
Abstract
Small and isolated populations are prone to future extinctions and thus perceived as ‘living dead’. Although generally considered to be of low conservation value, their existence can still enhance species survival at the landscape scale through improving the connectivity of other populations and facilitating some (even if little) gene flow. We investigated the demography and genetic status of a tiny and highly isolated local population of Maculinea (= Phengaris) alcon near its distribution margin with the aim of identifying the features that allow it to persist. The study comprised intensive mark-recapture, surveys of Gentiana pneumonanthe foodplants and butterfly eggs laid on them, as well as genetic analyses. The population has been found to be characterised by low genetic diversity and estimated at only a few tens of individuals. The foodplant availability turned out to be the most obvious factor limiting M. alcon abundance. Nevertheless, the life expectancy of adult butterflies is fairly long, and their flight period very short, implying that most individuals occur within the same time window. Together with the relatively little protandry observed, i.e. almost synchronous emergence of males and females, this increases the chances of random mating among the individuals. Moreover, the butterflies move freely across the core habitat fragment. All things concerned, the effective population size is presumably not much lower than the recorded population size. Our findings provide guidelines for pinpointing those among ‘living dead’ populations that are likely to be the most persistent and thus worth conservation efforts aimed at preserving them.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Habitat loss and fragmentation is nowadays one of the most serious threats to biodiversity, with more and more species forced to live in increasingly fragmented landscapes (Caughley 1994; Hanski and Gaggiotti 2004). Small and highly isolated local populations experience a high risk of stochastic extinctions, and following such extinctions their habitat patches have little chance to become recolonized (Prugh et al. 2008). Below certain thresholds of area and connectivity, habitat patches have dramatically low probability of being continuously occupied (Hanski 1994). Consequently, populations inhabiting such habitat patches are perceived as ‘living dead’ (Hanski et al. 1996); even if they exist at present, they are bound to go extinct in the coming future.
It is sometimes argued that living dead populations have no conservation value, since being prone to extinctions they do not enhance directly the species persistence in fragmented landscapes (Hanski et al. 1996; Soga and Koike 2013). Nevertheless, their existence can still improve the chances of survival of other populations in metapopulation systems. First of all, as long as they exist they allow some (even if little) gene flow, thus facilitating the maintenance of genetic variability within the metapopulations. Furthermore, although due to their small size living dead populations should not be expected to produce many emigrants themselves, they may act as stepping stones, thus increasing the connectivity of other populations and the exchange of individuals among them (Haddad 1999; Skórka et al. 2013). Although habitat patches alone are needed to provide these stepping-stone effects, their occupancy is desirable for practical reason as it is simply much easier to convince decision makers that it is worth to protect certain fragments of land if they are actually inhabited by a focal species rather than if they merely constitute its habitat.
All in all, despite being dubbed as living dead, some small and relatively isolated local populations may be worth conservation efforts aimed at preserving them. The decision whether such populations deserve to be included in conservation programmes should depend on their persistence chances. Priority must be given to those are likely to persist for a long time despite their ultimate extinction risk, and not to those that face imminent extinction. According to a solid theoretical rationale, it is easy to link extinction risk with the long-term trend in population size as well as population stability, negatively associated with the extent of its year-to-year fluctuations (see the “Discussion”), but in reality its quantitative assessment through population viability analysis is problematic unless very good monitoring data are available, which is rarely the case (Schtickzelle and Baguette 2009). Consequently, a highly useful solution could be simple but reliable proxies in the form of easy to measure population characteristics which translate into viability.
In the present study, we investigated a tiny and highly isolated local population of the Maculinea (= Phengaris) alcon butterfly near its distribution margin in north-eastern Poland using intensive mark-recapture sampling and genetic analysis, with the aim of identifying the features that allow it to persist against all the odds. Based on the results obtained, we discuss their implications for the viability assessment of the investigated population. More generally, we provide guidelines for pinpointing those among ‘living dead’ populations that are likely to be the most persistent and thus worth conservation efforts.
Materials and methods
Study species
Myrmecophilous butterflies of the genus Maculinea (= Phengaris), and M. alcon among them, are considered useful indicators of grassland biodiversity and flagships of insect conservation in Europe (Maes and van Dyck 2005; Settele et al. 2005). To complete their life cycle they require two crucial resources, which comprise specific foodplants, on which Maculinea larvae feed in their initial stages, and specific host ants of the genus Myrmica, in the colonies of which the larvae finish their development, acting as social parasites (Thomas 1995; Witek et al. 2010). There is a controversy surrounding the taxonomic status of M. alcon, with its two distinct forms associated with wet meadows and xerothermic grasslands, regarded as separate species or, more frequently, merely as different ecotypes (Als et al. 2004; Pecsenye et al. 2007; Sielezniew et al. 2012; Bereczki et al. 2018). Consequently, the conservation status of the species is uncertain, and it is classified as least concern (LC) in Europe (van Swaay et al. 2010), even though many authors regard both forms to be clearly under threat (WallisDeVries 2004; Tartally et al. 2008; Czekes et al. 2014).
