
Abstract
Purpose
Mixtures of cover crops are gaining importance in farming systems. While complementarity between species is a potentially important mechanism underlying mixture benefits, there is little quantitative evidence about complementarity of root growth. Therefore, we aimed to gain detailed knowledge about mixture effects of specific root traits.
Methods
Blue lupin (Lupinus angustifolius L.) and winter rye (Secale cereale L.) were grown as sole and intercrops in a two-year organically managed field experiment in Germany. Crops were analysed for shoot biomass and nutrient uptake as well as for root mass density (RMD), root length density (RLD) and specific root length (SRL). We used Fourier transform infrared (FTIR) spectroscopy to discriminate the root mass species-specifically in the mixtures.
Results
In the shoot, overyielding was only found for N and P uptake in the 75:25 mixture of lupin and rye, whereas positive mixture effects of RLD, RMD, and root mass complementarity occurred only in the 50:50 mixture in one year. Consistently positive mixture effects of SRL were found. Root biomass proportion of rye was higher in topsoil, that of lupin higher in subsoil. Vertical niche differentiation in RMD between sole crops and mixtures was not observed.
Conclusion
Among all root traits analysed, consistent mixture effects were observed only for SRL. Mechanisms behind mixture effects of root traits need further research. FTIR spectroscopy discriminated intercrop root mass well. Discrimination of mixture partners’ root length would help to better understand complementarity of root growth in mixtures.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Benefits of cover crops in arable farming systems such as nitrogen (N) retention and green manuring (Thorup-Kristensen et al. 2003), weed suppression (Hartwig and Ammon 2002), improvement of soil physical quality (Bodner et al. 2014), carbon sequestration (Jian et al. 2020) and climate change mitigation (Kaye and Quemada 2017) are widely understood and described. The significance of cover crop mixtures has increased in practical farming in the last years.
Benefits and synergistic interactions of crop mixtures (Bedoussac et al. 2015) and their management (Yu et al. 2016) are well understood, especially cereal-legume intercrops due to their complementary N use (Hauggaard-Nielsen et al. 2001a; Jensen 1996). Many studies examined mixture effects in shoot growth of cover crops (Couëdel et al. 2018; Elhakeem et al. 2019; Tosti et al. 2012, 2014; Tribouillois et al. 2016; Wendling et al. 2017). Compared to pure stands, cover crop mixtures can have positive effects for shoot biomass and shoot N uptake (Elhakeem et al. 2019) and transgressive overyielding of shoot biomass is possible due to complementarity (Wendling et al. 2017).
Cover crop mixtures of legumes and non-legumes can effectively be used to minimize the trade-offs between N retention and N supply for the subsequent crop (Couëdel et al. 2018; Tosti et al. 2012, 2014; Tribouillois et al. 2016). For crucifer-legume cover crop mixtures it was shown by Tribouillois et al. (2016) and Couëdel et al. (2018) that the catch crop service, i.e. the nitrate leaching reduction, of the mixtures is as high as of crucifer sole crop and that green manure service, i.e. the N availability for the next crop, is higher than for crucifer sole crop.
Despite the known benefits of cover crop mixtures and the importance of belowground residues for soil quality, especially soil organic carbon (Poeplau et al. 2021), and the significance of root intensity for N retention (Thorup-Kristensen 2001), less is known about root traits of cover crop mixtures from field experiments. Heuermann et al. (2019) used a qPCR-based method to analyse root biomass in mixtures of white mustard (Sinapis alba L.), Egyptian clover (Trifolium alexandrinum L.), phacelia (Phacelia tanacetifolia Benth.), bristle oat (Avena strigosa Schreb.) and corresponding sole crops. They found root biomass overyielding in the mixture, root biomass dominated by mustard and phacelia which suppressed clover and oat as well as only low niche differentiation. Bukovsky-Reyes et al. (2019) investigated mixtures and sole crops of hairy vetch (Vicia villosa Roth.) and cereal rye (Secale cereale L.). They determined increased total root length and specific root length (SRL) in root traits in mixture compared to sole crop for vetch but not for rye.
More general knowledge about roots of intercrops is recognised from field experiments with main crops or from rhizotron studies. Using 32P technique in a field experiment with a mixture of pea (Pisum sativum L.) and barley (Hordeum vulgare L.), Hauggaard-Nielsen et al. (2001b) determined that barley root growth was faster than that of pea, and barley rooted deeper in mixture. This explains the competitive ability of cereals for soil inorganic N. Similarly, dominance of root growth of barley over vetch (Vicia sativa L.) was shown in a rhizotron experiment with a barley-vetch-mixture using microscopy for species identification (Ramirez-Garcia et al. 2015); moreover, they determined higher root intensity in deep layers in the mixture compared to barley pure stand. Streit et al. (2019) detected higher overyielding of root biomass than of shoot biomass in intercrops of winter wheat (Triticum aestivum L.) and winter faba bean (Vicia faba L.). Using Fourier transform infrared (FTIR) spectroscopy, which enables separation of root mass from intercropping partners, they did not find dominance of cereal biomass in root and shoot over legume biomass in contrast to other studies. Tosti and Thorup-Kristensen (2010) demonstrated in rhizotron experiment that legume roots were highly affected by interspecific competition with red beet (Beta vulgaris L.) and that fast initial root growth was important for competition. Similarly, Andersen et al. (2014) showed that more roots were observed in the mixtures than predicted, and that red clover (Trifolium pratense L.) root growth dominated in the topsoil, while red beet was dominant in deep layers.
In faba bean mixtures with oil crops root length density (RLD) in the subsoil increased in contrast to faba bean sole crop (Schröder and Köpke 2012). In intercrops of wheat and maize (Zea mays L.), wheat roots spread underneath maize mainly in deeper soil layers, RLD of wheat and maize was higher when intercropped than in sole crops (Li et al. 2006). Faba bean had shallow roots in mixture with maize, whereas roots of intercropped maize spread underneath faba bean. Thus, Li et al. (2006) concluded that greater soil exploration in the mixtures contributed to the grain yield advantage. The ‘three sisters’ maize, bean (Phaseolus vulgaris L.) and squash (Cucurbita pepo L.) yielded higher in mixture compared to sole crops and their root volume density was higher in mixture than expected from the sole crops (Zhang et al. 2014). Grassland species mixtures showed root biomass overyielding and below-ground complementarity, but no evidence for vertical niche differentiation (Mommer et al. 2010).
