
Abstract
Analysis of Cladocera and organic matter from the sediments of the Rzecino paleolake are presented. These are probably among the first detailed published data from the Eemian Interglacial (MIS 5e) organic matter. The Rzecino paleolake is located in NW Poland in an area covered by the last glaciation. Approximately 5.8 m of sediment was analyzed. Previous palynological data indicate a continuous accumulation from the final phase of the Wartanian to the early Vistulian Glaciation. Data on the Cladocera correlated with the results of the analyses of organic matter, which enabled the reconstruction of varying environmental conditions and geneses of organic matter. In the final phase of the Wartanian and at the beginning of the Eemian Interglacial, low frequencies of Cladocera specimens and species were identified, which suggests unfavorable conditions for the development of zooplankton. Organic matter (OM) was characterized by a small amount of organic carbon and nitrogen, a high value of C/N (18–27) and a δ13C of approximately −26 ‰, which suggests a higher plant source and a predominantly terrestrial origin for OM. An improvement in environmental conditions occurred during the pollen zone E3. The number of Cladocera species and individuals increased. An increase in TOC and TN suggests an enhancement of primary productivity and the development of macrophytes in the lake. A lower C/N ratio indicates a primary role of algae in the production of organic matter. During the Eemian optimum (pollen zones E4–E5), mostly Cladocera species preferring warm water occurred. Algae remained a significant component of OM. The early Eemian post-optimum (pollen zone E6) was characterized as a period of maximal abundance of zooplankton (20 species). During this period, rapid decreases in TOC, TN and the C/N ratio and an increase in δ13C values suggest the reduced abundance of higher plants in the lake. In pollen zone E7, the frequency and diversity of Cladocera rapidly decreased. Species that tolerate cold water were dominant, which reflects unfavorable conditions for the development of zooplankton but a high water level. The low content of organic carbon and nitrogen, a C/N ratio in the range of 13–17 and δ13C values in the range of −26.2 to −28 ‰ suggest the inhibition of terrestrial plant delivery. During the Early Vistulian (pollen zone EV1), rare faunal remains and the features of OM (the lowest amounts of TOC and TN, and the highest value of C/N) reflect the decline of lake conditions and a lack of primary production and aquatic vegetation in the lake.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Eemian lacustrine successions are common in central and eastern Poland (Mamakowa 1989; Kupryjanowicz 2008), but these successions are rare in northwestern Poland (West Pomerania Lakeland). The Rzecino paleolake is an interesting site in this region. The uniqueness of the Rzecino paleolake is due to its location in NW Poland, i.e., in the area strongly modified and covered by the last glaciation deposits. This location means that both the oceanic influence and the expanding Scandinavian ice sheet had an effect on climate and lake conditions. Sediments from Rzecino have been the subject of pollen, petrographic, sedimentological, magnetic susceptibility, carbonate oxygen and carbon isotope analyses (Winter and Dobracka 2006; Winter et al. 2008; Mirosław-Grabowska 2008). Previous investigations focused on the: (1) estimation of biostrathigraphy of sediments and a terrestrial vegetation history (based on pollen analyses), (2) lithological characteristics of the Eemian deposits (petrographic and sedimentological analyses), (3) climatic reconstruction (pollen and magnetic susceptibility analyses), (4) carbonate deposits and evolution of lake (carbonate oxygen and carbon isotope analyses).
The aim of this study was to examine changes in the Cladocera community and organic matter in sediments from the Rzecino palaeolake and their significance. During the Eemian Interglacial and the Early Vistulian (Weichselian), the Rzecino paleolake was transformed by natural processes, such as sediment infilling and overgrowth, which were partially caused by climate deterioration. At that time, changes occurred in the quantity and diversity of the Cladocera species, isotopic features and the origin of organic matter. We compare Cladocera with selected geochemical and isotopic data and the reconstructions of environmental changes throughout the Eemian Interglacial and the Early Vistulian. We present the results of our studies of lake deposits with respect to the background of the vegetation history of this area. Detailed data on the Eemian organic deposits from Poland are first published herein.
Study site
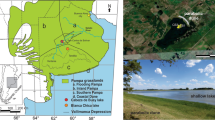
The Rzecino paleolake (53°50′27″N, 15°59′57″E) is located near Połczyn-Zdrój, which is situated in the West Pomerania Lakeland (NW Poland), as shown in Fig. 1, at an elevation of 104.5 m a.s.l. The sediments fill a tectonic graben, which was formed along a large faulting zone that borders the Połczyn-Zdrój salt dome in the southwest (Winter and Dobracka 2006). The detailed geological structure were presented by Dobracka (2001) and by Winter et al. (2008). During the Eemian Interglacial and the Early Vistulian (Weichselian), this area had features of the lakeland (Winter and Dobracka 2006).

Location of Rzecino paleolake. Asterisk Rzecino paleolake; circle Eemian sites citied in text: Be Besiekierz (Mirosław-Grabowska and Niska 2005); Ka Kaliska (Mirosław-Grabowska and Niska 2007a); Ku Kuców IIc (Niska 2008); Ru Ruszkówek (Mirosław-Grabowska et al. 2009); So Solniki (Kupryjanowicz 2008, Niska 2008), Sl Sławoszewek (Pawłowski 2011); St Studzieniec (Mirosław-Grabowska and Niska 2007b); marked area West Pomerania Lakeland
Recently the area of the Rzecino paleolake is associated with a moderate marine-continental climatic region with a clear predominance of marine influence. Due to the proximity of the Baltic Sea (about 45 km), the climate is characterized by high humidity. The annual precipitation is 650 mm (an average 135 days year−1 with precipitation). The mean annual temperature is 7.5 °C less than western Poland as a result of the cooling influence of the Baltic Sea. The mean temperature in the warmest month (July) is 14 °C. The second half of the summer and the early autumn are characterized by significant precipitation. The mean temperature in the coldest month (January) is −3 °C. Due to the warming influence of the sea, the winters are characterized by mild temperatures with snow cover of 40 days. Westerly winds, with an average wind speed of 1.8 m s−1, predominate in the growing season, which begins in early April and persists for 210–220 days (http://www.lobez.pl/pl/Polozenie.html).