It should be underlined that in the present study we focus on the wet meadow form of M. alcon, which uses almost exclusively the marsh gentian Gentiana pneumonanthe as its foodplant (Nowicki et al. 2007; Sielezniew et al. 2012; Czekes et al. 2014). Unlike the foodplants of other Maculinea species, which tend to grow in high densities within relatively large meadow fragments (Bonelli et al. 2013; Nowicki et al. 2014), the marsh gentians are typically scarce and occur in highly localised places, and thus their availability constitutes a limiting factor for M. alcon abundances (Nowicki et al. 2007). Consequently, the butterfly quite often exists in very small and spatially isolated local populations, although fairly large metapopulation systems with many interconnected populations can be found as well (WallisDeVries 2004; Nowicki et al. 2007, 2009).
Study area
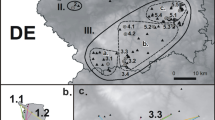
The study was carried out at the Bokiny site (N53°02′02″, E22°54′45″; 115–116 m a.s.l.), located in the Narew National Park, where M. alcon was discovered in 2012 (Sielezniew et al. 2015). The butterflies inhabit an elevated ‘grassland island’, i.e. a small patch (5.47 ha in area) of higher, drier land overlying mineral soils, surrounded by wetlands (Fig. 1). The vegetation of this area includes a mosaic of Molinio-Arrhenatheretea grasslands, with strips of light pine and aspen forest. Only some discrete fragments of the site can be regarded as the potential habitat of M. alcon as defined by the presence of its G. pneumonanthe foodplant. The core habitat fragment with the highest foodplant density is relatively small (0.32 ha in area) and situated in the southern part of the site (Fig. 1).

Source of the orthophotomap: Digital Globe (acquired by Google Earth). (Color figure online)
The distribution of M. alcon capture points (black crosses), its eggs (white dots with the size linearly corresponding the number of eggs), and G. pneumonanthe foodplants (blue/grey dots) within the investigated site. It should be noted that the numbers of capture points, eggs, and foodplants within the core southern fragment of the site are somewhat visually underestimated, because they often overlap one another.
In the past the entire site was grazed by cattle with low intensity and occasionally burnt, but both these practices were ceased in the 1990s. At the moment, only marginal low-lying parts of the site (as well as surrounding wetlands) are mown every second year in late summer. The population of M. alcon is the only one found in the Narew National Park (Sielezniew et al. 2016) and it is highly isolated, with the nearest other known localities with the species being about 50 km away (Sielezniew unpubl. data).
Field sampling
Although the population has been surveyed for several years since its discovery in 2012, the field surveys conducted were intensive enough only in 2015 to allow detailed investigations. Consequently, in the present paper we restricted our analyses to the demographic data collected in that year. Nevertheless, it must be stressed that the information gathered in other years clearly suggests that the results obtained in 2015 are representative for the situation of the focal population in recent years.
Mark-release-recapture (MRR) sampling was conducted between 12 July and 4 August 2015, covering the entire flight period of M. alcon. No butterflies were encountered during visits performed just before and after this period. Sampling was performed every day, weather permitting, between 10 AM and 5 PM. One or two people spent about 2 h on the site during each sampling day. Butterflies were captured with entomological nets, marked on the underside of their hind-wings with unique identity numbers using a fine-tipped waterproof pen, and then immediately released at the place of capture. Date, time and GPS coordinates of each (re)capture as well as sex and ID number of each butterfly were recorded.
In the same year the entire site was thoroughly searched in order to record all the G. pneumonanthe foodplants present as well as M. alcon eggs laid on them. This survey was greatly facilitated by a preliminary search performed in the previous year, when all the gentians found were marked with bamboo sticks to avoid trampling on them while conducting the MRR study, but we also looked for additional gentians. To be sure that all of them were spotted we repeated the search several times during the season so as to account for potential differences in the phenology of individual plants flowering period. Until mid-August all the gentians were marked with sticks and their position was measured with GPS. Subsequently, i.e. after the adult flight period was already well finished, all the eggs/eggshells of M. alcon were counted separately for each plant. Both the foodplant and egg count are standard methods applied in the monitoring M. alcon populations, and they allow perfect detectability (Maes et al. 2004; Nowicki 2017).
