
Abstract
The Trás-os-Montes agrarian region (TM; north-eastern Portugal) is a traditionally growing area of almond trees. However, climate change may significantly alter the current growing conditions and threaten sustainability. Chilling and forcing conditions in TM are assessed herein, also considering different varietal phenological timings. The dynamic model/chill portions (CP) and chilling hour (CH) models are used to assess the chilling phase. For the forcing phase, growing degree hours (GDH) and growing degree days (GDD) models are selected, hinting at differences between simpler (CH and GDD) and more complex models (CP and GDH). Furthermore, the climate change projections for these models are assessed. The models are computed using daily temperatures for baseline (Iberia01, 1971–2015) and two future periods (EURO-CORDEX: medium-term, 2041–2060, and long-term, 2061–2080), following two anthropogenic forcing scenarios (RCP4.5 and RCP8.5). For the future, a five-member ensemble of regional-global climate model chains is used. Projections for CP and CH show decreases, mostly for RCP8.5, whereas projections for GDH and GDD reveal an increase in heat conditions until the beginning of summer. GDH shows that potentially damaging high temperatures during summer, effectively lead to a decrease in heat accumulation for almond trees. CP and GDH, more biologically effective, seem to be more accurate in capturing climate change impacts. For the three varietal groups, the late cultivar may experience a greater reduction of CP and GDH compared to early and mid-season varieties. Nonetheless, the increase in heat forcing should be more detrimental than the decrease in chilling conditions. Although almond production may be compromised in the future, adequate adaptation options are suggested to mitigate future losses of yield/quality and warrant its sustainability.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Almond species [Prunus dulcis Mill. (D.A Webb) syn. P. amygdalus Batsch] belongs to the family Rosaceae, sub-family Prunoideae and subgenre Amygdalus (L.) (AJAP 2017; Bento et al. 2020). Almond trees are native to warm and arid regions of southwest Asia and lower mountain slope regions of central Asia (Benmoussa et al. 2017; Oliveira et al. 2018). Over the centuries, the commercial interest in almonds increased, contributing to the growth of trade routes (Cabo and Matos 2017). Therefore, the almond expansion occurred in 3 main areas: Asia, America (especially in California) and the Mediterranean region. In 2020, the worldwide almond production was 3.9 × 106 t (FAOSTAT 2022). America is the continent with the largest almond production (2.2 × 106 t), followed by Asia (6.48 × 105 t), Europe (5.61 × 105 t), Africa (2.97 × 105 t) and Oceania (2.21 × 105 t) (Cabo and Matos 2017; FAOSTAT 2022). In America, California has been the largest producing region due to its suitable pedoclimatic conditions and structured commercial organization, supporting the demands of both national and international markets (Aguiar et al. 2017; Jin et al. 2020). In some Southern European regions, the prevailing Mediterranean-type climatic characteristics, with mild rainy winters and warm and dry summers, also benefit the almond tree development (NPR 2021). Spain (4.17 × 105 t), Italy (8.1 × 104 t) and Portugal (3.2 × 104 t) are indeed the largest producers in Europe.
In 2020, Portugal ranked 15th in the world in terms of production, with an almond cultivation area of 5.2 × 104 ha (FAOSTAT 2022). Besides the fact mainland Portugal has favourable climatic conditions for almond tree growing, the country has also benefited from the implementation of modern irrigation systems and the change from traditional to intensive cropping systems, particularly in the south of the country. Nonetheless, in the northern part of the country, the Trás-os-Montes (TM) agrarian region has a long tradition of almond production, which has led to a great diversity of varieties (Santos and Teixeira 2020; Doll et al. 2021). TM was the region that suffered fewer changes in traditional practices, remaining mostly rainfed, with emphasis on the preservation of native varieties (Doll et al. 2021). These varieties show different development and growth characteristics (Alonso et al. 2005; Sakar et al. 2019), resulting in differences in terms of productivity. The traditional varieties that are representative of this region are “José Dias”, “Duro da Estrada Grado” and “Duro Amarelo Grado” (Oliveira et al. 2018; Santos and Teixeira 2020). Over the decades, other varieties have been cultivated, focusing on a higher resistance to climate change and pests/diseases, performances and productivity, such as the “Ferragnès”, “Penta” and “Tardona” (Cordeiro et al. 2005).
Successful flowering and yield production require the fulfilment of dormancy, which is directly related to the air temperature (Díez-Palet et al. 2019). Dormancy can be divided into two main stages: endodormancy and ecodormancy (Prudencio et al. 2018b). The chill accumulation is associated with the endodormancy phase, when cool temperatures guarantee that plants retain the nutrients incorporated over the growing season, to promote floral differentiation and allow the branches to harden which makes them more resistant to frost (Díez-Palet et al. 2019; Guillamón et al. 2020; Djaman et al. 2021). Once the chill requirements are fulfilled, budbreak may occur (Paper et al. 2010; Prudencio et al. 2018a), and the endodormancy phase gives place to the ecodormancy phase, where heat accumulation is essential. Warm temperatures contribute to the sprouting and flowering buds, fruit ripening and vegetative growth also known as forcing phase (Tabuenca et al. 1972; Gaeta et al. 2018). Chill and heat requirements are species- and cultivar-specific (Prudencio et al. 2018b).
For the next decades, a significant regional temperature rise is predicted (Benmoussa et al. 2017; Salama et al. 2021; Freitas et al. 2022). These changes may have an impact on the dormancy stages and endanger growth and physiological development (Santos et al. 2017). In case the chilling requirements are not fulfilled, it could be manifested with delayed budburst and crop growth, partial anthesis and fruit development may be threatened (Egea et al. 2003; Prudencio et al. 2018b; Djaman et al. 2021). For the forcing phase, warmer temperatures can lead to phenological advancements, unbalanced fruit maturation, enhancing stress conditions, causing disturbances in fruit production and bloom-advancing impacts (Fraga and Santos 2021; Lorite et al. 2020). Moreover, higher temperatures throughout the growing season may also increase the frequency of occurrence of late spring frost, which can damage the flowers (Lorite et al. 2020). According to Guillamón et al. (2022), late frosts can affect 31–40% of the flowering of both extra-early and early varieties.
Several mathematical models have been used to calculate chill and heat accumulation. In general, these models are applied to different species and climatic conditions (Luedeling and Brown 2011). Moreover, these are useful for predicting the impacts of climate change and fundamental for outlining mitigation and adaptation measures (Salama et al. 2021). To determine chill conditions, the chilling hour (CH) and the dynamic (DM, henceforth CP) models are two of the most used models (Fishman et al. 1987a, b; Weinberger 1950). The former is a simpler model, widely used, while the latter is a complex model, which more accurately simulates the plant’s physiological response to cold temperatures (more biologically effective). To calculate heat accumulation, the growing degree day (GDD) and the growing degree hour (GDH) models are also widely used, also representing a simple model and a more elaborated model, respectively (Anderson et al. 1986). The CP and the GDH use optimum temperature thresholds and compensation mechanisms to consider the effects of unfavourable extreme or unusually warm temperatures (Gaeta et al. 2018; Díez-Palet et al. 2019; Salama et al. 2021). Due to these factors, the CP and the GDH do not respond linearly to temperature, which is particularly important under future warmer climates.