Methods
Sediment core
The sediments were drilled during cartographic field works for the Detailed Geological Map of Poland at a 1:50 000 scale on a Dobrowo sheet (Dobracka 2001). The core was obtained using a rotary drilling rig. The sediment core was a form of massive, cylindrical sections of a 1-m length. The deposit core consists of a 20.1-m series of lacustrine and fluvial sediments, which are composed of sands, silts and gyttja.
In this study, sediments thicker than 600 cm from a depth range of 3,460–4,040 cm were analyzed. Sandy silts were detected on the bottom (below 3,960 cm) and next calcareous gyttja (3,960–3,500 cm). Organic silts were detected at the depth range of 3,640–3,760 cm. The upper part (above 3,500 cm) of this succession contains organic sands.
Chronology
The sediments from the Rzecino palaeolake are only “palynologically dated”. There are no radiometric dates because the entire Eemian Interglacial fall outside the range of radiocarbon dating, and the carbonate detritus lacustrine sediments are very problematic when addressed by other absolute chronometric dating methods. Therefore only the results of palynological analysis confirm the Eemian age of the Rzecino paleolake deposits. The Eemian Interglacial is correlated with Marine Isotope Stage, MIS 5e (Oxygen Isotope Stages—OIS 5e; Shackleton et al. 2003).
Cladocera analysis
Cladocera analysis was performed using 58 samples from the core depth 3,460–4,040 cm. Samples of 1.5 g, were prepared according to a slightly modified standard procedure (Frey 1986). After the removal of carbonates using HCl, each sample was boiled in 10 % KOH for 20 min. After washing with distilled water, the residue was sieved through a 40-μm-mesh sieve. The fine material was transferred into a polycarbon test tube. Prior to counting, the remains were colored with safranine. The samples were analysed under light microscope with 100–400× magnification. A minimum of 200 remains of Cladocera (3–5 slides) were examined from each sample. Firstly all remains from each slide were enumerated (headshield, shell, postabdomen, postabdominal claws, exopods and antennules) and the most abundant remains of each taxa were chosen to represent the number of individuals.
The identification and ecological interpretation of the Cladocera remains were conducted based on the studies by Duigan (1992); Frey (1958, 1962); Goulden (1964); Hofmann (1986, 2000); Korhola (1990); Flössner (2000); Szeroczyńska (1985); Szeroczyńska and Sarmaja-Korjonen (2007). The concentration diagram (Fig. 2), the percentage diagram (Fig. 3), the total number of Cladocera specimens and species (Fig. 4), and the ratio of planktonic to littoral form P/L (Fig. 4) were performed using PolPal computer program (Nalepka and Walanus 2003). For statistical analyzes, the content of the remains of Cladocera was converted into 1-g−1 units of sediment.

Diagram of the absolute number of Cladocera individuals in the sediments of Rzecino paleolake. CAZ Cladocera Assemblage Zone; Lithology: 1 gyttja, 2 silt/sandy silt, 3 sand; E1–E7 regional pollen assemblage zones of the Eemian Interglacial

Percent species composition of Cladocera fauna in the sediments of Rzecino paleolake. CAZ Cladocera Assemblage Zone; Lithology: 1 gyttja, 2 silt/sandy silt, 3 sand; E1–E7 regional pollen assemblage zones of the Eemian Interglacial

Diagram of total number of Cladocera specimens and number of species diversity and ratio of planktonic to littoral form—P/L in the sediments of the Rzecino profile. CAZ Cladocera Assemblage Zone; Lithology: 1 gyttja, 2 silt/sandy silt, 3 sand; E1–E7 regional pollen assemblage zones of the Eemian Interglacial
The Cladocera species were classified into three habitat preference groups —bottom-dweller species, species associated with vegetation, species restricted to vegetation (Korhola 1990; Flössner 1964; Whiteside 1970; Hofmann 1987; Whiteside and Swindoll 1988) and were also categorized as planktonic (offshore) and littoral (meiobenthic) groups on Mueller (1964). The species Chydorus sphaericus and Bosmina longirostris were classified into littoral and open water zones (Whiteside 1970). Lake water acidity was estimated on the basis of the classification of species into five pH group developed by Krause-Dellin and Steinberg (1986). However, distribution of Cladocera is also affected by factors, such as trophic state, acidification, water temperature, and fish predation (Korhola and Rautio 2001; Krause-Dellin and Steinberg 1986).
Subfossil Cladocera are commonly used to paleolimnological reconstructions. Due to their characteristics (small size, ability to produce ephippia, which may be transported by birds to colonize new water bodies, and the ability to reproduce by parthenogenesis), Cladocera can be widely dispersed than the other representatives of aquatic fauna. Therefore, their response to climatic and environmental changes may be faster and more sensitive than responses by larger organisms (Frey 1962).
Isotopic and geochemical analyses
Geochemical analyses of 49 samples of carbon and nitrogen stable isotopes were collected every 10 cm from 3,520 to 3,990 cm. The sediments were dried at 60 °C and ground. The carbonate fraction was removed with hydrochloric acid. The carbon and nitrogen isotope compositions were analyzed using the Flash Elemental Analyzer 1112 and Thermo MAT 253 mass spectrometer, which were calibrated based on an internal nicotinamide standard and reported as per mill (‰) deviations versus atmospheric N2 (δ15N) and Vienna Pee Bee Belemnite (δ13C). The analytical errors (l SD) for the δ13C and δ15N measurements were 0.17 and 0.24 ‰, respectively. The organic carbon and nitrogen concentrations were analyzed using the Elemental Analyzer Vario Micro Cube. The analyses were performed in the Laboratory for Isotope Dating and Environmental Studies at the Institute of Geological Sciences of the Polish Academy of Sciences in Warsaw, Poland.
The δ15N and δ13C ratios of bulk organic matter (OM) in lake sediments can be used to infer changes in organic matter (aquatic or terrestrial source). Changes in δ13C and δ15N can also provide information on nutrient cycling, lake mixing regimes and water column stability (e.g., Meyers and Ishiwatari 1993; Meyers and Lallier-Vergès 1999).
The carbon and nitrogen stable isotopes were used to define the changes in the quantity and source of organic matter transported into the paleolake. Lake morphology, catchment topography and the relative abundances of lake and catchment plants significantly influence the relative contributions from terrestrial and aquatic sources to lake sediments (Meyers and Ishiwatari 1993).