As indicated in these studies, possible positive mixture effects on roots are higher root intensity, root length and volume density as well as root biomass in mixtures compared to sole crop. In some studies, also the spatial distribution of the roots was described, and in some cases complementary root distribution was observed with one mixture partner spreading mainly in the upper soil layers and the other partner mainly in the subsoil (Andersen et al. 2014; Li et al. 2006). Complementarity occurs when plants in a mixture require resources differently in space, time or form or one mixture partner facilitates the resource availability (Cardinale et al. 2007; Justes et al. 2021; Tilman et al. 2001), and thus, reduces interspecific compared to intraspecific competition (Mulder et al. 2001). Hence, complementarity consists of both niche differentiation and facilitation (Loreau and Hector 2001). Vertical root niche differentiation is a possible underlying mechanism of spatial complementarity. Besides mixture effects of root traits, possible vertical niche differentiation of root growth which may allow below-ground complementarity is interesting to be investigated for cover crop mixtures. In our study, we attempted to maximize complementary root distribution by mixing homorhizous and allorhizous cover crops as suggested in Kemper et al. (2020), because different vertical rooting patterns may intensify niche differentiation. As a homorhizous crop we chose winter rye (Secale cereale L.) and blue lupin (Lupinus angustifolius L.) as an allorhizous crop. Besides vertical rooting patterns, roots of lupin and rye differently affect soil pore dynamics (Bodner et al. 2014) and access complementary nitrogen pools. Blue lupin can fix atmospheric N, thus, provide additional N input to the cropping system and may improve growth conditions for the subsequent crop. We chose a rye-lupin mixture since cover crop mixtures of non-legumes and legumes are known to minimize the trade-off between nitrate leaching reduction and N supply for the subsequent crop (Couëdel et al. 2018; White et al. 2017).
The aim of our study was to investigate cover crop mixtures of blue lupin and winter rye and its corresponding sole crops for the root traits root mass, root mass density, root length density, specific root length as well as shoot biomass and shoot nutrient uptake. We used FTIR spectroscopy to analyse the share of root mass of the mixture partners. We hypothesized that:
-
(1)
cover crop mixtures of lupin and rye show a positive mixture effect for shoot biomass and shoot nutrient accumulation,
-
(2)
cover crop mixtures of lupin and rye show a positive mixture effect for RLD and root mass,
-
(3)
in cover crop mixtures root biomass proportion of rye in topsoil is higher compared to lupin and that root biomass proportion of lupin in subsoil is higher compared to rye
-
(4)
vertical RMD profile of lupin and rye differs between sole crops and mixture.
Materials and methods
Experimental site
The field trials were carried out on a Fluvisol with a silt loam texture on gravel layers beginning between 60 and 200 cm of soil depth and a shallow groundwater table at the experimental station for Organic Farming ‘Wiesengut’ of the University of Bonn in Hennef (50°48′ N, 7°17′ E), Germany, with a mean annual precipitation of 840 mm and a mean temperature of 10.3 °C. More detailed information about site, soil and climate are given in Kemper et al. (2020).
Experimental design and management practices
The field trials were laid out as a randomized complete block design with four replicates. Plot size was 1.5 m × 12.5 m in 2018 and 1.5 m × 20 m in 2019. Three replicates were examined for root traits. Pre-crop history of the trial in 2018 was spring wheat (Triticum aestivum L.) in 2015 (18 t ha−1 farmyard manure), potatoes (Solanum tuberosum L.) in 2016 (30 t ha−1 farmyard manure), spring wheat in 2017 (18 t ha−1 lime) and spring faba bean (Vicia faba L.) as a direct pre-crop in 2018. Pre-crop history of the trial in 2019 was spring oat (Avena sativa L.) in 2016 (18 t ha−1 farmyard manure), potatoes in 2017 (30 t ha−1 farmyard manure), spring oat in 2018 and spring faba bean as a direct pre-crop in 2019. After harvest of pre-crop, tillage, irrigation and mechanical and hand weeding were performed as described in detail in Kemper et al. (2020). Briefly, tillage was done with a mouldboard plough and a rotary harrow; 104 mm and 42 mm were irrigated in 2018 and 2019, respectively. The cover crops were sown with a plot seeder with row distance of 17 cm on 12th August 2018 and on 14th August 2019. Sole crops of fibrous rooted winter rye (WR, Secale cereale L., cv. Bonfire) and taprooted blue lupin (LU, Lupinus angustifolius L., cv. Boruta) were grown each with a seed rate of 120 kg/ha. Intercrops were sown as mixed and substitutive intercrops (Hauggaard-Nielsen et al. 2008) with proportions of 50:50 (LU_WR_50_50) in both years and additionally 75:25 (LU_WR_75_25) in 2019. The content of mineral nitrogen at the time of sowing for both experimental years is given in Table 1.
Shoot harvest and nutrient analysis
Shoot biomass was harvested on 15th November in 2018 and on 13th November in 2019 on 1.0 m2 per plot and was dried at 60 °C for 24 h and at 105 °C for further 24 h. Dry matter samples were milled with a vibratory disc mill (RS 200 Retsch, Haan, Germany). Milled samples were analysed for N content with Dumas method using an Elemental analyser (Eurovector EA 3000, Pavia, Italy). After microwave digestion milled samples were analysed for phosphorous (P) content photometrically using a continuous flow analyser (Seal QuAAtro 39, Norderstedt, Germany) and for potassium (K) content with atomic absorption spectrometry using Analyst 200 (PerkinElmer, Waltham, USA).
Soil mineral nitrogen
Soil samples for analysis of soil mineral nitrogen (Nmin) were taken using a Puerckhauer auger. Samples in soil depths 0–30 cm and 30–60 cm were taken in both years and in 2019 also for 60–90 cm. Four samples per plot were mixed for 30–60 cm and 60–90 cm, eight samples per plot were mixed for 0–30 cm. In 2018 soil samples were taken on 8th and 9th November and in 2019 on 13th November. Both nitrate and ammonium were determined photometrically using a continuous flow analyser (Seal QuAAtro 39, Norderstedt, Germany) after extraction with potassium sulphate.
Monolith method
In October 2018 and 2019 a trench (1.2 deep; 1.0 m wide) was established with an excavator transversely to the plots for three field replications. The monolith method by Böhm (1979) was used to extract monoliths down to a soil depth varying between 60 and 100 cm depending on the gravel layer appearing. Monoliths in 10 cm intervals of soil depth were collected. In 2018 two monoliths next to each other of 10 cm × 10 cm × 20 cm and in 2019 two monoliths next to each other in the topsoil (0-30 cm) and three monoliths next to each other in the subsoil (30-100 cm) of 10 cm × 10 cm × 17 cm were taken. With regard to the width of the plot, the monoliths were taken in the plot centre to minimize edge effects. After sampling in the field and before washing, monoliths were kept refrigerated at 5° C.
Subsequently, roots were washed from monoliths by soaking in water for easier disaggregation by hand. The water with the dissolved monolith was poured through a stack of sieves until the soil was completely dissolved. In 2018 sieves of 2.0 and 1.0 mm mesh size were used. In 2019 sieves of 4.0, 2.0, 1.0, 0.71, 0.63 and 0.5 mm mesh size were used. The roots recovered by the single sieves were washed off from the sieves and soil, stones and some particular organic matter were removed by repeated sedimentation and decantation. Until further processing, roots were kept refrigerated in water at 5° C. Roots were manually separated from remaining non-root material, especially particular organic matter, by tweezers. In 2019, roots recovered in the 0.5 mm sieve were not separated from particular organic matter. Thus, these roots were analysed for root length but not for root mass, to minimize the error by the mass of the non-root material, as roots recovered in sieves with smaller mesh size contribute to a high extent to root length, but their contribution to root mass is negligible (Livesley et al. 1998).
