
Abstract
Background
Nutrient cycle in tropical forests is mainly driven by litter fall amounts and by litter decay due to litter physical and chemical properties. Apart from differences in site conditions, different successional stages of tropical rainforests may influence nutrient inputs to the soil via differences in litter fall and decay.
Methods
We studied leaf litter fall, decomposition and related nutrient input to the soil in two secondary tropical rainforest types differing in land-use intensity and history in the lowlands of Sumatra (Indonesia).
Results
Mean annual litter fall was by 29.5% higher in the old than in the young secondary forests. In contrast, annual litter decomposition differed only marginally (54.9% vs. 52.1%) in young vs. old secondary forests. Litter chemical composition (C, N, P, K contents) did not differ between the forest types. Litter decomposition was also not different between the forest types (4.6% vs. 4.3% loss of initial weight per month). Consequently, annual nutrient input to the soil was primarily affected by differences in leaf litter production, thus, old secondary forests exceeded that of young secondary forests by 30% and 24% for C and N, but was not different regaring P and K among the two forest types.
Conclusions
The results indicate that litter nutrient inputs to the soil in the two types of secondary tropical lowland forests are most strongly influenced by aboveground litter production rather than litter chemical quality or litter decay. We conclude that the restoration status of secondary tropical lowland forests is crucial for the nutrient status of these forest ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil organic matter and soil nutrient sources are strongly influenced by decomposing leaf litter fall (beside of root litter inputs), therefore leaf litter material plays a major role in carbon and nutrient cycles of ecosystems (Vitousek 1984; Vitousek and Sanford 1986; Rapp et al. 1999; Lohbeck et al. 2015). Organic matter forms the source of energy for microorganisms and as it decomposes nutrients bound in litter are mineralized (Pfeiffer et al. 2013; Hicks et al. 2018). Plants are main drivers of nutrient input via litter fall and rhizodeposition, and litter quality is an important driver of decomposition processes (De Deyn et al. 2008; Vesterdal et al. 2008; Hertel et al. 2009; Pfeiffer et al. 2013; Lohbeck et al. 2015). This is in accordance to studies on leaf litter decomposition in tropical moist forest worldwide (Vitousek 1984; Burghouts 1993; Kavvadias et al. 2001; Hertel et al. 2009; Xiogai et al. 2013; Kotowska et al. 2015b). Results of these studies suggest that nutrient cycling is crucial for the functioning of tropical forests, but the effect of forest conversion on leaf litter fall and nutrient cycling is not fully understood so far.
Tropical lowland rainforests of South East Asia count among the most threatened ecosystems in the world due to the persistent forest conversion into agricultural production systems such as rubber and oil palm plantations (Wilcove et al. 2013; Krashevska et al. 2015). Indonesia has become the world’s largest palm oil (Koh et al. 2011) and second largest rubber producer during the last decades (Marimin et al. 2014). Based on the National Forest Inventory (NFI) from 1985–1997 22.5 million hectars have been deforested which is equivalent to a deforestation rate of 1.8 million ha/year (Indonesian ministry of forestry 2002). Differences in logging intensity, frequency and time since logging affect the structure of secondary forests. The removal of large trees creates canopy gaps (Tsingalia 2010) and the use of heavy machinery results in compacted topsoils (Hillel 1982; Donahue et al. 1983; Kasel and Bennet 2007 and 2011). Consequently, a logged over forest tends to become dominated by pioneer species which are adapted to early successional conditions with their dominance increasing with the intensity of disturbance (e.g. Rutten et al. 2015). Logged rainforest has been classified according to their successional stage and timber production value according to stem size distribution, total basal area and the successional status of the species present (Vilá et al. 2013; Martin et al. 2013; Swinfield et al. 2016; Bertoncello et al. 2016). However, few studies examined variations in aboveground litter fall, litter decomposition and related nutrient input to the soil depending on the successional stage of tropical secondary lowland forests (e.g. Kotowska et al. 2015b).
Harapan Rainforest Reserve (HRF) in Sumatra, Indonesia, is one of the few remaining tropical lowland rainforests in this part of South-East Asia. A total of 607 species of trees have been recorded in this forest reserve (research data of HRF, 2012). Rembold et al. (2017) found 1382 plant species and morpho-species from 148 vascular plant families, including 724 tree, 379 shrub species, and 258 herb species in the lowland rainforest reserves Harapan rainforest and Bukit 12 National Park (northwest of Harapan forest) emphasizing the high biodiversity status of these two tropical lowland forest reserves on Sumatra. However, the Harapan Rainforest Reserve was and still is under the pressure of logging activities by legal and illegal timber harvesting dating back to the 1970s. The differences in these logging activities has created forest areas that are more of a natural tree species composition and stand structure, while others are more strongly affected by logging.