In Portugal, studies considering the almond tree chill and heat conditions are still incipient, considering the high importance of almonds for the country’s economy. As such, the present study aims to fill this knowledge gap. The objectives are four-fold: (1) to describe the climatic characteristics of the study area; (2) to understand the evolution of chill and heat accumulation under climate change; (3) to compare simple and complex models for chill and heat estimation; (4) to propose adaptative measures towards climate change risk reduction.
2 Material and methods
2.1 Study area
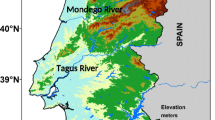
The TM agrarian region, located in north-eastern Portugal (Fig. 1a), is a territorial division with traditional cultural practices regulated by the Portuguese government. It integrates the agricultural subregions of “Alto Tâmega-Barroso”, “Marão-Padrela”, “Terra Fria”, “Planalto Mirandês” and “Terra Quente” (Lima 1999). The almond tree is one of the species that stands out in the region, traditionally rainfed and mostly located in the “Terra Quente” (translates to “warm land”) subregion. Herein, production is mostly limited by weather conditions and water availability (Doll et al. 2021). According to the Portuguese Statistics Office (INE 2022), almond production has been systematically recorded since 1986, reaching 1.7 × 104 t in 2021 (Fig. 1c). The characterization of the almond area was based on the digital inventory provided by the Geographical Institute of Portugal (Carta de Uso e Ocupação do Solo de Portugal Continental—COS 2007) (DGT 2018). Areas with the designation “Pomares de amendoeira”, “Pomares de amendoeira com vinha” and “Pomares de amendoeira com olival” were selected as data source (Fig. 1b).

Location of a mainland Portugal in the Mediterranean region. Almond b distribution in the Trás-os-Montes agrarian region and c yield in the Trás-os-Montes agrarian region from 1980 to 2020 (source: DGT (2007), INE (2022))
2.2 Meteorological variables and climate projections data
In the present study, three time periods were considered, the recent-past baseline (1971–2015), mid-term (2041–2060) and long-term future (2061–2080). For the baseline, the Iberia01 high-resolution observational gridded dataset was selected. This dataset provides climatic variables on a 0.11° latitude × 0.11° longitude (approximately 12.5 km at the equator) and it was considered to be one of the most accurate observational datasets available (Herrera et al. 2019; Freitas et al. 2022). Temperature data for the future periods was retrieved from the EURO-CORDEX project (also 0.11° latitude × 0.11° longitude) and downloaded from the Climate Data Store of the Copernicus Climate Change Service (Jacob et al. 2014).
For future periods, a 5-member ensemble of the global circulation model–regional climate model (GCM-RCM) chain experiment was used (Table 1). This multi-model ensemble covers inter-model variability and reduces single-model uncertainties, explicitly in model setup, initializations and parameterizations (Santos et al. 2017; Freitas et al. 2022). Additionally, two future anthropogenic radiative forcing scenarios were taken into account: RCP4.5 (intermediate emission pathway) and RCP8.5 (high-emissions pathway) (IPCC 2019). These scenarios cover a wide range of uncertainties regarding future greenhouse gas emissions, atmospheric concentrations and climate change magnitudes, with global mean near-surface temperature increases until the end of the twenty-first century ranging from ~ 2 to ~ 4 °C, in RCP4.5 and RCP8.5, respectively (IPCC 2019). RCP4.5 corresponds to an additional (concerning pre-industrial levels) radiative forcing of 4.5 Wm−2 with stabilization after the mid-twenty-first century, while an additional radiative forcing of 8.5 Wm−2 is foreseen until the end of the twenty-first century for RCP8.5 (Thomson et al. 2011; Barredo et al. 2017).
A latitude–longitude sector was defined to optimize the study area, which contains vast continuous almond orchards (Fig. 1b). Gridded daily minimum and maximum air temperatures were extracted from the abovementioned datasets, for both baseline and future periods. Subsequently, the Iberia01 gridded temperatures were extracted for the recent-past period to calibrate the climate model variables with the “Empirical Quantile Mapping” bias-correction method (Cofiño et al. 2018). This methodology was critical to ensure that the bias between observed and modelled variables was correctly adjusted (Fraga et al. 2020; Fonseca et al. 2023). Subsequently, this methodology was then applied to future climate model data.
2.3 Climatic characterization
To characterize the climate of TM, the Iberia01 database (variables: daily mean temperature, in °C, and daily precipitation, in mm) was used to calculate specific parameters. The mean temperature for January (JanMT, in °C) and July (JulMT, in °C) were calculated, as these months correspond to the extremes of the mean seasonal cycle of temperature in the target region. The annual accumulated precipitation (AAP, in mm) and the ratio between the precipitation in the winter half and the summer half of the year (QWS, dimensionless) were also computed. QWS was a simple measure of precipitation seasonality. In TM, on average, from October (previous year) to March (next year) (winter half) there was much higher precipitation than from April to September (summer half), which was a typical feature of northern hemisphere Mediterranean-type climates. The regional orographic characteristics (elevation, in meters above mean sea level) were determined by a digital elevation model (DGT 2018).
2.4 Estimating chill and heat accumulation
The chill and heat requirements of almond trees have been determined using several models. To analyse the differences between them, simple and more complex models were selected herein. In the present study, the chilling during winter dormancy was evaluated using the chilling hours (CH) model (Weinberger 1950) and the dynamic model (DM, henceforth CP) (Fishman et al. 1987a, b). The heat forcing during the growing season was calculated according to the growing degree days (GDD) and growing degree hours (GDH) models (Miranda et al. 2021; Miranda 2020; Anderson et al. 1986). According to Paper et al. (2010) and Weinberger (1950), the CH model is a simple and extensive system to calculate the chill conditions. The model quantifies hours between 0.0 and 7.2 °C (Luedeling and Brown 2011). However, CH does not recognize the adverse effect of warm temperatures on the chill accumulation period (Salama et al. 2021). The chillR package (version 0.72.7; Luedeling 2022) was used to apply the DM in R Core Team (2022), expressed in chill portions (CP) (Fishman et al. 1987a, b). The DM provided accurate estimates at various sites and conditions and takes into consideration the disruptive effect of intermittent higher temperatures in chill accumulation, which is particularly important in warm climate conditions (Díez-Palet et al. 2019). Its response curve is a function of hourly temperatures, with an optimum temperature of 4 °C (Fraga and Santos 2021). In the case of high temperatures, the model applies a neutralizing effect (Luedeling and Brown 2011). The hourly temperatures were estimated from daily data using the built-in package functions.