Washed roots were scanned with a photo scanner (Expression 12000XL, Epson, Suwa, Japan) at 800 dpi and root-length was analysed with WinRHIZO (Version WinRhizoPro 2019a 64bit, Regent Instruments, Quebec, Canada). For determination of root dry matter, roots were dried at 55 °C for 48 h.
Fourier transform infrared spectroscopy
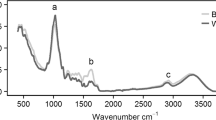
The roots of winter rye (WR) and blue lupin (LU) of the years 2018 and 2019 were used to evaluate the species root proportion of the mixed species samples. The dried roots of the single species samples and mixed species samples harvested with the monolith method were ground to 0.2 mm (ultra-centrifugal mill, ZM 200, Retsch, Haan, Germany). Absorption spectra were measured by the FTIR-ATR spectrometer (Alpha-P with a diamond crystal attenuated total reflection (ATR) device, Bruker Optics, Ettlingen, Germany) with a resolution of 4 cm−1 and 32 scans in the spectral range of 4,000–400 cm−1. Each sample was measured 5 times. The evaluation of the FTIR-ATR spectra was conducted with the Opus software Quant 2 (version 7.2, Bruker Optics, Ettlingen, Germany).
The FTIR spectra of the single sample species were used for cluster analysis (Opus software, version 7.2, Bruker Optics) to separate the species. For the cluster analyses, the spectra were pre-processed by the second derivative and vector normalization, the frequency range was reduced to 1800–850 cm−1 and the Euclidian’s distance and Ward’s algorithm was applied (see Figs. 1 and 2, supplementary material). For 2018 and 2019, the interspecific heterogeneity for both species (9.426 and 4.303, respectively) was higher than the intraspecific heterogeneity (1.068 and 0.523 for WR, and 4.711 and 1.169 for LU, respectively), permitting separation of the two species.
For the quantification of the proportion of the species in the mixed species root samples, the FTIR spectra of the single species samples were used to generate a model. The details in establishing and evaluating the model were added in the supplementary material.
With the chosen model for each year, the FTIR spectra of the mixed species samples (of the corresponding year) were evaluated with the associated model. The output of this evaluation was the percental share of each species within the mixed species root samples. The percentage of the single species were used for further calculations.
Calculations
For each monolith, root length density (RLD), root mass density (RMD) and specific root length (SRL) was calculated according to Eq. 1, 2 and 3. Units of RLD, RMD and SRL are cm cm−3, mg cm−3 and m g−1, respectively.
For the statistical comparison of RLD, RMD and SRL the following combined soil layers were used: 0–10 cm, 10–30 cm, 30–60 cm and 60–100 cm.
Shoot performance and root mass of cover crop mixtures were analysed using the land equivalent ratio (LER) concept (Mead and Willey 1980). LER is the relative land area under sole cropping that is required to produce the same yields as under intercropping. The land use efficiency of an intercrop will be superior to that of corresponding sole crops if LER is > 1. LER is the sum of the partial land equivalent ratios pLER (Eq. 4). Yi is shoot or root dry mass or shoot nutrient uptake (g m−2) in the intercrop of species i and Mi is shoot or root dry mass or shoot nutrient uptake (g m−2) of the sole crop i.
Root and shoot performance of cover crop mixtures was analysed using the relative mixture effect (RME) concept. RME (Eq. 5) was calculated for the root traits RMD, RLD and SRL as well as for shoot biomass and shoot nutrient uptake. RME is the percentage difference between the observed value in the cover crop mixture and expected value, which is the average of the sole cover crops weighted by relative density pi. Relative density is the density in mixture divided by density in sole crop.
Additionally, we use the concept of Loreau and Hector (2001) partitioning the net biodiversity effect into complementarity and selection effect and the notation of Elhakeem et al. (2019). NE is the net biodiversity effect (g m−2) which is equal to the absolute mixture effect, i.e. the difference between observed and expected yield of the mixture. Yi is the yield of species i in mixture (g m−2), Mi is the yield of species i as sole crop (g m−2) and pi is the relative density (Eq. 6). NE is the sum of CE and SE (Eq. 7). CE is the complementarity effect (g m−2) and SE is the selection effect (g m−2).
CE is calculated as the number of species (N) multiplied with the average relative yield gain (∆R̅Y) and M̅ which is the average yield of sole crops (Eq. 8). The relative yield gain (∆RY) is the difference between observed and expected relative yield (Eq. 9).
CE indicates whether the mixed species benefit from facilitation or niche differentiation or whether species interfere with each other. CE is positive if the yield of both partners is higher than expected or the relative yield gain of one partner is higher than the relative yield loss of the other partner.
SE is calculated by the number of species multiplied by the covariance between relative yield gain and sole crop yields (Eq. 10). SE shows the dominance by one species. It indicates the relation between the yield of sole crops and the relative yield gain in the mixture of the corresponding species. SE is positive if the relative yield gain of the species with the higher yield in sole crop is relatively higher. SE is negative if the relative yield gain of the species with the lower yield in sole crop is relatively higher.
Statistical analysis
Data were analysed with the programme R version 3.5.2 with R studio version 1.1.463 (R Core Team 2018).
Shoot biomass, root biomass, root-shoot-ratio, mean of RLD, RMD, SRL and Nmin in combined soil layers were analysed by a one-factorial analysis of variance (Anova). Homogeneity of variances was checked with a plot of residuals versus fitted values and normal distribution was tested with a normal quantile–quantile (QQ) plot (Kozak and Piepho 2018). If necessary, data were square root or logarithm transformed. After Anova, mean values of treatments were compared with a Tukey test at a significance level of α = 0.05 by using the agricolae package (Mendiburu 2019).
The percentage share of root mass and the difference between expected and observed shoot biomass, root biomass, RMD and pLER was tested with a two sample Welch t-test. The significance of LER > 1 was tested by a one sample t-test against 1. The significance of RME was tested by a one sample t-test against 0. Pearson’s correlation between RME of root traits were calculated.
Results
The growth period of 2018 was characterized by an extreme dry spell from July to November leading to extremely dry soil conditions in autumn. The summer of 2019 was dry as well, especially in June and July, but less extreme than 2018. Both growing periods were warmer compared to the long-term mean. Soil moisture content, recorded at a nearby weather station, was much higher in October and November in 2019 than in 2018 (Fig. 1).

Soil moisture in % of plant-available water capacity under grass on a sandy loam between 0 and 60 cm soil depth at the nearby (12 km) weather station from the German Meteorological Service at Cologne Bonn Airport in the period from two weeks before sowing until biomass sampling of cover crops for both experimental years
Root and shoot biomass
In both years total shoot dry mass (DM) did not differ significantly between cover crops (Fig. 2). The shoot proportion of lupin was significantly lower than expected from sowing proportion in both years, whereas the shoot proportion of rye was significantly higher than expected in both years.

