
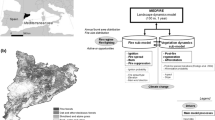
Abstract
Context
Planted pine forests are highly abundant communities in the Mediterranean Basin. Being the result of past reforestation, these forests show high species and structural homogeneity. Diversification to conifer-broadleaved mixed forests is recommended to promote adaptation to climate change and increase their resilience to perturbations.
Objectives
This study aims at evaluating how these planted pine forests will develop in the future as a result of the combined impacts of management and climate.
Methods
We applied a forest landscape model (LANDIS-II) to simulate different climate scenarios and management strategies designed in cooperation with forest managers to assess their effects on the performance of planted pine forests and their change in terms of forest composition.
Results
Climate change scenarios caused a shift in the phenological growth pattern of planted pine forests by reducing forest growth during summer and increasing photosynthetic productivity in spring and fall, particularly under high emission scenarios. Biomass increased through time and more strongly under climate change, but this increase differs among species, resulting in changes of forest types across the landscape. Our results portray natural succession as the main driver of forest change, but intensive management accelerated this process by limiting pine growth and promoting growth of oak species.
Conclusions
Our results highlight the importance of active management on planted pine forests to favour mixed and climate-adapted ecosystems in shorter time scales than offered by succession alone. Moreover, our spatially explicit modelling approach helps to identify areas where lack of seed dispersal and/or competitive exclusion prevent natural diversification, providing useful recommendations for interventions. However, the modelling approach has some limitations since it does not consider natural disturbances.
Graphical abstract

Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Monospecific and even-age forests constitute the most abundant forest types in Europe as a result of intensive anthropogenic management over the last centuries (EEA 2016). These ecosystems were created or promoted for purposes such as timber production, soil erosion protection, hydrological management and as facilitators for the subsequent establishment of more shade-tolerant broad-leaved species (Coll et al. 2021; Waitz and Sheffer 2021). In Mediterranean areas, such monospecific forest communities are often composed of pine species (Pinus spp.) which have received limited post-planting management (Coll et al. 2021). As a result, these planted pine forests frequently present high densities, low structural heterogeneity and low tree species diversity, which make them vulnerable to perturbations such as forest fires and pest outbreaks (Senf and Seidl 2022). These factors also limit their natural diversification over time (Gómez-Aparicio et al. 2009; Waitz and Sheffer 2021). Moreover, climate change is expected to cause forest dieback in these planted pine forests as a result of increased competition under water shortage in highly dense stands (Navarro-Cerrillo et al. 2019; Peñuelas and Sardans 2021). Actually, drought-induced mortality events have already been reported for different species in western Mediterranean pine forests (Navarro Cerrillo et al. 2007; Navarro-Cerrillo et al. 2018; Waitz and Sheffer 2021).
Current management goals aim to increase the adaptive capacity of Mediterranean pine forests to climate change and its expected risks (Navarro-Cerrillo et al. 2019). In this regard, mixed forests (i.e., conifer-broadleaved mixtures) show better capacities to face drought events and climate change in dry areas (Pardos et al. 2021) due to the presence of multiple species with different functional traits in terms of water use, post-fire behaviour or sensitivity to pests. Thus, Mediterranean mixed forests show an improved capacity to deal with climate change impacts (Coll et al. 2021; Waitz and Sheffer 2021). Therefore, diversification of pure conifer stands to mixed conifer-broadleaved forests is recommended as a suitable strategy to promote adaptation to climate change and increase resilience to disturbances such as fires (de-Dios-García et al. 2015; Coll et al. 2021; Oettel and Lapin 2021; Messier et al. 2022).
Under these circumstances, there is an urgent need to better understand how Mediterranean planted pine forests might develop under future and uncertain climate regimes. Moreover, assessing the long-term effects of different management scenarios on Mediterranean planted pine forests can provide hints to define which are the best management strategies to promote diversification. Focusing on a 390,000 ha forested landscape in southeastern Spain, we herewith evaluate how Mediterranean planted pine forests might change through time as a result of the combined impacts of management and climate scenarios. These forests constitute a good example to study the effects of climate and management since they cover a large surface (Rubio-Cuadrado et al. 2018), they are located in highly sensitive areas to climate change (Giorgi 2006) and they cover a wide range in terms of dominant species and climate regimes, from xeric to alpine conditions. We assessed different trajectories of these forests using a forest landscape model that incorporates climate and management effects on forest dynamics. Management scenarios were designed in collaboration with foresters to reproduce realistic management strategies. With this work we aim to answer the following questions:
-
How will future climate and management affect the performance (i.e., photosynthesis and biomass) of planted pine forests?
-
How will different climate and management scenarios affect the forest composition change of these pine dominated systems?
-
To what extent different management strategies can help to speed up the transition towards mixed forests?
Methodology
Study area
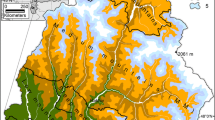
The study area is located in southeastern Spain (Fig. 1). It includes Sierra Nevada, Sierra de Arana and Sierra de Baza-Filabres mountain ranges, resulting in a complex landscape ranging from 400 to 3479 m a.s.l. Planted pine forests considered in this study represent 17% of the surface and are located at intermediate elevations (1578 ± 358 m a.s.l.). Dominant pine species vary with elevation, from Pinus halepensis Miller. (Aleppo pine) and Pinus pinaster Aiton (Maritime pine) at the lowest elevations, to Pinus nigra Arnold. (Black pine) and Pinus sylvestris L. (Scots pine) at higher elevations (Mesa Garrido 2019). Most of the existing pine forests stands were planted during the first half of twentieth century to avoid soil erosion. These plantations are currently covering the natural niche of several Quercus species (Quercus ilex L.; Holm oak, Q. pyrenaica Willd.; Pyrenean oak, and Q. faginea Lam.; Portuguese oak). The study area still has some remaining patches of this natural vegetation. Apart from pines and oaks, large areas are covered by sclerophyll shrublands in lowlands and sparse alpine vegetation at the high altitudes. Climate is highly seasonal, with dry hot summers and mild winters typical of the Mediterranean climate. The planted pine forests considered in this study are mainly located at intermediate elevations, what we define as subhumid and mesic climate areas, with some patches also in the low-elevation xeric climate area (Fig. 1).