The fruclimadapt package (version 0.4.4) was used to estimate GDD and GDH values (Miranda et al. 2021; Miranda 2020; Anderson et al. 1986). GDD and GDH were determined from the daily mean minimum and maximum temperatures. The GDD model considers temperatures above the base temperature (Tbase temperature = 4.5 °C), which was calculated as a daily temperature summation (Blanco-Ward et al. 2019). The GDH model was based on the assumption that heat accumulates when the hourly temperature is between a base temperature (4 °C) and the optimum temperature (25 °C), leading to a maximum heat accumulation (Benmoussa et al. 2017; Díez-Palet et al. 2019). Moreover, the model design takes into account warm temperatures with a critical temperature (36 °C). Temperatures above 36 °C cause slow and premature growth and may bring negative effects on the plant stomatal closure (Gu 2015; Santos et al. 2017).
A simulation was performed for the study area, considering the parameters highlighted above. Given the large spatial variability, some assumptions were introduced. For this reason, the chilling phase was estimated to start accumulating on October 1st (of the previous year) and end on February 28th (of the following year). The chilling requirement is often fulfilled in January, though it may shift to February under warm conditions (Santos et al. 2017). For heat accumulation, the period between January 1st and September 30th was considered, from flowering to fruit maturity (Sakar et al. 2019). These relatively long periods were adjusted to the thermal needs of several varieties, according to their different maturation periods, being slightly overlapped. The CP, GDH, CH and GDD were computed separately for each climatic dataset. Subsequently, for future climates, multi-model means were used, to take into account model uncertainty. Finally, the results of the present period were compared to the two future periods: mid- and long-term. For the sake of succinctness, the CH and GDD model results can be found in Supplementary Material.
2.5 Complementary analysis
As already explained, in the heat accumulation phase, temperatures above 36 °C can lead to heat stress, leading the GDH/day values to decrease. To understand these effects, the percentage of days where GDH/day values decrease after June 1st was calculated. In addition, the difference between historical and future scenarios/periods was computed to evaluate to what extent CP and GDH accumulated values will change in the future (increase/decrease). These parameters are useful to growers because they indicate the possibility of insufficient chill and heat conditions during the crop lifetime (Fraga and Santos 2021).
2.6 Chill and heat conditions for almond varieties
The almond tree species has a large diversity of varieties and clones, with a distinct duration of phenological phases. Herein, we chose three representative groups of almond varieties, selected based on their flowering time classification (Table 2): early (represented by “Desmayo”, “Duro da Estrada Grado” and “Duro Amarelo Grado”), mid-season (“Casa Nova”, “Nonpareil” and “Ferraduel”) and late (“Tarraco”, “Penta” and “Tardona”). Therefore, we computed the amount of CP and GDH required by each group, based on their average phenological timings (Table 2), and compared the baseline with the future (mid- and long-term). In this study, we assume that the phenological timings (chilling and forcing phase and flowering) are the same in the present and future scenarios/periods.
3 Results
3.1 Regional characterization
January is usually the coldest month of the year. According to the results, the JanMT (mean temperature for January) values range from < 2 to 9 °C (Fig. 2a). The region shows 40% of the areas that correlate with JanMT between 4.1 and 5 °C, standing out on a large area in the northeast of TM and 28% and 25% of areas corresponding to 3.1–4 °C and 5.1–6 °C classes, respectively. In the southern part of TM, temperatures are higher (6.1–9 °C), due to the influence of the Atlantic Ocean. Considering the grid box, previously defined for the almond tree distribution (Fig. 1b), the orchard is especially distributed in the southeast of TM. This means that the JanMT values corresponding to the almond distribution range between 4 and 7 °C.

A January mean temperature (JanMT; 1971–2015; °C); b July mean temperature (JulMT; 1971–2015; °C); c annual accumulated precipitation (AAP; 1971–2015; mm); d ratio between the precipitation of the winter half and summer half months of the year (QWS; 1971–2015; dimensionless); e elevation (m). For representation purposes, a downscaling methodology was applied, following the work by Freitas et al. (2022)
In contrast, July temperatures are usually the highest of the year, and JulMT (mean temperature for July) values range from 16 to 26 °C for TM (Fig. 2b). According to JulMT, the warmer temperatures are observed in the southeast of the TM region (22.1–25 °C), which coincides with a vast area of almond trees. Overall, 47% of the total area is represented by 22.1–24 °C and 14% of the area corresponds to temperatures above 24 °C, in the southeast. Temperatures below 21 °C are seen in the west of the region. The physiological activity of almond trees can be strongly reduced at temperatures below 15 °C and above 35 °C (Bento et al. 2020).
Regarding AAP (annual accumulated precipitation) (Fig. 2c), for the region, the values range between 250 and 2000 mm. Most of the region (70%) corresponds to values between 501 and 1000 mm. The AAP values tend to decrease from the west (2000 mm) to the east (250 mm). According to the results, 38% and 34% of the area correspond to 501–750 mm and 751–1000 mm classes, respectively. The orchard area is characterized by AAP of 251–750 mm. Almond trees are very resistant to drought periods, being native to regions with 300–600 mm (Aguiar et al. 2017).
The QWS (ratio between the precipitation in the winter half and the summer half of the year) values range from < 1.7 to 2.7 (Fig. 2d), which represents the precipitation seasonality. For this parameter, 34% of the TM corresponds to 1.91–2.1 QWS, in the southeast, whereas the highest values, from 2.61 to 2.7 QWS, stand out to the west. Apropos the almond orchard, QWS values vary between < 1.7 and 2.3, which means that the difference in precipitation between winter and summer is not as high as elsewhere in the region.
Elevation ranges roughly from < 250 to 2000 m (Fig. 3e). In total, 23% of the area corresponds to the elevation class 251–500 m, 44% corresponds to 501–750 m, 24% corresponds to 751–1000 m and 4% corresponds to < 250 m and 1001–1250 m, the other class are residuals. Lower elevation regions are found in the southwest. The mountainous areas are in the north, depicting higher elevations. The almond tree areas correspond to elevations from 64 to 971 m (southeast).