Carbon to nitrogen ratio (C/N) is an indicator for nitrogen limitation of plants and other organisms. The C/N ratio indicates the primary source of organic matter in the lake. For example, C/N ratios in the range 4–10 are usually from marine sources, whereas higher ratios are likely to come from a terrestrial source. C/N ratios less than 10–12 are typical for a mainly algal source and those greater than 20 suggestive of a terrestrial plant origin. Therefore the C/N ratios of sediments have frequently been used to infer the relative proportions of organic detritus derived from algae and terrestrial plants (Smeltzer and Swain 1985; Tyson 1995; Meyers and Lallier-Vergès 1999).
Results
Cladocera data
The subfossil Cladoceran fauna of deposits from the Rzecino paleolake are represented by 22 species that belong to four families: Bosminidae, Chydoridae, Sididae and Daphnidae. The majority of the remains consist of Chydoridae and Bosminidae. In the deposits, the ephippial eggs of the Daphnia longispina group, Ceriodaphnia spp., Chydorus spp., and Bosmina spp. are also identified. Six faunal zones (Cladocera Assemblage Zone—CAZ) were identified based on quantitative and qualitative analysis supported by numerical analysis—Constrained Single link of sample method (ConSLink) provides by PolPal (Nalepka and Walanus 2003). The indicators of the importance of Cladocera species were considered to determine the faunal zones and especially the subzones (Figs. 2, 3). Bosmina longirostris is the dominant species during all development periods of the Rzecino paleolake.
Cladocera Assemblage Zone I (3,985–4,040 cm) is characterized by a low diversity of Cladocera species; however, the frequency of specimens was numerous (>330 specimens g−1, Fig. 4). Five pioneer species were identified (Fig. 2). Alona affinis is strongly associated with the macrophytes, which tolerate cool water. Bosmina longirostris (O.F. Müller) and Chydorus sphaericus are eurybiont and can exist in various environments and in different ecological niches in the lake, especially in water with a high trophic level (Frey 1986). At a depth of 4,010 cm, a high frequency of planktonic Bosmina (E.) coregoni (Baird) and Bosmina longirostris were reported.
Cladocera Assemblage Zone II (3,908–3,985 cm) is characterized by a low frequency of Cladocera specimens (less than 100 specimens g−1, Fig. 4) and only 3 species. Two of these species—Alona affinis and Eurycercus lamellatus (O. F. Müller)—are associated with water plants and can tolerate low temperatures and clear oligotrophic water. The third species is Bosmina longirostris, which is characterized by a broad environmental tolerance (Fig. 2).
Cladocera Assemblage Zone III (3,853–3,908 cm) differs from the other zones by the occurrence of 8 species, e.g., Alona affinis, Eurycercus lamellatus which are cold water-tolerant, and Chydorus sphaericus which is tolerant of broad range of environmental condition. The number of individuals in this species is high (maximum of 400 specimens g−1). At a depth of 3,880 cm, the frequencies of B. longirostris and B. (E.) coregoni increase (Figs. 2, 3). The end of this zone is characterized by a decline in the frequency of B. longirostris and Ch. Sphaericus, and an increase in the frequency of Chydorus spp. ephippium (maximum 10 %, Fig. 3) and other species.
Cladocera Assemblage Zone IV (3,742–3,853 cm) is characterized by maximal abundance of zooplankton (approximately 930 specimens g−1) in which 20 species of Cladocera have been identified (Fig. 4). Based on the faunal composition, significant differences in the succession of species and the number of individuals as well as the occurrence of indicator species of Cladocera four subzones have been distinguished (Fig. 2).
In subzone IVa (3,810–3,853 cm), 18 species including both littoral and planktonic species occur. The following species that prefer warm water appeared: Camptocercus rectirostris (Schoedler), Pleuroxus (P.) truncatus (O.F. Müller), Pleuroxus spp., and Graptoleberis testudinaria (Fischer). A characteristic feature of this subzone are benthic species, e.g., Pleuroxus uncinatus, Alona quadrangularis and Monospilus dispar. Monospilus dispar and Alonella exigua are also associated with mesotrophic conditions (Adamska and Mikulski 1969). Graptoleberis testudinaria was detected at a depth of 3,830 cm.
In Subzone IVb (3,788–3,810 cm), 16 species were identified (Fig. 4). The abundance of Bosmina longirostris and Bosmina (E.) coregoni decrease and two benthic species, Monospilus dispar and Alonopsis elongata, increase. High abundance of Alona affinis, Alona guttata, Eurycercus lamellatus and Monospilus dispar, which are typical of mesotrophic water, are noted. In this subzone, two taxa that prefer water with high temperatures also occur: Camptocercus rectirostris and Pleuroxus (P.) truncatus (Poulsen 1944).
Subzone IVc (3,768–3,788 cm) is characterized by a decrease in the abundance and diversity of the cladoceran fauna (6 species, >60–100 specimens g−1 excluding B. longirostris). Although B. longirostris dominates (Fig. 3, 60 %), its abundance decreases. The frequency of the benthic species Alona quadrangularis is high (35 %, Fig. 3). Alonella excisa is present at the beginning and at the end of this subzone. Only species that are tolerant of cold water (Chydorus sphaericus and Acroperus harpae) are present in the middle of this subzone.
In Subzone IVd (3,742–3,768 cm), an increase in both abundance and diversity of Cladocera is observed. Total abundance of Cladocera reached a maximum of 2,667 specimens g−1 (Fig. 4). An increase species which prefer warm water (e.g., Camptocercus rectirostris and Pleuroxus trigonellus) and tolerate low water temperatures (e.g., Acroperus harpae and Eurycercus lamellatus) is observed. The maximum abundance in Bosmina longirostris and Bosmina (E.) coregoni (930 individuals g−1) and an occurrence of the Daphnia longispina group is noted.