Schematic view of the study area: A location; B planted pine forests; C details of study area, with colours referring to climate areas and planted pine forests overlapped and highlighted in dark green. Figure SI1 shows the current distribution of all forest types
Modelling approach
LANDIS-II is a widely-applied forest landscape model created to simulate forest growth and disturbance across large landscapes (Scheller et al. 2007). The spatially-explicit structure allows simulating forest succession and seed dispersal among interconnected grid-cells at decadal to multi-century time scales, as well as the potential influence of perturbations such as management interventions, among others. Trees are grouped into species- and age-specific cohorts in each cell. LANDIS-II is built on a core module interacting with multiple model extensions handling ecological processes at various levels of complexity.
Model initialization
The methodology and data sources for generating 1-ha resolution initial vegetation conditions and biophysical inputs are extensively described in Suárez-Muñoz et al. (2021). Within initial vegetation conditions—i.e., current species presence and age representing the starting point for the simulations—shrub species groups were also included as they are an abundant land cover in the study area as well as important elements affecting forest dynamics in drought-prone Mediterranean areas (Cassell et al. 2019). Biophysical inputs (i.e. ecoregions) were obtained by combining homogeneous climate areas (see Fig. 1), derived from historical climate maps, with maps of soil texture.
Forest growth and succession
To simulate future succession, we used the LANDIS-II PnET-Succession extension v4.1 (De Bruijn et al. 2014). PnET-Succession simulates cohorts’ competition for light and water, so that photosynthetic processes are explicitly linked to changing environmental drivers such as temperature, precipitation and CO2 (De Bruijn et al. 2014). A mechanistic process-based approach is used to simulate forest dynamics, which is being considered better suited for long-term projections under unprecedented environmental conditions (Gustafson et al. 2015). PnET-Succession simulates species-cohort growth integrating competition with other cohorts and the effect of changing climate. Gross photosynthesis is calculated as a function of foliar nitrogen multiplied by a series of reduction factors, which control how water availability, temperature, age or light limit potential photosynthesis. Species-specific parameters and monthly climate series are considered in the calculation of these reduction factors. Cohorts mortality is a result of either gross photosynthetic rate reduction with age or depletion of carbon reserves below a certain threshold. We calibrated PnET-Succession parameters by running single-cell simulations with different species combinations and compared outcomes with empirical biomass values derived from silvicultural schemes proposed by Campos and Díaz-Balteiro (2015). Further details on calibration of species parameters are given in Suárez-Muñoz et al. (2021) and Appendix SI1.
Climate change
We considered three climate scenarios: contemporary climate (current), moderate emissions (RCP4.5) and high emission (RCP8.5). Climatic series for LANDIS-II simulations were obtained from the MIROC5 global circulation model and precipitation data were further bias-corrected based on meteorological stations data in the study area. Monthly precipitation and temperature data were obtained from CHELSA (Karger et al. 2020). This dataset contains historical series (1950–2005) as well as future projections (2006–2100) downscaled to ~ 5 km spatial resolution. Photosynthetically active radiation (PAR) data were obtained from Cornes et al. (2018) version 20.0e and CO2 concentrations were obtained from Mina et al. (2021). Future projections showed a clear increasing gradient in temperatures and CO2 from current to RCP8.5 scenario, while trends for precipitation were less divergent between emission scenarios, despite showing a slight decreasing gradient under the highest emissions scenario (Fig. 2 and SI18). More details on the data sources, the generation of climate scenarios and the precipitation bias correction are provided in Appendix SI2.

Annual average temperature (A) and precipitation (B) under current, RCP4.5 and RCP8.5 climate scenarios. Data refer to subhumid climate area, where the highest proportion of planted pine forests is found (see map of climate areas in Fig. 1). Trend line and shaded area indicate smoothed conditional mean and confidence intervals based on loess method. Monthly average temperatures are shown in Figures SI17–SI19
Forest management
Management scenarios were designed in close collaboration with local managers and forest experts to ensure realism and applicability of model outputs. Authors and forest managers met in iterative meetings to describe the current management of planted pine forests in the study area, as well as to discuss the limitations in place to apply alternative management strategies. Among limitations, managers highlighted the limited budget available for interventions as well as concerns regarding harvesting impacts which call for precaution when designing management strategies. Considering these ideas, four management scenarios were applied in this study, representing an intensity gradient in terms of proportion of area harvested and biomass removed (Table 1). The ‘non-management’ scenario represents a common situation in large portions of the territory, as a result of limited budget allocated to forest management. The ‘conservative’ scenario is highly driven by the precautionary principle: managers prevent post harvesting mortality due to increased wind shear and snowfalls impacts by applying low thinning intensities in planted pine forests and lightly promoting Quercus spp. growth through coppice. The ‘proactive’ scenario assumes that precautionary management removes an insufficient amount of biomass to trigger the desired conditions in species composition. Therefore, under this scenario, higher amounts of biomass are removed within stands while the proportion of area harvested is comparable with the conservative scenario. Finally, the proactive-plus scenario represents a situation where the amount of biomass removed within stands is the same as in the proactive scenario, but more resources are allocated to forest management and therefore broader areas of the landscape are harvested.
Forest management was implemented with the Biomass Harvest extension v4.4 (Gustafson et al. 2000), which reproduces silvicultural activities by harvesting biomass from the landscape based on user-defined rules. Prescriptions define which cells, species and age ranges are targeted and which proportion of biomass is removed from each cohort. The study area was divided into forest stands (Figure SI24) grouped into management units (Figure SI25), each with its own set of silvicultural prescriptions and harvest treatment frequency. Management units were based on land property (public/private) and conservation use (reserve/non-reserve). Non-reserve public areas, where most of the planted pine forests are located (81% of the surface), were managed more intensively than private ones, and reserve areas were not managed under any scenario, reflecting what is commonly occurring in the territory (Junta de Andalucía 2004, 2011). Further details on the implementation of management scenarios and silvicultural prescriptions are provided in the Appendix SI3.