Chilling portions (CP) index for a recent-past period (1971–2015), and future scenarios: RCP4.5 for b 2041–2060 and d 2061–2080, and RCP8.5 for c 2041–2060 and e 2061–2080
3.2 Chill accumulation
The annual mean evolution of daily CP and CP accumulated for historical and future periods are demonstrated in Fig. 3. The chilling phase was estimated to start on October 1st of the previous year and ends on February 28th of the current year (Díez-Palet et al. 2019; Prudencio et al. 2018b, b; AJAP 2017). Figure 3a corresponds to the 1971–2015 period. The daily CP increases from October (0 CP) to December (0.7 CP) and stabilizes until the end of February, decreasing afterward. The accumulated values from October 1st to the end of February reaches 81 CP. From April 1st onwards, the CP accumulation line stabilizes at ~ 110 CP until the end of the season. For the future scenarios and periods (mid- and long-term), the CP data are analysed according to mean, minimum and maximum values attained from the 5 climate models (CP–model range henceforth). For the RCP4.5 scenario and the mid-term period (Fig. 3b), the end of the stabilization period of the daily CP is anticipated (December to January). During the chilling phase (October to February), the accumulated CP is reduced to 76 CP, a decrease of − 5 CP compared to the baseline (81 CP), which is only reached after 9 days (March 9th). For 2061–2080 (Fig. 3d), the accumulated value is 75 CP (− 6 CP) at the end of February, requiring 11 additional days (March 11th) to obtain the same CP as in the past. The CP reduction is more severe under RCP8.5 (stronger anthropogenic forcing) and more evident in the long-term period. In the chilling phase, the accumulated values of 75 CP and 69 CP correspond to 2041–2060 (Fig. 3c) and 2061–2080 (Fig. 3e), respectively. The CP/day stabilization periods are also reduced. For 2041–2060, to obtain the same 81 CP of the recent-past, 17 additional days are required (− 8 CP), while for 2061–2080, 33 days (April 2nd) are needed. According to the climate model means, in all future scenarios/periods the baseline value of 81 CP will not be met until the end of February. Nonetheless, regarding the climate model range for the accumulated CP, these values may be reached, taking into account the maximum CP values, while the model minimum can have very low values of CP. These very low CP values are particularly noticeable under RCP8.5 and more pronounced in the long-term period.
The results of the CH model (SM 1) indicate that the chill accumulation will be lower in the future compared to the recent-past period. SM 1a is following the baseline, the total number of chilling hours is 1375 CH. The CH/day values vary from 0 and 15 CH. For RCP4.5, 2041–2060 (SM 1b), during the chilling phase 1082 CH, a lower CH value compared recent-past period (− 293 CH). In the 2061–2080 period (SM 1d), the accumulated value obtained at the end of the chilling phase is 1079 CH (− 296 CH). For RCP8.5, the values are 1022 CH (− 353 CH) and 894 CH (− 481 CH), corresponding to the mid- (SM 1c) and long-term periods (SM 1e), respectively. Contrarily to the CP, the CH model does not consider the disruptive effect of a warm temperature period on the chill accumulation. Therefore, the results may not be as realistic as in the DM. Hence, the CH/day continues to increase from December to February.
3.3 Heat accumulation
The forcing phase follows the chilling phase and it is estimated between January 1st and September 30th, in agreement with the GDH model (Fig. 4), from flowering to fruit maturity. The evolution of GDH/day and accumulated GDH for the recent-past and future periods under both scenarios are illustrated in Fig. 4. For the baseline (Fig. 4a), the reference value is 5.8 × 104 GDH during the period from January to September. Overall, from January to June there is a continuous increase in daily GDH values (384 GDH), followed by a subtle negative slope until August, decreasing constantly afterward. The GDH/day curve shows two peaks, in June and September. These two peaks may correspond to the upper-temperature limit that is considered for GDH accumulation. In the RCP4.5 scenario, the accumulated GDH is 6.1 × 104 GDH for the two periods (Fig. 4b and d). For RCP8.5, the increase in GDH is more pronounced (Fig. 4c and e) for the 2061–2080 period, 6.2 × 104 (Fig. 4e). Generally, in the future scenarios/periods, the accumulated GDH tends to increase, though differences appear to be small (compared to recent-past values), possibly due to the GDH temperature thresholds. From June to September, there is a negative curve when the GDH value reduces due to high temperatures (> 36 °C). This effect is particularly visible in 2061–2080, RCP8.5. Nonetheless, all future scenarios/periods indicate that the reference value 5.8 × 104 GDH will be reached before the end of the forcing phase.

Growing degree hours (GDH) index for the a recent-past period (1971–2015), and future scenarios: RCP4.5 for b 2041–2060 and d 2061–2080, and RCP8.5 for c 2041–2060 and e 2061–2080
Concerning the results of the GDD model (SM 2), heat conditions show an increase in the future compared to the baseline. It should be noted that the GDD model does not have an upper-temperature threshold, such as the GDH. For the baseline, the daily GDD values range between 0 and 22 GDD, and the accumulated GDD reached 2665 GDD (SM 2a). For RCP4.5, in 2041–2060 (SM 2b), during the forcing phase a value of 3010 GDD is achieved, higher when compared to recent-past (344 GDD). For the same scenario, in 2061–2080 (SM 2d), the accumulated value obtained at the end of the chilling phase is 3046 GDD (381 GDD). For RCP8.5, the values are 3095 GDD (430 GDD) and 3291 GDD (626 GDD) for 2041–2060 (SM 2c) and 2061–2080 (SM 2e), respectively.
3.4 Complementary analysis
In this analysis, we compare the percentage of days when the value is lower than June 1st (Fig. 5), which usually corresponds to the highest GDH value (1st peak date) (Fig. 4), for recent-past and future periods/scenarios. These results indicate that, from June to October, the temperature will rise significantly, resulting in lower GDH values, as temperatures rise above the critical threshold (36 °C) (Miranda 2020; Fraga et al. 2019). In the baseline, the maximum percentage of 64% is reached. For RCP4.5, the maximum is 81% and 89% for 2041–2060 and 2061–2080, respectively. For RCP8.5, the maximum is 79% and 99% for 2041–2060 and 2061–2080, correspondingly. As previously mentioned, this future percentage increase is associated with the limits set by the model when temperatures reach 36 °C.

Percentage of days when the GDH value is lower than the GDH value on the June 1st (1st GDH peak)
The CP (Fig. 6a) and GDH (Fig. 6b) accumulated and the respective differences between each future period/RCP and historical are illustrated in Fig. 6. From October 1st (previous year) to September 30th (next year), the CP accumulated is calculated. From October until January, a sharp decrease is visible, with a stabilization from January until the end of February, followed by a strong decrease in CP until June, when a stabilization occurs. At the end of the chilling phase (end of February), anomalies are already observed, especially for the 2061–2080 period, with − 8 and − 12 for RCP4.5 and RCP8.5, respectively. Conversely, in the GDH case (Fig. 6b), future values tend to increase until June, decrease until September and then increase again. The decrease in the difference values, from June to September, is associated with the high summer temperature and the increase projected for the future. At the end of December, the anomalies reach their maximum value between 4.8 × 103 and 6.2 × 103, for RCP4.5, 2041–2060 and 2061–2080, and RCP8.5, 2041–2060. In RCP8.5, 2061–2080, the difference corresponds to 8.5 × 103.