At the beginning of Cladocera Assemblage Zone V (3,625–3,742 cm), a rapid deterioration in environmental conditions occurred that may be associated with a decrease in water level. The presence of benthic species Alona quadrangularis (O. F. Müller), whose abundance reached 30 % (Fig. 3), and Oxyurella tenuicaudis (G. O. Sars) appeared. This finding was indicated on the mud-vegetation substrates in Ireland lakes (Duigan and Kovach 1991). A decrease in open-water species was also noted. In this zone, the abundance of cladoceran remains are low. Only 11 species of Cladocera were identified (Fig. 4). The majority of the following species can tolerate cold water: Chydorus sphaericus, Acroperus harpae, Alona affinis (G. O. Sars), Eurycercus lamellatus. The disappearance of Cladocera that thrive in warm water is observed (Pleuroxus spp., Camptocercus rectirostris).
Cladocera Assemblage Zone VI (3,485–3,625 cm) exhibits few faunal remains and only 1 species—Bosmina longirostris and frequently Bosmina spp. ephippia (maximum 50 %, Fig. 3)—are identified. No Cladocera remains have been identified above a depth of 3485 cm.
Isotopic and geochemical data
TOC, TN, and C/Natomic
The lowest sediments (below 3,890 cm) are characterized by a concentration of organic carbon in the range of 0.1–0.5 % and a nitrogen content below 0.1 % (Fig. 5). The C/Natomic ratio oscillates from 18 to 27. The content of both TOC and TN (maximum depth of 3,860 cm) rapidly increases to 16.4 and 1.2 %, respectively, but the C/N ratio decreases to 15. In the deposits at depths ranging from 3,860 to 3,800 cm, the Corg content fluctuates from 14.1 to 18.7 % and the N content varies from 0.9 to 1.31 %. The C/N ratio oscillates at approximately 17. In the overlying deposits, the TOC and TN attain maxima of 33.6 and 1.6 %, respectively, (maximum depth of 3,740 cm), which rapidly reduces to the ranges of 1–3.3 % and 0.1–0.2 %, respectively (at a depth above 3,740 cm). These values slightly increase at depths of 3,550 and 3,670 cm. The C/N ratio initially reduces to a range of 13–17 and systematically increases above 38. The C/N ratio decreases to 30 above a depth of 3,540 cm.

Results of selected geochemical and isotopic analyses of the sediments of the Rzecino paleolake. R PAZ regional pollen assemblage zones; E1–E7 Eemian Interglacial, EV early Vistulian, W Wartanian; Lithology: 1 gyttja, 2 silt/sandy silt, 3 sand
δ13 and δ15N
In the lowest sediments (below 3,880 cm), the δ13C values slightly oscillate at approximately −26 ‰ (Fig. 5). Due to a low content of nitrogen (below 0.1 %), the δ15N values were not measured. The values of δ13C (depth: 3,860–3,880 cm) systematically reduce to approximately −29 ‰. In the overlying deposits (maximum depth of 3,810 cm), values of δ13C continue to decrease and attain a minimum of −31.5 ‰ but δ15N fluctuates from 1.5 to 2.4 ‰ (maximum values). From a depth range of 3,745–3,810 cm, a maximum increase in carbon isotopic values approximately −29 ‰ is observed. δ15N is constant of 1.7 ‰. Above 3,740 cm, δ13C fluctuates from −26.2 to −28 ‰ and (beginning at a depth of 3,630 cm) slightly oscillates at about −28 ‰.
Pollen analysis
Palynological data indicate that these sediments accumulated during the late Wartanian Glaciation, the Eemian Interglacial and a subsequent part of the Vistulian Glaciation (Winter and Dobracka 2006; Winter et al. 2008). The 9 local pollen assemblage zones (Rze 1-Rze 9 L PAZ) were distinguished and correlated with regional pollen assemblage zones (E1-EV1 R PAZ) defined by Mamakowa (1989)—Table 1.
Discussion
The ecological preferences of Cladocera were used to reconstruct changes in water level, trophic status and temperature of the lake water of the study paleolake. The isotopic and geochemical data identified the characteristics of organic sediments and defined the source of organic matter.
Final phase of the Wartanian
In the final phase of the Wartanian (maximum depth of 3,970 cm; pollen zones Rze 1 and Rze 2—Winter et al. 2008), the Rzecino lake was formed. Pioneer fauna, which were characterized by a low frequency of Cladocera species (5) and high abundance of specimens (CAZ I), existed in this newly developed, postglacial lake. This composition of Cladocera species indicates cool water with moderate concentrations of nutrients. The cold climatic conditions are confirmed by a threefold increase in NAP values, which are primarily related to an increase in the pollen of Artemisia (maximum of 23 %), Chenopodiaceae, Poaceae, Ericaceae and Betula (Winter and Dobracka 2006; Winter et al. 2008). The significant occurrence of Chydorus sphaericus and Bosmina longirostris (at the depth of 4,010–4,020 cm) is probably associated with a higher trophic level, which suggests an increase in productivity. At that time, the nutrients were supplied to the lake from land. This result is supported by the isotopic composition of lake carbonates, which is typical for dispersed allochthonous carbonates from the underlying morainic till (Mirosław-Grabowska 2008). An intensification of surface runoff is a consequence of the reduced vegetation cover and is indicated by a low pollen abundance and the presence of damaged sporomorphs that predate the Quaternary (Winter et al. 2008).
Eemian Interglacial
Beginning of the Eemian Interglacial (E1–E2)
At the beginning of the Eemian Interglacial (Pinus sylvestris type-Betula zone, pollen zone Rze 3—Winter et al. 2008; regional pollen zone E1, Table 1—Mamakowa 1989), no Cladocera remains were identified. Only at a depth of 3,935 cm, a low abundance of Cladocera specimens and merely 3 species were noted (CAZ II). This finding suggests unfavorable conditions for the development of zooplankton, which may be attributable to the cold water and low water level. During this period, small quantities of organic carbon and nitrogen were detected (Fig. 5). The deposits were characterized by a high value of C/N (18–27) and δ13C values of approximately −26 ‰. The C/N ratios indicate a higher plant component during this period and the low δ13C values may reflect the dominance of C3 plants in the catchment. These isotopic and geochemical data suggest that the organics present in sediments at this time have a predominantly terrestrial origin. At that time, the appearance of heliophilous plants, such as Helianthemum, Hippophae and Ephedra distachya t. correlate with an increase in NAP (primarily Artemisia and Cyperaceae), which indicates cooling of the climate. The high proportion of Artemisia pollen may be attributed to this cooling and high continentality (Winter and Dobracka 2006; Winter et al. 2008). At the end of this pollen period, an improvement in environmental conditions for Cladocera was observed (the beginning of CAZ III). An increase in evaporation and a lowering of the water level in this lake were suggested by oxygen isotopic data (Mirosław-Grabowska 2008). The low-carbonate silts were gradually replaced by authigenic carbonate gyttja.