Experimental design and analysis
A combination of climate and management scenarios was simulated in this study for the period 2005–2100, for a total of 60 simulations (3 climate scenarios × 4 management scenarios × 5 replicates). Model replicates are run to account for the variability due to stochastic processes within LANDIS-II, such as establishment probability and seed dispersal (Hong and Mladenoff 1999). Model outputs were analysed by extracting variables for the area covered by the planted pine forests according to the Andalusian Map of Vegetation Land Cover (REDIAM 2006) (Fig. 1). Thus, we refer to the results at landscape level as the aggregated values of all cells within planted pine forests and to cell level when data for each cell were analysed independently, allowing to evaluate the existence and changes in spatial patterns.
Model outputs were assessed by analysing changes in average monthly net photosynthesis (hereafter photosynthesis), aboveground biomass (hereafter biomass) and mortality of cohorts. We compared photosynthesis at the beginning and at the end of the simulation period. Biomass was analysed both at cell- and landscape-level. The temporal trend in cell-level biomass was analysed by applying the non-parametric Mann–Kendall trend test (Mann 1945). This test evaluates the significant trend of a variable over time based on the Kendall rank correlation, which measures the strength of a monotonic association between two variables. Kendall’s tau statistic ranges from −1 and 1, and indicates a negative and a positive, respectively, trend in the analysed variable through time. Landscape-level biomass was evaluated by analysing total and species-specific biomass densities through time. Mortality of cohorts per cell was calculated as the difference between the number of cohorts of a species at a certain time step and the number of surviving cohorts of the species in the subsequent time step. The values were obtained by combining PnET-Succession and Biomass by Age extension outputs.
We also evaluated changes in overall forest composition by computing the difference in forest types coverage between the initial and final years of the simulation. We reclassified cell species biomass into 7 forest type categories: pure pines (pines biomass > 90% of the cell biomass), pure oaks (oaks biomass > 90%), mixed no dominance (no specific group accounts for more than 90% of the biomass), mixed pine-dominated (pines 50–90%), mixed oak-dominated (oaks 50–90%), shrublands (shrubs > 50% of total biomass) and empty (no biomass). All analyses were performed using the software R v4.1.1 (R Team 2022).
Results
Net photosynthesis changes
Our results indicate a seasonal growth pattern in concordance with tree species seasonal growing in the Mediterranean climate (e.g. two active growing periods and summer growth stagnation, Fig. 3). Monthly climate clearly defined such intra-annual growth pattern while changes caused by management interventions did not alter the photosynthetic pattern of planted pine forests at landscape level (Figure SI27). Under current climate, the intra-annual growth pattern showed maximum photosynthesis in May (86.78 ± 22.19 g m−2 at the beginning of the simulation). Changes due to forest succession only—changes due to vegetation development without management or climate change influences, i.e. those observed between the beginning and the end of the simulation under current climate and non-management scenario—caused a slight decrease in photosynthesis during early summer and a slight increase in autumn, resulting in a small net decrease between the beginning and the end of the simulation (−6.8%).

Average monthly net photosynthesis of the area covered by planted pine forests (elevation 1578 ± 358 m a.s.l.) across climate scenarios. Since differences between management scenarios were minimal, results are shown for non-management. See Figure SI27 for all scenarios. Beginning and End periods correspond to the average among the first and the last three simulation timesteps, respectively (years 2005, 2010 and 2015 and years 2090, 2095 and 2100). Ribbon represents the standard deviation among timesteps from the mean values of five replicates
Compared to current climate, climate change increased intra-annual photosynthesis both at the beginning and at end of the simulation. Under RCP4.5, spring growth was higher at the beginning of the simulation (114.72 ± 20.57 g m−2 in May), while at the end the summer stagnation started earlier and was more pronounced, but the autumn growing period lasted longer than under current climate. Overall, annual photosynthetic productivity under RCP4.5 decreased through time (−29.9% between the beginning and the end of the simulation). Under RCP8.5, the spring growing period reached similar values than under current climate at the beginning of the simulation (95.34 ± 57.69 g m−2). Climate change stressed summer stagnation: photosynthesis in August was 11.39 ± 10.57, 7.74 ± 0.99 and 2.17 ± 1.02 g m−2 at the end of the simulation under current, RCP4.5 and RCP8.5 climate scenarios, respectively. At the end of the simulation, the RCP8.5 scenario resulted in much earlier spring growth and much later autumn growth. Specifically, spring growing phase started in February, reaching its maximum in April (108.42 ± 32.77 g m−2), while growth continued during late autumn until December. This resulted in an overall increment of annual photosynthesis between the beginning and the end of the simulation under RCP8.5 (+31.9%).
Biomass and mortality trends
We found a significantly increasing trend in total biomass in large areas of the landscape under all simulated scenarios, especially under RCP8.5 climate, as shown in Table 2. This table shows the proportion of area where a significant positive, negative or absent trend in total aboveground biomass was found. Under current climate, mesic and subhumid areas showed increasing trends in total biomass while xeric areas showed decreasing trends (Figure SI30). Management modulated the increase in biomass by limiting its increase under more intense management scenarios. As such, the increasing trend signal was stronger under non-management than under the more intense management. For example, 63% of the area shows a significantly increasing trend and 12% a no significant trend under the conservative RCP4.5 scenario, while under the proactive-plus RCP4.5 scenario 47% of the area shows a significantly increasing trend and 18% of the area shows no significant trend.