Analysis
The mark-recapture data collected were analysed using constrained open population models in the program MARK 8.0 (White and Burnham 1999) as described by Osváth-Ferencz et al. (2017). First, Cormack-Jolly-Seber (CJS) models were applied to evaluate the patterns in survival (φ) and capture probability (p), and subsequently the best supported pattern in those parameters served as a fixed basis for estimating seasonal population size (\(\hat {N}\)) with Jolly-Seber (JS) models (Arnason and Schwarz 1999; Schwarz and Seber 1999). The performance of the candidate model variants was assessed with the Akaike Information Criterion corrected for small sample size (AICc) (Hurvich and Tsai 1989), and the model φ(.) p(s), i.e. the one assuming constant survival rate and sex-dependent capture probability, was found to be the best supported. Consequently, due to the intersexual difference in capture probability the seasonal population sizes were estimated separately for both sexes, and then summed. In addition, we estimated mean life span of adult butterflies as \(\hat {e}={(1 - \hat {\varphi })^{ - 1}} - 0.5\) (Nowicki et al. 2005).
Based on the UTM co-ordinates of the capture points we calculated the distances moved by individual butterflies between consecutive captures, and analysed them against the day in season. Whenever the date of a particular capture and the following one differed (i.e. in most cases), the day in season for such a movement was derived as the mid-point between the capture dates. In addition, for the individuals captured a minimum of three times we established minimum convex polygons encompassing all the capture points and calculated their areas using GIS software Idrisi 2.0 (Eastman 1997). It can be assumed that the minimum convex polygon area (MCP) should grow asymptotically with the number of capture points (k) for an individual, and the asymptote value would reflect the home range size. Therefore, we evaluated the mean home range of adult butterflies in the investigated population through fitting the asymptotic function of \({\text{MCP}}=a(k - b)/k\), where a represents the asymptote value, and the b parameter was set at 3 so as to account for the fact that for less than three points MCP is essentially zero. It is worth noting that when as an alternative we fitted the function version with unconstrained b, this parameter value was anyway estimated at very close to 3 (\(\hat {b}\) ± SE = 3.035 ± 0.220), and the overall model fit remained virtually unchanged, which increases the reliability of our approach. While the investigations of movement distances were conducted separately for males and females, the home range analysis relied on the data pooled together for both sexes, because otherwise the sample sizes were too small.
Genetic study
A total number of 25 individuals of M. alcon were sampled in 2014 for genetic studies at the Bokiny site. Small pieces of wings were torn off using tweezers and placed into containers with 95% ethanol. DNA was extracted using a Genomic Mini Kit (A&A Biotechnology) as described in Rutkowski et al. (2009). DNA polymorphism was assessed using microsatellite loci. We PCR-amplified five microsatellite loci, characterized by Zeisset et al. (2005) as Macu5, Macu8, Macu11, Macu15 and Malc169, using primers as described by the authors and reagents and conditions as described by Rutkowski et al. (2009). The length of the amplified fragments was estimated using a CEQ8000 Beckman Coulter automated sequencer (Beckman Coulter, Inc., Brea, CA, USA; agency in Poland: Comesa, Warsaw), and subsequently the chromatograms were analysed using Beckman Coulter Fragment Analysis Software in order to obtain into the individual genotypes.
For comparative purposes we also used datasets from eight other previously studied populations of M. alcon in Poland (for details see Sielezniew et al. 2012), for which the same microsatellite markers were applied and thus the datasets are directly comparable. For each population we calculated the values of basic genetic indices, including the number of alleles (A), allelic richness defined as the number of alleles corrected for sample size using the rarefaction method with a sample of 9 individuals (R; Petit et al. 1998), observed (HO) and expected heterozygosity (HE; Nei 1978), as well as inbreeding coefficient (FIS). These analyses were performed using GenAlEx v. 6.503 (Peakall and Smouse 2012) and FSTAT version 2.9.3.2 (Goudet 2001). The deviation from the Hardy–Weinberg equilibrium (hereafter HWE) was evaluated using Fisher’s exact test in Genepop v.4 (Raymond and Rousset 1995; Rousset 2008), with the following settings: 10,000 dememorisations, 1000 batches and 10,000 iterations. Genetic differentiation among populations was expressed as FST based on the Infinite Allele Model of mutation (Weir and Cockerham 1984). Pairwise FST values and their significance, as well as the overall FST with corresponding 95% confidence intervals, were calculated in FSTAT.
Results
Throughout the entire season, we recorded 15 males and 11 females of M. alcon, which were captured 43 and 30 times respectively. While the daily capture probability was slightly (but insignificantly) higher in males than in females (\(\hat {p}\) ± SE = 0.80 ± 0.10 vs. 0.67 ± 0.09), the estimated sex ratio was still moderately male-biased. The seasonal population size was assessed at 40 adults individuals (with the 95% CI of 33–52 adults), including 23 males and 17 females.
The butterfly survival rate was estimated at a relatively high level of 0.814 (± SE = 0.051) per day, which corresponds to adult life expectancy of 4.88 (± SE = 1.59) days. In turn, the flight period was fairly short, and with the exception of a single male flying a few days earlier, it spanned 20 days. There was little indication of protandry (i.e. earlier emergence of adult males, typical for butterflies) with the peaks of male and female abundance occurring in the same period (Fig. 1), although it should be noted that due to low butterfly numbers both peaks were poorly pronounced.
