigr 55(2):199–230. https://doi.org/10.1127/nos/2021/0662
Pérez-Valera F, Pérez-López A (2008) Stratigraphy and sedimentology of Muschelkalk carbonate s of the Southern Iberian Continental Palaeomargin (Siles and Cehegín Fromations, Southern Spain). Facies 54:61–87. https://doi.org/10.1007/s10347-007-0125-1
Platt NH, Wright VP (1991) Lacustrine carbonates: facies models, facies distributions and hydrocarbon aspects. In Anadón P, Cabrera L, Kelts K (eds) Lacustrine Facies Analysis. International Association of Sedimentologists Special Publication 13, Blackwell, Oxford, pp 57–74
Platt NH, Wright V (1992) Palustrine carbonates and the Florida Everglades: towards and exposure index for the fresh-water environment. J Sed Petrol 62(6):1058–1071. https://doi.org/10.1306/D4267A4B-2B26-11D7-8648000102C1865D
Riveline J, Berger JP, Bilan W, Feist M, Martín-Closas C, Schudack M, Soulié-Märsche I (1996) European Mesozoic-Cenozoic charophyte biozonation. Bull Soc Géol France 167(3):453–468
Ruffell AH, Batten DJ (1990) The Barremian-Aptian arid phase in western Europe. Palaeogeogr Palaeoclim Palaeoecol 80:197–212. https://doi.org/10.1016/0031-0182(90)90132-Q
Salas R (1987) El Malm i el Cretaci inferior entre el Massís de Garraf i la Serra d’Espadà. Anàlisi de Conca. PhD Thesis, Universitat de Barcelona, Barcelona
Salas R, Guimerà J (1996) Rasgos estructurales principales de la cuenca cretácica inferior del Maestrazgo (Cordillera Ibérica oriental). Geogaceta 20:1704–1706
Salas R, Guimerà J, Mas R, Martín-Closas C, Meléndez A, Alonso A (2001) Evolution of the Mesozoic Central Iberian Rift System and its Cainozoic inversion (Iberian Chain). In: Ziegler PA, Cavazza W, Robertson AFH, Crasquin-Soleau S (eds) Peri-Tethys Memoir 6: PeriTethyan Rift/Wrench Basins and Passive Margins. Mém Mus Nat Hist Nat, Paris 186:145–185
Sanjuan J, Martín-Closas C (2012) Charophyte palaeoecology in the Upper Eocene of the Eastern Ebro Basin (Catalonia, Spain). Biostratigraphic implications. Palaeogeogr Palaeoclim Palaeoecol 365–366:247–262. https://doi.org/10.1016/j.palaeo.2012.09.031
Schäfer A, Stapf KRG (1978) Permian Saar-Nahe Basin and Recent Lake Constance (Germany): two environments of lacustrine algal carbonates. In: Matter A, Tucker ME (eds) Modern and Ancient Lake Sediments. I.A.S Special Publication 2, Blackwell, Oxford, pp 83–197. https://doi.org/10.1002/9781444303698.ch5
Schudack ME (1993) Die Charophyten im Oberjura und Unterkreide Westeuropas. Mit einer phylogenetischen Analyse der Gesamtgruppe. Berliner Geowissensch Abh (a) 8:1–209
Shinn EA, Lloyd RM, Ginsburg RN (1969) Anatomy of a modern carbonate tidal-flat, Andros Island, Bahamas. J Sed Petr 30:1202–1228. https://doi.org/10.1306/74D71DCF-2B21-11D7-8648000102C1865D
Short AD (2020) Australian Coastal Systems: Beaches, Barriers and Sediment Compartments. Coastal Research Library. 32. Switzerland: Springer. https://doi.org/10.1007/978-3-030-14294-0
Spence GH, Tucker ME (2007) A proposed integrated multi-signature model for peritidal cycles in carbonates. J Sed Res 77:797–808. https://doi.org/10.2110/jsr.2007.080
Strasser A (1988) Shallowing-upward sequences in Purbeckian peritidal carbonates (lowermost Cretaceous, Swiss and French Jura Mountains). Sedimentology 35:369–383. https://doi.org/10.1111/j.1365-3091.1988.tb00992.x
Strasser A, Védrine S (2009) Controls on facies mosaics of carbonate platforms: a case study from the Oxfordian of the Swiss Jura. IAS Spec Publ 41:199–213. https://doi.org/10.1002/9781444312065.ch13