The aim of our study was to compare leaf litter production and decomposition between rather natural forest sites ('old secondary forest', 'OSF') with those where the structure and species composition have been changed more strongly ('young secondary forest', 'YSF'') in the Harapan forest reserve. We selected six pairs of the respective forest types and investigated leaf litter fall, litter decomposition and the associated nutrient return to the soil.
More explicitly, the aims of this research study were:
-
1)
to quantify the amount of leaf litter fall in 'young' and 'old' secondary forests in the HRF,
-
2)
to analyze the nutrient contents in the leaf litter of the YSF and OSF in the HRF,
-
3)
to determine the nutrient stocks in the soil of both forest types in the HRF,
-
4)
to analyze the correspondence of leaf litter fall production and climate conditions (precipitation and temperature),
-
5)
to investigate the rate of decomposition rate of leaf litter in both forest types In the HRF,
-
6)
to assess the proportion of nutrients in the leaf litter that is transferred to the soil through litter decomposition.
We wanted to test the hypotheses (i) that leaf litter production differs between the two types of secondary lowland rainforest types, (ii) that litter chemical properties differ between the forest types due to differences in species composition, and thus (iii) that element return via leaf litter production and litter decay to the soil differs between the two lowland secondary forest types.
Material and methods
Study site description and plot selection
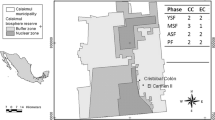
The research was conducted in the Harapan Rainforest Reserve (HRF), district of Batanghari, Jambi province, Sumatra, Indonesia (Figs. 1 and 2). The forest plots grow on rather flat area and are characterized by high plant biodiversity. Mean annual rainfall is 2643 mm and mean monthly temperature is 28°C (Table 1). The soil type is predominantly medium nutrient-rich Oxisol (Allen et al. 2016; Drescher et al. 2016; for more soil chemical data see Table 1).

Map showing the location of the Harapan forest reserve in the Jambi province on Sumatra, Indonesia

View of one of the young (left) and the old (right) secondary forest sites, respectively, in the Harapan tropical lowland rainforest reserve in Sumatra (Indonesia). Photo credits: D. Hertel
For each secondary forest type ('old' vs. 'young'), six pairs of replicated plots were established in spring 2014 within a proximity of 300 m up to seven km that guarantee statistical independence under similar site conditions regarding soil chemical properties, and climate conditions in this climatical and geological homogenous tropical lowland area. At each site, a pair of 'young' secondary forest, i.e. apparent logging activities during the last 5 years characterized by a basal area (BA) less than 6 m2 ha−1 with > 30% of the BA comprising pioneer tree species, and 'old secondary forest', i.e. only slightly logged over of the largest canopy trees in the past 10 years characterized by BA > 12 (up to 30) m2 ha−1 with only < 10% of the BA comprising pioneer tree species (for details, see Rembold et al. 2017), were selected.
Study plots soil chemical properties
The soil samples were taken at 0–5 cm, 5–30 cm, 30–60 cm and 60–80 cm soil depth, respectively. The soil samples were analyzed for carbon, nitrogen, plant-available phosphorus, and exchangeable cations (CEC). Soil and litter samples were analyzed in Jambi at Andalas University’s laboratory. Total N was measured by using the Kjeldahl method, i.e. 5 g soil was digested in H2SO4, using catalyst mixture (CuSO4, Na2SO4 and selenium powder) and distillation. Total C was determined by 5 g soil was digested in K2Cr2O7 and H2SO4 (Black 1965). Available P was measured by using the Bray method, i.e. 2.5 g soil was digested Bray and Kurt extraction (0.025 N HCL + NH4F 0.03 N liquid), (Bray and Kurtz 1945). Total P was measured by wet combustion/dectruction method, i.e. 2.5 g litter sample were digested in 2.5 ml H2SO4 and determined by Spectrophotometer (Jones 1984). Total K was analyzed by using flame photometry, HCL 25% as a reagent (Jones 1984). Exchangeable cation concentrations were determined by atomic absorption spectrophotometer after soil percolation with NH4-Ac (Chapman 1965).