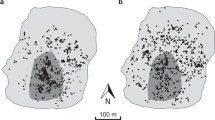
Overall we found 263 G. pneumonanthe foodplants at the investigated site, however a substantial fraction of them (97 i.e. 36.9%) were concentrated within a relatively small southern fragment of the site. The food plant density within this southern fragment thus reached 303 per ha, whereas within the remaining area it was only 48 per ha, i.e. almost an order of magnitude lower. Not surprisingly a majority of butterflies and their eggs were observed within the southern core fragment (Fig. 2). The total number of eggs found was 581 on 56 (21%) gentian plants, which translates into 10.2 eggs per gentian with eggs, and 34.2 eggs per average M. alcon female.

Within-season dynamics of the estimated daily numbers of M. alcon adults (males: solid squares, solid line; females: empty squares, broken line). Whiskers represent 95% confidence intervals of the estimates
The maximum recorded distance between consecutive captures was 156 m, which roughly corresponds to the maximum dimension of the southern fragment of the site predominantly used by M. alcon butterflies, and the mean movement distance was 50 m. There was no indication of any differences in the distances covered by both sexes (Student’s t test for ln-transformed distances to achieve normality: n = 42 for males and 31 for females, t = 0.850, P = 0.398). Similarly, the movement distances were not affected by the duration of the period between consecutive captures (Pearson’s correlation: r = − 0.145, P = 0.221). However, they depended on the date in season, and remarkably the pattern of this dependence was opposite in both sexes (Fig. 3). Male mobility decreased with the season progression (Pearson’s correlation: r = − 0.317, P = 0.041) while female one increased (Pearson’s correlation: r = 0.405, P = 0.024).

Movement distances between consecutive captures of M. alcon adults (males: solid dots; females: empty dots) in relation to the season progression, with day 0 representing 10 July. Lines show the best fit linear regressions for males (solid line; y = 87.6 − 2.10x) and females (broken line; y = − 15.3 + 4.40x)
As expected, the area of minimum convex polygons encompassing capture points of individual butterflies increased with the number of points, and the increase fitted very well to the asymptotic function applied (R2 = 0.7071, P < 0.0001; Fig. 4). The asymptote value, assumed to reflect the average butterfly home range size, was estimated at 0.294 (± SE = 0.028) ha, which is in perfect agreement with the area of the southern fragment of the site where most G. pneumonanthe foodplants grow. The above result, together with the aforementioned recorded movement distances, indicate that even though the butterflies do not utilise the whole grassland site to the same extent, they move freely and frequently within its entire core fragment comprising the most suitable habitat.

The area of minimum convex polygons encompassing individual butterfly capture points in relation to the number of captures. Solid line represents the best fit asymptotic function (y = 0.264(x − 3)/x), while broken like indicates the asymptote value
Several indices reflected relatively low genetic diversity of the Bokiny population and in the case of the observed heterozygosity the value was lowest among all the populations considered (Table 1). Moreover significant departure from Hardy–Weinberg Equilibrium (HWE) was detected in three out of five investigated loci as well as in the overall test across all the loci (P < 0.05 in all the cases), indicating heterozygote deficit. Overall FST reached 0.093 (with 95% CI of 0.076–0.108). The Bokiny population differed significantly from all other previously studied populations of M. alcon and its pairwise FST ranged between 0.078 and 0.180 (Table 2).
Discussion
Our findings confirm that the investigated population of M. alcon is a very small one, comprising only a few tens of breeding adult individuals. Admittedly, the results we presented are restricted to an intensive single-year fieldwork campaign, and the data we gathered for other years are not comprehensive enough to allow similar analyses. Nevertheless, it should be underlined that these data clearly indicate that the abundances of M. alcon butterflies and their eggs in other years were comparably low or even lower. All things concerned, it appears that the tiny size we recorded for the investigated population reflects its typical status rather than an occasional drop of a usually much larger but highly fluctuating population (cf. Nowicki et al. 2009).
The most obvious factor limiting our focal population is the low availability of G. pneumonanthe foodplants. The foodplants are in very short supply in most of the locality, and even within its southern core fragment their density (ca. 300 gentians per ha) is well below the typical levels recorded at M. alcon sites (WallisDeVries 2004; Nowicki et al. 2007, 2009; Radchuk et al. 2012; Czekes et al. 2014; but see; Maes et al. 2004). It is worth pointing out that 1000 gentians per ha has been suggested as the threshold value, below which the foodplant density becomes a critical resource for M. alcon populations (WallisDeVries 2004). In this context, it may seem somewhat contradictory that only a fraction of foodplant in our study were found to have M. alcon laid on them. However, such a situation is quite common, and stems from the fact that many gentians are either hidden by the surrounding higher vegetation and thus not visible for butterflies, or not in the appropriate phenological state to be used for oviposition (Küer and Fartmann 2004; Nowicki et al. 2007). In turn, the relatively high average number of eggs laid per used gentian implies a serious possibility of intraspecific competition among M. alcon larvae developing in foodplants. Besides, it cannot be excluded that host ant availability constitutes an additional limiting factor for M. alcon in Bokiny. Although an earlier study found that the primary host species Myrmica scabrinodis is the most common Myrmica ant at this study, the abundance of its nests was not quantified in absolute terms (Sielezniew et al. 2015).