At landscape level, total aboveground biomass increased during the first decades under current climate and then decreased, while under RCP4.5 it stabilised around year 2050 and under RCP8.5 it increased through time (Figure SI28). For individual species, we observed differences across simulated scenarios (Fig. 4). Pines remained the most abundant species group at landscape level throughout the entire simulation under all climate and management scenarios. Biomass of Pinus nigra increased under all climate scenarios but its biomass accrue was limited by intensified management interventions. On the other hand, biomass of P. halepensis decreased towards the end of the simulation and this decrease was more pronounced under RCP8.5. Biomass of P. sylvestris and P. pinaster, however, remained quite stable through the simulation under all scenarios, although P. pinaster was limited by intensive management and seemed to be benefited under RCP8.5 climate. Among oaks, Quercus ilex—and less clearly Q. pyrenaica—increased in biomass during the final 50 years of the simulation under all climate and management scenarios, but more strongly under more intense management and more rapidly under RCP8.5.

Aboveground biomass of pines and oaks through time at landscape level (the region initially covered by planted pine forests) for all management scenarios under Current and RCP8.5 climate scenarios. Values correspond to mean across 5 replicates and error bars, when visible, represent standard deviation. See Figure SI28 for all climate scenarios
Biomass accumulation show different spatial patterns across the landscape (Figures SI32–SI35). Pine biomass decreased in xeric areas under all management and climate scenarios, but more clearly under more intense management and current and RCP4.5 climate scenarios. On the other hand, oak biomass declined at mesic and subhumid areas under current and RCP4.5 climate scenarios and more so under non-management and conservative management scenarios.
Oaks and pines showed different mortality patterns among scenarios through time (Fig. 5). Climate had a stronger effect on mortality, while management slightly influenced the mortality pattern of oaks and pines (Figure SI36). Pines died more than oaks, and their mortality was higher under climate change, contrary to oaks, which had a lower mortality under the more intense climate change scenarios.

Number of dead cohorts of pines and oaks through time under different climate and non-management scenarios. Data refer to the area initially covered by planted pine and values correspond to one model replicate. Figure SI36 shows all combinations of climate and management scenarios
Forest composition change under climate and management scenarios
The opposing trends in the biomass of oaks and pines and the differential mortality patterns resulted in changes of forest types for some areas. Planted pine forests, initially pure or mixed pine-dominated stands, partially developed into oak-dominated woodlands by the end of the century. Our results show a reduction in the area initially covered by pure pine and mixed pine-dominated stands, and an increase of pure oak and mixed oak-dominated stands (Fig. 6). This pattern was consistent among all management and climate scenarios but was promoted by more intense management. Under the proactive-plus scenario the highest increase in oak stands and the highest reduction in pure pine stands was achieved, followed by the proactive scenario. Conservative and non-management scenarios showed similar increase in oak stands, but the decrease in pure pine stands was more pronounced under the conservative scenario.

Change in percentage of the most abundant forest type area between initial and final year of the simulation (2005 and 2100, respectively). Error bars depict the standard deviation among the five replicates. See Table SI4 for the minor forest types
Climate interacted with management by amplifying the above-mentioned effects. The increase in oak stands and the decrease in pure pine stands under each management scenario were generally more pronounced under RCP8.5 scenario, followed by RCP4.5 compared to current climate. Only under current climate and non-management we observed a slight increase in pure pine stands, likely due to a shift from mixed pine-dominated to pure pine stands.
The change in forest types shows a characteristic spatial pattern (Figures SI36–SI37). Pine stands which were transformed into oak stands (either pure or mixed oak-dominated) were mainly located in xeric areas, while pine stands in mesic and subhumid areas remained so by the end of the century. As an example, there was a clear transformation from pine stands to oak stands in the northwestern area (Sierra de Arana in Fig. 1) while the large east-to-west patch of pine stands found in the centre of the study area (Sierra Nevada) mainly remained as pine stands.
Discussion
Climate controls biomass accruing in the landscape while management determines forest composition
Our analysis revealed that climate change had a large impact on the seasonal photosynthetic pattern of Mediterranean planted pine forests. Climate change generally increased annual growth, causing a higher net annual productivity by the end of the century under extreme climate change (+ 31.9% under RCP8.5). The differences between climate change scenarios were due to the fact that growth would no longer be limited during winter under extreme climate change (Fig. 2) (e.g., see Di Filippo et al. 2021 for P. halepensis forests), compensating additional growth losses due to more pronounced summer droughts. Our findings corroborate other studies showing enhanced growth due to climate change on Mediterranean forests. For instance, Madrigal-González et al. (2018) reported net positive effects of climate change on P. pinaster growth over the last decades in the central part of the Iberian Peninsula, while Gea-Izquierdo et al. (2011) found that Q. ilex stands on cold areas in the Iberian Peninsula may be benefited under climate change by analysing tree-rings over the last 100 years.
An increasing trend in biomass was observed not only under extreme climate change but also under current climate and moderate climate change. This was likely due to the natural ageing of these planted pine forests, initially composed of relatively young stands (initial average age 42 ± 7 years old). Our results showed that planted pine forests will likely accrue biomass through time in mesic and subhumid areas. Meanwhile, in xeric areas the increasing trend of biomass was only observed under extreme climate change. This increase in biomass is the result of higher projected temperatures and CO2 enrichment, which allowed the continuation of growth for most part of the winter. An overall increase in biomass under RCP8.5 scenario was also simulated by García-Duro et al. (2021) in the Southern Carpathian temperate forests. García-Duro et al. (2021) and other authors (de Wergifosse et al. 2022) have suggested that this simulated growth increment could be an effect of increased CO2 concentrations stimulating higher carbon uptakes and primary productivity, as well as the effect of milder winter temperatures lengthening the growing season (Ueyama et al. 2020). Nevertheless, some authors have expressed their concerns regarding the uncertainties involved when modelling growth under CO2 concentrations above calibration range (Gea-Izquierdo et al. 2017). Additionally, forest landscape models like ours rarely account for sink limitations due to higher CO2 concentration (e.g., lack of turgor for cell expansion).
