
Abstract
Context
Spatial conservation prioritization (SCP) has most often been applied to the design of reserve network expansion. In addition to occurrences of species and habitats inside protected area candidate sites, one may also be interested about network-level connectivity considerations.
Objectives
We applied SCP to the identification of ecological networks to inform the development of a new regional plan for the region of Uusimaa (South-Finland, including the Finnish capital district).
Methods
Input data were 59 high-quality layers of biotope and species distribution data. We identified ecological networks based on a combination of a Zonation balanced priority ranking map and a weighted range size rarity map, to account for both relative and absolute conservation values in the process. We also identified ecological corridors between protected areas and other ecologically high-priority areas using the corridor retention method of Zonation. Furthermore, we identified candidate sites for habitat restoration.
Results
We found seven large ecological networks (132–1201 km2) which stand out from their surrounding landscape in terms of ecological value and have clear connectivity bottlenecks between them. Highest restoration needs were found between large high-priority sites that are connected via remnant habitat fragments in comparatively highly modified areas.
Conclusions
Land conversion should be avoided in areas of highest ecological priorities and network-level connectivity. Restoration should be considered for connectivity bottlenecks. Methods described here can be applied in any location where relevant spatial data are available. The present results are actively used by the regional council and municipalities in the region of Uusimaa.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The current biodiversity crisis demands that biodiversity should be systematically accounted for in land-use planning (Newbold et al. 2015). A key question in biodiversity protection is how landscape connectivity should be considered; a debate that still continues (Gippoliti and Battisti 2017; Foltête 2019; Miller-Rushing et al. 2019). Biodiversity-friendly land-use planning requires information about connectivity for ensuring sustainability of populations (Opdam et al. 2006; Hodgson et al. 2009) and maximizing the benefits of restoration for the entire landscape (Volk et al. 2018). Requirements for ecological connectivity are repeated in global (CBD 2010), continental (European Commission 2011), and national policy documents (e.g. the Finnish Biodiversity Action Plan 2012), but accounting for it systematically in operational land-use planning is difficult (Boitani et al. 2007). Building on a real-life regional planning case, this paper introduces a method for identifying well-connected ecological structures in a human-modified landscape to inform land-use planning.
Land-use planning should account for all fundaments of ecology that determine the carrying capacity of a landscape for local species populations, habitat area, quality, and connectivity, in order to effectively preserve biodiversity (Hodgson et al. 2009, 2011). Out of those, habitat area and quality are the main factors for preservation of populations and they should be given a clear priority over connectivity (Hodgson et al. 2011). Recent analyses have concluded that the importance of connectivity based on island biogeography has generally been overestimated at least to some extent (Fahrig 2013; Martin 2018; Wintle et al. 2018). Connectivity itself essentially derives from the pattern of habitat quality over space (Hodgson et al. 2009). Habitat quality is a continuum that ranges from completely unsuitable to optimal reproduction areas (Fischer and National 2006). Somewhere in-between these two extremes are environments that support dispersal but not reproduction (Puth and Wilson 2001; Fischer and National 2006; Hodgson et al. 2009; Moilanen 2011). What counts as suitable for dispersal is specific to species, which makes specification of strict connectivity definitions and recommendations difficult in operational land-use planning (Puth and Wilson 2001; Chetkiewicz et al. 2006; Boitani et al. 2007; Gippoliti and Battisti 2017). Furthermore, ecological corridors, while greatly emphasized in conservation and land-use planning, are merely one special case of path-like connectivity (Chetkiewicz et al. 2006; Rayfield et al. 2011). In reality, an area defined as a corridor can act as a dispersal route for some species, as breeding habitat to others, and neither to some (Puth and Wilson 2001; Chetkiewicz et al. 2006). Identifying corridors that best maintain overall biodiversity is therefore a challenging task in land-use planning (Puth and Wilson 2001; Boitani et al. 2007).
While connectivity itself is highly complicated and species-specific, complexities around the application of connectivity can become greatly reduced when the contrast between natural and human-modified areas is high. In this case, the human-modified parts of the landscape are of little value to most species and habitat quality, connectivity and biodiversity have become highly concentrated in the remaining less degraded part of the landscape (Opdam et al. 2006; Prugh et al. 2008; Reider et al. 2018). In these areas, ecological networks can be considered as large, semi-continuous, remnants of biodiversity concentrations that stand out from the more degraded environments. Fragmentation of these areas should be avoided. Ecological corridors, on the other hand, can be most clearly distinguished as bottlenecks between high-quality biodiversity areas through environments of significantly reduced quality. These parts of the landscape are most important for regional-scale connectivity of species that are influenced by fragmentation, i.e., species that avoid human-modified landscapes during dispersal (Puth and Wilson 2001; Fischer and National 2006). With this approach, the aim becomes identification of connectivity bottlenecks that should not be weakened, rather than search for corridor-like structures per se.
Spatial (conservation) prioritization has developed from the need for spatial conservation planning, including in the context of land use planning (Kullberg and Moilanen 2014). Spatial prioritization allows biodiversity to be assessed systematically, and in a well-balanced manner, across the region of interest (Lehtomäki and Moilanen 2013). Originally, spatial prioritization (reserve selection; reserve network design) was developed for the planning of protected area networks (Margules and Pressey 2000). Later on, methodology has been expanded to cover both conservation and environmental impact avoidance (Kareksela et al. 2013), with obvious implications for general land use planning and zoning. Given that over 80% of the world is unprotected, impact avoidance is arguably an even more important application of prioritization than planning of conservation area networks.
Connectivity can be accounted for in many ways in spatial prioritization, but most methods would focus on comparatively simple forms of aggregation rather than evaluation of network-level connectivity. Many methods base on metapopulation theory (for distance-scaled aggregation) or different edge penalties (for structural compactness or unity) (Moilanen et al. 2009). On the other hand, graph-theoretic approaches are better in the identification of network-level connectivity and e.g. connectivity bottlenecks (Rayfield et al. 2011; Correa Ayram et al. 2016). However, on their own these connectivity methods miss many factors inherent to spatial prioritization, including the balance over a great number of biodiversity features, inclusion of costs and threats, or analyses on high-resolution grids (Moilanen 2011). Spatial prioritization has also been coupled with graph-theoretic or least-cost methods with the aim of locating important ecological networks (Rouget et al. 2006; Albert et al. 2017; Álvarez-Romero et al. 2018; Meurant et al. 2018).