Measurement of aboveground litter production
In every plot, five places were selected randomly where the soil samples for chemical analyses were taken, litter traps (60 cm × 60 cm size) were placed and litter bags for investigating seasonal leaf litter decay were placed (see below). The leaf litter was collected every month between May 2014 and April 2015. Litter fall samples were collected and separated into different litter components, containing leaf litter, and dry weight was analyzed by drying the litter material at 65°C to constant weight for 48 h. Only leaf litter was used for the further analyses in our study. The leaf litter samples were then grinded and analyzed for element contents.
Measurement of litter decomposition and element input to the soil via leaf litter production and decomposition
For the measurement of leaf litter decomposition, the leaf litter was air dried and put into litter bags (20 cm × 10 cm, mesh size for the bottom side 1 mm and upper side 2.5 mm) with 10 g litter per bag. The litter bags were put in the field at the same location as the litter trap sites. In total, 300 litter bags were installed in the field at six locations in old secondary forest and six locations in young secondary forest. At each location five litter bags replications were installed to allow for five dates of resampling the bags; i.e. zero time (starting conditions), and after 3, 6, 9, and after 12 months of field incubation. To avoid the samples being lost by erosion or other events, the litter bags were pinned to the soil using a nail 7 inch in every corner of the litterbag to the ground. After recollecting the litter bags from the field we analyzed the remaining dry weight to determine the mass loss and rate decomposition and then to estimate the nutrient input to the soil.
Element input to the soil via leaf litter fall and decomposition
Annual input of chemical elements (C, N, P, K) to the soil via leaf litter fall and decomposition was calculated based on the annual leaf litter fall, leaf litter decomposition, and the chemical composition of the leaf litter.
Statistical analyses
An analyses of variance (ANOVA) was conducted in form of one-way procedures with post-hoc Tuckey HSD test were used to determine the different variables. All analyses were conducted using SPSS software (version 16) with statistical significance at p < 0.5% level. Additionally, a Principal Componant Analysis (PCA) was conducted in order to detect differences between the six young (YSF) and old secondary (OSF) lowland forests, respectively, in Jambi Province, Sumatra, regarding annual leaf litter fall (Lif), leaf litter chemical composition regarding leaf litter C, N, and P concentration (LlC, LlN, LlP), element return via leaf litter fall (LlfC, LlfN, LlfP), annual leaf litter decomposition rate (percentage of remaining dry mass; Lld), annual element return to the soil of C, N, P via leaf litter fall and decomposition (LlfdC, LlfdN, LlfdP), and soil C, N, and P concentration (0–5 cm soil depth, SC, SN, SP), respectively. The analysis was conducted using CANOCO software, version 5.02 (Biometris, Wageningen, the Netherlands).
Results
Chemical properties of the soil
The concentration for all chemical elements investigated (C, N, P, Ca, Mg, K) was highest in the top soil except for potassium values, and clearly decreased with increasing soil depth (Table 1). Except for this element, the two secondary forest types showed no significant chemical element concentrations in each soil depth.
Leaf litter production
The quantity of leaf litter produced significantly varied between old and young secondary forests. The highest litter fall was measured in October 2014 and the lowest production was measured in March 2015 which followed the annual rainfall regime (Table 2). Old secondary forests had higher litter fall production than young secondary forests. The monthly litter fall production at young and old secondary forests varied from 29 to 144. and 41 to 178 g m−2 year −1, respectively (Table 2). The total annual litter fall total in old secondary forests (949.8 g m−2 yr−1) was by 29.6% higher than in young secondary forests (733.3 g m−2 yr−1, Table 2 and Fig. 4). The lowest litter fall production occurred in the rainy season (with highest rainfall), while the highest production of litter fall occurred in October, which was the month following after the season of lowest rain fall (Table 2). The temperature in the research area, in contrast, was quite invariate during the study period.
Leaf litter element concentration
The chemical composition of leaf litter in old secondary forest and young secondary forests did not show statistically significant differences except of somewhat higher C concentrations in the old vs. young secondary forests (Table 3).
Decomposition of leaf litter
After three months of incubation of leaf litter in litter bags, the dry weight loss was more than 25% of the initial weight (Table 4). However, there was no significant difference in litter decay between the two forest types, although the difference in litter decomposition between the two forest types seemed to increase with time with higher litter decay in young secondary forests (Table 4).
After one of year, litter decay was found to be somewhat faster in the young secondary forest compared to old secondary forest, but this result was only marginally significant (p < 0.10; Table 4 and Fig. 3). Within 1 year, incubated litter of old and young secondary forest decomposed more than half of the initial litter dry weight i.e. 54.9% in the young secondary forest vs. 52.1% in the old secondary forest (Fig. 4).