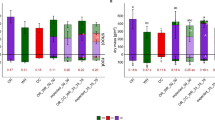
Mean values ± standard deviation of shoot (n = 4) and root (n = 3) dry mass [g/m2] and root-shoot-ratio (n = 3) of sole cover crops blue lupin (LU), winter rye (WR), 50:50 and 75:25 cover crop mixtures of blue lupin and winter rye (LU_WR_50_50 and LU_WR_75_25) and of expected mixtures (corresponding proportions of sole crops) in 2018 (A) and 2019 (B). Bars above 0 represent shoot dry mass, bars below 0 show root dry mass. Bar colours refer to species: green for rye, blue for lupin. Transparent bars represent expected values. Red numbers below bars represent root-shoot-ratio. Different lowercase letters indicate significant differences (p ≤ 0.05, HSD Tukey-test) between different cover crops for total shoot or total root dry mass or root-shoot-ratio within one year. Different uppercase letters indicate significant differences (p ≤ 0.05, t-test) between observed and expected dry mass for shoot or root of intercrop partner within one year
Total root DM did not differ significantly in 2018, but differed in 2019. For the 50:50-mixture, in 2018 root DM of winter rye was significantly higher than expected. In 2019 expected and observed root DM of mixtures did not differ significantly. Rye dominated root and shoot DM compared to lupin in both years and mixtures. The 75:25-mixture showed more balanced partners in shoot and root DM than the 50:50-mixture. In 2018, the root-shoot-ratios of rye and the 50:50-mixture were significantly higher than that of lupin. In 2019, the root-shoot-ratios of lupin and both mixtures were significantly lower than that of rye.
In the LER diagram (Fig. 3) all shoot values were below the diagonal dashed line for both mixtures. Thus, rye had competitive advantage over lupin in the shoot. For the 50:50-mixture, all shoot values for pLER of lupin were below 0.5 and for pLER of rye were above 0.5. For the 75:25-mixture all shoot values for pLER of lupin were below 0.75 and for pLER of rye were above 0.25. In the shoot blue lupin was suppressed by winter rye. For the shoot in 2018 and 2019, mean LER of the 50:50-mixture was 1.04 and 1.03, respectively and mean RME were -0.3% and -0.4%, respectively. For the 75:25-mixture in 2019 mean shoot LER was 1.22 and mean shoot RME was 17.3%. For the root mass, LER values for the 50:50 mixture in 2018 were all above 1 (mean 1.50), whereas in 2019 they were below 1 (mean 0.52). Two root LER values for the 75:25 mixture were below 1, one was above 1. The mean was 0.65.

Partial land equivalent ratio of shoot dry mass (circle) and root dry mass (triangle) of blue lupin (pLER LU) and winter rye (pLER WR) and relative mixture effect [%] (RME) of shoot dry mass in a 50:50-mixture (A) and in a 75:25-mixture (B) in two years 2018 (black symbols) and 2019 (grey symbols). RME of root mass is given in Fig. 6. The solid line corresponds to a land equivalent ratio of 1 (LER = pLER LU + pLER WR). The dashed horizontal and vertical lines represent the expected pLER for the mixture partners. Numbers adjacent to symbols represent RME
Shoot nutrient uptake and soil mineral nitrogen depletion
LER and RME were only significant for N and P shoot uptake in the 75:25-mixture (Table 2). All pLER of shoot nutrient uptake were higher than expected for rye, lower than expected for lupin.
In 2018, N shoot uptake of lupin was significantly higher than that of rye. In 2019, N shoot uptake of the 75:25-mixture was significantly higher than that of rye. In both years, in the 50:50-mixture pLER of lupin for N shoot uptake was significantly lower than 0.5, and pLER of rye for N shoot uptake was significantly higher than 0.5. In 2019, in the 75:25-mixture pLER of rye for N shoot uptake was significantly higher than expected, LER (1.53 ± 0.16) was significantly > 1 and RME (29.60% ± 9.88%) significantly > 0.
In 2018, P shoot uptake was significantly lower in lupin than in rye and the 50:50-mixture. In 2019, a significantly higher P shoot uptake in the 75:25-mixture than in lupin was detected. In both years and for both mixtures, pLER of lupin for the P shoot uptake was significantly lower than expected and pLER of rye was significantly higher than expected. In 2019, in the 75:25-mixture LER (1.40 ± 0.17) was significantly > 1 and RME (50.53% ± 12.86%) significantly > 0.
K shoot uptake did not significantly differ between any of the cover crops in any of the years. In both years and for both mixtures, pLER of lupin for K shoot uptake was significantly lower than expected and pLER of rye was significantly higher than expected. None of the LER or RME values significantly differed from 1 or 0, respectively.
Cover crops significantly reduced soil Nmin compared to the control by 80–130 kg/ha (Fig. 4). Mixtures of blue lupin with winter rye reduced soil Nmin significantly more compared to lupin sole crop and were as effective as rye sole crop in reduction of inorganic N. Differences were especially high in the soil layers of 30–60 cm and 60–90 cm.

Mean values ± standard deviation (n = 4) of soil mineral nitrogen [kg/ha] (Nmin) under a weed-free control, sole cover crops blue lupin (LU), winter rye (WR), 50:50 and 75:25 cover crop mixtures of lupin and rye (LU_WR_50_50 and LU_WR_75_25) in 2018 (A) and 2019 (B). Bar colours refer to soil layer: light grey for 0–30 cm, white for 30–60 cm, dark grey for 60–90 cm. Different lowercase letters indicate significant differences (p ≤ 0.05, HSD Tukey-test) between different treatments for total Nmin within one year. Different uppercase letters indicate significant differences (p ≤ 0.05, HSD Tukey-test) between different treatments for Nmin within each soil layer within one year
Root length density, root mass density and specific root length
Topsoil RLD was high for rye and low for lupin. In 2018, mixtures were similar to rye, and intermediate in 2019 (Fig. 5). Lupin had significantly lower RLD in 0–30 cm than rye and 50:50-mixture in 2018. In 2019, in 0–10 cm soil depth rye and 50:50-mixture had significantly higher RLD than lupin, whereas 75:25-mixture and lupin had significantly lower RLD than rye. In 10–30 cm soil depth rye had significantly higher RLD than lupin, both mixtures were intermediate. No significant differences for subsoil RLD were recorded.

Mean values ± standard deviation (n = 3) of root length density [cm/cm3] (RLD) (A, B), root mass density [mg/cm3] (RMD) (C, D) and specific root length [m/g] (SRL) (E, F) of sole cover crops blue lupin (LU), winter rye (WR), 50:50- and 75:25-cover crop mixtures of lupin and rye (LU_WR_50_50 and LU_WR_75_25) in 2018 (A, C, E) and 2019 (B, D, F) at different soil depth. Colours refer to cover crops: dark green for rye, light green for 50:50-mixture, light blue for 75:25-mixture and dark blue for lupin. Different lowercase letters indicate significant differences (p ≤ 0.05, HSD Tukey-test) between different cover crops within the different soil depth layers (0–10 cm, 10–30 cm, 30–60 cm and 60–100 cm) within one year
Topsoil RMD was high for rye and low for lupin. In 2018, RMD of mixtures were similar to rye, and similar to lupin in 2019. In 2018, lupin had significantly lower RMD than rye and 50:50-mixture in 0–30 cm soil depth. In 2019, RMD of lupin and both mixtures was significantly lower than that of rye in topsoil. No significant differences for RMD below 30 cm soil depth were recorded.