Here, we show how to use the spatial prioritization approach software, Zonation, to directly identify network-level connectivity to support regional planning. Our study area is the province of Uusimaa (South-Finland), which includes the capital district of Helsinki. We show how to use Zonation to identify comparatively well connected ecological networks, to identify corridor-like elements, and to find restoration opportunities. Furthermore, we assessed the expected connectivity effects of a proposed regional plan. Assuming that data typically underlying spatial prioritization are available (Kullberg and Moilanen 2014; Kujala et al. 2018), our methods can be replicated anywhere.
Materials and methods
Study area and Uusimaa 2050 regional plan
The province of Uusimaa (henceforth Uusimaa) is located at the Southern coast of Finland and includes the capital district of Helsinki. With 1.7M people, it is the most populous province in Finland. Uusimaa covers an area of 9600 km2, including heavily-modified areas such as cities, towns, and agricultural and forestry areas. It also hosts significant natural values: old-growth forests, mires, coastal habitats, rivers, and other areas in natural or semi-natural state. There are many national parks, Natura 2000 areas, and other types of protected areas in the region (Regional Council of Uusimaa 2018).
In Finland, regional councils are responsible for province-level regional zoning that steers municipal land-use planning. The Regional Council of Uusimaa started to develop a new regional plan in 2016 (Regional Council of Uusimaa 2018). This so-called Uusimaa 2050 plan aims to accommodate 500,000 new people in Uusimaa by 2050, while balancing preservation of ecological and cultural values. The plan is strategic and aims to define only the key zones for regionally balanced development. The plan also defines ecological connections that must be accounted for in further land-use planning. To support this aim, information of ecological networks and connections was produced using spatial prioritization. The original report is in Finnish by Jalkanen et al. (2018a).
Zonation analysis for supporting regional planning in Uusimaa
Our study builds directly upon spatial prioritizations that were done in 2015 to inform regional planners about biodiversity priority areas in Uusimaa for the previous version of the regional plan (established in 2017; Regional Council of Uusimaa 2017), shown in Fig. 1. These prioritizations were implemented with the Zonation software (Moilanen et al. 2005; Lehtomäki and Moilanen 2013), which produces a complementarity-driven priority ranking of the given landscape. First, it assumes that ecologically best would be to have the full landscape protected. Then, it identifies, ranks and removes the grid cell that can be given up with smallest aggregate loss for biodiversity, conditional on what remains in the landscape. This step is iterated until the entire landscape has been ranked. It is fundamental to Zonation that it maintains full dimensionality of biodiversity through the process, which enables it to maintain a balance between all species and other biodiversity features throughout. (The balancing is based on the tracking and use of remaining distribution sizes in the calculations.) As a result, a priority ranking is produced, ranging from 0 (least importance for balanced maintenance of biodiversity features) to 1 (highest importance). Connectivity can also be incorporated into Zonation process in several ways (Lehtomäki and Moilanen 2013).

Zonation priorities in the Uusimaa province by Kuusterä et al. (2015). As shown by the mean performance curve, biodiversity is rather aggregated in fragmented Uusimaa as the top 20% of the landscape (dark red) hold roughly 70% of the distributions of biodiversity features (note that distributions do not start from full 100% because the use of a condition layer lowers the initial distribution levels of all features). Generally, biodiversity priorities are biased towards the western parts of the province, although top-priority areas do exist also in the east. A clear weakened zone can be seen in central Uusimaa (dark blue) due by major cities, a major motor way, the main railroad, and heavy agriculture. This zone was used to divide Uusimaa to eastern and western parts for the corridor analyses (see “Corridor-Zonation for structural connectivity through bottlenecks” section). Furthermore, a 15 km buffer area was included in corridor analyses, which also included some “new” top-priority sites near the provincial border. (Color figure online)
The prioritizations in Uusimaa have been described (in Finnish) by Kuusterä et al. (2015). They included 59 layers of information about habitat and species distributions and geodiversity across Uusimaa (Supplementary Table S1). The input data consists of various high-quality biodiversity data currently used by the Finnish environmental administration, and they were originally sourced from several organizations including national, regional, and municipal authorities and environmental NGOs. The present work is based on analysis variant 5 of Kuusterä et al. (2015), which includes species and habitats as input data, matrix connectivity between major habitat types, and effects of current land-use generalized for all features as a condition layer. See Supplementary Tables S1–S3, and Kuusterä et al. (2015) for a detailed description of the input data and the Zonation settings.
Two spatial Zonation outputs: complementarity-based rank and scoring-based weighted range-size rarity
The present work combines two different Zonation outputs in a manner that has not been used before. These are the standard priority rank map and the weighted range-size rarity map, which was called “weighted range size normalized richness” starting from first version of Zonation (Moilanen et al. 2005; the .wrscr layer). The priority ranking ranks all areas between 0 and 1 and it has a special characteristic that any given top or bottom fraction of the map holds a balanced coverage across all biodiversity features (etc.) included in analysis. A ranking will be developed even if there are relatively small differences between areas (the performance curves provide additional quantitative information). In comparison, the weighted range size rarity map is completely different (Williams 2000; Veach et al. 2017). It is a weighted sum of the fractions of features occurring in a grid cell, which means the map has a clear interpretation in the absolute scale. However, it has a deficiency that all biodiversity information has been squashed into one dimension (by a weighted sum), which means that e.g. top areas chosen from a range-size rarity map do not necessarily hold a balanced coverage across features. This would be most apparent when the study area includes multiple environments (forest, mire, wetland), which have very different levels of species richness and endemicity. Consequently, we here use a combination of the priority rank map and the weighted range size rarity map as the basis of identifying ecological networks.