Seasonal course of leaf litter decomposition rate (percentage of remaining dry mass) in the six young secondary (YSF) and old secondary forests (OSF), respectively. No statistical differences were found between the the two forest systems (Tukey’s honest significant difference; p < 0.05)

Annual leaf litter decomposition rate (percentage of remaining dry mass) and annual leaf litter production in the six young secondary (YSF) and old secondary forests (OSF), respectively. Given are means and standard error. Means within a column followed by different letter are significantly different using Tukey’s honest significant difference (p < 0.05)
Element input to the soil via leaf litter fall and decomposition
Element input to the soil via leaf litter decomposition in the presented study showed that the C and N input was significantly higher in the old secondary forests than the young secondary forests (Table 5). In contrast, no differences were found in the annual nutrient input of P and K via litter input and decomposition between old secondary forests and young secondary forests. However, all four elements investigated showed higher input values in the old secondary than in the young secondary forest (Table 5).
Analysis of overall interactions (PCA)
Ordination of the two lowland rainforest types following a principal component analysis (PCA) based on the investigated soil and leaf litter chemical composition, leaf litter fall amount, and litter decomposition patterns showed a clear separation of the forest types. Figure 5 and Table 6. Thus, the first principal component axis was significantly associated by annual litter fall amount, leaf litter C amount, leaf litter C and N flux via litter fall, as well as annual input of C and N via leaf litterfall and decomposition. On the other hand, leaf litter decomposition was strongly positively associated with the second axis and strongly negatively associated with the third axis. Similarly, leaf litter P concentration as well as annual leaf litter fall P and input of leaf litter P to the soil via litter fall and decomposition was strongly associated to the second axis of the PCA. Notably, 4 of 6 YSF plots were associated negativly with the first component axis, while 5 of 6 OSF plots were positively associated with the first component axis indicating a clear differentation of the YSF and OSF plots.

Results from a Principal Component Analysis (PCA) of the differences between the six young (YSF) and old secondary (OSF) lowland forests, respectively, in Jambi Province, Sumatra, regarding annual leaf litter fall (Lif), leaf litter chemical composition regarding leaf litter C, N, and P concentration (LlC, LlN, LlP), element return via leaf litter fall (LlfC, LlfN, LlfP), annual leaf litter decomposition rate (percentage of remaining dry mass; Lld) annual element return to the soil of C, N, P via leaf litter fall and decomposition (LlfdC, LlfdN, LlfdP), and soil C, N, and P contents (0–5 cm soil depth; SC, SN, SP), respectively
Discussion
Supporting our first hypothesis, the results of our study showed that the old secondary forests had a significantly higher total leaf litter production than the young secondary forests in the Harapan forest reserve area. Thus, in line with our expectations, the old secondary forest revealed a higher annual amount of leaf litter fall that, most likely, is due to the higher tree stem density, size of trees and higher leaf area index in the old compared to the young secondary forest stands. With regard to the seasonality of leaf litter fall, the highest leaf litter production was measured in October 2014 for both forest types and the lowest litter fall was measured in March 2015 again for both forest types, independently. It is a common observation that trees in tropical rainforests sites with a more or less seasonal rainfall pattern shed the old leaves after the end of the dry season, thus our observation that the litter fall increased in the dry season in both forest types were in accordance with similar results of other similar studies (e.g. Vitousek 1984; Kotowska et al. 2015b; Lohbeck et al. 2015; N’Dri et al. 2018).
The chemical element concentration of the leaf litter material was found to be somewhat but not statistically significantly higher in young secondary forests than in old secondary forests (regarding N, P and K concentration), but this was statistically not significant. This trend might be due to the fact that fast growing pioneer tree species have found to show higher leaf nutrient concentrations compared to slower growing late successional species (e.g. Dusenge et al. 2015). However the leaf litter chemical properties were similar compared to other studies in tropical lowland forests (e.g. Kotowska et al 2015b).