Difference between the mean value of each scenario/future period and the recent-past period value for a chilling portions and b growing degree hours
3.5 Evaluation of chill and heat conditions for flowering
The chill and heat conditions are now assessed taking into account the 3 varietal groups defined for the flowering stage: early, mid-season and late (Fig. 7). For the baseline, for budbreak the early varietal group shows 42 CP, the mid-season 63 CP and the late season 81 CP. For flowering, during the forcing phase, early varieties accumulate up to 2.5 × 103 GDH, mid 3.0 × 103 GDH and late 4.6 × 103 GDH. Concerning the RCP4.5 and regarding budbreak, a reduction in CP values occurs for all varietal groups and is more significant in the long-term. In general, compared to the baseline, in 2061–2080 a reduction of around − 8 CP can be observed. Conversely, for flowering, heat accumulation tends to increase, with higher intensity for the late varieties, as they endure the warming later in the season. For RCP8.5, the most pertinent CP reduction is found in 2061–2080, where compared to the baseline, there is a more pronounced decrease in late (− 12 CP) than in early varieties (− 9 CP). In comparison with recent-past, the GDH increase is around 1.5 × 103 at the flowering period, for all varieties.

Chill (CP) and heat (GDH) accumulation throughout the almond tree annual cycle, for the recent-past and RCP4.5 and RCP8.5 future scenarios (annual means for each period: 1971–2015, 2041–2060 and 2061–2080). The flowering periods for early, mid-season and late varieties are also represented according to Table 2
4 Discussion
The constant search, by producers and scientific teams, for tools to identify the challenges and present solutions is increasing, and the evolution of technology has contributed to the effectiveness of this task. Climatic and phenological descriptions help the producers select the cultivar and the most effective agricultural practices for each situation (Valentini et al. 2001). As is known, the success of the chill and forcing phase is essential for the well-being of the plant and productivity. There is an inter-dependent relationship between the chill and forcing phase, e.g. the later the chill phase ends, the later the forcing phase begins (Díez-Palet et al. 2019). Adequate development of floral buds and fruits, efficient pollination process and fruit set depend on the thermal requirements being fulfilled (Prudencio et al. 2018b). In the future, the predicted climate change may have detrimental impacts on the chill and heat conditions, threatening the species’ stability, and the almond tree sector as a whole.
The TM agrarian region, where winters are relatively cool and rainy while summers are hot and dry, is considered suitable for almond orchards, particularly in the southeast (“Terra Quente”) (Bento et al. 2020). Chilling and forcing conditions were analysed for the almond trees in this region, for recent-past and future scenarios, also taking into account different varietal requirements. Given the results for the recent-past, the chill and heat conditions in the TM region may exhibit different behaviours in the future, chill accumulation tends to decrease and heat accumulation tends to increase, which may influence the prevalence of this crop.
Currently, this region shows CP values ranging from 75 to 95 CP, which is in agreement with Santos et al. (2017). For the future, the CP reduction is almost − 12 CP, which may have negative impacts. The chill reduction also applies to the results of the CH model. In the baseline, the value is 1375 CH and, in the future, the reduction is − 482 CH. It should be noted that the CH (− 35%) reduction is much stronger than the CP (− 15%). Even though the results express a chill reduction over the TM region under future climates, almond trees should still be able to fulfil their chilling requirements. However, if a stronger chill reduction indeed occurs, this could lead to complications, such as delayed phenological stages, asynchronous flowering and loss of flower buds and flowers (Egea et al. 2003; Prudencio et al. 2018b; Djaman et al. 2021; Tominaga et al. 2022). Moreover, it is fundamental to highlight the main difference between these chill models, the DM takes into account that in the case of high temperatures, the model applies a compensating effect, which is not the case with the CH model (Luedeling and Brown 2011). For this reason, it is believed that the DM is more accurate than the CH model, particularly regarding climate change projections.
Regarding forcing conditions, heat accumulation is projected to rise due to higher temperatures. According to the baseline, heat accumulation is 5.8 × 104 GDH. Considering the most severe scenario, the GDH will rise 2.8 × 103 and 3.5 × 103, for mid- and long-term, respectively. In baseline, the GDD value is 2665 and the results show a rise of 430 GDD and 626 GDD, for periods mid- and long-term, in the RCP8.5. The increase in the GDD (+ 24%) is much more prominent than the GDH (+ 7%). This is due to a reduction in GDH during the June to September period (RCP8.5; long-term). Effectively, higher temperatures during this period (temperatures above 36 °C) affect the plant’s stomatal closure (Gu 2015) and negatively influence fruit development, vegetative growth and flower differentiation which will influence the following year’s yield (Cantin et al. 2020; Fraga and Santos 2021), and thus are discarded by the GDH model. Conversely, the GDD model does not establish a limit for maximum temperatures in summer. As the results suggest, regarding climate change projections, the use of the CP and GDH models, compared to CH and GDD, is recommended, as these models use statistical tools to reduce the unfavourable effect of warm temperatures (Gaeta et al. 2018; Salama et al. 2021). The most noticeable differences, between CP and GDH models, occur between the baseline and the RCP8.5 (long-term). According to the results and analysis, future simulations for the DM and GDH models could be useful tool in crop management and in forecasting climate change impacts, and growers should take this into account.
Regarding the linkage between the chilling and forcing models and almond phenology, 3 varietal groups, from the TM region, were analysed: early, mid-season and late varieties (Table 2). Almond trees require relatively low chilling to budbreak, already in late winter, well before other fruit trees (Aguiar et al. 2017; Prudencio et al. 2018b). For chill accumulation, in the baseline period, CP ranges from 42 to 81 (early to late seasons). In other studies, the CP values range from 8 to 55 CP (early to extra-late—41 to 106 days) (Benmoussa et al. 2017; Prudencio et al. 2018a, b; Díez-Palet et al. 2019; El Yaacoubi et al. 2019; Djaman et al. 2021). In future conditions (RPC8.5; long-term), a reduction in CP values is observed, being more significant for late varieties (− 12 CP). This reduction should not compromise the almond tree chilling requirements in the TM region, even under the most severe climate scenario. Nonetheless, a stronger reduction in chill may still occur. If this is the case, late varieties may experience greater impacts, possibly leading to more pronounced changes in phenology, such as delayed flowering, and other physiological disorders (Prudencio et al. 2018a, b). As an example, in south-eastern Spain, studies found that late varieties, such as Tardona and Penta, can have yield losses associated with chill requirements not being met (Prudencio et al. 2018a, b). Conversely, early and mid-season varieties should be less affected by a chill reduction, due to the lower chilling requirements.
From budbreak onwards, heat accumulation will increase under both future scenarios, with increased severity under RCP8.5. Regarding the varietal groups, late varieties will tend to experience this enhanced warming more than early or mid-season. This fast and intensive increase in heat accumulation may contribute to the advancement of flowering and maturity dates (Prudencio et al. 2018b). The occurrence of earlier flowering dates can increase the exposure of flowers and fruits to the frost damage, especially early flowering cultivars. Other effects of the temperature increase in the forcing phase are photoassimilates inhibition, disturbances in fruit formation with a smaller kernel, anticipated harvest and yield, and quality attributes reduction (Luedeling et al. 2013; Lorite et al. 2020). Studies report that for the heat accumulation up to flowering, GDH values can vary between 5.5 × 103 (e.g. Desmayo) and 9.0 × 103 (Penta) GDH (Egea et al. 2003; Prudencio et al. 2018b; Díez-Palet et al. 2019). Overall, the increase in temperatures during the season, which contribute to the decreased chilling and increased forcing will tend to shorten the duration of the phenological stages, which has already been reported by several studies (Fraga and Santos 2021).