Identification of well-connected networks
According to the shape of the Zonation performance curves (Fig. 1), biodiversity is rather aggregated in Uusimaa. The top 20% and 40% priority areas of the landscape harbor 61.6% and 74.7% of the input feature distributions, respectively. Therefore, the loss of top 20% priority areas would result in a rapid loss of biodiversity values, whereas land-use changes in the lowest 60% priority areas will have a comparatively mild impact on biodiversity feature distributions. These landscape fractions were therefore used as robust thresholds for core (top 20% priority) and supporting (top 40–20% priority) biodiversity areas.
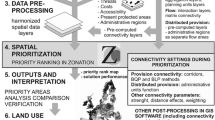
Figure 2 describes the workflow for the identification of ecological networks. To gain information on both the amount of biodiversity in areas and the complementarity between different areas, we combined the Zonation rank and wrscr layers with the Raster Calculator in ArcGIS 10.3. (ESRI, Redlands, CA, USA) by multiplying the log-transformed wrscr layer with the rank layer (see Supplementary Information S2.1 for full syntax). The log-transformation was made to broaden the range of values visible on a computer screen so that we were able to visually delineate ecological networks from otherwise very skewed raster map (Supplementary Information S2.1.; Supplementary Fig. S1). We then used the Focal Statistics tool in ArcGIS 10.3. for kernel-type smoothing of the resulting ‘balanced feature density layer’ using a declining-by-distance smoothing kernel that ended at zero at a 2 km radius (Supplementary Fig. S2). The distance of 2 km was chosen so it is relevant for the identification of regional ecological networks (Supplementary S2.2.; Supplementary Fig. S3). Following the thresholds identified from the previous Zonation performance curves (Kuusterä et al. 2015), we then separated the top 20% and 40% areas from the balanced feature density map to locate the core and supporting areas of the networks, respectively. The resulting map describes large and semi-continuous aggregations of biodiversity, with focus on both absolute distribution and complementarity (balance) between features inside the networks.

The workflow for identifying large ecological networks using the Zonation rank and weighted range size rarity (wrscr) layers. The final networks are contiguous areas that belong to top-fractions of the aggregated balanced feature density layers (i.e. harbor much biodiversity in an aggregated manner) and harbor great amount of top-priority sites (i.e. constitute to balanced coverage of biodiversity feature distributions in Uusimaa)
The large networks were then defined using the following criteria: they consisted of continuous surfaces of top 20% and 40% areas of the balanced feature density layer, they included a high density of top 20% and 40% priority ranking areas (from earlier analysis), and they were clearly distinguishable from the surrounding landscape, which has been modified by human activity. Areas close to a shoreline involved special treatment, because these areas had a negative bias in the smoothed, balanced feature density map. (Water bodies were not included in analysis, which lead to lowered estimated value near water when smoothing using the Focal statistic tool.) Consequently, the large ecological networks were identified semi-manually from the top-fractions of the balanced feature density map (Fig. 2). The human eye is very proficient in edge effect correction and in identifying networks that stand out from the rest of the landscape.
Once the regional networks were defined, we used landscape identification analysis (LSM) in Zonation (Moilanen et al. 2005, 2014) to quantify and characterize biodiversity in each network. Furthermore, to compare biodiversity concentration between networks, we calculated a feature density index, the density of features in the network compared to what the density would be if features were randomly distributed across the landscape. This index was calculated for network j as
where Sj is the sum of weighted feature distribution fractions in the network j (received from the LSM analysis; see Moilanen et al. 2014), Aj is the area of the network j, St is the weighted feature distribution sum of the entire study landscape (in our case, Uusimaa), and At is the area of the entire landscape. The feature density index thus compares concentration of biodiversity in a regional network to the average across the full study area.
Corridor-Zonation for structural connectivity through bottlenecks
To locate structural connectivity across the connectivity bottlenecks between the large ecological networks, or subnetworks inside them, we ran a new Zonation analysis using the corridor building method (henceforth Corridor-Zonation; Pouzols and Moilanen 2014). Corridor-Zonation utilizes a penalty for decrease in structural connectivity that is embedded in the general prioritization process, i.e. it allows balancing between local habitat quality and structural connectivity without any pre-defined habitat patches or starting-points for corridors. Corridor-Zonation is one of Zonation’s connectivity methods, and it can be used together with many other Zonation methods and settings (Pouzols and Moilanen 2014).
The two main parameters of Corridor-Zonation are corridor width and the strength of the corridor loss penalty (Pouzols and Moilanen 2014). In the case of Uusimaa, a 300 m corridor width was considered appropriate after discussions with the regional planners. We tested different options for the corridor loss penalty (Supplementary S3.1.; Supplementary Fig. S4) and eventually came to use a case-specific value of 0.0001. Our test runs showed that the locations of the corridors had only minor variation between different Corridor-Zonation variants whereas the priorities given for the corridors varied (higher corridor loss penalty translates into higher priorities for the corridors themselves). As we were not interested of the priorities of the corridor areas per se but rather their locations, we then used a value sufficiently high to make the corridors stand out from the landscape (see below).
Our Corridor-Zonation analysis included the same input data and weightings as described by Kuusterä et al. (2015). We also used the same matrix connectivity setting between habitat types as well as the effects of the current land-use as condition layer (Supplementary S1). We excluded lakes from analysis. However, including Corridor-Zonation required some changes to data and analysis settings. Because we wanted to avoid edge effects (Zonation not finding corridors to high-priority patches just outside regional borders), our analysis area included a 15 km buffer around Uusimaa. (The same biodiversity data was available for the buffer area).
In the present work, the primary function of ecological corridors was taken to be connection of the most secure and ecologically important habitat patches (core areas), including ecologically high-quality protected areas. To facilitate this outcome, we used a (standard) hierarchical analysis structure, where core areas had highest priority (Mikkonen and Moilanen 2013). Core areas were defined as the top 20% priority areas that were larger than 50 ha, which was considered by the local regional planners as the standard minimum size for regionally important nature areas. Protected areas were not exclusively included to the hierarchical mask, as the main question was about remaining ecological networks irrespective of present conservation status. Consequently, Corridor-Zonation “focused” on maintenance of corridors between those core areas. Furthermore, test analyses showed that corridors were highly biased towards western Uusimaa (Supplementary S3.2.; Supplementary Fig. S5), because of the overall higher aggregation of biodiversity in the western parts of the region (as can be seen in Fig. 1). As corridors should nevertheless benefit the local ecological networks throughout the whole region, we did the prioritization separately for the eastern and western parts. The division between the subareas (Fig. 1) followed the main rail- and motorway corridor accompanied by residential zones and intensive agriculture, which constitute a significant barrier to the movement of many if not most animals. Finally, to avoid locating utterly unrealistic corridors through densely built areas (e.g. through city centers), we set Corridor-Zonation to only start maintaining corridors after the lowest 15% fraction of Uusimaa had been prioritized and removed from analysis.