Very small populations of M. alcon are not unique, but previous studies reported their existence in metapopulation systems (WallisDeVries 2004; Maes et al. 2004; Nowicki et al. 2007; Radchuk et al. 2012), where they are likely to be supported by a continuous influx of immigrants from more abundant populations located nearby (Hanski 1994; Gonzalez et al. 1998). In contrast, it is rather unusual that the population we investigated manages to persist in complete isolation. In this context we believe that our findings offer practical hints on demographic features that improve persistence chances of local butterfly populations and thus to identify those that deserve conservation efforts the most.
Long-term trend in population size is the most evident predictor of extinction risk (Caughley 1994; Keith et al. 2015). In the absence of a clear trend, the extent of population size fluctuations, serving as an inverse measure of stability, makes a useful indicator of population viability, as more fluctuating populations are more vulnerable to extinctions (Caughley 1994; Lundberg et al. 2000; Nowicki 2017). However, the assessment of both trend as well as fluctuation level requires relatively long time series of quantitative data for a particular population, which are rarely available, especially in the case of insects (Schtickzelle and Baguette 2009; Nowicki 2017). Therefore the assessments of population persistence chances often have to be based on proxies in the form of population characteristics which are simple and straightforward to evaluate. The present study may provide useful guidelines in this respect for assessing the status of butterfly populations.
Our focal population shows several clearly undesirable characteristics. Apart from its sheer size, strong isolation and resource limitation as discussed above, it apparently suffers from low genetic diversity, suggesting genetic drift due to isolation and/or that it underwent a prolonged bottleneck or possibly even a series of such events in the past (Saccheri et al. 2001). One potential consequence of reduced genetic diversity is low adaptability to changing environmental conditions, e.g. ongoing climate change (Cobben et al. 2012). Furthermore, low genetic variability may also lead to decreased individual fitness (Lacy 1997; Ortego et al. 2007). A potential manifestation of the above principle could be the very low female fecundity that we recorded in the present study, although we do not have any direct evidence for its genetic basis. Moreover, the correlation between variation in neutral markers, such as microsatellites, and fitness components remains hotly debated issue (Väli et al. 2008; Ljungqvist et al. 2010; Szulkin et al. 2010). Nonetheless, most previous studies estimated the number of eggs laid per an average female of M. alcon at 80–100 (WallisDeVries 2004; Maes et al. 2004; Radchuk et al. 2012), whereas our reported value is about three-fold lower. Such a situation is clearly disadvantageous for the population persistence chances, since it translates into much lower achievable population growth rate, thus slowing down the recovery from occasional declines in ‘bad years’, caused by environmental stochasticity (Lande 1993).
Nevertheless, the M. alcon population in Bokiny also has some promising features. First is the relatively short flight period and long adult life expectancy. Obviously, both parameters are known to exhibit some inter-annual variability, mostly due to different weather patterns (Bubova et al. 2016), and thus using only single-year estimates it is impossible to ascertain whether they are population specific or perhaps they merely reflect a favourable season effect. However, it is worth noting that another much larger M. alcon population (Brzeźno in E Poland), surveyed in the same year for comparison, was characterised by substantially longer flight period and shorter adult life span (35 and 2.87 ± 0.14 days respectively; Pałka and Sielezniew unpubl. data). Moreover, the estimate of adult life span that we obtained for the Bokiny population is among the highest ever found for M. alcon across Europe (and clearly the highest if the values for the xerophilous ‘rebeli’ ecotype are excluded), whereas the flight period length is near the lower limit of the reported range (see review in Bubova et al. 2016). Both short flight period and long adult life expectancy imply low level of temporal fragmentation (sensu Bubova et al. 2016). In other words, all adult individuals emerging in a season, even in its initial or final part, have good opportunities to find mating partners within the time window they fly. In the Bokiny population, the situation is further improved by little protandry, i.e. almost synchronous emergence of males and females. Consequently, no females should be expected to die unmated, which is often a problem in butterfly populations with clearly pronounced protandry (Calabrese and Fagan 2004). Nevertheless, the above finding does not necessarily imply completely random mating in the focal population. Some individuals may still be more successful in mating than others, or simply their offspring may be more successful in survival due to patchy distribution of host ant nests.