It should be mentioned that the current study does not consider other factors that may also influence almond development and growth (Wolkovich et al. 2022). The impact of the increase in pest and disease pressure (Aguiar et al. 2017). The impact of spring frost, which can also significantly affect almond productivity, especially in cold regions, such as TM (Segara and Ossama 2007; Lorite et al. 2020). Early-bloom cultivars are more vulnerable to frost impacts compared to late-bloom varieties (Guillamón et al. 2022). Moreover, other climatic variables could affect the species’ growth, such as the decreased annual precipitation that may increase drought stress (Doll et al. 2021). Additionally, this current study provides a simulation based on climatic factors that are prone to several uncertainty sources, such as model and scenario uncertainty (Bedia et al. 2013). Furthermore, we assume that the phenological timings (chilling and forcing phase and flowering) are the same in the present and future scenarios/periods, which may not occur.
Adaption measures may be used to reduce potential yield/quality losses and ensure the sector’s long-term viability. To address the lack of chill, growers can take advantage of the application of bio-stimulants or even adopt more suitable scion-rootstock combinations (Prudencio et al. 2018a, b). To decrease heat accumulation, the implementation of sunscreens or shading systems is recommended (Fraga and Santos 2021). In the case of almond species, growers focus on developing varieties that are more resistant to prevent the effects of spring frosts. Earliest varieties are more sensitive to frost events, since the flowers are already in development, and can be easily damaged by these extreme events (Guillamón et al. 2022). Contrarily, late varieties present good adaptability to different climatic conditions, which show naturally more resistance to this event, owing to the later flowering period (Martínez-Gómez et al. 2017). To reduce the decreased precipitation impacts, in a sustainable manner, the implementation of irrigation systems can be an option that helps during the hottest and driest months, or crop relocation (Bento et al. 2020).
Heatwaves can also be harmful to the species, and to reduce their impact select more heat-resistant varieties and apply for fertilizer and bio-stimulating plant protection under environmental stress are proposed (Freitas et al. 2021). To combat the other climate impacts, it is recommended to cultivate varieties that are most suited to the local climate and are more resilient to climate change, as well as to employ modern breeding techniques, like mapping the genetic basis of chilling requirements, to improve the viability and productivity of almond trees (Luedeling 2012). Additionally, the support for the breeding of new enhanced varieties for predetermined chilling and heat condition values is essential for climate change risk reduction (Guillamón et al. 2022). To address these issues, additional studies are required, such as calculating the occurrence of extreme weather events (e.g. spring late frost and heatwaves), precipitation irregularity over the year, water supply in the future and the respective impacts they have on the almond trees.
5 Conclusions
Climate challenges are being experienced with each passing day. According to the climatic projections, the TM agrarian region, defined by traditional agricultural practices and almond varieties, may be more susceptible to significant climate change impacts. As the temperature is expected to increase, chill and heat conditions tend to change. As seen in this study, increased temperature during the forcing phase should be more problematic than during the chilling phase. As can be seen, the use and development of mathematical models are fundamental, since they are auxiliary tools for crop prediction and management. These models can also be considered as a decision-support tool for farmers and agricultural consultants, namely, concerning the selection of varieties and cultural and management practices. Adaptation measures must be timely planned and implemented so that the sector does not suffer economic and biodiversity losses. Therefore, investment should be made in more resilient and sustainable varieties and more sophisticated and adapted agricultural practices to adapt to climate change, reducing the negative impacts. Lastly, the present study contributes to a better understanding of the evaluation of thermal conditions for almond species in the TM agrarian region. Furthermore, since the orographic and climatic characteristics of the TM region may be similar to other regions of the world, this study may be used as a reference. It is hoped that this study will be useful in almond management and inspire further work.
References
Aguiar C, Pereira J, Arrobas M, et al (2017) Estado de produção - Centro Nacional de Competências dos Frutos Secos. Portugal
AJAP (2017) Manual Boas Práticas para Culturas Emergentes - A cultura da amêndoa, Associação de Jovens Agricultores de Portugal. Portugal
Alonso JM, Ansón JM, Espiau MT, Socias I, Company R (2005) Determination of endodormancy break in almond flower buds by a correlation model using the average temperature of different day intervals and its application to the estimation of chill and heat requirements and blooming date. J Am Soc Horticult Sci 130:308–318. https://doi.org/10.21273/jashs.130.3.308
Alonso JM, Espiau MT, Socias R (2010) Increase in the chill and heat requirements for blooming of the new almond cultivars. Opt Méditerranéennes Sér A Mediterranean Sem 69:65–69
Anderson JL, Richardson EA, Kesner CD (1986) Validation of chill unit and flower bud phenology models for “Montmorency” sour cherry. Acta Horticulturae 71–78
Barredo JI, Caudullo JI, Mauri G (2017) Mediterranean habitat loss under RCP4.5 and RCP8.5 climate change projections. European Commision; JRC Technical Reports 38. https://doi.org/10.2760/622174
Bedia J, Herrera S, Gutiérrez JM (2013) Dangers of using global bioclimatic datasets for ecological niche modeling. Limitations for future climate projections. Global Planet Change 107:1–12. https://doi.org/10.1016/j.gloplacha.2013.04.005