Our study also showed that the effects of climate on total biomass were modulated by management, which had different influences on the dynamics of each tree group. The management scenarios represent a gradient of intervention intensity limiting the growth of pines and favouring oak expansion. This strategy resulted in different patterns depending on the climate area. In xeric areas, the decreasing pine biomass trend responded to a combination of climate and management impact, but also to the mortality of P. halepensis, the most common pine species in these areas (Figure SI39), due to its shorter longevity (150 years, therefore reaching longevity limit within simulations). In mesic and subhumid areas, overcompetition of pines over oaks and limited capacity of oaks to establish and develop under such conditions may have diluted the effects of management.
The underestimation of the incidence of extreme weather events (see Fischer et al. 2013) and specifically of droughts in the Mediterranean Basin (Tramblay et al. 2020) is a known issue in climate projections. In this area, weather extremes (e.g., drought events) are expected to become more frequent together with the reduction in annual precipitation (Zittis et al. 2021). Since precipitation is the main limiting factor of Mediterranean forest growth (Durante et al. 2009), an increment in drought-induced mortality is expected under climate change (Peñuelas and Sardans 2021). In our study area, some drought events have already been reported to cause mortality and dieback of conifer stands (Navarro Cerrillo et al. 2007; Sánchez-Salguero et al. 2013). These dieback events allow us to infer that the modelled biomass accrue would be less important if we would have been able to include the impact of droughts. Besides, we can hypothesize that this lower biomass accumulation would be more relevant in pines than in oaks. Thus, the lack of a clear decreasing trend in accumulated precipitation in climate projections used here as model inputs (see Figures SI14 and SI18) could explain the relatively low mortality observed in our results under extreme climate change (García-Duro et al. 2021). This aspect has been explored by Mina et al. (2022), who artificially modified climate scenarios by introducing a drought signal to simulate the incidence of extreme weather events in Quebec’s forests. Further studies in our area could use a similar approach by simulating extended drought periods and observing the effects on forest dynamics. In this work, nevertheless, the focus was on management and projected climate scenarios instead of extreme events and therefore such an approach fell out of the scope of the study. Apart from the influence of climate series, simulated mortality in LANDIS-II could as well be limited by the internal structure of the model, i.e., cohort senescence and mortality function. PnET-Succession considers drought mortality only due to carbon starvation and not hydraulic-related mortality—a process rarely integrated in simulation models (Bugmann et al. 2019). Therefore, underestimation of drought-induced mortality could be expected in these simulations. Despite this, our results still indicate a higher mortality rate of pines over oaks, which is expected given the ecological characteristics of each tree genus, especially regarding drought resistance (Waitz and Sheffer 2021). Further studies comparing multiple models using different approaches for drought-induced mortality could deliver crucial insights on such processes and the uncertainties around them. This kind of studies could also give hints for further model improvements, which would therefore provide managers with more robust recommendations to support decision-making (Boulanger et al. 2021).
Nevertheless, increased tree vulnerability due to climate change remains a complex topic (Allen et al. 2015). We acknowledge that our results apply in the lack of potential disturbances (e.g. pest outbreaks, fires), which constitutes a limitation of this study. Disturbance events such as fires or forest pests are common processes subtracting important amounts of biomass in Mediterranean landscapes. In the case of the planted pine forests such as the ones considered in this study, their sensitivity to fire is highly influenced by the specific pine species considered (Pausas et al. 2004), and therefore assessing the potential impacts of fire requires a detailed analysis of species distribution. Moreover, increasing temperatures are expected to favour some relevant pest species affecting pine trees such as Thaumetopoea pityocampa (Pureswaran et al. 2018). Since the incidence of these disturbances is expected to increase under climate change (Fyllas and Troumbis 2009; Seidl 2017), in order to assess the potential beneficial effects of climate change on forest productivity (Walker et al. 2019) more studies are required to investigate the interaction between disturbances, climate and management. Moreover, similarly to other process-based model studies, our landscape initialization and experiment design does not consider all possible processes, and therefore our results apply under some level of uncertainty. However, it is possible to hypothesize the impact of forest fires and other disturbances in our results if they would be included in our simulation experiment. The different growth and reproduction patterns between pines and oaks are key to make such as an inference (Gómez-Aparicio et al. 2009; Ruíz-Benito et al. 2012). We would expect higher mortality rates and lower biomass accrue for pines than for oaks if forest fires and other disturbances were considered.
The transformation of planted pine forests is driven by forest succession and accelerated by intensive management
Changes from pure pine forests towards mixtures of conifer and oaks and pure oaks arose as an emergent process from the above-described dynamics in photosynthesis, biomass and mortality in planted pine forests. Other studies also suggested that natural succession is causing a slow transformation of these forests into pine-oak mixtures (Dios et al. 2019). The marginal presence of oaks with the ability to resprout, the capacity of Q. ilex to colonise pine stands and the limited regeneration capacity of pines under dense canopies could explain this pattern (Gómez-Aparicio et al. 2009). Moreover, as pines reach their longevity limits (e.g. P. halepensis stands) without sufficient regeneration, windows of opportunity appear for new species to become dominant. Longer simulation experiments could give insights into this process. Once more, the previously mentioned absence of disturbances in our modelling approach, can be partially overcome by including indirectly current scientific evidence. By doing so, the disturbances would foster the pattern describe above. Forest fires and pest outbreaks would impact pine plantations more intensely and therefore acting as “natural management actions”.
Moreover, the highest rate of transformation occurred under extreme climate change due to the increased growth rate. Our findings suggest that climatic patterns across space can play an important role in the transformation of planted pine forests towards mixed forests. For instance, xeric areas showed clear transformation dynamics while in mesic and subhumid areas this alteration was less pronounced (see climate areas in Fig. 1). Xeric areas have shown to be more affected by recent climate change than milder areas (Peñuelas and Sardans 2021), which would explain the decreasing trend in biomass and the change in forest types under current and moderate climate change. One should note, however, that the distribution of species among climate areas is not homogeneous (Supplementary Figures SI39–SI45), and therefore these outcomes are the combined result of climatic effects and species distribution.