The identification of corridor-like ecological connections was done by inspection of the difference between the basic Zonation analysis and corridor Zonation priority rank maps, separately for eastern and western Uusimaa. We identified three kinds of ecological connections (Fig. 3): (i) linear connections that belonged to the top 20% Zonation priorities already based on biodiversity alone (mainly riverbanks), (ii) homogeneous areas of high connectivity, and (iii) corridors. Linear top-priority sites were defined as long and narrow elongated areas that belonged to the highest 20% priority of Uusimaa. Homogeneous areas of high connectivity were areas that belonged effectively homogeneously to the ecological mid-to-high priorities of Uusimaa, and where Corridor-Zonation had located dense concentrations of corridors. We interpreted that those areas had overall high potential for connectivity, but locating single narrow corridors within them would have been arbitrary due to the high number of alternative connectivity routes. Corridors were narrow structures that could be clearly distinguished from the Corridor-Zonation results and that connected core biodiversity areas (top 20% priority areas, min. 50 ha) and/or homogeneous areas of high connectivity.

Different types of ecological connections, identified by comparing the regular Zonation rank (a) with the Corridor-Zonation rank (b). Different connection types (c) include: linear elements that belong to the top-priorities of Uusimaa; homogeneous areas of high connectivity which show clear concentrations of corridors (i.e. areas that Corridor-Zonation generally recognizes beneficial for structural connectivity); and ecological corridors that are narrow corridor-shape elements, clearly distinguished in Corridor-Zonation rank map, which combine large top-priority areas and/or homogeneous areas of high connectivity
To aid land-use planners to focus on the most relevant connections in the strategic plan, we finally identified key ecological connections throughout Uusimaa. Key connections join large ecological networks, or large subnetworks, across otherwise degraded zones. Key connections also have few, if any, options remaining in the landscape. All different types of ecological connections (Fig. 3) could act as parts of the key connections.
Restoration needs of the structural connections
We defined the restoration needs of the ecological connections simply as the difference between Zonation ranks with and without Corridor-Zonation. The resulting layer describes restoration need from high to low, with high need identified for connections that are degraded but have high priority in corridor Zonation. This means a degraded piece of land is the best remaining connection between two areas of significant biodiversity content.
Assessing connectivity in the Uusimaa 2050 plan
The large ecological networks, as well as structural connections and their restoration needs were used as a background information for regional planning. After the initial Uusimaa 2050 plan proposal (Regional Council of Uusimaa 2018) had been compiled, we assessed the plans’ expected impacts on ecological connectivity against both the present results and the earlier analysis of biodiversity core areas by Kuusterä et al. (2015). To quantify the possible impacts of the plan to the networks, we did a new Zonation post-processing analysis (LSM), which extracts both summary and detailed biodiversity data for areas where networks and development zones intersect. For visualization, we did a post-hoc GIS overlay of the ecological networks and connections against the proposed new development zones.
The primary focus of the 2050 assessment was to identify locations where ecological core areas (top-priority sites) or large ecological networks might be damaged. Here we focus on the results concerning ecological networks, because overlays with the core areas are easy to interpret and biodiversity found in the core areas is not the focus of the present study. Furthermore, we assessed the adequacy of green connections proposed in the plan against our Corridor-Zonation results. Special focus was given to a “green belt” suggested around the capital district.
Results
Ecological networks and connections
We identified seven large ecological networks (132–1201 km2) that consist of aggregated mosaics of high-priority biodiversity areas. These networks span many municipalities and may enclose heavily-modified areas such as small towns or agricultural fields (Fig. 4a). These networks host a total of 63% of the distributions of biodiversity features of Uusimaa. Isolated high-priority sites also exist outside these networks.

a Seven large ecological networks in Uusimaa. These networks are highly aggregated and host a high proportion of the biodiversity found in Uusimaa. Small, isolated, high-priority areas are located outside the large networks. Local land-use planning should try to maintain connectivity inside and between the large networks, in addition to preserving the top-priority areas throughout Uusimaa (including the smaller and isolated ones). b Example of a key connection between the Sipoonkorpi and Porvoo networks, the only connection between these two networks along the shoreline
Table 1 characterizes the major ecological networks of Uusimaa. In general, biodiversity values are biased towards western Uusimaa: the networks are larger, they include higher proportions of top-priority sites, and they have higher feature density than the eastern networks. However, also the eastern networks are clearly distinguished from the surrounding landscape, as their feature density indices are 61–76% higher the average of Uusimaa. Therefore, all major networks identified here should be considered at least locally important for the ecological landscape of Uusimaa.
Some biodiversity features such as forests are found in all networks, but some features are characteristic to individual networks (Table 1). Examining features covered by networks provides useful information for local land-use and conservation planning. For example, the West-Uusimaa network harbors over 70% of the known natural sand beaches in Uusimaa, so they should be a specific target for biodiversity preservation along the western coast of Uusimaa.
Use of Corridor-Zonation in prioritization had only minor effect on the coverage of features by the network: at greatest, the difference between the average feature distributions was 0.0097%-units in western and 0.0087%-units in eastern Uusimaa when 81% and 78% of the landscape were prioritized, respectively. This implies that Corridor-Zonation did not identify corridors at significant expense in terms of coverage of biodiversity features. (Some loss is expected by default, as some corridors may need to traverse sections of land that have moderate human impacts already.)
