
Abstract
This study aimed at determining shoot and root characteristics of cassava as affected by root yield and the influence of soil moisture on vegetative growth and yield. Thirty cassava genotypes were evaluated for morphological and physiological characterization in three locations in Nigeria: Ibadan, Mokwa and Zaria. Randomized complete block design was used with four replicates. Studies on the pasting properties of the genotypes were also carried out. Data were collected on plant height, stem girth, stay-green ability, garri and fresh root weight. Genotypes differed significantly (P < 0.05) across and within locations for shoot and root characteristics. Across locations, genotype 011663 had the highest plant height (132.4 cm); 30572 had the largest stem girth (8.6 cm); and 010040 was the best stay-green (2.2). Genotype 011086 had the highest number of roots per plot (95.7), 950289 had the highest fresh root yield (24.3 t/ha), and 990554 had the highest percentage of dry matter (35.2). Trends in root yields across locations were Ibadan (28.9 t/ha), Mokwa (20.3 t/ha), and Zaria (8 t/ha). Five genotypes IITA-TMS-IBA950289, 010034, 990554, 011807, and 980581 had negligible interactions with the environment and so have broad adaptation and are considered stable; and two clones 011807 and 950166 were found to be the best for pasting properties. Breeding strategies that consider root size, total root number, harvest index, dry matter, with applications for household foods and industrial uses, will be an effective and efficient way to select genotypes for high yield.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cassava is a major food crop in sub-Saharan Africa (Hahn et al. 1989). In the past, production in most countries was mainly for the starchy tuberous roots that are valuable sources of cheap calories particularly for low-income earners and resource-poor farmers. However, in recent times, the crop is gradually gaining a strategic position in world trade as a result of the efforts by various governments and the private sector in developing novel, value-added cassava-based products for human consumption and industrial uses (Dixon et al. 2005). Indeed, cassava can serve as an important engine for growth in many countries if production, diversification, and commercial uses are improved (Anon 2000).Nigeria stands out as the world’s largest producer with increased production and average output of 42.5million tons and 12.3t/ha in 2010 to 54 million tons and 14.03 t/ha in 2012 harvested from 3.85 million hectares of land (FAOSTAT 2013). Data from the Collaborative Study of Cassava in Africa (COSCA) showed that 80% of Nigerians in the rural areas eat a cassava meal at least once a week with increased average per capital consumption over 5 years from 88 kg/person/year to 120 kg/person/year between 1965 and 1998 (Nweke et al. 2002).
Cassava performs five main roles: famine reserve crop, rural food staple, cash crop for foreign exchange, industrial raw material, and foreign exchange earner. This suggests that Nigeria is the most advanced of the African countries poised to diversify the use of cassava as a primary industrial raw material and livestock feed (Nweke et al. 2002). Nutritionally, cassava contains potassium, iron, calcium, vitamin A, C, and B-6, folic acid, sodium, and protein, all vital in the human diet (IITA 2005). It provides some protein, minerals, (iron and calcium) and vitamins (A and C) through the consumption of the leaves. It gives stable yields even in the face of drought, low soil fertility, and low intensity management (IITA 1998). It can be used in products of many types, such as food (abacha, fufu, lio-lio, tapioca, cassava flour and Garri (Ihenkoronye and Ngoddy 1985; Iwuoha et al.1996), confectionery, sweeteners, glues, plywood, textiles, papers, and biodegradable products, also in the manufacture of monosodium glutamate and medicines, dry chips, and alcohol (IITA 2005).
Cassava grows and produces well in the Nigerian environment, especially in the southern parts. However, it shows different growth behaviors and yields in different years as a result of differences in annual weather conditions. The water regime of an environment is another important factor which affects growth. Differential soil water and nutrient regimes have been reported to affect yield stability (Cock 1985). Although cassava is extremely tolerant to water stress, prolonged moisture deficiency leading to reduced growth, development, and root yield have been reported (CIAT 1980; Connor et al. 1981; El-Sharkawy et al. 1992).
Cassava is relatively drought resistant (Onwueme 1978; Cock 1985). A good proportion of the crop is grown in areas with less than 750 mm of annual rainfall. Evidence that cultivation has increased in recent years in the savanna and semiarid areas in Nigeria, Tanzania, and northern Ghana, suggests that it may have an increasingly important role to play as a reliable source of staple food supplies (Berry, 1993). Only a few studies have examined how cassava copes with drought (Connor et al. 1981; El-Sharkawy et al. 1984; Baker et al. 1989).
Global warming due to greenhouse gases can also change the variability of climate. These changes are effected through climate-related parameters which include temperature, precipitation, soil moisture, and evapotranspiration. The degree to which climate variability occurs is still uncertain but potentially serious changes have been identified in some regions. These include events of extremely higher temperature, floods, and droughts with associated bush fires, pest outbreaks, ecosystem composition, and primary productivity. Consequently, there is need to research the cassava plant’s response to environments while also identifying the indices that will enable the crop to cope well with changes in the environment in the very near future.
International Institute of Tropical Agriculture (IITA) has some selections which are stay-green varieties with the ability to retain leaves during the dry season when many cassava varieties would have shed their leaves. This present research attempts to find out morphological and physiological characteristics which cause the stay-green characteristics and to relate them to yield.
The expectation from the result is to inform breeders in developing genotypes with improved drought resistance leading to increase food production and sustainable agriculture in the drier areas.
Materials and methods
Thirty stay-green cassava genotypes were evaluated in a randomized complete block design with four replicates in three locations. The plot size was 36 m2 and plants were spaced at 1 m apart in six rows of 6 m each. The three locations were:
-
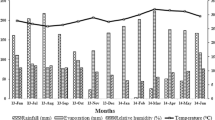
Ibadan [(Derived Savanna (DS) with Lat 7°26′ N and Long 3°54′ E, and 243 masl], with mean annual temperature (minimum (22.7–23.5) °C and maximum (22.8–34.5) °C and rainfall (1260 mm) with length of growing period of (211–270) days. The soil in this area is an Alfisol classified as Oxic Haplustalf or ferric Luvisol. There are two rainy seasons, one from March to July and the other from mid-August to November. The months of December, January and February are cold due to harmattan wind blowing across the State from the North-East (IITA, Ibadan).