Another advantageous attribute of our focal population is the considerable mobility of the investigated butterflies. Our results concerning both the recorded movement distances as well as the estimated home range size suggest that M. alcon adults move freely within the southern fragment of the site, constituting the core habitat patch, and they occasionally venture to more distant areas. At first glance, unconstrained movements within a habitat patch seem nothing peculiar, but in Maculinea this is not necessarily the case, because these butterflies have been found to establish restricted home ranges within their habitat patches (Hovestadt and Nowicki 2008; Kőrösi et al. 2008). The fact that in Bokiny they are able to mix freely is a further feature enhancing individual mating chances, and thus population viability. Interestingly, mobility of males was found to decrease and that of females to increase with season progression, which appears in accordance with optimal breeding strategies of both sexes. Female tendency to fly further distances towards the end of season may reflect their increased effort to find foodplants with no eggs previously laid by conspecifics in order to reduce intra-specific competition experienced by their offspring. In turn, higher male mobility at the start of season enables them to search more intensively for newly emerging females, which is particularly important in a population with a very low density of flying adults.
As discussed above, several attributes of the investigated population (short flight period, long adult life expectancy, little protandry, relatively good mobility) ensure that all individuals have a good chance to mate as adults. Consequently, unlike in many wild populations (Frankham 1995; Luikart et al. 2010), the effective population size is presumably not much reduced as compared with the census population size, i.e. the sheer number of individuals making the population. We expect that this may be the main reason for which M. alcon is able to persist in Bokiny in spite of its very low abundance and strong isolation there. To conclude, our study indicates that very small size does not necessarily translate into low viability, and thus low conservation value of an isolated local population. Therefore, the prioritisation of populations to be included in conservation programmes should not rely on population census alone, but instead it should be based on more detailed analyses of demographic parameters, in the case of insects including their fecundity, mobility, adult longevity and occurrence period.
References
Als TD, Vila R, Kandul NP, Nash DR, Yen SH, Hsu YF, Mignault AA, Boomsma JJ, Pierce NE (2004) The evolution of alternative parasitic life histories in large blue butterflies. Nature 432:386–390
Arnason AN, Schwarz CJ (1999) Using POPAN-5 to analyse banding data. Bird Study 46:157–168
Bereczki J, Pecsenye K, Varga Z, Tartally A, Tóth JP (2018) Maculinea rebeli (Hirschke)—a phantom or reality? Novel contribution to a long-standing debate over the taxonomic status of an enigmatic Lycaenidae butterfly. Syst Entomol 43:166–182
Bonelli S, Vrabec V, Witek M, Barbero F, Patricelli D, Nowicki P (2013) Selection on dispersal in isolated butterfly metapopulations. Popul Ecol 55:469–478
Bubová T, Kulma M, Vrabec V, Nowicki P (2016) Adult longevity and its relationship with conservation status in European butterflies. J Insect Conserv 20:1021–1032
Calabrese JM, Fagan WF (2004) Lost in time, lonely, and single: reproductive asynchrony and the Allee effect. Am Nat 164:25–37
Caughley G (1994) Directions in conservation biology. J Anim Ecol 63:215–244
Cobben MMP, Verboom J, Opdam PFM, Hoekstra RF, Jochem R, Smulders MJM (2012) Landscape prerequisites for the survival of a modelled metapopulation and its neutral genetic diversity are affected by climate change. Landscape Ecol 27:227–237
Czekes Z, Markó B, Nash DR, Ferencz M, Lázár B, Rákosy L (2014) Differences in oviposition strategies between two ecotypes of the endangered myrmecophilous butterfly Maculinea alcon (Lepidoptera: Lycaenidae) under unique syntopic conditions. Insect Conserv Divers 7:122–131
Eastman JR (1997) Idrisi for Windows. User’s Guide Version 2.0. Clark University, Worchester
Frankham R (1995) Effective population size/adult population size ratios in wildlife: a review. Genet Res 66:95–107
Gonzalez A, Lawton JH, Gilbert FS, Blackburn TM, Evans-Freke I (1998) Metapopulation dynamics, abundance, and distribution in a microecosystem. Science 281:2045–2047
Goudet J (2001) FSTAT V2.9.3, a program to estimate and test gene diversities and fixation indices. http://www.unil.ch/izea/softwares/fstat.html