In 2018, mean SRL ranged from 93 m/g in lupin to 543 m/g in rye, and in 2019 from 139 m/g in lupin to 793 m/g in 50:50-mixture. SRL in 2018 was significantly higher for rye than for lupin in 0–30 cm soil depth, the mixture was intermediate. In 2019, SRL of lupin was significantly lower than rye and both mixtures within 0–10 cm.
Below-ground mixture effects
In 2018, relative mixture effects (RME) for all three root traits were positive in 0–60 cm, except SRL in 30–60 cm (-0.13%). Positive RME for RLD was significantly different from 0 in 10–30 cm (Fig. 6). In 60–100 cm, RME for RLD and RMD were negative, whereas RME for SRL was positive. Across the total profile, RME of all three root traits were positive.

Mean values ± standard error (n = 3) of relative mixture effect [%] for root length density (RLD), root mass density (RMD) and specific root length (SRL) for 50:50 and 75:25 cover crop mixtures of lupin and rye (50_50 and 75_25) in 2018 (A) and 2019 (B) at different soil depth and for the total soil depth profile. Colours refer to cover crop mixtures: light green for 50:50-mixture and light blue for 75:25-mixture. Asterisks indicate a significant difference in RME from 0 (p ≤ 0.05, t-test)
In 2019, for the 50:50-mixture RME for RLD varied around 0, while RME for RMD were negative, and significantly lower than 0 in 0–10 cm and 60–100 cm. RME for RLD and RMD in the 75:25 were negative, except in 60–100 cm. Both mixtures showed positive RME for SRL in all depth layers.
In the three mixtures RME of SRL was consistently positive and was significant for 0–10 cm across both mixtures and both years (t-test, df = 81, p < 0.0001). Positive RME of RLD and of SRL were observed without trade-off with each other or RLD and SRL itself (Table 2 Supplementary Material).
Results based on the FTIR spectroscopy (Fig. 7) show that the share of rye root mass in the mixture was always significantly higher than of lupin in topsoil and share of root mass in mixture of lupin was significantly higher in subsoil in two out of three cases. In 2018 RMD of rye in topsoil (20–30 cm) was significantly higher than expected (Fig. 8). In all other cases there were no significant differences between observed and expected RMD of rye and lupin.

Mean values (n = 3) of share of root mass [%] of blue lupin and winter rye (A, C, E) and mean values ± standard deviation (n = 3) of root mass density [mg/cm3] (RMD) at different soil depth of blue lupin and winter rye (B, D, F) in a 50:50 cover crop mixtures of lupin and rye in 2018 (A, B), in a 50:50 cover crop mixtures of lupin and rye in 2019 (C, D) and for a 75:25 cover crop mixtures of lupin and rye in 2019 (E, F). Colours refer to species: green for rye, blue for lupin. Different uppercase letters indicate significant differences (p ≤ 0.05, t-test) between share of root mass of blue lupin and winter rye within one soil depth layer

Mean values ± standard deviation (n = 3) of observed (black) and expected (grey) root RMD of rye (A, C, E) and lupin (B, D, F) in a 50:50 cover crop mixtures of lupin and rye in 2018 (A, B), in a 50:50 cover crop mixtures of lupin and rye in 2019 (B, C) and in a 75:25 cover crop mixtures of lupin and rye in 2019 (E, F). Different lowercase letters indicate significant differences (p ≤ 0.05, t-test) between observed and expected RMD within one soil depth layer
The calculation of the diversity effects gave the following results (Table 3): For the shoot DM and shoot nutrient uptake NE was clearly positive (75:25 in 2019) to slightly negative (shoot DM 50:50 in 2019 and N uptake in 2018). CE was always positive, whereas the SE was negative for shoot DM and N uptake and positive for P and K uptake. Significant positive diversity effects for shoot DM and shoot nutrient uptake were only found for the 75:25-mixture in 2019.
For the root DM NE and CE were positive in 2018 and negative in 2019. SE behaved the other way around. Significant diversity effects for root DM were negative NE and CE for the 50:50-mixture in 2019.
Discussion
Shoot biomass, shoot nutrient uptake and soil mineral nitrogen depletion
In the mixtures, our field experiment revealed lower shoot proportion and shoot nutrient uptake for lupin than expected from sole crops, while for rye these values were higher than expected from sole crops (Fig. 2; Table 2). Thus, rye had competitive advantage over the suppressed lupin. However, this interaction was examined three months after seeding while it may be useful to examine early competitive interactions on a single plant level as well, which may be recommended for further studies to understand competitiveness more in detail. Our field experiment confirmed results of other studies, showing the cereal to be dominant in cereal-legume intercrops (Hauggaard-Nielsen et al. 2001a; Yu et al. 2016). Complementarity of N resource use is discussed to be the driver of these intercrops (Corre-Hellou et al. 2006; Jensen 1996). This is shown in our study by a positive complementarity effect (CE) for shoot DM and shoot nutrient uptake (Table 3). Also, Elhakeem et al. (2019) reported positive CE for cover crop shoot mass and N uptake.
The best performing mixture and only one with significant overyielding for N- and P-uptake was the 75:25-mixture (Table 2). Likewise, the positive CE for shoot DM, N and P uptake were only significant in this mixture (Table 3). Moreover, for the shoot mass the mean LER of the 50:50-mixture was only slightly above 1 in both experimental years, whereas mean LER of the 75:25-mixture was 1.22 (Fig. 3). For N and P shoot uptake, LER and RME were only significant in the 75:25-mixture. The 75:25-mixture performed best as the proportion of the dominant partner was reduced for the benefit of the suppressed partner (Yu et al. 2016). Thus, our hypothesis (1) is confirmed only for the 75:25-mixture with more balanced partners. In the 50:50-mixture the biomass gain of rye in the mixture was countervailed by the biomass loss of lupin. However, in all mixtures LER was higher for shoot nutrient uptake than for shoot biomass. Thus, the hypothesis was also confirmed for shoot nutrient accumulation. With a view on nutrient retention in cover cropping LER of nutrient uptake might be more important than LER of biomass itself.
As shown in other studies with mixtures of legumes and non-legumes (Hauggaard-Nielsen et al. 2001a, 2003), in our field experiments cover crops significantly reduced soil inorganic N compared to the control; and soil inorganic N was significantly reduced by mixtures compared to the legume sole crop (Fig. 4). Furthermore, mixtures were as effective as the non-legume sole crop in Nmin-reduction. This confirms the well-known advantage of non-legumes in cover crop mixtures with legumes to reduce the risk of nitrate leaching (Tosti et al. 2014; Tribouillois et al. 2016). We see here a link to increased SRL in mixtures, as high N retention was possible in mixtures although root mass was lower than in non-legume sole crop in 2019. Moreover, N retention and N supply to the subsequent crop present a trade-off which is minimized in mixtures compared to sole crops (Couëdel et al. 2018; White et al. 2017).
Root mass: Below-ground complementarity and vertical root niche differentiation
Topsoil root dry mass and RMD were dominated by rye compared to lupin in both years and mixtures (Fig. 2; Fig. 7 B, D, F). This could be expected from the relations in the shoot and from what is known from other studies showing competitiveness and dominance of cereals intercropped with legumes (Corre-Hellou et al. 2006; Hauggaard-Nielsen et al. 2001a). Beyond this, there is the question whether vertical niche differentiation of root growth, which may allow complementarity in soil resource use (Mommer et al. 2010), can be found.