We identified many important ecological connections in Uusimaa (Supplementary Fig. S6). Some connections also cross regional borders to neighboring provinces, especially to the north-west and east. Key connections (Fig. 4b) that should be secured in land-use planning have few alternatives and connect major networks and large subnetworks. They were especially concentrated in western and northern parts of Uusimaa, coastline, and urban fringe of the capital district.
Identification of connectivity restoration needs
The estimated restoration needs for ecological corridors are highest in areas that connect high-priority sites (i.e. important for structural connectivity in the landscape) through modified, comparatively low-priority areas (Supplementary Fig. S7). A low restoration need would be identified when a corridor would be identified between generally low-priority areas. Also a high-quality connection between high-quality areas would not show, as there is no need for restoration. Note that this analysis does not tell which restoration actions should be carried out in a given site and there may be sites where restoration is not feasible. Choosing appropriate management is always case-specific and can vary from allowing of passive recovery to one-time restoration action to continuous management (Fig. 5).

Examples of corridors that would require different restoration actions. In (a), forest connection goes through a managed forestry site. The connection would most benefit from limitations on forest management and harvesting. In (b) a forest connection finds the only remaining path through agriculture fields. Reforestation of those fields would enhance the structural connectivity of forests at that location
Assessment of the Uusimaa 2050 plan proposal
The Uusimaa 2050 plan emphasizes densification of current residential zones and the growth inside the capital district. The plan also proposes new or upgraded highways and railways. Some green connections and a green belt around the capital district are proposed, motivated by recreation and maintenance of the dispersal routes of animals. Nevertheless, the extent of these connections has been reduced from the previous regional plan.
Our connectivity analyses enabled a general-level connectivity impact assessment of the Uusimaa 2050 plan proposal (for full report and maps see Jalkanen et al. 2018b). Table 2 summarizes the expected losses of biodiversity inside the large ecological networks. Out of the 21 current residential zones, 18 are allowed to expand into large ecological networks. Out of the 9 proposed new residential zones, 6 overlap large networks, potentially reducing their area by 5800 ha (1.3%) in total. The most significant losses would be expected around the Nuuksio and Sipoonkorpi networks at the fringes of the capital district (Table 2, Fig. 6). In the north, expansion of the town of Hyvinkää eats into the already narrow bottleneck right in the middle of the North-Uusimaa network (Fig. 6). New proposed highways and railways would fragment especially Nuuksio, Sipoonkorpi, and North-Uusimaa networks, inside of which the plan proposes a total of 178, 91, and 56 new highway or railway kilometers, respectively.

Overlay of the large ecological networks and proposed expanding future land-use in the Uusimaa 2050 plan. The overlay shows that new residential zones would harm ecological connectivity in several locations, particularly the western and eastern sides of the capital district, and in the middle of the North-Uusimaa network (see also Table 2). The proposed new highways and railways interact with impacts from the new residential zones and would contribute to fragmentation of the large networks. Regional green areas are excluded from this figure for the sake of visual clarity
Figure 7 shows Corridor-Zonation results against the Uusimaa 2050 plan proposal’s green areas and green connections. Direct comparison is, however, difficult because the plan is very general-scale and strategic and it is not supposed to show exact borders of any zones or corridors. The plan proposal includes 89 green corridors, with brief descriptions about areas they are intended to connect. According to the descriptions, 42 are based on our Corridor-Zonation results. The strategic plan mainly identifies corridors in the proximity of major cities and towns (46) and corridors that cross major highways via wildlife crossings (14). However, connectivity bottlenecks between large ecological networks, i.e. key connections identified by the present work, have mostly been missed by zoning: only one of the identified key connections in rural Uusimaa was marked in the Uusimaa 2050 plan proposal. Maintenance of additional green corridors would benefit regional connectivity in several parts of Uusimaa (Fig. 7). The Uusimaa 2050 plan proposal allows 66 out of 599 core connectivity areas (large top-20% sites) to disappear or decrease. Development zones threaten 85 (12.0%) of ecological connections. Furthermore, new highways and railways may (further) cut 146 (20.8%) of connections. The green belt around the capital district emphasizes both recreation and the connectivity needs of biodiversity. However, given that these green corridors are long and narrow connections through comparatively large ecologically high-priority sites that will be negatively impacted by a large population increase in the capital district and construction of new highways and railways (Fig. 6), the sufficiency of the proposed connections can easily be questioned.

Comparison of Corridor-Zonation results and the green connections in the Uusimaa 2050 plan proposal (UM2050 in the map legend). The green connections are concentrated around the capital district. Red circles mark areas where new corridor zones would significantly benefit regional connectivity by connecting many top-priority sites or large networks (Fig. 4a)
Discussion
This work shows how to use spatial prioritization for the identification of large ecological networks. The methods proposed were applied in a real-world case study, which is about the need to account for ecological networks in zoning in the province of Uusimaa, S-Finland, including the capital district. We identified seven large semi-contiguous mosaics of ecologically good-quality areas, which stand out from the rest of the landscape and harbor significant proportions of the biodiversity found in the Uusimaa region. We also identified connectivity bottlenecks, where ecological connectivity should be preserved or enhanced. We were also able to evaluate the impacts of a proposed Uusimaa 2050 regional plan to regional connectivity. The methods described here are of utility to land-use planners who are concerned about the maintenance of ecological values and the connectivity of the landscape.
It is rather straightforward to use spatial prioritization to identify the most important areas of a landscape for conservation or the least important areas for impact avoidance (Kareksela et al. 2013). However, in the context of ecological networks and land-use planning, one would be interested in delineating the parts of the landscape where habitat quality remains sufficient for biodiversity in general. As opposed to significantly human-degraded environments, these areas support reproduction and dispersal of a large fraction of the regional flora and fauna (Opdam et al. 2006; Hodgson et al. 2009, 2011; Reider et al. 2018). Connected networks can be difficult to identify from the priority maps alone. The Zonation rank map, for example, is always a map with values linearly scaled 0 to 1 and further information about the concentration of biodiversity is needed before areas that maintain biodiversity can be identified (Lehtomäki and Moilanen 2013; “Two spatial Zonation outputs: complementarity-based rank and scoring-based weighted range-size rarity” section). We used a combination of the complementarity-driven ranking and the weighted range-size rarity map to address this need. In terms of Zonation technique, this can be considered as a way to combine the so-called performance curves and the rank map. In the case of Uusimaa, log-transformation of the wrscr layer was required for visual delineation of the networks, which might not be required in all cases. Zonation allows use of feature-specific connectivity transformations, and it can be coupled with e.g. graph-theoretic methods in the input data pre-processing phase (Albert et al. 2017) if required, but the method presented here does not rely on external connectivity modelling methods.