-
Mokwa [(Southern Guinea Savanna (SGS) with Lat 9°29′ N and Long 5°04′ E and 152 masl], it has underlain rocks of pre-Cambrian Basement Complex with ferric Luvisol soil type. The zone has sub-humid climate with mean annual temperature (minimum (18.8–23.5) and maximum (28.8–35.8) and rainfall (1120 mm) with length of growing period of (181–201) days and about 5 months’ dry season. (National Crop Research Institute (NCRI) sub-Station, Mokwa).
-
Zaria [Northern Guinea Savanna (NGS) with Lat 11°11′ N and Long 11°78′ E and 610 masl]. The soil in the area is orthic Luvisol classified as Typic Haplustalf (USDA Soil Taxonomy as cited by Ogunwale et al. 1975) or Acrisol (FAO-UNESCO legend as cited by Valette and Ibanga 1984). Rainfall pattern is mono modal with mean annual rainfall (941 mm) and temperature (minimum (14.3–25.0) and maximum (29.1–36.6) with (151–189) days length of growing period. (Ahmadu Bello University (ABU) Teaching and Research Farm, Shika, Zaria).
Healthy stakes of 25 cm long were planted at a slanting position on 30 cm high ridges with 2/3 of the length buried in the soil. Planting was done in each location when soil moisture was sufficient to sustain establishment. Weeding was done manually using hoes and cutlasses at 1, 3, 6, and 9 months after planting (MAP). Harvesting was done at 12 months after planting (MAP).
Samples of the garri product from the genotypes were taken to the Biochemistry Laboratory for determination of pasting properties (cooking property).
Sample preparations
The dried garri samples were ground to pass through a 1-mm sieve using an analytical mill (A10 Janke & Kunkel, Staufen, Germany) and stored at 4 °C until analysis (Sanni et al. 2008).
Data collection
Physiological and growth parameters were evaluated at 3, 6, and 9 MAP using four plants selected at random within the plot as samples. The variable measured at various growth stages include leaf stomata conductance of the abaxial area, chlorophyll contents, leaf area, plant height, stem girth, number of nodes, level of branching, leaf retention ability (stay-green). Genotypes were screened for resistance to diseases (CMD: cassava mosaic, CBB: cassava bacterial blight, CAD: cassava anthracnose).
Yield parameters: fresh shoot weight per plot (kg). fresh root weight per plot (kg and t/ha) and total number of storage roots per plot.
Root size was visually scored using scale 3 to 7: 3 = small, 5 = moderately big, and 7 = very big. Dry matter percentage (DM %) of the storage root cortex was determined from a random bulk sample of four plants selected from the inner four rows. The roots were washed to remove soil and shredded. One hundred grams of fresh root cortex was taken in the form of small chips and dried at 70 °C for 72 h in a forced air oven until constant weight was achieved. The dried sample was weighed and root DM% was calculated as follows.
Percentage root cortex DM = dry weight/ Fresh weight × 100
Garri production: 10 kg of cassava roots were taken from each genotype harvested. Cassava roots were peeled, washed with water, and grated with grating machine. The pulp was put in jute bag and pressure exerted on it to remove water from the pulp. Dewatering took up to three days and as the pulp was being dewatered it was also undergoing fermentation. The pulp was sieved to remove chaff and toasted in a pot until gelatinized grains are formed.
Peak viscosity (PV): maximum viscosity developed during or soon after the heating portion of the test, in Relative Value Units (RVU). Setback viscosity (SV) from peak: final viscosity minus peak viscosity, in RVU. Final viscosity (FV): viscosity at the end of the test, in RVU. Pasting temperature (PV): Temperature at which the peak viscosity occurred in °C. Pasting time: time in minutes at which the peak viscosity occurred, in minutes.
Determination of pasting properties
The pasting properties were determined using a Rapid Visco Analyzer (RVA) (Model RVA 3D +). The RVA is a heating and cooling viscometer configured especially for starch-based products and others requiring a precise control of temperature and shear. It is ideal for determining rapidly the pasting profile of cooked starch-based samples in a controlled manner. The sample was turned into slurry by mixing 3 g with 25 mL of water inside the RVA Can. This was inserted into the tower, which was then lowered into the system. The slurry was heated from 50 to 95 °C and cooled to 50 °C within 12 min. The Can was rotated at a speed of 160 rpm with continuous stirring of the content with a plastic paddle. Results on pasting properties were obtained following the procedure explained by Sanni et al. (2008).
Statistical analysis
All data generated were analyzed using the Statistical Analytical System (SAS 1999). Analysis of variance (ANOVA) was carried out on plot mean basis, for combined ANOVA over locations and seasons and pooled as environments using the Generalized Linear Model (GLM) procedures for Randomized Complete Block Design (RCBD). All factors were considered as random. Phenotypic variances for the combination of years and locations were calculated from formulae (Falconer and Mackey 1996). Principal Components Analysis (PCA) was obtained from genotype and genotype-by-environment (GGE) biplot analysis.
Data generated from the laboratory measurements of pasting properties were analyzed using the GLM procedure (SAS 1999). ANOVA was conducted to generate mean squares (MS) using proc core procedure of SAS.
Results
Variations in shoot and root characters within and across locations
The MS from the pooled ANOVA for shoot, root, and yield traits of 30 stay-green genotypes across three locations in two seasons are presented in Table 1. The MS for year were highly significant for all characters (P < 0.001) except for number of roots, harvest index (HI), and dry matter (DM). For locations, the MS was not significant for DM but highly significant for all traits (P < 0.01). Concomitantly, the MS for the environments (location × year) were also significant for all those shoots, root and yield related, except for garri weight. The MS for genotypes were highly significant (P < 0.001). All the traits showed significant genotype × location MS except for DM. For genotype × year interaction MS, plant height, garri weight, shoot weight, and number of roots were significant (P < 0.05) while root size was highly significant (P < 0.001). The genotype × year × location MS component was also significant for all traits except for DM and root size.