Haddad NM (1999) Corridor and distance effects on interpatch movements: a landscape experiment with butterflies. Ecol Appl 9:612–622
Hanski I (1994) A practical model of metapopulation dynamics. J Anim Ecol 63:151–162
Hanski I, Gaggiotti OE (eds) (2004) Ecology, genetics and evolution of metapopulations. Elsevier, San Diego
Hanski I, Moilanen A, Gyllenberg M (1996) Minimum viable metapopulation size. Am Nat 147:527–541
Hovestadt T, Nowicki P (2008) Investigating movement within irregularly shaped patches: Analysis of mark–release–recapture data using randomization procedures. Isr J Ecol Evol 54:137–154
Hurvich CM, Tsai C (1989) Regression and time series model selection in small samples. Biometrika 76:297–307
Keith D et al (2015) Temporal correlations in population trends: Conservation implications from time-series analysis of diverse animal taxa. Biol Conserv 192:247–257
Kőrösi A, Örvössy N, Batáry P, Kövér S, Peregovits L (2008) Restricted within habitat movement and time-constrained egg laying of female Maculinea rebeli butterflies. Oecologia 156:455–464
Küer A, Fartmann T (2004) Prominent shoots are preferred: microhabitat preferences of Maculinea alcon ([Denis & Schiffermüller], 1775) in Northern Germany (Lycaenidae). Nota Lepid 27:309–319
Lacy RC (1997) Importance of genetic variation to the viability of mammalian populations. J Mammal 78:320–335
Lande R (1993) Risks of population extinction from demographic and environmental stochasticity and random catastrophes. Am Nat 142:911–927
Ljungqvist M, Åkesson M, Hansson B (2010) Do microsatellites reflect genome-wide genetic diversity in natural populations? A comment on Väli et al. (2008). Mol Ecol 19:851–855
Luikart G, Ryman N, Tallmon DA, Schwartz MK, Allendorf FW (2010) Estimation of census and effective population sizes: the increasing usefulness of DNA-based approaches. Conserv Genet 11:355–373
Lundberg P, Ranta E, Ripa J, Kaitala V (2000) Population variability in space and time. Trends Ecol Evol 15:460–464
Maes D, van Dyck H (2005) Habitat quality and biodiversity indicator performances of a threatened butterfly versus a multispecies group for wet heathlands in Belgium. Biol Conserv 123:177–187
Maes D, Vanreusel W, Talloen W, Van Dyck H (2004) Functional conservation units for the endangered Alcon Blue butterfly Maculinea alcon in Belgium (Lepidoptera: Lycaenidae). Biol Conserv 120:229–241
Nei M (1978) Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89:583–590
Nowicki P (2017) Survey precision moderates the relationship between population size and stability. Biol Conserv 212:310–315
Nowicki P, Richter A, Glinka U, Holzschuh A, Toelke U, Henle K, Woyciechowski M, Settele J (2005) Less input same output: simplified approach for population size assessment in Lepidoptera. Popul Ecol 47:203–212
Nowicki P, Pepkowska A, Kudlek J, Skórka P, Witek M, Settele J, Woyciechowski M (2007) From metapopulation theory to conservation recommendations: lessons from spatial occurrence and abundance patterns of Maculinea butterflies. Biol Conserv 140:119–129
Nowicki P, Bonelli S, Barbero F, Balletto E (2009) Relative importance of density-dependent regulation and environmental stochasticity for butterfly population dynamics. Oecologia 161:227–239
Nowicki P, Vrabec V, Binzenhöfer B, Feil J, Zakšek B, Hovestadt T, Settele J (2014) Butterfly dispersal in inhospitable matrix: rare, risky, but long-distance. Landscape Ecol 29:401–412
Ortego J, Calabuig G, Cordero PJ, Aparicio JM (2007) Egg production and individual genetic diversity in lesser kestrels. Mol Ecol 16:2383–2392
Osváth-Ferencz M, Bonelli S, Nowicki P, Peregovits L, Rákosy L, Sielezniew M, Kostro-Ambroziak A, Dziekańska I, Kőrösi A (2017) Population demography of the endangered large blue butterfly Maculinea arion in Europe. J Insect Conserv 21:411–422
Peakall R, Smouse P (2012) GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research—an update. Bioinformatics 28:2537–2539
Pecsenye K, Bereczki J, Tihanyi B, Tóth A, Peregovits L, Varga L (2007) Genetic differentiation among the Maculinea species (Lepidoptera: Lycaenidae) in eastern Central Europe. Biol J Linnean Soc 91:11–21
Petit RJ, El Mousadik A, Pons O (1998) Identifying populations for conservation on the basis of genetic markers. Conserv Biol 12:844–855
Prugh LR, Hodges KE, Sinclair ARE, Brashares JS (2008) Effect of habitat area and isolation on fragmented animal populations. Proc Natl Acad Sci USA 105:20770–20775
Radchuk V, WallisDeVries MF, Schtickzelle N (2012) Spatially and financially explicit population viability analysis of Maculinea alcon in The Netherlands. PLoS ONE 7:e38684
Raymond M, Rousset F (1995) GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. J Hered 86:248–249