The seven large networks shown in Fig. 4a cover a large fraction of the known biodiversity of Uusimaa. It turns out that some biodiversity features are highly concentrated into specific networks, which has implications for conservation planning and management (Table 1). Some of the networks go around small towns or other human-degraded areas, which was communicated to municipal-level planning offices. We specially focused on areas where the general habitat quality remains relatively high, but where ecologically high-quality areas have been narrowed down to connectivity bottlenecks. Overall, our results inform local planners about where new development would harm areas that are not only locally high-quality but also important for the regional ecological network as a whole (Opdam et al. 2006). Important isolated biodiversity sites do exist outside these networks (Wintle et al. 2018), but isolated areas are less likely to be influenced by changes in regional connectivity. Furthermore, we were able to assess the potential connectivity impacts of a newly proposed high-level regional plan proposal (Regional Council of Uusimaa 2018). Direct comparison of our results and the plan proposal is difficult due to the strategic level of the plan. We were, however, able to identify areas where new development would most severely impact ecological networks (Table 2, Fig. 6) or diminish core connectivity areas and connections (Fig. 7). In some places, negative ecological effects of new developments could be mitigated by careful planning, but in others this is practically impossible, as is the case for new residential zones at the western fringes of the capital district (Fig. 6). Spatial prioritization tools can support planning for mitigation and compensation of the new development (Kareksela et al. 2013). Top-priority areas should, for example, be avoided in municipal zoning. Zonation can also be used to identify places suitable for the expansion of core protected areas (Lehtomäki and Moilanen 2013). Our analysis regarding restoration needs of the connections can also aid planning for mitigation of connectivity losses: if some high-quality connections are lost and alternative ones exist, the connections with next-lowest restoration need should be restored and enhanced (Supplementary Fig. S7). We expect similar analyses to be broadly useful also in any zoning exercise that wishes to account for ecological connectivity effects.
Our interpretation of ecological corridors as short connectivity bottlenecks between larger areas of higher habitat quality (networks) somewhat challenges the traditional Finnish land-use planning tradition of describing corridors as links between specific core areas, e.g. reserves. In reality, long and narrow connections that encompass areas of varying habitat quality in human-modified landscapes might often not be the most appropriate way of describing how some areas relate to regional connectivity (Puth and Wilson 2001; Gippoliti and Battisti 2017). Many ecological studies have found doubtful benefits for long and narrow corridors (Mutanen and Mönkkönen 2003; Gilbert-Norton et al. 2010; Pérez-Hernández et al. 2014). Furthermore, long corridors can be seen as an unsafe approach to maintenance of biodiversity in Finland, because corridors are vague objects in legal terms and do not limit e.g. forestry activities (Salomaa et al. 2017). For example, in the Green belt of Helsinki, where human population growth puts very high pressure on biodiversity, a zone-type marking would better describe connectivity requirements than (very) long linear corridors (Fig. 7). We also identified target areas for habitat restoration in corridors (Fig. 5), although we do suspect that it is quite unrealistic to expect large-scale habitat restoration action to take place in Uusimaa. It is also worth noticing, that because restoration comes with operational uncertainties and time lags, preservation of high-quality area should in general be given priority over restoration (Maron et al. 2012; Spake et al. 2015). Nevertheless, our methods can help planners and managers to prioritize limited investment into habitat restoration.
The delineation of the networks from the balanced feature density map was based on two thresholds chosen from the Zonation performance curves (Fig. 2)—details of networks would vary if different thresholds were used. Here, the thresholds were chosen in an ecologically informed manner based on concentration of biodiversity, as shown by the mean performance curve (Fig. 1). The manual delineation of the large networks and connections (corridors, homogeneous areas of high connectivity) is another source of subjectivity in our approach. However, instead of mechanistically generalizing networks and connections, we wanted to include our local expert knowledge in e.g. adjusting the borders of the networks near major cities or connections in intensively managed areas. As spatial planning inevitably requires some subjective decisions and verification (Margules and Pressey 2000; Opdam et al. 2006; Lehtomäki and Moilanen 2013), we find the present approach adequate for the needs of regional-level planning. Technically, if such considerations are relevant and data exists, Zonation prioritizations like ours could easily be expanded to cover costs (e.g. land price, opportunity costs for forestry, etc.), current land-use or natural resource extraction plans, or other similar factors relevant for local zoning. Furthermore, the present work complements a recent prioritization of Finnish marine areas (Virtanen et al. 2018), allowing identification of sections of coastline that are important for both terrestrial and marine ecosystems.
The current discussion about connectivity, summarized by e.g. Miller-Rushing et al. (2019), has questioned the view that fragmentation, i.e. lack of structural connectivity, directly decreases landscapes’ support for species and populations which may have implications to e.g. land-use planning. In the long term, support of the landscape for populations is determined by the habitat quality of the matrix (Reider et al. 2018), which itself should be considered as a continuum. We therefore agree with Boitani et al. (2007) that strict focus on core areas and links in-between them may be problematic. As Boitani et al. (2007) state, it may lead to neglect of the habitat matrix at large, which too provides breeding habitats for species and supports connectivity. Human activities can obviously induce sharp changes (deterioration) in habitat quality. To minimize future degradation of the landscape, connectivity conservation should focus on impact avoidance (Kareksela et al. 2013). New developments should be concentrated into those parts of the landscape where overall habitat quality is already so low that only limited biodiversity is supported (highly-urbanized areas, areas of intense agriculture or forestry, etc.). In these areas, further human development leads to comparatively small ecological impacts—e.g., the lowest ranked 40% of the landscape in the present analysis. We emphasize that the present approach should be used in addition to, not instead of, “typical” prioritizations, which aim at identifying top-priority areas for inclusion in a protected area network. The present work shows that spatial prioritization can be used to achieve three types of analyses at once that comprehensively cover different aspects of biodiversity preservation in general land-use planning. They include: (i) analyses for core areas of conservation, (ii) analyses for impact avoidance, and (iii) analyses for regional connectivity. Together, these analyses should lead to ecologically well informed land-use planning with benefits for sustainable development (United Nations 2015).