Table 2 presents the pooled analysis of variance for morphological traits of 30 stay-green genotypes across three locations and two seasons. The MS for year were highly significant (P < 0.0001) for all the traits. The MS of location was significant for all traits except leaf area. Genotypes, however, showed significant MS only for chlorophyll content. The MS for genotype × location was also significant (P < 0.001) for all traits except for stem girth. The MS for genotype × year component was all highly significant (P < 0.01). The three-order interactions (genotype × location × year) were also highly significant for all traits except for stem girth.
Morphological and physiological variabilities among cassava genotypes for traits for 2 years in three locations
The mean performance of 30 cassava genotypes in relation to 19 morphological and physiological traits in three locations evaluated for 2 years is shown in Table 3.
Tested genotypes performed better in Ibadan in respect to level of branching, chlorophyll, plant height, number of leaves, gari weight, CBB resistance, fresh root yield, harvest index. Mokwa is favorable to tested genotypes in relation to stomata, stem girth, number of nodes, cad resistance, shoot weight and number of roots. Genotypes respond better in Zaria in relation to leaf area, CMD resistance, stay green and dry matter content while genotypes respond the same way in Ibadan and Mokwa in respect to root size.
Table 4 shows overall mean of selected morphological and physiological traits performance of 30 cassava genotypes evaluated for 2 years in three locations in Nigeria. 90.3% of tested genotypes recorded LAI value above grand mean of 0.01 with 930267 had the highest value of 0.032. More than half of the tested genotypes recorded mean value above grand mean for stem girth (73.3%), chlorophyll content (60%), number of node (56.7%) and stomata conductance (53.3%) with genotype 30572 (8.6 cm), 920289 (40.0), 980510 (50.5) and 950166 (144.6 mol m−2 s−1) recorded highest mean value, respectively. Half of the tested genotypes had Crop Growth Rate (CGR) and Stay Green (SG) value above grand mean of 0.29 and 2.76 with genotype 92B00068 (0.710) and I0040 (2.2) recorded highest value, respectively. Less than half of the 30 genotypes recorded mean value above grand mean for number of leaf (16.7%) with genotype 30572 (49.3) recorded the highest mean value across locations.
Yield and yield related traits of 30 cassava genotypes tested in three locations in Nigeria for 2 years is shown in Table 5. The mean plant height across locations ranged from 132.4 cm for G4 to 86.1 cm for G25 (MH950414). Ibadan recorded the highest mean plant height (134.1 cm) which was 14.2% higher than that of Mokwa and 48.9% higher than value obtained in Zaria. Across the locations, the mean fresh root yield ranged from 24.3 t/ha for G19 to 13.5 t/ha for G30. In 2010, Ibadan recorded the highest mean yield (35.1 t/ha) whereas in 2011, Mokwa recorded the highest mean yield (23.3 t/ha) and Zaria recorded the lowest (4.0 t/ha).
Across locations the mean DM ranged from 35.2% in G24 to 24.10% in G4 which was a yellow fleshed variety. Mean dry matter in Zaria was 12.4% higher than that in Ibadan and 21% higher than in Mokwa. Across locations, the mean gari weight ranged from 1.2 to 1.9 kg and 43% of the genotypes had mean value below the recorded average gari weight across locations. Ibadan recorded the highest mean gari weight (2.1 kg) followed by Zaria (1.5 kg), and Mokwa (0.9 kg) (Table 5).
The mean number of roots ranged from 95.0 for G3 and G5 to 53.0 for G21. The trend of number of roots across the three locations was Mokwa (97.7), Ibadan (96.8), and Zaria (42.4). Across the locations, the mean root size ranged from 6.8 (big) in G19 to 4.3 (small) in G14. The biggest root size of 6.43 was recorded at Mokwa in 2010; the smallest root size of 4.34 was recorded in Zaria in 2011.
The mean harvest index (HI) across locations ranged from 0.60 for G12 to 0.42 for G4. 43.3% of the genotypes evaluated across the three locations recorded mean HI above the across-location mean. The mean shoot weight across locations ranged from 34.3 kg in G19 to 20.0 kg for G30. At Ibadan, Mokwa and Zaria G8, G22 (46.9 kg/plot) and G19 (19.7 kg/plot) recorded highest mean shoot weight (Table 5).
Across the locations, mean CMD score ranged from 1.10 for G21 to 3.10 for G13 (91934). At Mokwa, the mean score for CMD was higher (1.6) in 2010 (1.6) than in 2011 (1.10). Zaria recorded the same lowest mean CMD score (1.1) in both years. Across locations, mean CBB score ranged from 1.60 (G14) to 2.62 (G30). Among all the six environments, Ibadan in 2011 recorded the lowest mean score for CBB (1.40) whereas Zaria recorded the highest mean CBB score of 2.70 (Table 5).
The correlation coefficients for traits measured over the years and locations are presented in Table 6. Most of the traits show significant correlation inter se. Notably, fresh root yield was positively correlated (P < 0.001) with leaf number (0.46), gari weight (0.23), HI (0.43), shoot weight (0.79), number of roots (0.77) and root size (0.43). It is also positively correlated with chlorophyll content (0.25), stem girth (0.24), plant height (0.49), number of nodes (0.44), and CMD (0.13). Fresh root yield was however negatively related (P < 0.01) to mean scores of CBB (− 0.28) and CAD (− 0.29).
Chlorophyll content was significantly and positively correlated with stem girth (0.19), plant height (0.23), node number (0.11), fresh root yield (0.25), shoot weight (0.19), and root size (0.11) but negatively correlated with stomata (− 0.15), garri weight (− 0.18), and DM (− 0.14). Stomata were significantly and positively correlated (P < 0.001), with node number (0.55), garri weight (0.26), and root size (0.12), but had a significantly negative correlation with stem girth (− 0.11), leaf number (− 0.32), HI (− 0.11), and number of roots (− 0.15).
In Table 6 Plant height was significantly and positively correlated with node number (0.41), leaf number (0.68), garri weight (0.18), fresh root yield (0.49), H1 (0.09), shoot weight (0.54), number of roots (0.37), and root size (0.24) but significantly and negatively correlated with DM (− 0.25). Node number was significantly and positively correlated with fresh root yield (0.44), shoot weight (0.56), number of roots (0.33), and root size (0.22), but significantly and negatively correlated with leaf number (0.17), H1 (− 0.08), and DM (− 0.19).