Rousset F (2008) Genepop’007: a complete reimplementation of the Genepop software for Windows and Linux. Mol Ecol Resour 8:103–106
Rutkowski R, Sielezniew M, Szostak A (2009) Contrasting levels of polymorphism in cross-amplified microsatellites in two endangered xerothermophilous, obligatorily myrmecophilous, butterflies of the genus Phengaris (Maculinea) (Lepidoptera: Lycaenidae). Eur J Entomol 106:457–469
Saccheri IJ, Nichols RA, Brakefield PM (2001) Effects of bottlenecks on quantitative genetic variation in the butterfly Bicyclus anynana. Genet Res 77:167–181
Schtickzelle N, Baguette M (2009) Metapopulation viability analysis: a crystal ball for the conservation of endangered butterflies? In: Settele J, Shreeve TG, Konvicka M, Van Dyck H (eds) Ecology of butterflies in Europe. Cambridge University Press, London, pp 339–352
Schwarz CJ, Seber GAF (1999) Estimating animal abundance: review III. Stat Sci 14:427–456
Settele J, Kühn E, Thomas JA (eds) (2005) Studies on the ecology and conservation of butterflies in Europe. vol 2. Species ecology along a European gradient: Maculinea butterflies as a model. Pensoft, Sofia
Sielezniew M, Rutkowski R, Ponikwicka-Tyszko D, Ratkiewicz M, Dziekańska I, Švitra G (2012) Differences in genetic variability between two ecotypes of endangered myrmecophilous butterfly Phengaris (= Maculinea) alcon—the setting of conservation priorities. Insect Conserv Divers 5:223–236
Sielezniew M, Bystrowski C, Deoniziak K, Da Costa JM (2015) An unexpected record of Myrmica schencki Emery, 1895 as a secondary host ant of the hygrophilous form of a small and isolated population of the Alcon Blue butterfly Phengaris (= Maculinea) alcon (Denis et Schiffermüller, 1775) (Lepidoptera, Lycaenidae) in NE Poland. Pol J Entomol 84:49–59
Sielezniew M, Bystrowski C, Deoniziak K (2016) Motyle dzienne Narwiańskiego Parku Narodowego. In: Banaszuk P, Wołkowycki D (eds) Narwiański Park Narodowy. Krajobraz, przyroda, człowiek. Agencja Wydawnicza Ekopress. Białystok–Kurowo, Poland, pp 149–159
Skórka P, Nowicki P, Lenda M, Witek M, Śliwińska EB, Settele J, Woyciechowski M (2013) Different flight behaviour of the endangered scarce large blue butterfly Phengaris teleius (Lepidoptera: Lycaenidae) within and outside its habitat patches. Landscape Ecol 28:533–546
Soga M, Koike S (2013) Mapping the potential extinction debt of butterflies in a modern city: implications for conservation priorities in urban landscapes. Anim Conserv 16:1–11
Szulkin M, Bierne N, David P (2010) Heterozygosity-fitness correlations: a time for reapprasial. Evolution 64:1202–1217
Tartally A, Nash DR, Lengyel S, Varga Z (2008) Patterns of host ant use by sympatric populations of Maculinea alcon and M. rebeli in the Carpathian Basin. Insect Soc 55:370–381
Thomas JA (1995) The ecology and conservation of Maculinea arion and other European species of large blue butterfly. In: Pullin AS (ed) Ecology and conservation of butterflies. Chapman and Hall, London, pp 180–197
Väli Ü, Einarsson A, Waits L, Ellegren H (2008) To what extent do microsatellite markers reflect genome-wide genetic diversity in natural populations. Mol Ecol 17:3808–3817
van Swaay C et al (2010) European red list of butterflies. Publications Office of the European Union, Luxembourg
WallisDeVries MF (2004) A quantitative conservation approach for the endangered butterfly Maculinea alcon. Conserv Biol 18:489–499
Weir BS, Cockerham CC (1984) Estimating F-statistics for the analysis of population structure. Evolution 38:1358–1370
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46:120–138
Witek M, Nowicki P, Sliwinska E, Skórka P, Settele J, Schönrogge K, Woyciechowski M (2010) Local host ant specificity of Phengaris (Maculinea) teleius butterfly, an obligatory social parasite of Myrmica ants. Ecol Entomol 35:557–564
Zeisset I, Als TD, Settele J, Boomsma JJ (2005) Microsatellite markers for the large blue butterflies Maculinea nausithous and Maculinea alcon (Lepidoptera: Lycaenidae) and their amplification in other Maculinea species. Mol Ecol Notes 5:165–168
Acknowledgements
The fieldwork was conducted with a proper permission from the Ministry of Environment. The analysis was supported by the Polish National Science Centre Grant DEC-2013/11/B/NZ8/00912. Christine Richards improved the English of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Nowicki, P., Deoniziak, K., Dziekańska, I. et al. What keeps ‘living dead’ alive: demography of a small and isolated population of Maculinea (= Phengaris) alcon. J Insect Conserv 23, 201–210 (2019). https://doi.org/10.1007/s10841-018-0078-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-018-0078-4