In addition to climatic effects, intense management practices can accelerate the conversion of xeric planted pine forests towards pine-oak mixed woodlands. Although no management scenario caused a full transformation from pine-dominated to mixed stands, scenarios with more intense management interventions promoted a higher rate of transformation than less intensive scenarios. This confirms that targeting management actions on low-diverse forests and therefore more vulnerable forests to climate change can proactively facilitate transition to better-adapted forest ecosystems in shorter time scales than by natural succession (Millar and Stephenson 2015). Moreover, our results support previous findings showing that management practices may have stronger impacts on forest structure and composition than the direct effects of climate change (Mina et al. 2017).
Conclusions and implications for management
This work aimed to deepen our understanding on how Mediterranean planted pine forests might evolve under different management and climate scenarios. Climate change is likely to induce a shift in the phenological growth pattern of these forests by reducing growth during summer but anticipating the spring growing phase. In the absence of natural disturbances, forest biomass will increase through time as stands mature and are further promoted by a longer growing season under climate change. However, this increase is different among species groups, resulting in a future landscape that will be more diverse in terms of forest types.
Nevertheless, the uncertainty associated with the projection of extreme weather events (droughts mainly) and the expected higher incidence of disturbances—herewith not considered—calls for precaution in driving optimistic conclusions about extreme climate change scenarios and will require managers to account for these uncertainties. Moreover, managers will be faced with the decision of promoting and enforcing forest composition change at the price of reducing biomass content. In this sense, uses such as carbon sequestration may be limited, but this limitation could become a perk under a context of expected higher incidence of forest fires.
Management can have a strong influence on future forest dynamics, likely promoting a shift from pure pine to oak and conifer-oak mixed woodlands, especially in xeric areas under more intense management scenarios. Although our outcomes portray natural succession as an important driver of change, they also confirm that choices in management regimes can accelerate changes in future landscape trajectories. In large, diverse and complex landscapes, however, such transformations do not occur homogeneously across spaces, such as in our mountain regions where changes will likely be more pronounced in xeric areas and small patches in subhumid and mesic areas. Our spatially-explicit modelling analysis could allow a prioritisation of areas to intervene, which could save precious resources by avoiding unnecessary interventions in areas where natural succession is already occurring. Instead, management should focus on those areas where such diversifying processes are being limited to promote more climate-adaptive ecosystems. In such areas, where lack of seed dispersal or excess of competition prevents natural diversification, small-scale highly intense interventions could be considered. Moreover, intensive management could still be causing a positive effect in all areas as it reduces competition, which could help these forests avoid the drought stress caused by climate change.
Finally, the underestimation of droughts in the projected climate time series and the absence of disturbances in our experiment would likely have important impacts in our results. These limitations may be resulting in biomass accrue overestimation. We expect that the inclusion of disturbances within the simulation would yield a more intense transformation of pine plantations into mixed stands. Nevertheless, further work is needed to explicitly overcome these limitations.
Data availability
The data supporting this study are archived in https://github.com/MARIASUAM/harvest_x_climate_LANDIS/releases/tag/v1.1 and will be made permanently available upon acceptance of this manuscript. LANDIS-II technical documentation is available at https://www.landis-ii.org/. The code is distributed under an open source licence at https://github.com/LANDIS-II-Foundation.
References
Allen CD, Breshears DD, McDowell NG (2015) On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6(8):art129
Boulanger Y, Pascual J, Bouchard M, D’Orangeville L, Périé C, Girardin MP (2021) Multi-model projections of tree species performance in Quebec, Canada under future climate change. Glob Change Biol 28:1884–1902
Bugmann H, Seidl R, Hartig F, Bohn F, Brůna J, Cailleret M, François L, Heinke J, Henrot A-J, Hickler T, Hülsmann L, Huth A, Jacquemin I, Kollas C, Lasch-Born P, Lexer MJ, Merganič J, Merganičová K, Mette T, Reyer CPO (2019) Tree mortality submodels drive simulated long-term forest dynamics: assessing 15 models from the stand to global scale. Ecosphere 10(2):e02616
Campos P, Díaz-Balteiro L (2015) Memorias científicas de RECAMAN Volumen 1 Economía y selviculturas de los montes de Andalucía. Consejo Superior de Investigaciones Científicas, Junta de Andalucía
Cassell BA, Scheller RM, Lucash MS, Hurteau MD, Loudermilk EL (2019) Widespread severe wildfires under climate change lead to increased forest homogeneity in dry mixed-conifer forests. Ecosphere. https://doi.org/10.1002/ecs2.2934
Coll L, Ameztegui A, Calama R, Lucas-Bor ME (2021) Dynamics and management of western Mediterranean pinewoods. In: Ne’eman G, Osem Y (eds) Pines and their mixed forest ecosystems in the Mediterranean basin. Springer, Berlin, pp 659–677. https://doi.org/10.1007/978-3-030-63625-8
Cornes RC, van der Schrier G, van den Besselaar EJM, Jones PD (2018) An ensemble version of the E-OBS temperature and precipitation data sets. J Geophys Res Atmos 123(17):9391–9409
De Bruijn A, Gustafson EJ, Sturtevant BR, Foster JR, Miranda BR, Lichti NI, Jacobs DF (2014) Toward more robust projections of forest landscape dynamics under novel environmental conditions: embedding PnET within LANDIS-II. Ecol Model 287:44–57
de Dios RS, Velázquez J, Ollero HS (2019) Classification and mapping of Spanish Mediterranean mixed forests. iForest Biogeosci for 5:480–487
de Wergifosse L, André F, Goosse H, Boczon A, Cecchini S, Ciceu A, Collalti A, Cools N, D’Andrea E, De Vos B, Hamdi R, Ingerslev M, Knudsen MA, Kowalska A, Leca S, Matteucci G, Nord-Larsen T, Sanders TG, Schmitz A, Jonard M (2022) Simulating tree growth response to climate change in structurally diverse oak and beech forests. Sci Total Environ 806:150422
de-Dios-García J, Pardos M, Calama R (2015) Interannual variability in competitive effects in mixed and monospecific forests of mediterranean stone pine. For Ecol Manag 358:230–239
Di Filippo A, Baliva M, Brunetti M, Di Fiore L (2021) Long-term tree-ring response to drought and frost in two Pinus halepensis populations growing under contrasting environmental conditions in peninsular Italy. Forests 12(3):305.