Leaf area was positively correlated with garri weight (0.18) and significant, fresh root yield (0.09), and shoot weight (0.31), but significantly and negatively correlated with leaf number (− 0.22). Shoot weight was also notably significantly and positively correlated (P < 0.001) with stem girth (0.55), chlorophyll contents (0.19), plant height (0.56), node number (0.31), leaf area (0.26) and fresh root yield (0.79) but had a significantly negative correlation with DM (− 0.17). Number of roots (0.77) and root size (0.43), chlorophyll content (0.25), stem girth (0.24), plant height (0.49), number of nodes (0.44), and CMD (0.13) but negatively correlated to CBB (− 0.28) and CAD (− 0.29) mean scores. Chlorophyll content correlated (P < 0.001) with stem girth (0.19), plant height (0.23), node number (0.11), shoot weight (0.19), and root size (0.11) but negatively correlated with stomata (− 0.15), gari weight (− 0.18), and DM (− 0.14). Stomata correlated (P < 0.001), with node number (0.55), gari weight (0.26), and root size (0.12) but had a significantly negative correlation with stem girth (− 0.11), leaf number (− 0.32), HI (− 0.11), and number of roots (− 0.15). Plant height (P < 0.01) correlated with node number (0.41), leaf number (0.68), gari weight (0.18), H1 (0.09), shoot weight (0.54), number of roots (0.37), and root size (0.24) but negatively correlated with DM (− 0.25). Node number correlated with shoot weight (0.56), number of roots (0.33), and root size (0.22) but negatively correlated with leaf number (0.17), H1 (− 0.08), and DM (− 0.19). Leaf area correlated with gari weight (0.18) and shoot weight (0.31) negatively correlated with leaf number (− 0.22). Shoot weight correlated (P < 0.001) with stem girth (0.55) but had a negative correlation with DM (− 0.17) (Table 6).
Genotype by Environment Interaction (G × E) analysis of genotypes for root yield using AMMI model analysis
The AMMI 1 analyses for fresh root weight and DM are presented (Figs. 1, 2). Cassava production per plant was significantly (P < 0.01) influenced by treatment effect as contained in the genotype, environment, and interaction components and these jointly captured 95.8% of the Total Sum of Squares (TSS) for fresh root weight and 88.1% for DM. The interaction and the genotype portions were 5.08 and 4.7% for fresh root yield and 16.12 and 7.8% for DM. Principal component axes 1 and 2 were significant (P < 0.0001 and 0.001). The two axes jointly explained significant of the interaction sum of squares for fresh root weight and DM (63.91% and 67.17%) (Figs. 1, 2) leaving a non-significant of 26.09% and 32.83% in the residual (Tables 7, 8). The mean squares (MS) from the analysis represented the variance of components and captured 88.05% and 64.22% of the variation with environment for fresh root weight and DM accounting for the largest portion, 95.8% and 88.1% of the total variation for fresh root weight and DM respectively. The genotype main effects had 5.08% and 16.05% of the variation for fresh root weight and DM (Figs. 1, 2). Genotypes or environments on the same parallel line relative to the y-axis have similar yields, and a genotype or environment on the right side of the midpoint of this axis has higher yields than those on the left. For DM the interaction accounted for 85.7% of the total sum of squares due to treatments; genotypes accounted for 12% while environment captured 2.3%.

Biplot of first AMMI interaction-IPC1 (Stability Analysis) scores (Y-axis) plotted against fresh root yield (x-axis) for 30 cassava genotypes in 6 environments (3 × 2) in Nigeria

Biplot of first AMMI interaction-IPC1 (Stability Analysis) scores (Y-axis) plotted against fresh dry matter (x-axis) for 30 cassava genotypes in 6 environments (3 × 2) in Nigeria
Genotype compatibility to different environments
From Fig. 1 the environments of Zaria in 2010 and 2011 and of Mokwa in 2010 were similar in having lower main effects (yield) and negative IPC1. Genotypes 2, 17, 29, and 30 were grouped along with the environments; G7 was grouped along with Mokwa environment in 2011 with both having high main effects and positive but near zero IPC1. Genotypes 6, 21, and 9 had negative IPC1 and below average main effect; G23 and G25 had main effects and IPC values closest to the average main effect and zero IPC interaction; G3 and G26 had above average main effect and near zero IPC1.The remaining genotypes and environments clustered together within negative IPC1 and above-average main effect.
Genotype × Environment (G × E) and Stability Analysis of 30 cassava genotypes for fresh root and DM using the Genotype and Genotype × Environment (GGE) model
Figure 3 presents the GGE-Biplot polygon for cassava root yield, showing related genotypes for each trait across environments. For the fresh root yield, seven rays divide the Biplot into five sectors in which all the environments fell except one and the vertex genotype for this sector was 19 (950289), The identified sector with G19 as vertex genotype also include G7, G4, G3 G16, G14, G22, G13; the highest yielding genotype was G23. Results showed that no environment fell into sectors with G9, G6, G29, and G30 as the vertices, indicating that these genotypes were not the best in terms of yield in any environment.

GGE Biplot for best genotypes (G) in term of fresh root yield in different environments. IB10 = Ibadan 2010, IB11 = Ibadan 2011; MK10 = Mokwa 2010, MK11 = Mokwa 2011; ZA10 = Zaria 2010, ZA11 = Zaria 2011
For dry matter (Fig. 4) even though, there were five sectors in all, only one mega- environment was identified. The identified sector with G24 as the vertex genotype also includes G3, G14, G22, G20, G1, G7, G25. No environments fell in the sector with G29, G16, G27; G12, G15, G5, G26 G8, G9, G2; G6, G18, G4, G19, G13, G21 and G23, G10, G28.as vertex cultivars.

GGE Biplot for best genotypes (G) in term of dry matter in different environment. IB10 = Ibadan 2010, IB11 = Ibadan 2011; MK10 = Mokwa 2010, MK11 = Mokwa 2011; ZA10 = Zaria 2010, ZA11 = Zaria 2011
Table 9 shows the pasting properties of 30 stay-green clones planted in 2010 and 2011 at different agro-ecological locations (Ibadan, Mokwa, and Zaria).