After one year of litter decomposition in the field, the young secondary forests had an only marginally higher decomposition rate than the old secondary forests: overall, high and young secondary forests lost more than half of original litter material via litter decay. The leaf litter decomposition process has been found to be mainly driven by differences in climate and soil conditions among sites (e.g. temperature, soil fertility, and bulk density) as well as by the leaf litter chemical composition (Swift et al. 1979; Tripathi et al. 2006; Rapp et al.1999, van Straaten et al. 2014, Hicks et al. 2018). Differences in leaf litter decomposition rates of tropical tree species have been reported in several studies (Swift et al. 1979; Aranguren et al. 1982; Vitousek 1984; Hedge 1995; Rapp et al.1999; Tripathi et al. 2006; Wang et al. 2008; Hayashi et al. 2012; Lohbeck et al. 2015; Kotowska et al. 2015b; Hicks et al. 2018). Beside differences in site factors, it has been observed that the initial lignin/N ratio of the leaf litter material is decisive for controlling the earlier phases of the litter decomposition process (Harmon et al. 1990; Coûteaux et al. 1995; Ribeiro et al. 2002; Yang et al. 2004 and Tripathi et al. 2006; Zang et al. 2018). Unfortunately, litter lignin concentrations were not been determined in our study. However, since the forest sites in the study by Meriem et al. (2016) and in our study were the same, we can use this information for an assessment of the importance of the lignin/N ratio for our decomposition rates. Therefore, we assume that the slightly higher carbon concentration in the leaf litter material of the OSF compared to the YSF may indicate a higher lignin/N ratio in the litter material of the OSF that may at least partly explain the trend of a lower decomposition rate in the old secondary forest. Furthermore, the difference of a lower basal area and thus lower canopy closure of the YSF compared to the OSF should both have led to an increase in solar radiation to the floor and therefore an increase in the temperature at the ground. Higher soil temperature is known to enhance the decomposition process (Vitousek and Sanford 1986; Kavvadias et al. 2001; Steffan-Dewenter et al. 2007; Lebrija-Trejos et al. 2010; Kotowska 2015b; Krishna and Mohan 2017). Thus, the similar results in litter decay in the young and the old secondary tropical lowland forests in our study may be mainly a result of similar soil nutrient conditions, particularly in the top soil of our study sites, as well as small differences in the leaf litter chemical composition, but could be also due to the difference in the thermal regime at the forest floor that might counterbalance the above mentioned differences i.e. in chemical leaf litter composition.
The soil fertility is often influenced by the activities of microorganisms, decomposers and enzymes (Coûteaux et al. 1995; Raubuch and Beese 1995; Rejmánková and Sirová, 2007; Agegnehu and Amede 2017). The nutrient stock of the soil and the litter fall production has an important impact on the nutrient cycling in the forest ecosystems. Nutrient concentration of the soil can influence the decomposition process of litter fall (Swift et al. 1979; Tripathi et al. 2006; Hobbie 2015; Meriem et al. 2016; Hicks et al. 2018) and vice versa litter fall also influence the nutrient input to soil via decomposition (Wang et al. 2008).
Nutrient and C flux to the soil via litter fall and litter decomposition in the present study was significantly higher in old secondary forest compared to the young secondary forest. The nutrient input to the soil is generally influenced by the litter fall amount, the litter chemical composition, and the litter decay rate (e.g. Krishna and Mohan 2017; Kavvadias et al. 2001). Annual C, N, P and K return to the soil through litter production and decomposition was by 30.1, 24.1 percent higher in the old secondary forests compared to the young secondary forest plots, but not significantly higher for P and K between the tow forest types, respectively. The main factor, however, for the higher nutrient inputs to the soil via litter fall and decomposition in this study was the differences in total leaf litter fall between the two forest types rather than differences in leaf litter nutrient concentrations or the leaf litter decomposition rates. This has been similarly been found by Kavvadias et al. 2001. They found that difference in nutrient concentration of litter fall and nutrient input to the soil between two sites in undisturbed natural forest, Greece (high site quality and low site quality) were mainly due to the different litter fall amounts.
The total soil organic carbon in the top soil (0–5 cm) of old secondary forests was higher than that of young secondary forests (5.78% vs. 4.19%, respectively), while the organic carbon tended to decrease with soil depth in both forest types. It is a general phenomenon that soil organic carbon is always accumulated mainly in the top soil (Dieter, et al. 2010) as a result of soil carbon input via leaf litter fall as well as via root litter input as fine root biomass has been shown to strongly decrease with soil depth as well (Hertel et al. 2009; Violita et al. 2015; Angst et al. 2015, 2018; Kotowska et al. 2015a; Heinze et al. 2018). The higher concentration of organic carbon in the topsoil of the old secondary forests compared to the young secondary forest is therefore most likely caused by the higher litter fall production in this system. The same is true for the differences in nitrogen concentrations in the topsoil of the two different secondary forest types. Soil N in the uppermost soil was significantly higher in old secondary forests than in young secondary forests, and as the organic carbon concentration, decreased with soil depth in both forest types. The higher level of N in the topsoil of the old secondary forests is therefore due to the higher level of organic carbon input and storage there. This is in accordance with findings e.g. by Rapp et al. (1999); Mendham et al. (2002); Martínez-Mena et al. (2008); N’Dri et al. (2018); Allen et al. (2018).