Durante P, Oyonarte C, Valladares F (2009) Influence of land-use types and climatic variables on seasonal patterns of NDVI in mediterranean iberian ecosystems. Appl Veg Sci 12(2):177–185
EEA (2016) European forest ecosystems, p 129
Fischer EM, Beyerle U, Knutti R (2013) Robust spatially aggregated projections of climate extremes. Nat Clim Change 3(12):1033–1038
Fyllas NM, Troumbis AY (2009) Simulating vegetation shifts in north-eastern Mediterranean mountain forests under climatic change scenarios. Glob Ecol Biogeogr 18(1):64–77
García-Duro J, Ciceu A, Chivulescu S, Badea O, Tanase MA, Aponte C (2021) Shifts in forest species composition and abundance under climate change scenarios in southern Carpathian Romanian temperate forests. Forests 12(11):1434
Gea-Izquierdo G, Cherubini P, Cañellas I (2011) Tree-rings reflect the impact of climate change on Quercus ilex L. Along a temperature gradient in Spain over the last 100 years. For Ecol Manag 262(9):1807–1816
Gea-Izquierdo G, Nicault A, Battipaglia G, Dorado-Liñán I, Gutiérrez E, Ribas M, Guiot J (2017) Risky future for Mediterranean forests unless they undergo extreme carbon fertilization. Glob Change Biol 23(7):2915–2927
Giorgi F (2006) Climate change hot-spots. Geophys Res Lett 33(8):L08707
Gómez-Aparicio L, Zavala MA, Bonet FJ, Zamora R (2009) Are pine plantations valid tools for restoring Mediterranean forests? An assessment along abiotic and biotic gradients. Ecol Appl 19(8):2124–2141
Gustafson EJ, Shifley SR, Mladenoff DJ, Nimerfro KK, He HS (2000) Spatial simulation of forest succession and timber harvesting using. Can J for Res 30:32–43
Gustafson EJ, De Bruijn AMG, Pangle RE, Limousin JM, Mcdowell NG, Pockman WT, Sturtevant BR, Muss JD, Kubiske ME (2015) Integrating ecophysiology and forest landscape models to improve projections of drought effects under climate change. Glob Change Biol 21(2):843–856
Hong SH, Mladenoff DJ (1999) Spatially explicit and stochastic simulation of forest-landscape fire disturbance and succession. Ecology 80:81–99
Junta de Andalucía CdeMA (2004) Plan de Ordenación de Recursos Naturales del Parque Natural Sierra de Baza
Junta de Andalucía CdeMA (2011) Plan de Ordenación de los Recursos Naturales de Sierra Nevada
Karger DN, Schmatz DR, Dettling G, Zimmermann NE (2020) High-resolution monthly precipitation and temperature time series from 2006 to 2100. Sci Data 7(1):1–10
Madrigal-González J, Andivia E, Zavala MA, Stoffel M, Calatayud J, Sánchez-Salguero R, Ballesteros-Cánovas J (2018) Disentangling the relative role of climate change on tree growth in an extreme Mediterranean environment. Sci Total Environ 642:619–628
Mann HB (1945) Nonparametric tests against trend. Econometrica 13:245–259
Mesa Garrido MÁ (2019) Reforestación, silvicultura e incendios forestales en la dinámica del paisaje del Espacio Natural de Sierra Nevada (1881–2018). Reforestation, forestry and wildfires in the dynamics of the Sierra Nevada Natural Area (1881–2018). Investigaciones Geográficas 71:209–234
Messier C, Bauhus J, Sousa-Silva R, Auge H, Baeten L, Barsoum N, Bruelheide H, Caldwell B, Cavender-Bares J, Dhiedt E, Eisenhauer N, Ganade G, Gravel D, Guillemot J, Hall JS, Hector A, Hérault B, Jactel H, Koricheva J, Zemp DC (2022) For the sake of resilience and multifunctionality, let’s diversify planted forests! Conserv Lett 15(1):e12829
Millar CI, Stephenson NL (2015) Temperate forest health in an era of emerging megadisturbance. Science 349(6250):823–826
Mina M, Bugmann H, Cordonnier T, Irauschek F, Klopcic M, Pardos M, Cailleret M (2017) Future ecosystem services from European mountain forests under climate change. J Appl Ecol 54(2):389–401
Mina M, Messier C, Duveneck M, Fortin M-J, Aquilué N (2021) Network analysis can guide resilience-based management in forest landscapes under global change. Ecol Appl 31(1):e2221
Mina M, Messier C, Duveneck MJ, Fortin M-J, Aquilué N (2022) Managing for the unexpected: Building resilient forest landscapes to cope with global change. Glob Change Biol 28(14):4323–4341
Navarro Cerrillo R, Varo M, Lanjeri S, Hernández Clemente R (2007) Cartografía de defoliación en los pinares de pino silvestre (Pinus sylvestris L.) y pino salgareño (Pinus nigra Arnold.) en la Sierra de los Filabres. Ecosistemas 16(3):160–168
Navarro-Cerrillo RM, Rodriguez-Vallejo C, Silveiro E, Hortal A, Palacios-Rodríguez G, Duque-Lazo J, Camarero JJ (2018) Cumulative drought stress leads to a loss of growth resilience and explains higher mortality in planted than in naturally regenerated Pinus pinaster stands. Forests 9(6):358
Navarro-Cerrillo RM, Sánchez-Salguero R, Rodriguez C, Duque Lazo J, Moreno-Rojas JM, Palacios-Rodriguez G, Camarero JJ (2019) Is thinning an alternative when trees could die in response to drought? The case of planted Pinus nigra and P. sylvestris stands in southern spain. For Ecol Manag 433:313–324
Oettel J, Lapin K (2021) Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol Indic 122:107275