In the early Eemian Interglacial (Quercus-Fraxinus-Ulmus zone, pollen zone Rze 4—Winter et al. 2008; regional pollen zone E3—Mamakowa 1989), an increase in the number of species and the number of Cladocera individuals is noted, which reflects an improvement in the developmental conditions for fauna (CAZ III). No thermophilic species of zooplankton have been identified in sediments from this period, which suggests the lake was still cold. An increase in the frequency of B. longirostris and B. (E.) coregoni (at a depth of 3,880 cm) may be associated with a high water level (Fig. 4). The concentrations of organic carbon and nitrogen rapidly increase (Figs. 5, 6), which suggests an enhancement of primary productivity and the development of macrophytes in the lake (Ji et al. 2005). C/N rapidly falls to approximately 15 and remains at a level of about 17 (Figs. 5 and 7). The lower C/N ratios indicate a significant proportion of algae in the organic matter. This finding is confirmed by a decrease in δ13C below −30 ‰ (Fig. 5). δ13C values less than −26 ‰ are typical for freshwater algae (Leng et al. 2005). At the end of this pollen period, a rapid decline in the frequency of Cladocera species and an increase in the frequency of Chydorus spp. ephippia (Fig. 3, maximum 10 %) suggest the occurrence of environmental stress (end of CAZ III).

Plot of TOC versus TN of sediments of the Rzecino paleolake. E3-EV regional pollen assemblage zones; E3–E7 Eemian Interglacial, EV early Vistulian

Plot of C/Natomic versus δ13C of sediments of the Rzecino paleolake. E3-EV regional pollen assemblage zones; E3–E7 Eemian Interglacial, EV early Vistulian
Eemian optimum (E4–E5)
During the Eemian optimum, the Corylus-Tilia-Taxus zone (pollen zone Rze 5—Winter et al. 2008; E4—Mamakowa 1989) indicates optimal environmental conditions (warm water and a high level of nutrients) for the development of zooplankton (CAZ IVa–IVb). A maximum diversity in the Cladocera species and the presence of numerous species that prefer warm water—Camptocercus rectirostris, Pleuroxus (P.) truncatatus, Pleuroxus spp., and Graptoleberis testudinaria—are noted. An improvement in climatic conditions is reflected by an expansion of the thermophilous taxa of oak and hazel in forest communities (Winter and Dobracka 2006; Winter et al. 2008).The presence of Graptoleberis testudinaria (at a depth of 3,830 cm, Fig. 2) suggests a higher content of organic matter in the water (Duigan 1992). A significant proportion of benthic species (Monospilus dispar and Alonopsis elongata) most likely reflects a reduction in the water level. High frequencies of Alona affinis, Alona guttata, Eurycercus lamellatus and Monospilus dispar indicate the mesotrophic status of the water. The concentrations of TOC and TN are high and fluctuate between 15–33 % and 1–1.6 %, respectively (Figs. 5, 6). C/N remains at a level of about 17 (Fig. 5). Lower C/N ratios indicate a significant concentration of algae in the organic matter. The δ13C values decrease below −31 ‰ and attain values typical for freshwater algae (Leng et al. 2005). δ15N rapidly increases and attains a maximum value (of 2.4 ‰). The high δ15N values may suggest a high trophic status (Brenner et al. 1999).
In the Carpinus-Picea-Corylus zone (pollen zone Rze 6—Winter et al. 2008; pollen zone E5—Mamakowa 1989) probably a deterioration in the living conditions of zooplankton occurs, which may be due to a reduction in the water level in the lake and/or likely decrease in the pH of the water (presence of Alonella excisa is an indicator of the acidity of the water; Krause-Dellin and Steinberg 1986). At that time, the rapid decrease in the frequency and diversity of Cladocera are observed (CAZ IVc). In these deposits, the content of organic matter significantly increases (maximal value of TOC, Fig. 5). δ13C slightly oscillates at approximately −31 ‰ and δ15N is constant (1.7 ‰). The C/N ratio rapidly increases to 27 (Fig. 5). An increase in TOC and C/N suggest a supply of organic matter of terrestrial origin. The lake most likely began get overgrown at this time (lower water level, higher acidity). Climatic conditions remained favorable. The appearance of the pollen of Taxus, Ilex aquifolium, Tilia platyphyllos t. and Viscum, as well as a more frequent participation of Carpinus and Picea, reflect an increasing amount of rainfall and a more oceanic character of the climate (Winter and Dobracka 2006; Winter et al. 2008).
Eemian post-optimum (E6–E7)
The Eemian post-optimum, Pinus sylvestris type-Picea zone (pollen zone Rze 7—Winter et al. 2008; pollen zone E6—Mamakowa 1989), provided the best conditions for the development of zooplankton. The sudden increase in frequency and diversity of Cladocera is observed. The total abundance of individuals of Cladocera attain a maximum value (CAZ IVd). The composition of Cladocera, especially the presence of species that prefer warmer water (e.g., Camptocercus rectirostris and Pleuroxus trigonellus) and tolerate lower water temperatures (e.g., Acroperus harpae and Eurycercus lamellatus) indicates a moderate, temperature. The water level most likely increases (an occurrence of the Daphnia longispina group and a maximum frequency of Bosmina longirostris and Bosmina (E.) coregoni (Figs. 2, 3). In this period, the composition of organic matter significantly changes. The concentrations of TOC and TN and the C/N ratio rapidly decrease (at the depth of 3,740–3,747 cm, Fig. 5). The opposite trend is observed for δ13C values; these values increase to −28 ‰. The geochemical and isotopic data suggest a reduction of input of higher plants in the lake probably causing the higher clarity of water. In this period, climatic cooling begins, which causes the loss of plant cover. The distinct disappearance of taxa with higher climatic requirements and an increase in the proportion of Pinus sylvestris t. indicate a considerable change in climate (Winter and Dobracka 2006; Winter et al. 2008). The cooling documented by the pollen data, probably lowered the water temperature but it was still around 0 °C in the coldest month (Aalbersberg and Litt 1998). The lower temperature improved the oxidation of the water in the lake resulting also improve the conditions for the plankton development.