Conclusions
How well a landscape maintains biodiversity depends on remaining habitat quality and past, present, and future human impacts. Minimization of unnecessary ecological impacts is key to sustainable landscape and land-use planning. Identifying core areas and corridors for individual target or indicator species is not enough to halt biodiversity loss. A much safer approach is to concentrate further land-use development into areas that do little harm to complementarity-driven top-priority sites or to continuous or semi-continuous areas of generally high biodiversity. The methods described here provide land-use planners more tools for steering future developments in a nature-sensitive way. Assuming that data about the distributions of biodiversity features are available, our methods, which are based on spatial conservation prioritization techniques, can be replicated anywhere.
References
Albert CH, Rayfield B, Dumitry M, Gonzalez A (2017) Applying network theory to prioritize multispecies habitat networks that are robust to climate and land-use change. Conserv Biol 31:1383–1396.
Álvarez-Romero JG, Munguía-Vega A, Beger M, del Mar Mancha-Cisneros M, Suárez-Castillo AN, Gurney GG, Pressey RL, Gerber LR, Morzaria-Luna HN, Reyes-Bonilla H, Adams VM, Kolb M, Graham EM, VanDerWal J, Castillo-López A, Hinojosa-Arango G, Petatán-Ramírez D, Moreno-Baez M, Godínez-Reyes CR, Torre J (2018) Designing connected marine reserves in the face of global warming. Glob Change Biol 24:e671–e691
Boitani L, Falcucci A, Maiorano L, Rondinini C (2007) Ecological networks as conceptual frameworks or operational tools in conservation. Conserv Biol 21:1414–1422.
CBD (2010) Decision UNEP/CBD/COP/DEC/X/2 adopted by the conference of the parties to the convention on biological diversity at its tenth meeting. https://www.cbd.int/decision/cop/?id=12268. Accessed 29 Apr 2019
Chetkiewicz C-LB, St. Clair CC, Boyce MS (2006) Corridors for conservation: integrating pattern and process. Annu Rev Ecol Evol Syst 37:317–342.
Commission E (2011) Our life insurance, our natural capital: an EU biodiversity strategy to 2020. European Commission, Brussels
Correa Ayram CA, Mendoza ME, Etter A, Salicrup DRP (2016) Habitat connectivity in biodiversity conservation: a review of recent studies and applications. Prog Phys Geogr 40:7–37.
Fahrig L (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. J Biogeogr 40:1649–1663.
Finnish Biodiversity Action Plan (2012) Government resolution on the strategy for the conservation and sustainable use of biodiversity in Finland for the years 2012–2020. The Government of Finland, Helsinki, p 26
Fischer J, National TA (2006) Beyond fragmentation: the continuum model for fauna research and conservation in human-modified landscapes. Oikos 2:473–480
Foltête JC (2019) How ecological networks could benefit from landscape graphs: a response to the paper by Spartaco Gippoliti and Corrado Battisti. Land Use Policy 80:391–394.
Gilbert-Norton L, Wilson R, Stevens JR, Beard KH (2010) A meta-analytic review of corridor effectiveness. Conserv Biol 24:660–668.
Gippoliti S, Battisti C (2017) More cool than tool: Equivoques, conceptual traps and weaknesses of ecological networks in environmental planning and conservation. Land Use Policy 68:686–691.
Hodgson JA, Moilanen A, Wintle BA, Thomas CD (2011) Habitat area, quality and connectivity: striking the balance for efficient conservation. J Appl Ecol 48:148–152.
Hodgson JA, Thomas CD, Wintle BA, Moilanen A (2009) Climate change, connectivity and conservation decision making: back to basics. J Appl Ecol 46:964–969.
Jalkanen J, Moilanen A, Toivonen T (2018a) Uudenmaan ekologiset verkostot Zonation-analyysien perusteella. Uudenmaan liiton julkaisuja E 194, Helsinki, p 132
Jalkanen J, Moilanen A, Toivonen T (2018b) Uusimaa-kaavan 2050 luontovaikutusten arviointi Zonation-analyyseihin perustuen. Uudenmaan liiton julkaisuja E 205, Helsinki, p 44
Kareksela S, Moilanen A, Tuominen S, Kotiaho JS (2013) Use of inverse spatial conservation prioritization to avoid biological diversity loss outside protected areas. Conserv Biol 27:1294–1303.
Kujala H, Lahoz-Monfort JJ, Elith J, Moilanen A (2018) Not all data are equal: influence of data type and amount in spatial conservation prioritisation. Methods Ecol Evol 9:2249–2261.
Kullberg P, Moilanen A (2014) How do recent spatial biodiversity analyses support the convention on biological diversity in the expansion of the global conservation area network? Nat Conserv 12:3–10.
Kuusterä J, Aalto S, Moilanen A, Toivonen T, Lehtomäki J (2015) Uudenmaan viherrakenteen analysointi Zonation-menetelmällä. Uudenmaan liiton julkaisuja E 145—2015, Helsinki, p 77
Lehtomäki J, Moilanen A (2013) Methods and workflow for spatial conservation prioritization using Zonation. Environ Model Softw 47:128–137.
Margules DR, Pressey RL (2000) Systematic conservation planning. Nature 405:243–253
Maron M, Hobbs RJ, Moilanen A, Matthews JW, Christie K, Gardner TA, Keith DA, Lindenmayer DB, McAlpine CA (2012) Faustian bargains? Restoration realities in the context of biodiversity offset policies. Biol Conserv 155:141–148
Martin CA (2018) An early synthesis of the habitat amount hypothesis. Landsc Ecol 33:1831–1835.