Conclusions
The litter decomposition after a year of litter incubation in the field was more than 50% of the initial litter dry mass in both forest types. It had a large contribution to the nutrient input to the soil beside the differences in annual litter fall production between the forest types. In our study the nutrient input to the soil was significantly higher the in old secondary forests than in the young secondary forests. This indicates that old secondary forests in tropical lowland rain forests need to be kept preserved and protected from illegal logging not only from a biodiversity but also from a soil nutrient cycle perspective. Furthermore, the results from our study emphasize that young secondary forests in tropical lowlands need to be restored to reach the status of later successional forests in order to keep these forest ecosystems stable in terms of the ecosystems' soil nutrient status and cycle.
References
Agegnehu G, Amede T (2017) Integrated Soil Fertility and Plant Nutrient Management in Tropical Agro-Ecosystems: A Review. Pedosphere 27:662–680
Allen K, Corre MD, Kurniawan S, Utami SR, Veldkamp E (2016) Spatial variability surpasses land-use change effects on soil biochemical properties of converted lowland landscapes in Sumatra, Indonesia. Geoderma 284:42–50
Allen K, Hassler E, Kurniawan S, Veldkamp E, Core MD (2018) Canopy soil of oil palm plantations emits methane and nitrous oxide. Soil Biol Biochem 122:1–6
Angst G, Kögel-Knabner I, Kirfel K, Hertel D, Mueller CW (2015) Spatial distribution and chemical composition of soil organic matter fractions in rhizosphere and non-rhizosphere soil under European beech (Fagus sylvatica L.). Geoderma 264:179–187
Angst G, Messinger J, Greiner M, Häusler W, Hertel D, Kirfel K, Kögel-Knabner I, Leuschner C, Rethemeyer J, Mueller CW (2018) Soil organic carbon stocks in topsoil and subsoil controlled by parent material, carbon input in the rhizosphere, and microbial-derived compounds. Soil Biol Biochem 122:19–30
Aranguren J, Escalente G, Herrera R (1982) Nitrogen cycle of tropical perennial crops under shade trees: II Cacao. Plant Soil 67:259–269
Bertoncello R, Oliveira RR, Holl KD, Pansonato MP, Martini AM (2016) Cluster planting facilitates survival but not growth in early development of restored tropical forest. Basic Appl Ecol 17:489–496
Black CA (1965) Methods of Soil Analysis, Part 2, Agronomy 9. American Society of Agronomy, Madison, Wisconsin, pp 891–901
Bray RH, Kurtz LT (1945) Determination of total organic and available forms of phosphorus in soils. Soil Sci 59:39–45
Burghouts TBA (1993) Spatial variability of nutrient cycling in Bornean rain forest. Ph.D. Thesis Vrije University, Amsterdam
Chapman HD (1965) Cation exchange capacity. In: Blank CA, Evans DD, White JL, Ensminger LE, Clark FE (eds) Methods of soil analysis, vol II. American Society of Agronomy, Madison, Wisconsin, pp 891–901
Coûteaux MM, Bottner P, Berg B (1995) Litter decomposition, climate and litter quality. Trends Ecol Evol 10:63–66
De Deyn GB, Cornelissen JHC, Bardgett RD (2008) Plants functional traits and soil carbon sequestration in contrasting biomass. Ecological Letters 11:516–531
Dieter D, Elsenbeer H, Turner BL (2010) Phosphorus fractionation in lowland tropical rainforest soils in central Panama. CATENA 82:118–125
Donahue RL, Miller RW, Schicluna JC (1983) Soils; an Introduction to soil and plant growth, Fith. Prenticy-Hall. Inc., Englewood cliffs, New Jersey, p 667
Drescher J, Rembold K, Allen K, Beckschäfer P, Buchori D, Clough YFHFAM, Gunawan D, Herte D, Irawan B, Jaya INS, Klaner B, Kleinn C, Knohl A, Kotowska MM, Krashevska V, Krishna V, Leuschner C, Lorenz W, Meijide A, Melati D, Nomura M, Pérez-Cruzado C, Qaim M, Siregar IZ, Steinebach S, Tjoa A, Tschamtke T, Wick B, Wiegand K, Kreft H, Scheu S (2016) Ecological and socio-economic functions across tropical land use systems after rainforest conversion. Phil Trans R Soc B 371:20150275