During the Pinus sylvestris t. zone (pollen zone Rze 8—Winter et al. 2008; pollen zone E7—Mamakowa 1989), the composition of the Cladocera species and the low frequency of faunal remains (CAZ V) suggest a rapid deterioration in environmental conditions for the development of zooplankton. The presence of species that tolerate cold water and lived with water plant associated, such as Chydorus sphaericus, Acroperus harpae, Alona affinis, Alona guttata, Eurycercus lamellatus imply a reduction in water temperature and development of macrophyte zone. The cooling of the climate is indicated by the disappearance of Quercus, Ulmus, Tilia cordata, Taxus, Hedera and Viscum (Winter and Dobracka 2006; Winter et al. 2008). A decrease in the water level occurs at the beginning of this period, indicated by a high abundance of the benthic Alona quadrangularis and an appearance of Oxyurella tenuicaudis, which indicates the existence of mud-vegetation substrates (Duigan and Kovach 1991) and a decrease in open-water species. Then a change to colder and wetter climates is suggested by a decrease in the values of Pinus sylvestris t. and an increase in the values of Calluna vulgaris, Ericaceae, Cyperaceae and Poaceae (Winter and Dobracka 2006; Winter et al. 2008). At that time, the content of organic carbon and nitrogen is extremely low: 1–3.3 % and 0.1–0.2 %, respectively (Fig. 6). The C/N ratio oscillates between 13 and 17. δ13C fluctuates from −26.2 to −28 ‰ (Fig. 7). The isotopic and C/N values suggest a decline of terrestrial plants in organic matter. At the end of this period, carbonate sedimentation develops, which may be associated with an increase in water level (renew lake deposition) due to an increase in precipitation (Mirosław-Grabowska 2008).
Early Vistulian (EV)
In the Calluna vulgaris-Ericaceae-Betula zone (pollen zone Rze 9—Winter et al. 2008) of the Early Vistulian, the faunal remains are rare but Bosmina spp. ephippia frequently occur (CAZ VI). Bosmina longirostris has effective mechanism of reproduction, is also able to colonize different zones of the lake (Rybak and Błędzki 2010). Due to its thermal tolerance are widely spread and found in the Arctic regions as well as in the warmer ones (Poulsen 1944). Especially Bosmina development is associated with the ecological maturity of the lake to the eutrophic stage. The eutrophication in the Rzeciono paleolake is confirmed by the high proportion of Bosmina longirostris and increase of green algae Botryococcus (Winter et al. 2008). Above a depth of 3,485 cm, no Cladocera remains occur in the sediments. This indicates the occurrence of unfavorable conditions and environmental stress in the lake, which is possibly due to the progressive cooling suggested by pollen data. The large percentage of resting eggs may be due to a significant shortening of the period of favorable conditions for the development of zooplankton. The resting eggs inside ephippia are preserved throughout the winter (or other unfavorable periods) to enable re-establishment of populations in the following spring when environmental conditions will become favorable (Frey 1982; Sarmaja-Korjonen 2003). The highest values of Cyperaceae and Poaceae and a slight increase in Artemisia and Chenopodiaceae indicate an increase in herbaceous communities in open areas and reflects the progressive deterioration of the climate (Winter and Dobracka 2006; Winter et al. 2008). At this time, organic carbon and nitrogen are low, which is similar to conditions in zone E7 (Fig. 6). The C/N ratio rapidly increases to 38 and subsequently decreases to 30. δ13C is constant at approximately −28 ‰ (Fig. 5). This reflects a lack of primary production and water vegetation in the lake during this period. The organic matter was most likely derived from land via runoff. The presence of organic sands (above 3,500 cm) suggests a decline of the Rzecino paleolake (the depth and surface area).
Conclusions
The Rzecino paleolake is the first Polish paleolake dated to the Eemian Interglacial to provide a comparison of zooplankton, geochemical and isotopic data of organic deposits against background vegetation changes. The pollen analysis provides a scheme of the vegetation development and climatic changes during the Eemian Interglacial and the early Vistulian Glaciation in West Pomerania Lakeland (NW Poland). The Cladocera and geochemical data show the environmental conditions that occurred in this lake and confirm the climatic changes postulated by pollen data.
Climatic conclusions
Climatic warming is evident not only from the development of vegetation but also by an increase in primary production and an increase in the abundance and diversity of Cladocera (e.g., the Eemian optimum). Climatic cooling causes not only the disappearance of certain species of plants (e.g., deciduous trees) but also unfavorable conditions for the development of zooplankton and the production of small amounts of organic matter. Our data confirm short-term climatic cooling in the early Eemian Interglacial as suggested by pollen data, e.g., during the Rze 3 Pinus-Betula zone and at the beginning of the Rze 4 Quercus-Corylus-Fraxinus zone (a presence of Cladocera ephippium and a short-term decrease in the curves of thermophilous taxa). During pollen zone Pinus sylvestris t.-Picea (Rze 7 zone; E6), considerable environmental changes are observed in the Rzecino paleolake. The thermophilic plant taxa disappear and the proportion Pinus sylvestris t. increases (Winter and Dobracka 2006; Winter et al. 2008). The frequency and diversity of Cladocera increases (Fig. 4). The composition of organic matter significantly changes, i.e., the TOC and TN and the C/N ratio rapidly decrease but the values of δ13C increase. The changes could be caused by a gradual shift of the climate from interglacial to glacial conditions.
Zooplankton conclusions
The Cladocera species identified in the Eemian sediments from the Rzecino paleolake show close similarity to the Cladocera remains from the Holocene sediments and currently inhabiting Poland and other regions of Europe. The observed changes in the Eemian Cladocera species succession connected with climatic and environmental changes are comparable with contemporary geographic species distribution from north to the south of Europe (Frey 1962).