Benmoussa H, Ghrab M, Ben M, Luedeling E (2017) Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agric for Meteorol 239:34–46. https://doi.org/10.1016/j.agrformet.2017.02.030
Benmoussa H, Ben Mimoun M, Ghrab M, Luedeling E (2018) Climate change threatens Central Tunisian nut orchards. Int J Biometeorol 62:2245–2255
Bento A, Aires A, Silva AP, et al (2020) Manual prático da amendoeira - Centro Nacional de Competências dos Frutos Secos. Portugal
Blanco-Ward D, Monteiro A, Lopes M et al (2019) Climate change impact on a wine-producing region using a dynamical downscaling approach: climate parameters, bioclimatic indices and extreme indices. Int J Climatol 39:5741–5760. https://doi.org/10.1002/joc.6185
Cabo P, Matos A (2017) Amendoeira: Estado da Comercialização - Centro Nacional de Competências dos Frutos Secos. Portugal
Cantin CM, Wang XW, Almira M, et al (2020) Inheritance and QTL analysis of chilling and heat requirements for flowering in an interspecific almond x peach (Texas x Earlygold) F2 population. Euphytica 216:. https://doi.org/10.1007/s10681-020-02588-9
Cofiño AS, Bedia J, Iturbide M et al (2018) The ECOMS user data gateway: towards seasonal forecast data provision and research reproducibility in the era of climate services. Climate Services 9:33–43. https://doi.org/10.1016/j.cliser.2017.07.001
Cordeiro V, Alves C, Vieira J, Barroso MR (2005) Evaluation of almond cultivar adaptation in Trás-os-Montes region (Portugal). Options Méditérr 116:113–116
Costa J, Rosa A (2020) Colecção de amendoeiras do algarve caracterização morfológica de variedades, Direção Regional de Agricultura e Pescas do Algarve. Portugal
DGT (2018) Modelos Digitais de Terreno e de Superfície. https://www.dgterritorio.gov.pt/cartografia/cartografia-topografica/modelos-digitais-do-terreno. Accessed 15 Aug 2022
Díez-Palet I, Funes I, Savé R et al (2019) Blooming under Mediterranean climate: estimating cultivar-specific chill and heat requirements of almond and apple trees using a statistical approach. Agronomy 9:1–21. https://doi.org/10.3390/agronomy9110760
Djaman K, Koudahe K, Darapuneni M, Irmak S (2021) Chilling and heat accumulation of fruit and nut trees and flower bud vulnerability to early spring low temperatures in New Mexico: meteorological approach. Sustainability 13:23. https://doi.org/10.3390/su13052524
Doll D, Freire De Andrade J, Serrano P (2021) Produção de amêndoa em Portugal: Tendências de plantação e desafios de produção num sector em desenvolvimento, Agro.Ges Estudo e Projetos. Portugal
Egea J, Ortega E, Martínez-Gómez P, Dicenta F (2003) Chilling and heat requirements of almond cultivars for flowering. Environ Exp Bot 50:79–85. https://doi.org/10.1016/S0098-8472(03)00002-9
El Yaacoubi A, Oukabli A, Legave JM, et al (2019) Response of almond flowering and dormancy to Mediterranean temperature conditions in the context of adaptation to climate variations. Scientia Horticulturae 257. https://doi.org/10.1016/j.scienta.2019.108687
FAOSTAT (2022) FAOSTAT - crops and livestock products. In: 2022. https://www.fao.org/faostat/en/#data/QCL. Accessed 24 Feb 2022
Fishman S, Erez A, Couvillon GA (1987a) The temperature dependence of dormancy breaking in plants: computer simulation of processes studied under controlled temperatures. J Theor Biol 126:309–321. https://doi.org/10.1016/S0022-5193(87)80237-0
Fishman S, Erez A, Couvillon GA (1987b) The temperature dependence of dormancy breaking in plants: mathematical analysis of a two-step model involving a cooperative transition. J Theor Biol 124:473–483. https://doi.org/10.1016/S0022-5193(87)80221-7
Fonseca A, Fraga H, Santos J (2023) Exposure of Portuguese viticulture to weather extremes under climate change. Climate Services 30:. https://doi.org/10.1016/j.cliser.2023.100357
Fraga H, Santos JA (2021) Assessment of climate change impacts on chilling and forcing for the main fresh fruit regions in Portugal. Front Plant Sci 12:1–13. https://doi.org/10.3389/fpls.2021.689121
Fraga H, Pinto JG, Santos JA (2019) Climate change projections for chilling and heat forcing conditions in European vineyards and olive orchards: a multi-model assessment. Clim Change 152:179–193. https://doi.org/10.1007/s10584-018-2337-5
Fraga H, Pinto JG, Santos JA (2020) Olive tree irrigation as a climate change adaptation measure in Alentejo Portugal. Agric Water Manag 237:106193. https://doi.org/10.1016/j.agwat.2020.106193
Freitas TR, Santos JA, Silva AP, Fraga H (2021) Influence of climate change on chestnut trees: a review. Plants 10:1–17. https://doi.org/10.3390/plants10071463
Freitas TR, Santos A, Silva AP et al (2022) Climate change projections for bioclimatic distribution of Castanea sativa in Portugal. Agronomy 12:16. https://doi.org/10.3390/agronomy12051137
Gaeta L, Maria A, Losciale P (2018) Evaluation of three modelling approaches for almond blooming in Mediterranean climate conditions. Eur J Agron 97:1–10. https://doi.org/10.1016/j.eja.2018.04.005
Gu S (2015) Growing degree hours - a simple, accurate, and precise protocol to approximate growing heat summation for grapevines. Int J Biometeorol 60:1123–1134. https://doi.org/10.1007/s00484-015-1105-8
Guillamón JG, Egea J, Mañas F et al (2022) Risk of extreme early frosts in almond. Horticulturae 8:687. https://doi.org/10.3390/horticulturae8080687
Guillamón JG, Prudencio ÁS, Yuste JE, et al (2020) Ascorbic acid and prunasin, two candidate biomarkers for endodormancy release in almond flower buds identified by a nontargeted metabolomic study. Horticulture Research 7:. https://doi.org/10.1038/s41438-020-00427-5
Herrera S, Gutiérrez JM, Cardoso RM, et al (2019) Iberia01: daily gridded (0.1° resolution) dataset of precipitation and temperatures over the Iberian Peninsula. In: DIGITAL.CSIC. http://hdl.handle.net/10261/183071
INE (2022) Portuguese Statistical Office - Produção de amendoa. In: 2022. https://www.ine.pt/xportal/xmain?xpid=INE&xpgid=ine_indicadores&indOcorrCod=0000020&xlang=pt&contexto=bd&selTab=tab2. Accessed 8 Aug 2022
IPCC, 2019: Climate change and land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems [P.R. Shukla, J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H.-O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi, J. Malley, (eds.)]. In press.