The table presents the mean pasting properties by location. There were significant (P < 0.05) genotypic variations among the genotypes for all variables except trough and pasting temperature. The effect of the environment (locations) on the pasting parameters was significant (P < 0.05) in both years. The variations due to interactions between clone and location (clones × location) were significant (P < 0.05) except for pasting temperature.
Peak viscosity which is the maximum attainable during the heating cycle ranged between 130.3 RVU (MM972480) and 294.6 RVU (950166) in 2010 and 129.3 RVU (MM972480) and 439.3 RVU (011807) in 2011. The trough values ranged between 101.1 RVU (MM972480) and 189.6 RVU (011807) in 2010 and from 98.0 RVU (MM972480) to 385.6 RVU (011807) in 2011. Trough or hot paste viscosity is the minimum viscosity value in the constant temperature phase of the RVA profile and is an index of starch granule stability to heating. Breakdown viscosity ranged from 23.6 RVU (TMEB-1) to 106.6 RVU (MH950414) in 2010 and from 15.5 RVU (011663) to 115.7 RVU (950166) in 2011. The setback viscosity which is an index of the retrogradation of linear starch molecules during cooling ranged between 42.6 RVU (MM972480) and 81.4 RVU (020431) in 2010 and from 37.3 RVU (011807) to 77.9 RVU (011663) in 2011. Mean final viscosity across clones ranged from 143.7 RVU (MM972480) to 270.9 RVU (020431) in 2010 and from 141.3 RVU (MM972480) to 422.9 RVU (011807) in 2011.
Time to attain peak viscosity ranged from 4.4 min (MH950414) to 6.6 min (011663) in 2010 and from 3.8 min (011807) to 6.7 min (011663) in 2011. Pasting temperature (PT) of different cassava starches ranged from 47.7 °C (9102322) to 50.0 °C (011807) in 2010 and from 48.2 °C (010040 and 9102324) to 57.5 °C (011807) in 2011 (Table 10).
Discussion
The significant mean squares for most of the root shoot and yield traits confirm appreciable variations among the 30 cassava genotypes used in the study. This is indicative of possibility of evolution of further improved types that can replace the available cultivars. The non- significant effect of year, location, and some of their interactions for harvest index, dry matter, garri weight and a few root traits can be a direct consequence of the similarity in these traits over the years and locations, even with the inherent differences in the environmental indices, particularly moisture. Cassava being a fairly drought tolerant crop would still exhibit fairly stable performance where rainfall is not really limiting. It would appear that studies would still have to be done to determine the level of moisture difference that would elicit significant variation in the performance cassava, particularly with respect to harvest index and dry matter.
The significant morphological traits for the environments (as derived from location, year, and their interaction) showed ample variation that can be stimulated from the genotypes. The additional significant genotypic effect points to indicated genotypic differences for the traits and the possibility of selection for adaptation to specific environments. These observations are consistent with those of Aina et al. (2009) had observed similar results when 18 genotypes were evaluated across four locations in Nigeria. This consistency further prompts the need for further recombination of genes to select higher performing cultivars.
The high variability observed among genotypes, as indicated by the range of their mean performance, indicates the presence of sufficient genetic variability for the traits studied. Indeed, the high variation that was observed for storage root yield across the different environments in this study indicates that these environments differ greatly in environmental conditions categorized as different agroecological zones that have been reflected in each location. In this study the highest storage root yield at Ibadan (DS), while the lowest at Zaria (NGS) indicates that in some environmental locations there are more conditions that favor yield in cassava than in others. The higher storage root yields recorded at Ibadan and Mokwa were suggestive of their favorable climatic conditions while the lower root yield recorded at Zaria reflects the harsh climatic conditions evident in this location.
The significant interaction between genotype, location, and year (environment) (G × Y × L) for most traits supports the need for multilocational testing to identify good performers for specific locations. The significance of the year effects however alludes to the unpredictability of the Cassava growing seasons in Nigeria and suggests the need to evaluate for more than one year for reliable inferences to be made on performance (Semakula and Dixon 2007) within the overall focus of the development of stable cultivars in terms of morphological, yield and yield traits.
Cassava ability to serve as a drought resistant crop was also demonstrated in this study with root yield as high as 8.0 t/ha obtained in Zaria, a zone where annual precipitation is as low as 840 mm, and yet cassava could still produce an appreciable root yield, justifying it as a drought tolerant crop. The prolonged dry season enhances infestation of cassava fields by termites that eat up both the stems and roots. The termites bore into the stems and suck the sap content, thereby destroying the plant, reducing the number of stands at harvest, and resulting in a decreased final tuber yield. Therefore, moisture stress as observed by Okogbenin et al. (1999), was responsible directly or indirectly, for the decline in storage root yield observed at Zaria.
The high storage root yield obtained in Ibadan, however, was not indicative that the plants’ maximum genetic potential had been exhausted. Therefore, it is possible for the root yield to increase beyond the values that were obtained because cassava, like other crops, has the ability to thrive under favorable conditions when there are no major production constraints (Howeler and Cadavid, 1990).
The different locations favored the expression of different traits. However, the Ibadan environment appeared to have a combination of traits that eventually translated into higher harvest index and root yield. The overall higher shoot weight in Mokwa was largely accounted for by the relatively larger stem girth and this failed to translate into equally highest root yield. The Zaria environment with high insolation and limited moisture favored larger leaf area and stay green ability but this did not culminate in high root storage as a lot of the photosynthates were probably devolved into maintaining the vegetative part of the plants.
The high LAI, chlorophyll content and stomata conductance recorded by the varieties presents opportunity for advantageous selection and plant breeding efforts for further improvement. The ultimate focus in cassava production is the root harvest. The conditions that favor higher yield in Ibadan and the varieties that expressed the highest yield and related traits require pointed attention for further study and improvement. The complex interaction of environment and disease (CBB, CMD) development has to be concomitantly incorporated into such efforts. Egesi et al. (2007) had reported that low yielding cassava genotypes are related to high scores of the disease whereas high root yielders are not associated. Ibadan with higher moisture during growth had the least CBB and this may be due to the ability of the plants to take advantage of the more conducive growing condition and cope with the disease unlike the relatively poor growing conditions in the other locations.