FTIR spectroscopy analysis revealed a pattern of root mass allocation of cropping partners over soil depth within mixtures with higher RMD of rye in topsoil and higher RMD of lupin in subsoil (Fig. 7 A, C, E). Thus, our hypothesis (3) is confirmed. We assume that this results from the combination of the allorhizous lupin and the homorhizious rye. However, to examine vertical root niche differentiation we suggest not comparing root mass proportions of mixture partners, but rather comparing RMD of mixture partners in sole and intercrops. Thus, when the rooting profiles of sole and intercrops were contrasted, a clear pattern of vertical root niche differentiation was not found, as in all three mixtures observed and expected RMD was very similar, except for rye in topsoil in 2018 (Fig. 8). Thus, our hypothesis (4) cannot generally be confirmed. Hence, in our experiment the entire RMD profile of a cover crop was not changed when introducing this cover crop into a bispecific mixture.
Similar results were found by Mommer et al. (2010). For four grassland species they observed root biomass overyielding but no niche differentiation. Below-ground overyielding in their study mainly occurred in topsoil layers due to enhanced root growth of one species. Likewise, Heuermann et al. (2019) found rather low niche differentiation by comparing species-specific root mass distribution over depth compared for sole and intercrops. In their experiment they found root biomass overyielding in most cover crop mixtures, but dominating cover crops maintained the same niche as in sole crops, whereas suppressed cover crops were outcompeted for their niche.
Niche differentiation in our experiment might be not pronounced sufficiently to be clearly detected, because the growing time of two and a half month might have been too short for the differentiation to build up. This is similarly discussed for the grassland species of Mommer et al. (2010). Also, Heuermann et al. (2019) examined roots after a short growth period after sowing. In a meta-analysis Cardinale et al. (2007) showed that complementarity increases through time. Besides the impact of time, there is the question whether there is a need for vertical root niche differentiation. Possibly, cropping partners differentiate root growth in vertical niches not before strong competition occurs. Thus, due to the use of different N sources by legume and cereal, competition for N was reduced. The only indication for vertical root differentiation was found for RMD of rye in the topsoil in 2018, while no vertical root differentiation was seen in the second year. The mineral N content in 2019 at the time of sowing (Table 1) was more than three times higher than in 2018. Moreover, the soil moisture content, at least at a nearby weather station (Fig. 1), was much higher in 2019 than in 2018. Thus, growth conditions for cover crops were much better in the second years. We assume, at least in 2019, only moderate competition for water and N, so that vertical root niche differentiation was not a necessary strategy.
Alternatively to analysing rooting patterns for vertical niche differentiation, complementarity can be analysed on a more general level by the concept of Loreau and Hector (2001). For mixtures of grassland species, complementarity effects for root biomass were found (Mommer et al. 2010). In our study, indication of complementarity in root growth was found only in 2018 but not for the two mixtures in 2019, where CE for root mass was even negative (Table 3). These results reflect those for vertical root niche differentiation and may be explained by the soil conditions differing between years. Thus, the better aboveground performance of the 75:25 mixture in 2019 as compared to the 50:50 mixtures cannot be explained by root mass complementarity. Other explanatory approaches for the overyielding of N- and P-uptake are aboveground complementarity (Table 3) and belowground facilitative interactions regarding N or other nutrients (Hauggaard-Nielsen and Jensen 2005; Li et al. 2014). Enhanced N and P concentrations in intercropped compared to sole cropped rye in 2019 could be a hint for this (data not shown). In faba bean-maize relay intercrop it was shown that overyielding was higher when roots intermingled compared to separating species roots (Li et al. 1999). Thus, even if we could not find root mass complementarity, belowground effects are likely to be meaningful.
Contrary to the widespread view of complementary root growth and vertical root niche differentiation, we could not find a clear evidence of these concepts in our mixtures of rye and lupin. Thus, it may not be sufficient to measure vertical root niche differentiation or complementarity effects of root growth for root mass. Instead, we may need to run the calculations for root length, as it may have a different dynamic than root mass (Ryser 2006). Thus, techniques are needed to discriminate species-specific root length.
Relative mixture effects of root traits: RMD, RLD and SRL
We did not clearly observe overyielding of root biomass or RMD in our study. Only in 2018 we found higher than expected root mass and a positive RME of RMD (Fig. 6). The LER for root mass, which was only above 1 in 2018, reflects these results (Fig. 3). This contrasts with other studies, where overyielding was found in cover crop mixtures (Heuermann et al. 2019) and in intercrops of winter wheat and winter faba bean (Streit et al. 2019). Unlike in our experiment Streit et al. (2019) examined winter crops, having a longer growth period to build up overyielding. The difference to the study of Heuermann et al. (2019) may be due to strong differences in the methodology and the absolute amount of root mass. In our understanding, the high root mass and root-shoot-ratios of Heuermann et al. (2019) need to be interpreted with caution.
Focusing on root length instead of root mass does not change the picture: RME of RLD was positive only in 2018 and transgressive overyielding of RLD was found in none of the mixtures (Fig. 5 and 6). More complete exploration of total soil volume by RLD of mixtures could thus not be confirmed. Hence, we cannot generally accept our hypothesis (2). In other mixture experiments with main crops, higher RLD in mixtures compared to sole crops were found in faba bean-oil crop mixtures (Schröder and Köpke 2012) and in maize-wheat intercrops (Li et al. 2006). The growing conditions of the cover crops in our study might be hardly comparable to those of the main crops in these studies, especially when relay intercropping is used (Li et al. 2006). Moreover, due to the extreme different RLD of sole crops, rye was not fully able to compensate the low RLD of lupin. Besides, the question remains whether it is necessary for the intercropped rye to invest in more RLD when rye and lupin can access complementary N resources.
Contrary to our expectations, in our study better aboveground crop performance could neither be related to increased RLD nor to increased root mass or vertical niche differentiation in mixtures. Rather, it was higher SRL which may have increased the efficiency of the rooting system by same or higher nutrient uptake with less root mass, as positive RME of SRL was found in both mixtures and years (Fig. 6). A positive mixture effect of SRL may be desirable to improve nutrient uptake efficiency of cover crop mixtures. Similar to our study, Bukovsky-Reyes et al. (2019) examined mixtures and sole crops of hairy vetch and cereal rye, and reported for vetches higher SRL in mixture than in sole crop. Hill et al. (2006) showed that SRL increased with nutrient deficiency, especially for P. Due to interspecific competition for nutrients mixture partners may invest more in acquisitive root traits (Bukovsky-Reyes et al. 2019) such as SRL to cope with competition. If SRL increases, as much as or even more nutrients can be taken up with less investment of assimilates in roots. Hence, ideally higher RLD for better soil exploitation and simultaneously increased SRL for improved nutrient acquisition is found in mixtures. For synergistic interactions in mixtures, it is important that trade-offs between positive mixture effects of different root traits are minimized. In our study, positive mixture effects of RLD and SRL were possible without trade-off with RME of another root trait or those root traits themselves (Table 2 Supplementary Material).