Strasser A, Pittet B, Hillgärtner H, Pasquier JB (1999) Depositional sequences in shallow carbonate-dominated sedimentary systems: concepts for a high-resolution analysis. Sed Geol 128:201–221. https://doi.org/10.1016/S0037-0738(99)00070-6
Suárez-González P, Quijada IE, Benito MI, Mas R (2015) Sedimentology of ancient coastal wetlands: insights from a Cretaceous multifaceted depositional system. J Sed Res 85:95–117. https://doi.org/10.2110/jsr.2015.07
Trabelsi K, Soussi M, Touir J, Houla J, Abbes C, Martín-Closas C (2016) Charophyte biostratigraphy of the non-marine Lower Cretaceous in the Central Tunisian Atlas (North Africa): Palaeobiogeographic implications. Cret Res 67:66–83. https://doi.org/10.1016/j.cretres.2016.07.004
Tucker ME, Wright VP (1990) Carbonate depositional systems I: marine shallow-water and lacustrine carbonates. In: Tucker ME, Wright VP (eds) Carbonate sedimentology. Blackwell Scientific Publications, Oxford, pp 101–227
Tugend J, Manatschal G, Kusznir NJ (2015) Spatial and temporal evolution of hyperextended rift systems: Implication for the nature, kinematics, and timing of the Iberian-European plate boundary. Geology 43:15–18. https://doi.org/10.1130/G36072.1
Védrine S, Strasser A (2009) High-frequency palaeoenvironmental changes on a shallow carbonate platform during a marine transgression (Late Oxfordian, Swiss Jura Moutains). Swiss J Geosci 102:247–270. https://doi.org/10.1007/s00015-009-1326-0
Vicente A, Martín-Closas C (2013) Lower Cretaceous charophytes from the Serranía de Cuenca, Iberian chain: taxonomy, biostratigraphy and palaeoecology. Cret Res 40:227–242. https://doi.org/10.1016/j.cretres.2012.07.006
Vicente A, Martín-Closas C, Arz JA, Oms O (2015) Maastrichtian-basal Palaeocene charophyte biozonation and its calibration to the Global Polarity Time Scale in the Southern Pyrenees. Cret Res 52:268–285. https://doi.org/10.1016/j.cretres.2014.10.004
Vicente A, Expósito M, Sanjuan J, Martín-Closas C (2016) Small sized charophyte gyrogonites in the Maastrichtian of Coll de Nargó, Eastern Pyrenees: an adaptation to temporary ponds. Cret Res 57:443–456. https://doi.org/10.1016/j.cretres.2015.07.017
Yuste A, Bauluz B, Mayayo MJ (2017) Origin and geochemical evolution form ferrallitized clays to karst bauxite: an example from the Lower Cretaceous of NE Spain. Ore Geol Rev 84:67–79. https://doi.org/10.1016/j.oregeorev.2016.12.025
Acknowledgements
This study was funded by and is a contribution to the projects IBERINSULA (PID2020-113912GB-100), funded by MCIN/AEI/10.13039/501100011033 and the European Regional Development Fund (ERDF), and GEOLOGIA SEDIMENTÀRIA (2017SGR-824) of the AGAUR (Catalan Agency for Management of University and Research Grants). JP-C acknowledges the award of predoctoral grant BES-2016-076469 from the Spanish Ministry of Economy and Competitiveness. Drs. Ramon Salas and Khaled Trabelsi are acknowledged for their comments on an early draft of the manuscript. The authors appreciate the comments and suggestions given by the editor Prof. Dr. Maurice Tucker, and the reviewers Dr. Josep Sanjuan (Univ. de Barcelona), and Dr. Alba Vicente (Inst. Politécnico Nacional, La Paz, México) during the peer-review process. The English text was corrected by Michael Maudsley (Fundació Bosch i Gimpera, Universitat de Barcelona).
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Corresponding author
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pérez-Cano, J., Bover-Arnal, T. & Martín-Closas, C. Charophyte communities in Barremian Iberian wetlands. Facies 68, 13 (2022). https://doi.org/10.1007/s10347-022-00651-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10347-022-00651-6