The complex trait correlation obtained among several traits points to the need to focus more on root yield (and garri weight). Quite a number of the traits are correlated inter se and would be inherited depending on the direction of selection and breeding. High root yield for instance root size and garri weight are higher in varieties with lower chlorophyll content as the latter favors vegetative growth to the detriment of high tuber yield. Varieties that are susceptible to CBB and CAD also had reduced yield since leaf number and photosynthetic area are both affected.
Yield stability
Stable genotypes would promote good yields in both favorable and unfavorable environments while adaptable genotypes would increase economic gains and promote the improvement of such genotypes in specific environments. Results in this study showed that the performances of genotypes with respect to storage root yield differed across environments, indicating the presence of genotype by environment (g x e) interaction.
The GGE Biplot analysis enabled visual comparison of the locations and genotypes studied and their interrelationships. The portioning of GGE Biplot analysis of the fresh root yield showed that the first principal component interaction (PC 1) accounted for 55.8% of GGE sum of squares and the second principal component interaction (PC 2) accounted for 18.2%, explaining a total of 74% variation. The presence of GEI was emphasized by different yield performance among genotypes across the tested environments. A study by Egesi et al. (2007) on Cassava varieties in three locations within a relatively wet region in Nigeria had also separated genotypes based on location compatibility. The seven rays divide the Biplot into seven sectors into which all the environments fall except one and the vertex genotypes for this sector was G19 (950289), the genotype with highest yield. No environment fell into sectors with G9, G6, G29, and G30 as the vertices. These genotypes according to Yan et al (2000, 2001) are not the best in term of yield in any of the three locations in Nigeria.
Choice of genotypes
According to Yan and Tinker (2006), genotype with highest mean performance in the tested environments and is also stable is regarded as an ideal genotype and can also be used as a reference for genotypic evaluation. A genotype is more desirable if it is located closer to the ideal or farthest away from the standard, but lowest yielder compared with other genotypes. The cosine of the angle between the vectors of two genotypes also measures their similarity or dissimilarity in response to their interaction with the environments. In this consideration, G19 (TMS 950289) which fell farthest away from the least genotype (G30-TME1) was the ideal genotype with highest mean performance in the tested environments and stable compared with the rest of the genotypes. In addition, G7 (011807), G22 (980581), G16 (92B00068), G4 (010040), and G12 (9102322), located next may be regarded as desirable genotypes.
Mean performance and stability of genotypes
Following the direction of Yan and Raccan (2002), genotypes should be evaluated based on both mean performance and stability across environments. Therefore, the most stable genotypes in terms of fresh root yield were G19 and G7. Others with stable but had poor root yield were G16, G3, G26, G14, and G22. G28,. Additionally, G27. G29, G9, G17 were low yielding and unstable, but contributed significantly to the GEI.
Discriminating ability and representativeness of test environment
Discriminating power and representativeness view of the GGE-Biplot could be used to measure the testing environment (Dehghani et al. 2006). In this study, the GGE Biplot explained 78.3% of the G + GE data suggesting that the angles between the vectors of the environments are good indicators of correlation among the environments. For instance, the Mokwa environment in 2010 was strongly correlated with Zaria in the same year also Mokwa and Ibadan in 2011 were strongly correlated whereas Zaria in 2011 and Ibadan in 2010 were weakly correlated. The cosine of the angles between the vectors of Mokwa 2010 and Zaria 2010, also Mokwa 2011 and Ibadan 2011, were less than 90 Zaria’s environment in 2011 had the longest vector and was therefore the most discriminating and the least representative environment while the Mokwa and Ibadan locations in 2011 were the most representative and least discriminating. The Mokwa environment, which was averagely discriminating and the most representative, can therefore be used for genotypes with wide adaptation. The non-representative environments such as Zaria and Ibadan are useful for selecting specifically adapted genotypes. Evaluation of genotypes should be based on both mean performance and stability across environments (Yan and Rajcan 2002).
The additive main effect and multiplicative interaction (AMMI) model which can effectively handle the main effect and their interaction and identify stable and adaptable genotypes was therefore used in this study. G19 (980289) had the highest mean yield, followed by G7 (011807), and then G16 (92B00068), G4 (010040) and G22 (980581) that had mean yields similar to the grand mean while G30 (TME 1) was the lowest. In line with Yan and Tinker 2006 findings, G19 (980289), G7 (011807), G16 (92B00068), G4 (010040), and G22 (980581) were the highest yielding stable genotypes while G24, G5, and G27 were equally stable but with poor yield. G8, though high yielding, was highly unstable while G29 was both low yielding and unstable. Dixon and Nukeine (1997) and Ntawuruhunga et al. (2001) also engaged the use of the AMMI model. Result showed a high g x e interaction as a result of wide differences among the environments used in this study. This was in line with the findings of Ceccarelli (1996) and Annicchiarico (2000). The high influence of the environment on root yield was reflected in the proportion of the total variation due to environmental effects that exceeded that of the genotype and the g x e effect. In this study, five genotypes (950289, 010034, 990554, 011807, and 980581) had negligible interactions with the environment and have broad adaptation and were considered highly stable.
Large environmental interactions, however, resulted in location specificity and narrow adaptation; and genotype 011807 was specifically adapted to Zaria. Some genotypes were found to show specific adaptation to some environments, indicating that these would perform best in such locations, while others with high stability across environments (950289) showed that such varieties could be relied upon to give economic yields across all environments considered. The highest root yield of 35.1 t/ha observed at Ibadan in this study was below the maximum yield capacity of the crop that ranges between 25 and 45 t/ha (FAO 1999). Although the highest storage root yield was obtained in Ibadan, the maximum genetic potential of the plant in terms of yield has not been realized as soil analysis results revealed in all the locations.