Conclusion
Cover crop mixtures of lupin and rye showed positive aboveground mixture effects only for nutrient uptake in the 75:25-mixture of lupin and rye. This mixture performed best as the proportion of mixture partners in shoot and root mass was balanced. For the mixtures, FTIR spectroscopy revealed that rye had the highest proportion of root biomass in topsoil and that lupin had the highest proportion of root biomass in subsoil. However, a vertical differentiation in RMD between sole and intercrops was not found. Complementarity of root mass could not consistently be detected.
Mixture effects did occur on root-related variables, but they were not consistent across all depths or environments, and they could even be negative. Among the root traits measured, however, SRL had consistently positive mixture effects.
Future research may focus on mixture effects on root traits also for other intercrops and in other environments as we have conducted our experiments just on one site. Moreover, to enable optimization of cover cropping systems, research will need to elucidate the mechanisms leading to mixture effects of root traits and understand the conditions when mixture effects on root related variables occur.
FTIR spectroscopy gave valuable insights in the pattern of root mass allocation of mixture partners over soil depth. However, further research would profit tremendously from non-manual species-specific discrimination of root length and SRL to learn more about root growth of intercrops.
Change history
26 April 2022
A Correction to this paper has been published: https://doi.org/10.1007/s11104-022-05442-0
References
Andersen SN, Dresbøll DB, Thorup-Kristensen K (2014) Root interactions between intercropped legumes and non-legumes—a competition study of red clover and red beet at different nitrogen levels. Plant Soil 378:59–72. https://doi.org/10.1007/s11104-013-2014-4
Bedoussac L, Journet E-P, Hauggaard-Nielsen H, Naudin C, Corre-Hellou G, Jensen ES, Prieur L, Justes E (2015) Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron Sustain Dev 35:911–935. https://doi.org/10.1007/s13593-014-0277-7
Bodner G, Leitner D, Kaul H-P (2014) Coarse and fine root plants affect pore size distributions differently. Plant Soil 380:133–151. https://doi.org/10.1007/s11104-014-2079-8
Böhm W (1979) Methods of Studying Root Systems, vol 33. Springer, Berlin
Bukovsky-Reyes S, Isaac ME, Blesh J (2019) Effects of intercropping and soil properties on root functional traits of cover crops. Agr Ecosyst Environ 285:106614. https://doi.org/10.1016/j.agee.2019.106614
Cardinale BJ, Wright JP, Cadotte MW, Carroll IT, Hector A, Srivastava DS, Loreau M, Weis JJ (2007) Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc Natl Acad Sci 104:18123–18128. https://doi.org/10.1073/pnas.0709069104
Corre-Hellou G, Fustec J, Crozat Y (2006) Interspecific Competition for Soil N and its Interaction with N2 Fixation, Leaf Expansion and Crop Growth in Pea-Barley Intercrops. Plant Soil 282:195–208. https://doi.org/10.1007/s11104-005-5777-4
Couëdel A, Alletto L, Tribouillois H, Justes É (2018) Cover crop crucifer-legume mixtures provide effective nitrate catch crop and nitrogen green manure ecosystem services. Agr Ecosyst Environ 254:50–59. https://doi.org/10.1016/j.agee.2017.11.017
Elhakeem A, van der Werf W, Ajal J, Lucà D, Claus S, Vico RA, Bastiaans L (2019) Cover crop mixtures result in a positive net biodiversity effect irrespective of seeding configuration. Agr Ecosyst Environ 285:106627. https://doi.org/10.1016/j.agee.2019.106627
Hartwig NL, Ammon HU (2002) Cover crops and living mulches. Weed Sci 50:688–699. https://doi.org/10.1614/0043-1745(2002)050[0688:AIACCA]2.0.CO;2
Hauggaard-Nielsen H, Ambus P, Jensen ES (2001a) Interspecific competition, N use and interference with weeds in pea–barley intercropping. Field Crop Res 70:101–109. https://doi.org/10.1016/S0378-4290(01)00126-5
Hauggaard-Nielsen H, Ambus P, Jensen ES (2001b) Temporal and spatial distribution of roots and competition for nitrogen in pea-barley intercrops – a field study employing 32P technique. Plant Soil 236:63–74. https://doi.org/10.1023/A:1011909414400
Hauggaard-Nielsen H, Ambus P, Jensen ES (2003) The comparison of nitrogen use and leaching in sole cropped versus intercropped pea and barley. Nutr Cycl Agroecosyst 65:289–300. https://doi.org/10.1023/A:1022612528161
Hauggaard-Nielsen H, Jensen ES (2005) Facilitative root interactions in intercrops. Plant Soil 274:237–250. https://doi.org/10.1007/s11104-004-1305-1
Hauggaard-Nielsen H, Jørnsgaard B, Kinane J, Jensen ES (2008) Grain legume–cereal intercropping: The practical application of diversity, competition and facilitation in arable and organic cropping systems. Renew Agric Food Syst 23:3–12. https://doi.org/10.1017/S1742170507002025
Heuermann D, Gentsch N, Boy J, Schweneker D, Feuerstein U, Groß J, Bauer B, Guggenberger G, von Wirén N (2019) Interspecific competition among catch crops modifies vertical root biomass distribution and nitrate scavenging in soils. Sci Rep 9:11531. https://doi.org/10.1038/s41598-019-48060-0
Hill JO, Simpson RJ, Moore AD, Chapman DF (2006) Morphology and response of roots of pasture species to phosphorus and nitrogen nutrition. Plant Soil 286:7–19. https://doi.org/10.1007/s11104-006-0014-3
Jensen ES (1996) Grain yield, symbiotic N2 fixation and interspecific competition for inorganic N in pea-barley intercrops. Plant Soil 182:25–38. https://doi.org/10.1007/BF00010992
Jian J, Du X, Reiter MS, Stewart RD (2020) A meta-analysis of global cropland soil carbon changes due to cover cropping. Soil Biol Biochem 143:107735. https://doi.org/10.1016/j.soilbio.2020.107735
Justes E, Bedoussac L, Dordas C, Frak E, Louarn G, Boudsocq S, Journet E-P, Lithourgidis A, Pankou C, Zhang C, Carlsson G, Jensen ES, Watson C, Li L (2021) The 4 C approach as a way to understand species interactions determining intercropping productivity. Front Agric Sci Eng. https://doi.org/10.15302/J-FASE-2021414
Kaye JP, Quemada M (2017) Using cover crops to mitigate and adapt to climate change. A review. Agron Sustain Dev 37. https://doi.org/10.1007/s13593-016-0410-x
Kemper R, Bublitz TA, Müller P, Kautz T, Döring TF, Athmann M (2020) Vertical Root Distribution of Different Cover Crops Determined with the Profile Wall Method. Agriculture 10:503. https://doi.org/10.3390/agriculture10110503
Kozak M, Piepho H-P (2018) What’s normal anyway? Residual plots are more telling than significance tests when checking ANOVA assumptions. J Agron Crop Sci 204:86–98. https://doi.org/10.1111/jac.12220
Li L, Sun J, Zhang F, Guo T, Bao X, Smith FA, Smith SE (2006) Root distribution and interactions between intercropped species. Oecologia 147:280–290. https://doi.org/10.1007/s00442-005-0256-4