Pardos M, del Río M, Pretzsch H, Jactel H, Bielak K, Bravo F, Brazaitis G, Defossez E, Engel M, Godvod K, Jacobs K, Jansone L, Jansons A, Morin X, Nothdurft A, Oreti L, Ponette Q, Pach M, Riofrío J, Calama R (2021) The greater resilience of mixed forests to drought mainly depends on their composition: analysis along a climate gradient across Europe. For Ecol Manag 481:118687
Pausas JG, Blade C, Valdecantos A, Seva JP, Fuentes D, Alloza JA, Vilagrosa A et al (2004) Pines and oaks in the restoration of Mediterranean landscapes of Spain: new perspectives for an old practice—a review. Plant Ecol 171(1–2):209–220
Peñuelas J, Sardans J (2021) Global change and forest disturbances in the Mediterranean basin: breakthroughs, knowledge gaps, and recommendations. Forests 12(5):603
Pureswaran DS, Roques A, Battisti A (2018) Forest insects and climate change. Curr for Rep 4:35–50
R Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
REDIAM (2006) Mapa de Usos del Suelo y Coberturas Vegetales a escala 1:10.000 de la Masa Forestal de Andalucía, año 1996–2006. Consejería de Medio Ambiente, Junta de Andalucía
Rubio-Cuadrado Á, Camarero JJ, Aspizua R, Sánchez-González M, Gil L, Montes F (2018) Abiotic factors modulate post-drought growth resilience of scots pine plantations and rear-edge scots pine and oak forests. Dendrochronologia 51:54–65
Ruiz-Benito P, Gómez-Aparicio L, Zavala MA (2012) Large-scale assessment of regeneration and diversity in Mediterranean planted pine forests along ecological gradients. Divers Distrib 18(11):1092–1106
Sánchez-Salguero R, Camarero JJ, Dobbertin M, Fernández-Cancio Á, Vilà-Cabrera A, Manzanedo RD, Zavala MA, Navarro-Cerrillo RM (2013) Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For Ecol Manag 310:956–967
Scheller RM, Domingo JB, Sturtevant BR, Williams JS, Rudy A, Gustafson EJ, Mladenoff DJ (2007) Design, development, and application of LANDIS-II, a spatial landscape simulation model with flexible temporal and spatial resolution. Ecol Model 201(3–4):409–419
Seidl R (2017) To model or not to model, that is no longer the question for ecologists. Ecosystems 20(2):222–228
Senf C, Seidl R (2022) Post-disturbance canopy recovery and the resilience of Europe’s forests. Glob Ecol Biogeogr 31(1):25–36
Suárez-Muñoz M, Mina M, Salazar PC, Navarro-Cerrillo RM, Quero JL, Bonet-García FJ (2021) A step-by-step guide to initialize and calibrate landscape models: a case study in the Mediterranean mountains. Front Ecol Evol. https://doi.org/10.3389/fevo.2021.653393
Tramblay Y, Koutroulis A, Samaniego L, Vicente-Serrano SM, Volaire F, Boone A, Polcher J (2020) Challenges for drought assessment in the Mediterranean region under future climate scenarios. Earth Sci Rev 210:103348
Ueyama M, Ichii K, Kobayashi H, Kumagai T, Beringer J, Merbold L, Euskirchen ES, Hirano T, Marchesini LB, Baldocchi D, Saitoh TM, Mizoguchi Y, Ono K, Kim J, Varlagin A, Kang M, Shimizu T, Kosugi Y, Bret-Harte MS, Yasuda Y (2020) Inferring CO2 fertilization effect based on global monitoring land-atmosphere exchange with a theoretical model. Environ Res Lett 15:084009
Waitz Y, Sheffer E (2021) Dynamics of Mixed Pine-Oak Forests. In: Neeman G, Osem Y (eds) Pines and their mixed forest ecosystems in the Mediterranean basin. Springer, Cham, pp 345–362. https://doi.org/10.1007/978-3-030-63625-8
Walker XK, Baltzer JL, Cumming SG, Day NJ, Ebert C, Goetz S, Johnstone JF, Potter S, Rogers BM, Schuur EAG, Turetsky MR, Mack MC (2019) Increasing wildfires threaten historic carbon sink of boreal forest soils. Nature 572:520–523
Zittis G, Bruggeman A, Lelieveld J (2021) Revisiting future extreme precipitation trends in the Mediterranean. Weather Clim Extremes 34:100380
Acknowledgements
R.M.N.C. was funded by SILVADAPT.NET (RED2018-102719-T), EVIDENCE (Ref: 2822/2021) and REMEDIO (PID2021-128463OB-I00) projects. We acknowledge the data provided by E-OBS and ECA&D project (https://www.ecad.eu). We thank Dirk Karger from WSL Birmensdorf for the help in retrieving the climate data. We also thank Matthew Duveneck for his comments on earlier versions of this manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. M.S.M. was supported by the Spanish Ministry of Science and Innovation [FPU predoctoral grant]. M.M. acknowledges funding from the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie framework (Grant No. 891671, REINFORCE project).
Author information
Authors and Affiliations
Contributions
Conceptualization: M.S.M, M.M., R.N.C., F.J.B.G.; Methodology: M.S.M, M.M.; Visualisation, Formal analysis, Investigation and Writing—Original Draft: M.S.M; Data curation: M.S.M, J.H.; Writing—Review & Editing: M.S.M, M.M., R.N.C., J.H., F.J.B.G.; Resources: M.M., R.N.C., F.J.B.G.; Supervision: F.J.B.G., M.M.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Suárez-Muñoz, M., Bonet-García, F.J., Navarro-Cerrillo, R. et al. Forest management scenarios drive future dynamics of Mediterranean planted pine forests under climate change. Landsc Ecol 38, 2069–2084 (2023). https://doi.org/10.1007/s10980-023-01678-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-023-01678-y