Peak viscosity indicates the water-binding capacity of the starch. It is often correlated with the final product quality and also provides an indication of the viscous load likely to be encountered during mixing. The peak time is a measure of the cooking time. Pasting temperature has been described as the temperature above the gelatinization temperature when starch granules begin to swell and is characterized by an increase in viscosity on shearing. The period is sometimes called trough or hot paste viscosity. Following this, the rate of starch breakdown depends on the nature of the material, the temperature, and the degree of mixing and shear applied to the mixture. The ability of a mixture to withstand heating and the shear stress that is usually encountered during processing is an important factor for many processes, especially those requiring stable paste and low retrogradation/ senescence. Setback has been correlated with the texture of various products. High setback is also associated with syneresis or weeping during freeze/thaw cycles. Sanni et al. (2004) reported that lower setback during the cooling of paste from starch or a starch-based food indicates greater resistance to retrogradation. Final viscosity has been reported as the most commonly used parameter to characterize the ability of starch-based material to form a viscous paste or gel after cooking and cooling the resistance of the paste as well to shear force during stirring (Sanni et al. 2004).
From the above, we realized that apart from pasting temperature, that was preferred at the low temperature and peak time which are applicable to this study, the other pasting properties such as peak viscosity, trough, breakdown, final viscosity, and setback were preferred at high viscosity especially for household products such eba and fufu.
Ten out of the thirty clones studied were found to have higher pasting properties at a low temperature and peak time that are higher than the old improved (30572) and the local variety (TME1) used in this study. These clones are 950166, 92B00068, 020431, 011807, 9102324, 010040, 950289, 990554, 980510, and 98058. Two of them–011807 and 950166 –were found to be the best.
Conclusions and recommendations
Cassava genotypes performed differently across and within locations for all the trials evaluated, an indication of wide variability among them for morphological, physiological, and root characteristics. AMMI analysis was able to identify stable and adaptable genotypes across the different environments. Five genotypes—950289, 010034, 990554, 011807, and 980581 were identified as stable across different environments and combined a high garri weight. These genotypes could be recommended to farmers for stable yield potentials across different locations. The range of values obtained in the pasting profile of the roots studied is indicative of some level in genetic variations existing in the clones analyzed. By implication, some level of improvement could still be achieved for this trait. The presence of clone × agro-environment effect. However, necessitates the need to couple clone × environment interaction into the efforts to improve clones for this variable. This would ensure consistent performance over fairly wide cultivation zones. Two clones (011807 and 950166) were found to be the best for pasting properties. However, further work is needed to ascertain effect of pasting profile on food quality of these clones. Overall, selection strategies that consider root size alongside total root number, harvest index, dry matter, disease score, as well as wide applications of genotypes for household food and industrial uses will be an effective and efficient way to select for high yielding genotypes.
Data availability
All data are fully available.
Change history
26 May 2021
A Correction to this paper has been published: https://doi.org/10.1007/s10681-021-02855-3
References
Ahmadu Bello University (ABU) Teaching and Research Farm, Shika weather station, Zaria, Kaduna state, Nigeria, 2010/11 planting season weather data
Aina OO, Dixon AGO, Paul I, Akinrinde EA (2009) GxE interaction effects on components of cassava (land races and improved) genotypes in the savanna region of Nigeria. Agron J 82:887–891
Annicchiarico P (2000) Variety × location interaction and its implications on breeding of Lucerne a case study. In: Proc XIII Euphoria Medicago spp Univ. of Perugia Group Meeting 35–43
Anon (2000) Cassava can play a key role in reducing hunger and poverty. A press release on the International Forum on the tropical root crop. Organized by International Fund for Agricultural Development (IFAD) and the Food and Agriculture Organization (FAO),26–28
Baker GR, Fukai S, Wilson GL (1989) The response of cassava to water deficits at various stages of growth in the subtropics. Aust J Agric Res 40:517–528
Berry SS (1993) Socio-economic Aspects of Cassava Cultivation and use in Africa Implications for the development of appropriate technology. International Institute of Tropical Agriculture, Ibadan, Nigeria
Blum A (1993) Selection for sustained production in water-deficit environments. Internat. Crop. Sci. Soc. Am, Madison
Ceccarelli S (1996) Positive interpretation of genotype by environment interaction in relation to sustainability and biodiversity. In: Cooper M, Hammer GL (eds) Plant adaptation and crop improvement. Wallingford CABI, pp 467–486
Ceccarelli S, Grando S (1991) selection environment and environmental sensitivity in barley. Euphytica 57:157–167
CIAT (1980) Cassava Programme. Annual report 1979 centro internacional de agricultura tropical, AA 67–13 Cali Colombia
Cock JH (1985) Cassava New Potential for a Neglected Crop. Westview Press, IADS Publication. Colorado USA, p 192
Connor DJ, Cock JH (1981) The response of cassava to water shortage II Canopy dynamics. Field Crops Res 4:285–296
Connor DJ, Cock JH, Parra GE (1981) The response of cassava to water shortage. I growth and yield. Field Crops Res 4:181–200
Dehghani H, Ebadi A, Yousefi A (2006) Biplot analysis Genotype x Environment interaction for barley yield in Iran. Agron J 98:388–393
Dixon AGO, Nukeine EN (1997) Statistical analysis of cassava yield trials with the additive main effects and multiplicative interaction (AMMI) model. Afr J root tuber crop 3(1):46–450
Dixon AGO, OnyekaR. Bandy opadhyay, R.U. Okechukwu, and B. Bamkefa, TJ (2005) Distribution and status of bacterial blight and fungal diseases of cassava in Nigeria IITA. Ibadan Nigeria 2005:1–2p