Dusenge ME, Wallin G, Gårdesten J (2015) Photosynthetic capacity of tropical montane tree species in relation to leaf nutrients, successional strategy and growth temperature. Oecologia 177:1183–1194
Harmon ME, Baker GA, Spycer G, Greene SE (1990) Leaf litter decomposition in the Picea/Tsuga forest of Olympic National Park, Washington, USA. For Ecol Manage 31:55–66
Hedge R (1995) Nutrient content and decomposition of leaf litter of Accasia mangium Wild as affected by season and Field condition. Msc thesis, Kerala Agricultural University, unpublished, Thrissur
Heinze S, Ludwig B, Piepho HP, Mikutta R, Don A, Wordell-Dietrich P, Helfrich M, Hertel D, Leuschner C, Kirfel K, Kandeler E, Preusser S, Guggenberger B, Leinemann T, Marschner B (2018) Factors controlling the variability of organic matter in the top- and subsoil of a sandy Dystric Cambisol under beech forest. Geoderma 311:37–44
Hertel D, Harteveld MA, Leuschner C (2009) Conversion of a tropical forest into agroforest alters the fine root-related carbon flux to the soil. Soil Biol Biochem 41:481–490
Hicks LC, Meir P, Notthingham AT, Reay DS, Stott AW, Salinas N, Whitaker J (2018) Carbon and nitrogen inputs differentially affect priming of soil organic matter in tropical lowland and montane soils. Soil Biol Biochem 129:212–222
Hillel D (1982) Introduction to soil physics. Academic press, INC, California, p 364
Hobbie SE (2015) Plant species effects on nutrient cycling: revisiting litter feedbacks. Trends Ecol Evol 30:357–363
Jones JB Jr (1984) Laboratory guide of exercises in conducting soil tests and plant analysis. Benton Laboratories INC, Athens. Georgia
Kasel S, Bennet LT (2007) Land use history, forest conversion and soil organic carbon in pine plantation and native forest of south eastern Australia. Geoderma 137:401–413
Kasel S, Sonu S, Gregor JS, Bennet LT (2011) Species-specific effect of nativetrees on soil organic carbon in biodiverse plantings across north-central Victoria, Australia. Geoderma 161:95–106
Kavvadias VA, Alifragis D, Tsiontsis A, Brofas G, Stamatelos G (2001) Litterfall, litter accumulation and litter decomposition rates in four forest ecosystems in northern Greece. For Ecol Manage 144:127–133
Koh LP, Miettinen J, Liew SC, Ghazoul J (2011) Remotely sensed evidence of tropical peat land conversion to oil palm. Proceed Nat Acad Sci USA 108:5127–51232
Kotowska MM, Leuschner C, Meriem TT, S, Hertel D, (2015a) Quantifying above- and belowground biomass carbon loss with forest conversion in tropical lowlands of Sumatra (Indonesia). Glob Change Biol 21:3620–3634
Kotowska MM, Leuschner C, Triadiati HD (2015b) Conversion of tropical lowland forest reduces nutrient return through litterfall, and alters nutrient use efficiency and seasonality of net primary production. Oecologia 180:601–618
Krashevska V, Klarner B, Widyastuti R, Maraun M, Scheu S (2015) Impact of tropical lowland rain forest conversion into rubber and oil palm plantations on soil microbial communities. Biol Fertil Soils 51:697–705
Krishna MP, Mohan M (2017) Litter decompositionin forest ecosystem : A review. Energ Ecol Environ 2:236–249
Lohbeck M, Poorter L, Martinez-Ramos M, Bongers M (2015) Biomass is the main driver of changes in ecosystem process rates during tropical forest succession. Ecol Soc Am 96:1242–1252
Marimin M, Darmawan MA, Islam Fajar Putra MP, MachfundWiguna B (2014) Value chain analysis for green productivity improvement in the natural rubber supply chain: a case study. J Clean Prod 85:201–211
Martin PAA, Newton C, Bullock JM (2013) Carbon pools recover more quickly than plant biodiversity in tropical secondary forests. Proc R Soc B 280:1713