Li L, Tilman D, Lambers H, Zhang F-S (2014) Plant diversity and overyielding: insights from belowground facilitation of intercropping in agriculture. New Phytol 203:63–69. https://doi.org/10.1111/nph.12778
Li L, Yang S, Li X, Zhang F, Christie P (1999) Interspecific complementary and competitive interactions between intercropped maize and faba bean. Plant Soil 212:105–114. https://doi.org/10.1023/A:1004656205144
Livesley SJ, Stacey CL, Gregory PJ, Buresh RJ (1998) Sieve size effects on root length and biomass measurements of maize (Zea mays) and Grevillea robusta. Plant Soil 207:183–193. https://doi.org/10.1023/A:1026461107110
Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity experiments. Nature 412:72–76. https://doi.org/10.1038/35083573
Mead R, Willey RW (1980) The Concept of a ‘Land Equivalent Ratio’ and Advantages in Yields from Intercropping. Exp Agric 16:217–228. https://doi.org/10.1017/S0014479700010978
Mendiburu F de (2019) Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3–0. Available online at: https://CRAN.R-project.org/package=agricolae. (Accessed Jan 26, 2021)
Mommer L, van Ruijven J, de Caluwe H, Smit-Tiekstra AE, Wagemaker CAM, Joop Ouborg N, Bögemann GM, van der Weerden GM, Berendse F, de Kroon H (2010) Unveiling below-ground species abundance in a biodiversity experiment: a test of vertical niche differentiation among grassland species. J Ecol 98:1117–1127. https://doi.org/10.1111/j.1365-2745.2010.01702.x
Mulder CP, Uliassi DD, Doak DF (2001) Physical stress and diversity-productivity relationships: the role of positive interactions. Proc Natl Acad Sci 98:6704–6708. https://doi.org/10.1073/pnas.111055298
Poeplau C, Don A, Schneider F (2021) Roots are key to increasing the mean residence time of organic carbon entering temperate agricultural soils. Glob Change Biol 27:4921–4934. https://doi.org/10.1111/gcb.15787
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/. Accessed 19 May 2021
Ramirez-Garcia J, Martens HJ, Quemada M, Thorup-Kristensen K (2015) Intercropping effect on root growth and nitrogen uptake at different nitrogen levels. J Plant Ecol 8:380–389. https://doi.org/10.1093/jpe/rtu024
Ryser P (2006) The mysterious root length. Plant Soil 286:1–6. https://doi.org/10.1007/s11104-006-9096-1
Schröder D, Köpke U (2012) Faba bean (Vicia faba L.) intercropped with oil crops – a strategy to enhance rooting density and to optimize nitrogen use and grain production? Field Crop Res 135:74–81. https://doi.org/10.1016/j.fcr.2012.07.007
Streit J, Meinen C, Nelson WCD, Siebrecht-Schöll DJ, Rauber R (2019) Above- and belowground biomass in a mixed cropping system with eight novel winter faba bean genotypes and winter wheat using FTIR spectroscopy for root species discrimination. Plant Soil 436:141–158. https://doi.org/10.1007/s11104-018-03904-y
Thorup-Kristensen K (2001) Are differences in root growth of nitrogen catch crops important for their ability to reduce soil nitrate-N content, and how can this be measured? Plant Soil 230:185–195. https://doi.org/10.1023/A:1010306425468
Thorup-Kristensen K, Magid J, Jensen LS (2003) Catch crops and green manures as biological tools in nitrogen management in temperate zones. Adv Agron 79:227–302. https://doi.org/10.1016/S0065-2113(02)79005-6
Tilman D, Reich PB, Knops J, Wedin D, Mielke T, Lehman C (2001) Diversity and productivity in a long-term grassland experiment. Science (New York, N.Y.) 294:843–845. https://doi.org/10.1126/science.1060391
Tosti G, Benincasa P, Farneselli M, Pace R, Tei F, Guiducci M, Thorup-Kristensen K (2012) Green manuring effect of pure and mixed barley – hairy vetch winter cover crops on maize and processing tomato N nutrition. Eur J Agron 43:136–146. https://doi.org/10.1016/j.eja.2012.06.004
Tosti G, Benincasa P, Farneselli M, Tei F, Guiducci M (2014) Barley–hairy vetch mixture as cover crop for green manuring and the mitigation of N leaching risk. Eur J Agron 54:34–39. https://doi.org/10.1016/j.eja.2013.11.012
Tosti G, Thorup-Kristensen K (2010) Using coloured roots to study root interaction and competition in intercropped legumes and non-legumes. Plant Soil 3:191–199. https://doi.org/10.1093/jpe/rtq014
Tribouillois H, Cohan J-P, Justes E (2016) Cover crop mixtures including legume produce ecosystem services of nitrate capture and green manuring: assessment combining experimentation and modelling. Plant Soil 401:347–364. https://doi.org/10.1007/s11104-015-2734-8
Wendling M, Büchi L, Amossé C, Jeangros B, Walter A, Charles R (2017) Specific interactions leading to transgressive overyielding in cover crop mixtures. Agr Ecosyst Environ 241:88–99. https://doi.org/10.1016/j.agee.2017.03.003
White CM, DuPont ST, Hautau M, Hartman D, Finney DM, Bradley B, LaChance JC, Kaye JP (2017) Managing the trade off between nitrogen supply and retention with cover crop mixtures. Agr Ecosyst Environ 237:121–133. https://doi.org/10.1016/j.agee.2016.12.016
Yu Y, Stomph T-J, Makowski D, Zhang L, van der Werf W (2016) A meta-analysis of relative crop yields in cereal/legume mixtures suggests options for management. Field Crop Res 198:269–279. https://doi.org/10.1016/j.fcr.2016.08.001
Zhang C, Postma JA, York LM, Lynch JP (2014) Root foraging elicits niche complementarity-dependent yield advantage in the ancient ‘three sisters’ (maize/bean/squash) polyculture. Ann Bot 114:1719–1733. https://doi.org/10.1093/aob/mcu191
Acknowledgements
We thank the technical staff of the ‘Wiesengut’ organic farming research station and our students for help in the field.
Funding
Open Access funding enabled and organized by Projekt DEAL. The project is supported by funds of the Federal Ministry of Food and Agriculture (BMEL) based on a decision of the parliament of the Federal Republic of Germany via the Federal Office for Agriculture and Food (BLE) under the Federal Programme for Ecological Farming and Other Forms of Sustainable Agriculture.
Author information
Authors and Affiliations
Contributions
M.A. conceived the idea, planned the research and designed the experiments. R.K. conducted the experiments in the field, processed the root samples and collected the data. N.L. and C.M. performed the FTIR analysis. R.K. analysed the data and wrote the article. M.A., T.F.D. N.L. and C.M. contributed to data interpretation, writing and editing of the article. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Responsible Editor: Jiayin Pang.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kemper, R., Döring, T.F., Legner, N. et al. Root traits in cover crop mixtures of blue lupin and winter rye. Plant Soil 475, 309–328 (2022). https://doi.org/10.1007/s11104-022-05366-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05366-9