The Cladocera assemblages from the Rzecino paleolake are analogous to species from other Eemian Interglacial sites in Poland, such as Besiekierz, Studzieniec, Kaliska, Ruszkówek (Mirosław-Grabowska and Niska 2005, 2007a, 2007b; Mirosław-Grabowska et al. 2009). The dominance of the Chydoridae is commonly observed at these sites. The highest number of zooplankton, frequency as well as biodiversity, was recorded during the pollen zone E6-Picea. This increase of remains in Cladocera in paleolake Solniki from NE Poland, Kuców IIc (Niska 2008) and Kaliska from central Poland (Mirosław-Grabowska and Niska 2007a) indicates that a regional environmental factor affected the development of Cladocera succession. The cooling at that time, probably (1) lowered the water temperature and improved the oxidation of the lake water, and (2) blocked the input of higher plants causing a higher water clarity. The higher oxidation and clarity of lake water improved the conditions for the plankton development.
Unlike other Eemian paleolakes, there are no thermophilic species correlated with pollen zone E7-Pinus in the Cladocera succession in Rzecino paleolake. Commonly thermophilic species are present in E7-Pinus and in the successive pollen zone EV (Early Vistulian) in the areas of Sławoszewek—Konin (Pawłowski 2011), of Kaliska (Mirosław-Grabowska and Niska 2007a), of Studzieniec (Mirosław-Grabowska and Niska 2007b), and of Solniki (Niska 2008). Another difference is the presence of a high abundance of ephippia in the sediments, which indicates a progressive cooling and reflects the northerly location of this reservoir (closer to the adjacent growing ice sheet) in relation to other researched lakes. This cooling could shorten the summer (growing period) and stimulate a production of resting eggs (Frey 1982; Sarmaja-Korjonen 2003). In addition, compared with other paleolakes e.g., Studzieniec and Solniki (Mirosław-Grabowska and Niska 2007b; Niska 2008) in paleolake Rzecino occurred lower biodiversity of Cladocera species during the existence of the lake.
Geochemical conclusions
The geochemical and isotopic characteristics of organic matter from the Eemian deposits have not been previously analyzed in Poland and in the other European countries. We have deemed it useless to correlate our data with records of different climatic regions, such as Asia, due to the different lake origins, and to significant differences in climatic conditions that result in different vegetation and zooplankton development. Rzecino paleolake is excellent example of the influence of climatic conditions caused by geographical uniqueness of the local lake environment.
At the Eemian Interglacial beginning, the small amount of organic matter accumulated in Rzecino paleolake was mainly of terrestrial origin. When an improvement in environmental conditions occurred (pollen zone E3), an enhancement of primary productivity and the development of macrophytes in the lake took place. During the Eemian optimum, algae remained a significant component of OM. In the Eemian post-optimum (pollen zone E6), the abundance of higher aquatic plants declined and then (the pollen zone E7) a terrestrial plant delivery inhibited into the paleolake. During the Early Vistulian (pollen zone EV1), the deterioration of lake conditions and a lack of primary production and water vegetation in the lake took place.
References
Aalbersberg G, Litt T (1998) Multiproxy climate reconstructions for the Eemian and Early Weichselian. J Quat Sci 13:367–390
Adamska A, Mikulski JS (1969) Cladocera remains in the superficial sediments of lakes as a typologic indicator. Z. Naukowe UMK 25. Prace Stacji Limnolog w Iławie 5:41–48
Brenner M, Whitmore TJ, Curtis JH, Hodell DA, Schelske CL (1999) Stable isotope (δ13C and δ15N) signatures of sedimented organic matter as indicators of historic lake trophic state. J Paleolimnol 22:205–221
Dobracka E (2001) Detailed Geological Map of Poland at the 1:50 000 scale, Dobrowo sheet. Cent Arch Geol Pan Inst Geol, pp 1–40
Duigan CA (1992) The ecology and distribution of the littoral freshwater Chydoridae (Branchiopoda, Anomopoda) of Ireland with taxonomic comments on some species. Hydrobiologia 241:1–70
Duigan CA, Kovach WL (1991) A study of distribution and ecology of littoral freshwater chydorid (Crustacea, Cladocera) communities in Ireland using multivariate analysis. J Biogeogr 18:267–280
Flössner D (1964) Zur Cladoceranfauna des Stechlin-Gebietes. II. Őkologische Untersuchungen über die litoralen Arten. Limnologica 2(1):35–103
Flössner D (2000) Die Haplopoda und Cladocera (ohne Bosminidae) Mitteleuropas. Backhuys, Leiden 428
Frey DG (1958) The Late Glacial cladoceran fauna of a small lake. Arch Hydrobiol 54:209–275
Frey DG (1962) Cladocera from the Eemian Interglacial of Denmark. J Paleontol 36:1133–1154
Frey DG (1982) Contrasting strategies of gamogenesis in northern and southern populations of Cladocera. Ecology 63:223–241
Frey DG (1986) Cladocera analysis. In: Berglund BE (ed) Handbook of Holocene Palaeoecology and Palaeohydrology. Wiley, Chichester, pp 667–692
Goulden CE (1964) The history of the cladoceran fauna of Esthwaite Water (England) and its limnological significance. Arch Hydrobiol 60:1–53
Hofmann W (1986) Developmental history of the Grosser Plöner See and the Schöhsee (north Germany): cladoceran analysis, with special reference to eutrophication. Arch Hydrobiol Suppl 74:259–287
Hofmann W (1987) Cladocera in space and time: analysis of lake sediments. Hydrobiologia 145:315–321
Hofmann W (2000) Response of the chydorid faunas to rapid climatic changes in four alpine lakes at different altitudes. Palaeogeogr Palaeoclimatol Palaeoecol 159(3–4):281–292
Ji S, Xingqi L, Sumin W, Matsumoto R (2005) Palaeoclimatic changes in the Qinghai Lake area during the last 18,000 years. Quat Inter 136:131–140