Jacob D, Petersen J, Eggert B et al (2014) EURO-CORDEX: new high-resolution climate change projections for European impact research. Reg Environ Change 14:563–578. https://doi.org/10.1007/s10113-013-0499-2
Jin Y, Chen B, Lampinen BD, Brown PH (2020) Advancing agricultural production with machine learning analytics: yield determinants for California’s almond orchards. Front Plant Sci 11:1–15. https://doi.org/10.3389/fpls.2020.00290
Kiesel J, Stanzel P, Kling H et al (2021) Correction to: streamflow-based evaluation of climate model sub-selection methods. Clim Change 169:4–5. https://doi.org/10.1007/s10584-021-03252-4
Kjellström E, Bärring L, Nikulin G et al (2016) Production and use of regional climate model projections – a Swedish perspective on building climate services. Climate Services 2–3:15–29. https://doi.org/10.1016/j.cliser.2016.06.004
Larsson Ivanov O, Bärring L, Wilcke RAI (2022) Climate change impact on snow loads in northern Europe. Structural Safety 97:. https://doi.org/10.1016/j.strusafe.2022.102231
Lima C (1999) A agricultura de Tás-os-Montes e Alto Douro Diagnóstico Prospetivo, Estatística e Estudos Regionais. Portugal
Lorite IJ, Cabezas-Luque JM, Arquero O et al (2020) The role of phenology in the climate change impacts and adaptation strategies for tree crops: a case study on almond orchards in Southern Europe. Agric Forest Meteorol 294:108142. https://doi.org/10.1016/j.agrformet.2020.108142
Luedeling E (2012) Climate change impacts on winter chill for temperate fruit and nut production: a review. Sci Hortic 144:218–229. https://doi.org/10.1016/j.scienta.2012.07.011
Luedeling E, Brown PH (2011) A global analysis of the comparability of winter chill models for fruit and nut trees. Int J Biometeorol 55:411–421. https://doi.org/10.1007/s00484-010-0352-y
Luedeling E, Guo L, Dai J et al (2013) Differential responses of trees to temperature variation during the chilling and forcing phases. Agric for Meteorol 181:33–42. https://doi.org/10.1016/j.agrformet.2013.06.018
Luedeling, E (2022) chillR: statistical methods for phenology analysis in temperate fruit trees. R package version 0.72.7
Martínez-Gómez P, Prudencio AS, Gradziel TM, Dicenta F (2017) The delay of flowering time in almond: a review of the combined effect of adaptation, mutation and breeding. Euphytica 213:1–10. https://doi.org/10.1007/s10681-017-1974-5
Miranda C (2020) fruclimadapt: evaluation tools for assessing climate adaptation of fruit tree species. R package version 0.4.4. https://CRAN.R-project.org/package=fruclimadapt
Miranda C, Urrestarazu J, Santesteban LG (2021) fruclimadapt: an R package for climate adaptation assessment of temperate fruit species. Comput Electron Agric 180:105879. https://doi.org/10.1016/j.compag.2020.105879
NPR (2021) Climate change in California is threatening the world’s top almond producer. https://www.npr.org/2021/08/17/1028452988/climate-change-california-drought-heat-almond-production. Accessed 13 Sep 2022
Oliveira I, Meyer A, Afonso S et al (2018) Morphological, mechanical and antioxidant properties of Portuguese almond cultivars. J Food Sci Technol 55:467–478. https://doi.org/10.1007/s13197-017-2955-3
Paper C, Sagredo K, Reginato G (2010) Prediction models for chilling and heat requirements to estimate full bloom of almond cultivars in the Central Valley of Chile. Acta Horticulturae. https://doi.org/10.17660/ActaHortic.2010.872.12
Prudencio AS, P M-G, Dicenta F (2018a) Evaluation of breaking dormancy, flowering and productivity of extra-late and ultra-late flowering almond cultivars during cold and warm seasons in South-East of Spain. Scientia Horticulturae 235:39–46https://doi.org/10.1016/j.scienta.2018.02.073
Prudencio AS, Dicenta F, Martínez-Gómez P (2018b) Monitoring dormancy transition in almond [Prunus dulcis (Miller)Webb] during cold and warm Mediterranean seasons through the analysis of a DAM (dormancy-associated MADS-box) gene. Horticulturae 4:2–9. https://doi.org/10.3390/horticulturae4040041
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. URL https://www.R-project.org/
Sakar EH, El Yamani M, Boussakouran A, Rharrabti Y (2019) Codification and description of almond (Prunus dulcis) vegetative and reproductive phenology according to the extended BBCH scale. Sci Hortic 247:224–234. https://doi.org/10.1016/j.scienta.2018.12.024
Salama AM, Ezzat A, El-Ramady H, et al (2021) Temperate fruit trees under climate change: challenges for dormancy and chilling requirements in warm winter regions. Horticulturae 7:. https://doi.org/10.3390/horticulturae7040086
Santos AL, Teixeira SM (2020) Panorama nacional do amendoal. Revista de Ciências Agrárias 43:3–9. https://doi.org/10.19084/rca.19684
Santos JA, Costa R, Fraga H (2017) Climate change impacts on thermal growing conditions of main fruit species in Portugal. Clim Change 140:273–286. https://doi.org/10.1007/s10584-016-1835-6
Segara JMA, Ossama K (2007) Physical aspects of almond fruit quality. FAO-CIHEAM - Nucis-Newsletter
Tabuenca MC, Mut M, Herrero J (1972) The effect of temperature on flowering date in almond varieties. Spain Cons Super Invest Cient Estac Exp Aula Dei Anales 11:378–395
Thomson AM, Calvin KV, Smith SJ et al (2011) RCP4.5: a pathway for stabilization of radiative forcing by 2100. Clim Change 109:77–94. https://doi.org/10.1007/s10584-011-0151-4
Tominaga A, Ito A, Sugiura T, Yamane H (2022) How is global warming affecting fruit tree blooming? “Flowering (dormancy) disorder” in Japanese pear (Pyrus pyrifolia) as a case study. Front Plant Sci 12:1–17. https://doi.org/10.3389/fpls.2021.787638
Valentini N, Me G, Ferrero R, Spanna F (2001) Use of bioclimatic indexes to characterize phenological phases of apple varieties in Northern Italy. Int J Biometeorol 45:191–195. https://doi.org/10.1007/s004840100107
Weinberger J (1950) Chilling requirements of peach varieties. Proc Am Soc Hortic Sci 56:122–128
Wolkovich EM, Chamberlain CJ, Buonaiuto DM et al (2022) Integrating experiments to predict interactive cue effects on spring phenology with warming. New Phytol 235:1719–1728. https://doi.org/10.1111/nph.18269
Acknowledgements
This work was financed by the CoaClimateRisk “O impacto das alterações climáticas e medidas de adaptação para as principais culturas agrícolas na região do Vale do Côa” project (COA/CAC/0030/2019) financed by National Funds by the Portuguese Foundation for Science and Technology (FCT). This work was also supported by projects UIDB/04033/2020 and LA/P/0126/2020. A.F. thanks Fundo Europeu de Desenvolvimento Regional (FEDER), Programa Operacional Regional do Norte, under the project “Soil recover for a healthy food and quality of life” (SoilRec4+Health), NORTE-01-0145-FEDER-000083. HF thanks the FCT for contract CEECIND/00447/2017 and 2022.02317.CEECIND.
Funding
Open access funding provided by FCT|FCCN (b-on). This work was financed by the CoaClimateRisk project (COA/CAC/0030/2019) and financed by National Funds by the Portuguese Foundation for Science and Technology (FCT).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study’s conception and design. Conceptualization: T.R.F. and H.F.; methodology: T.R.F., J.A.S. and H.F.; software: T.R.F., A.F. and H.F.; validation: T.R.F. and H.F.; resources: T.R.F. and A.P.S.; data curation: T.R.F.; writing—original draft preparation: T.R.F.; writing—review and editing: J.A.S., A.P.S., A.F. and H.F.; visualization: J.A.S., A.P.S. and H.F.; supervision: H.F.; project administration: H.F.; funding acquisition: H.F. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Freitas, T.R., Santos, J.A., Silva, A.P. et al. Evaluation of historical and future thermal conditions for almond trees in north-eastern Portugal. Climatic Change 176, 89 (2023). https://doi.org/10.1007/s10584-023-03569-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10584-023-03569-2