Egesi CN, Ilona P, Ogbe FO, Akoroda DA (2007) Genetic Variation and Genotype ×Environment interaction for yield and other Agronomic Traits in Cassava in Nigeria. Soil Sci J Am 99(4):1137–1142
El-Sharkawy MA (1993) Drought tolerant cassava for Africa. Asia Lat Am BioSci 47(7):441–453
El-Sharkawy M, Cock JH, Cadena GD (1984) Stomata characteristics among cassava cultivars and their relation to gas exchange. Expl Agric 20:67–76
El-Sharkawy MA, Hernandez ADP, Hershey C (1992) Yield stability of cassava during prolonged mid-season water stress. Exp Agric 28:165–174
Falconer DS, Mackey T (1996) Introduction to quantitative genetics, 4th edn. Longman scientific and technical Co., Essex, p 464p
FAO (1999) FAOSTAT Database Collections. Web page, (accessed 1999)
FAOSTAT (2013) Food and Agriculture Organization of United Nations, Rome Italy
Hahn SK, JCG Isoba JCG, Ikotun T (1989) Resistance breeding in root and tuber crops at International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria. Crop Protection (p 147–168)
Howeler RH, Cadavid LF (1990) Short- and long-term fertility trials in Colombia to determine the nutrient requirements of cassava. Fertil Res 26:61–80
Ihenkoronye AI and Ngoddy PO (1985) Integrated Food Science and technology for the Tropics. London Macmillan Publisher Ltd; 1985. Tropical roots and tuber crops; (pp. 384–9)
IITA (International Institute of Tropical Agriculture) (2005) Integrated cassava project.20 cassava, the multipurpose crop flyer Designated and printed at IITA Ibadan Nigeria. 1p
IITA (International Institute of Tropical Agriculture) (1998) Annual report on cassava productivity in the lowland and mid-altitude agroecologies of sub-Saharan Africa. Project 14:2
IITA (International Institute of Tropical Agriculture (IITA) (2010) GIS Unit, Headquarter, Ibadan, Oyo state, Nigeria, 2010/11 planting season weather data
Iwuoha CI, Eke OS (1996) Nigerian indigenous fermented food, their traditional process operation, inherent problems, improvement, and current status. Food Res Int 29(5–6):527–540
Kang MS, Magari R (1995) Stable a BASICA program for calculating stability and yield- stability statistics. Agron J 87:276–277
Manjit KS (1993) Simultaneous selection for yield and stability in crop performance trilas: consequences for growers. Agron J 85(3):754–757
National Crop Research Institute (NCRI) sub-station weather station, Mokwa, Niger state, Nigeria, 2010/11 planting season weather data
Ntawuruhunga PPR, Rubaihaho JBA, WhyteDixon, and D.S.O. Sir, AGO (2001) Inter relationships among traits and path analysis for yield components of cassava A search for storage root yield indicators. Afr Crop Sci J 9(4):599–606
Nweke FI, Spencer DSC, Lyman JK (2002) The Cassava Transformation Africa’s best-kept secret. Michigan State University Press, East Lansing, p 273
Ogunwale JA and Ashaye TI (Soil Survey Staff. 1975) Soil taxonomy: A basic system of soil classification for making and interpreting soil surveys. Agric. Handb. 436. SCS, USDA. US Gov. Print. Office, Washington, DC.1975; 745
Okogbenin E, Ekanayake I, Porto MCM (1999) Effect of planting materials and soil moisture on cassava performance in the semi-arid Sudan Savannah belt of Nigeria. Afr Crop Sci J 1:21–33
Onwueme IC (1978) The tropical tuber crops. Wiley, Chichester, p 234p
SAS (1999) Statistical Analysis System (SAS) Institute Incorporate. Statistical Analysis System, software version 9.3; SAS: Cary, NC, USA, 2011.
Sanni LO, Kosoko SB, Adebowale AA, Adeoye RJ (2004) The influence of palm oil and chemical modification on the pasting and sensory properties of fufu flour. Int J Food Prop 7(2):229–237
Sanni LOAA, Adebowale B-D, Dixon AGO (2008) Chemical composition and pasting properties of CMD resistant cassava clones planted at different locations. J Food Agric Environ 6(2):97–104
Ssemakula G, Dixon A (2007) Genotype × environment interaction, stability and agronomic performance of carotenoid-rich cassava clones sci. Res Essay 2(9):390–399
Valette JA, Ibanga IJ (1984) The detailed soil survey of the experimental farm of the Institute for agricultural research farm, Samaru, Zaria, Nigeria. Soil Survey Bull. Ahmadu Bello University, Zaria, Nigeria
Yan W, Rajcan I (2002) Biplot analysis of test sites and trait relations of soybean in Ontario. Crop Sci 42:11–20
Yan W, Tinker NA (2006) Biplot analysis of multi-environment trial data: principles and applications. Can J Plant Sci 86:623–645
Yan W, Hunt LA, Sheng Q, Szlavnics Z (2000) Cultivar evaluation and mega- environment investigation based on the GGE biplot. Crop Sci 40:597–605
Yan W, Cornelius PL, Crossa J, Hunt LA (2001) Two types of GGE biplot for analyzing multi-environment trial data. Crop Sci 41:656–663
Acknowledgements
The authors thank the UK’s Foreign, Commonwealth & Development Office (FCDO) and the Bill & Melinda Gates Foundation (Grant INV-007637 http://www.gatesfoundation.org) for their financial support. Also the Cassava Breeding and Biochemistry units of the International Institute of Tropical Agriculture, Ibadan, Nigeria.
Funding
The research was financially supported by the UK’s Foreign, Commonwealth & Development Office (FCDO) and the Bill & Melinda Gates Foundation (Grant INV-007637 https://www.gatesfoundation.org).
Author information
Authors and Affiliations
Contributions
ANA was principal investigator, collected data and fully involved in the writing of the article, OOO and NAL supervised in 2. AA was responsible for statistical data analysis; PK is head cassava breeding unit. PE Cassava Breeder in the unit. OSA, AMG, AOO proofread; SLO did the review. M-D, B Head of Biochemistry unit; DA planned and facilitate the project as the Head of the unit at the onset of the project. Each author had participated sufficiently in the work to take public responsibility for appropriate portions of the content. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of Interest
The authors have not declared any conflict of interest.
Ethical approval and consent to participate
All experimental protocols were approved.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original version of this article was revised: the funding information and the acknowledgement section was incomplete.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Adetoro, N.A., Oworu, O.O., Nassir, A.L. et al. Evaluation of improved cassava genotypes for yield and related traits for a better breeding strategy under different agroecologies in Nigeria. Euphytica 217, 73 (2021). https://doi.org/10.1007/s10681-021-02798-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-021-02798-9