Meurant M, Gonzalez A, Doxa A, Albert CH (2018) Selecting surrogate species for connectivity conservation. Biol Conserv 227:326–334.
Mikkonen N, Moilanen A (2013) Identification of top priority areas and management landscapes from a national Natura 2000 network. Environ Sci Policy 27:11–20.
Miller-Rushing AJ, Primack RB, Devictor V, Corlett RT, Cumming GS, Loyola R, Maas B, Pejchar L (2019) How does habitat fragmentation affect biodiversity? A controversial question at the core of conservation biology. Biol Conserv. https://doi.org/10.1016/J.BIOCON.2018.12.029
Moilanen A (2011) On the limitations of graph-theoretic connectivity in spatial ecology and conservation. J Appl Ecol 48:1543–1547.
Moilanen A, Franco AMA, Early RI, Fox R, Wintle B, Thomas CD (2005) Prioritizing multiple-use landscapes for conservation: methods for large multi-species planning problems. Proc R Soc B Biol Sci 272:1885–1891
Moilanen A, Possingam H, Polasky S (2009) A Mathematical classification of conservation prioritization problem. In: Moilanen A, Wilson K, Possingham H (eds) Spatial conservation prioritization: quantitative methods and computational tools. Oxford University Press, Oxford, pp 28–42
Moilanen A, Pouzols FM, Meller L, Veach V, Arponen A, Leppänen J, Kujala H (2014) Zonation Version 4 user manual. C-BIG, University of Helsinki, Helsinki, p 288
Mutanen M, Mönkkönen M (2003) Occurrence of moths in boreal forest corridors. Conserv Biol 17:468–475
Newbold T, Hudson LN, Arnell AP, Contu S, De Palma A, Ferrier S, Hill SLL, Hoskins AJ, Lysenko I, Phillips HRP, Burton VJ, Chng CWT, Emerson S, Gao D, Pask-Hale G, Hutton J, Jung M, Sanchez-Ortiz K, Simmons BI, Whitmee S, Zhang H, Scharlemann JPW, Purvis A (2015) Global effects of land use on local terrestrial biodiversity. Nature 520:45–50
Opdam P, Steingröver E, Van RS (2006) Ecological networks: a spatial concept for multi-actor planning of sustainable landscapes. Landsc Urban Plan 75:322–332.
Pérez-Hernández CG, Vergara PM, Saura S, Hernández J (2014) Do corridors promote connectivity for bird-dispersed trees? The case of Persea lingue in Chilean fragmented landscapes. Landsc Ecol 30:77–90.
Pouzols FM, Moilanen A (2014) A method for building corridors in spatial conservation prioritization. Landsc Ecol 29:789–801.
Prugh LR, Hodges KE, Sinclair ARE, Brashares JS (2008) Effect of habitat area and isolation on fragmented animal populations. Proc Natl Acad Sci 155:20770–20775.
Puth LM, Wilson KA (2001) Boundaries and corridors as a continuum of ecological flow control: lessons from rivers and streams. Conserv Biol 15:21–30.
Rayfield B, Fortin MJ, Fall A (2011) Connectivity for conservation: a framework to classify network measures. Ecology 92:847–858.
Regional Council of Uusimaa (2017) Uudenmaan neljäs vaihemaakuntakaava: Selostus. Uudenmaan liitto, Helsinki, p 174
Regional Council of Uusimaa (2018) Uusimaa-kaava 2050: Helsingin seudun. Länsi-Uudenmaan ja Itä-Uudenmaan vaihemaakuntakaavojen ehdotukset, Helsinki, p 236
Reider IJ, Donnelly MA, Watling JI (2018) The influence of matrix quality on species richness in remnant forest. Landsc Ecol 33:1147–1157.
Rouget M, Cowling RM, Lombard AT, Knight AT, Kerley GIH (2006) Designing large-scale conservation corridors for pattern and process. Conserv Biol 20:549–561
Salomaa A, Paloniemi R, Kotiaho JS, Kettunen M, Apostolopoulou E, Cent J (2017) Can green infrastructure help to conserve biodiversity? Environ Plan C Gov Policy 35:265–288
Spake R, Ezard THG, Martin PA, Newton AC, Doncaster CP (2015) A meta-analysis of functional group responses to forest recovery outside of the tropics. Conserv Biol 29:1695–1703
United Nations (2015) Transforming our world: the 2030 agenda for sustainable development. https://sustainabledevelopment.un.org/post2015/transformingourworld. Accessed 29 Apr 2019
Veach V, Di Minin E, Pouzols FM, Moilanen A (2017) Species richness as criterion for global conservation area placement leads to large losses in coverage of biodiversity. Divers Distrib 23:715–726.
Virtanen EA, Viitasalo M, Lappalainen J, Moilanen A (2018) Evaluation, gap analysis, and potential expansion of the finnish marine protected area network. Front Mar Sci 5:1–19.
Volk XK, Gattringer JP, Otte A, Harvolk-Schöning S (2018) Connectivity analysis as a tool for assessing restoration success. Landsc Ecol 33:371–387.
Williams PH (2000) Some properties of rarity scores used in site quality assessment. Br J Entomol Nat Hist 13:73–86
Wintle BA, Kujala H, Whitehead A, Cameron A, Veloz S, Kukkala A, Moilanen A, Gordon A, Lentini PE, Cadenhead NCR, Bekessy SA (2018) Global synthesis of conservation studies reveals the importance of small habitat patches for biodiversity. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.1813051115
Acknowledgements
Open access funding provided by University of Helsinki including Helsinki University Central Hospital. J.J. was supported by the Kone Foundation. A.M. was supported by the Strategic Research Council project IBC-Carbon, grant #312559. T.T. was supported by the University of Helsinki, Kone foundation, and the Helsinki Metropolitan Region Urban Research Program. We thank Regional Council of Uusimaa for their interest and support in the use of ecologically informed spatial prioritization in land use planning and zoning.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jalkanen, J., Toivonen, T. & Moilanen, A. Identification of ecological networks for land-use planning with spatial conservation prioritization. Landscape Ecol 35, 353–371 (2020). https://doi.org/10.1007/s10980-019-00950-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-019-00950-4