Korhola A (1990) Paleolimnology and hydroseral development of the Kotasuo Bog, Southern Finland, with special reference to the Cladocera. Ann Acad Sci Fenn 155:5–40
Korhola A, Rautio M (2001) Cladocera and other branchiopod crustaceans. In: Smol JPH, Birks JB, Last WM (eds) Tracking environmental change using lake sediments, vol 4., Zoological indicatorsKluwer Academic, Dordrecht, pp 5–41
Krause-Dellin D, Steinberg C (1986) Cladoceran remains as indicators of lake acidification. Hydrobiologia 143:129–134
Kupryjanowicz M (2008) Vegetation and climate of the Eemian and Early Vistulian lakeland in northern Podlasie. Acta Palaeobot 48(1):3–130
Leng MJ, Lamb AL, Marshall JD, Wolfe BB, Jones MD, Holmes JA, Arrowsmith C (2005) Isotopes in Lake Sediments. In: Leng MJ (ed) Isotopes in Palaeonvironmental Research. Springer, The Netherlands, pp 147–184
Mamakowa K (1989) Late Middle Polish Glaciation, Eemian and Early Vistulian vegetation at Imbramowice near Wrocław and the pollen stratigraphy of this part of the Pleistocene in Poland. Acta Palaeobot 29:11–176
Meyers PA, Ishiwatari R (1993) Lacustrine organic geochemistry-an overview of indicators of organic matter sources and diagenesis in lake sediments. Org Geochem 7:867–900
Meyers PA, Lallier-Vergès E (1999) Lacustrine sedimentary organic matter records of Late Quaternary paleoclimates. J Paleolimnol 21:345–372
Mirosław-Grabowska J (2008) Reconstruction of lake evolution at Rzecino (NW Poland) during the Eemian Interglacial and Early Vistulian on the basis of stable isotope analysis. Ann Rep Pol Acad Sci 2008:90–92
Mirosław-Grabowska J, Niska M (2005) Isotopic and Cladocera records of climate changes of Early Eemian at Besiekierz (Central Poland). Geolog Quart 49:67–74
Mirosław-Grabowska J, Niska M (2007a) Isotope and Cladocera data and interpretation from the Eemian optimum and postoptimum deposits, Kaliska palaeolake (Central Poland). Quat Int 175:155–167
Mirosław-Grabowska J, Niska M (2007b) Reconstruction of environmental conditions of Eemian palaeolake at Studzieniec (Central Poland) on the basis of stable isotope and Cladocera analyses. Quat Int 162–163:195–204
Mirosław-Grabowska J, Niska M, Sienkiewicz E (2009) Evolution of the palaeolake at Ruszkówek (central Poland) during the Eemian Interglacial based on isotopic, cladoceran and diatom data. J Paleolimnol 42:467–481
Mueller WP (1964) The distribution of cladoceran remains in surficial sediments from three northern Indiana lakes. Invest. Indiana Lakes Streams 6: l–63
Nalepka D, Walanus A (2003) Data processing in pollen analysis. Acta Palaeobot 43(1):125–134
Niska M (2008) Interpretacja zmian środowiska jeziornego w interglacjale eemskim na podstawie analizy kopalnych Cladocera. PhD thesis, Archiv Akademia Pomorska w Słupsku
Pawłowski D (2011) Evolution of an Eemian lake based on Cladocera analysis (Konin area, Central Poland). Acta Geolog Polon 61(4):441–450
Poulsen E (1944) Entomostraca from a late-glacial lacustrine deposit at Næstved, Denmark. Meddelelser fra Dansk Geologisk Forening 10:405–416
Rybak JI, Błędzki LA (2010) Słodkowodne skorupiaki planktonowe. Klucz do oznaczania gatunków. Wyd UW, Warszawa, pp 1–368
Sarmaja-Korjonen K (2003) Chydorid ephippia as indicators of environemtal change-biostratigraphical evidence from two lakes in southern Finland. Holocene 13:691–700
Shackleton NJ, Sánchez-Goñi MF, Pailler D, Lancelot Y (2003) Marine Isotope Substage 5e and the Eemian Interglacial. Glob Planet Chang 36:151–155
Smeltzer E, Swain EB (1985) Answering lake management questions with paleolimnology. In: Lake and reservoir management—practical applications. In: Proceedings of the 4th annual conference and international symposium. North America Lake Manag Soc, pp 268–274
Szeroczyńska K (1985) Cladocera jako wskaźnik ekologiczny w późnoczwartorzędowych osadach jeziornych Polski Północnej (Cladocera as ecologic indicator in late Quaternary lacustrine sediments in Northern Poland). Acta Palaeontol Polon 30(1–2):3–69
Szeroczyńska K, Sarmaja-Korjonen K (2007) Atlas of Subfossil Cladocera from Central and Northern Europe. Tow Przyj Dolnej Wisły, Świecie, pp 1–84
Tyson RV (1995) Sedimentary organic matter: organic facies and palynofacies. Chapman and Hall, London
Whiteside MC (1970) Danish chydorid Cladocera: modern ecology and core studies. Ecol Monogr 40:79–118
Whiteside MC, Swindoll MR (1988) Guidelines and limitations to cladoceran paleoecological interpretations. Palaeogr Palaeoeclim Palaeoecol 62:405–412
Winter H, Dobracka E (2006) Multidisciplinary studies of Eemian and early Vistulian sediments at Rzecino site (Łobez Upland—Western Pomerania Lakeland). International Conference on Living Morphotectonics of the European Lowland, 28–30 August 2006, Cedynia, Poland. Abstracts, pp 98–110
Winter H, Dobracka E, Ciszek D (2008) Multiproxy studies of Eemian and early Vistulian sediments at Rzecino site (Łobez Upland, Western Pomerania Lakeland). Biul Pan Inst Geol 428:93–110
Acknowledgments
We would like to express our thanks to Dr. Hanna Winter for providing the sediment samples and very interesting cooperation. The studies were financed by the Pomeranian University in Słupsk (Rector grant) and the Institute of Geological Sciences Polish Academy of Sciences. The obtained data contribute to research project “Reconstruction of the development of lake environmental in the Eemian interglacial based on subfossil Cladocera (Crustacea) analysis” (Foundation for Polish Science, Bridge Program 2012).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Niska, M., Mirosław-Grabowska, J. Eemian environmental changes recorded in lake deposits from Rzecino (NW Poland): Cladocera, isotopic and selected geochemical data. J Paleolimnol 53, 89–105 (2015). https://doi.org/10.1007/s10933-014-9810-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-014-9810-6