Martínez-Mena M, López J, Almagro M, Boix-Fayos C, Albaladejo J (2008) Effect of water erosion and cultivation on the soil carbon stock in a semiarid area of South- East Spain. 2008. Soil Tillage Res 99:119–129
Meriem S, Tjitrosoedirjo S, Kotowska MM, Hertel D, Tridiati D (2016) Carbon and nitrogen stocks in dead wood of tropical lowland forests as dependent on wood decay stages and land-use intensity. Ann for Res 59:299–310
N’Dri JK, Guéi AM, Edoukou EF, Yéo JGN, Guessan KK, Lagerlöf J (2018) Can litter production and litter decomposition improve soil properties in the rubber plantations of different ages in Coˆte d’Ivoire? Nutrient Cycle Agroforestry Syst 111:203–215
Pfeiffer B, Fender AC, Lasota S, Hertel D, Jungkunst HF, Daniel R (2013) Leaf litter is the main driver for changes in bacterial community structures in the rhizosphere of ash and beech. Pllied Soil Ecol 72:150–160
Rapp M, Regina IS, Rico M, Gallego HA (1999) Biomass, nutrient content, litterfall and nutrient return to the soil in Mediterranean oak forests. Forest Ecol Manag 119:39–49
Raubuch N, Beese F (1995) Pattern of microbial indicator in forest soils along European transect. Biol Fertilty Soils 19:362–368
Rejmánková E, Sirová D (2007) Wetland macrophyte decomposition under nutrient condition : relationships beetwen decomposition rate, enzyme activities and microbial biomass. Soil Biol Biochem 39:525–538
Rembold K, Mangopo H, Tjitrosoedirdjo SS, Kreft H (2017) Plant diversity, forest dependency, and alien plant invasions in tropical agricultural landscapes. Biol Cons 213:234–242
Ribeiro C, Maidera M, Araújo MC (2002) Decomposition and nutrient release from leaf litter of Eucalyptus globules ground under different water and nutrient regimes. For Ecol Manage 171:31–41
Rutten G, Ensslin A, Hemp A, Fischer M (2015) Forest Structure and Composition of previously Selectively Logged and Non-Logged Montane Forest at Mt Kilimanjaro. For Ecol Manage 337:61–66
Steffan-Dewenter I, Kessler M, Barkmann J, Bos MM, Buchori D, Erasmi S, Faust H, Gerold G, Glenk K, Gradstein SR, Guhardja E, Harteveld M, Hertel D, Höhn P, Kappas M, Köhler S, Leuschner C, Maertens M, Marggraf R, Migge-Kleian S, Mogea J, Pitopang R, Schaefer M, Schwarze S, Sporn SG, Steingrebe A, Tjitrosoedirdjo SS, Tjitrisoemito S, Twele A, Weber R, Woltmann L, Zeller M, Tscharntke T (2007) Tradeoffs between income, biodiversity, and ecosystem functioning during tropical rainforest conversion and agroforestry intensification. Proceed Nat Acad Sci, USA (PNAS) 104:4973–4978
Swift MJ, Heal OW, Anderson JM (1979) Decomposition in Terrestrial ecosystem. Black Well Scientific Publications, Oxford
Swinfield T, Afriandi R, Antony F, Harrison RD (2016) Accelerating tropical forest restoration through the selective removal of pioneer species. For Ecol Manage 381:209–216
Tripathi SK, Sumida A, Shibata H, Ono K, Uemura S, Kodana Y, Hara T (2006) Leaf litter and decomposition of different above-and bellow-ground part of birch (Betulaermanil) trees and dwarf bamboo (sasa kurilensis) shrub in young secondary forest in Northern Japan. Biol Fertil Soil 43:237–246
Tsingalia M (2010) Impact of Selective Logging on Population Structure and Dynamics of Canopy Tree (Olea capensis) in Kakamega Forest. Afr J Ecol 48:569–575
Van Straaten O, Corre MD, Wolf K, Tchienkoua M, Cuellar E, Matthews RB, Veldkamp E (2014) Conversion of lowland tropical forests to tree cash crop plantations loses up to one-half of stored soil organic carbon. Proceed Nat Acad Sci, USA (PNAS) 12:9956–9960
Vasterdal L, Schmidt IK, Callessen I, Nilsson LO, Gundersen P (2008) Carbon and nitrogen in forest floor mineral soil under six common European tree species. For Ecol Manage 255:35–48
Vilá MA, Carrillo-Gavila´ N, J, Vayreda, Bugmann H, Fridman J, Grodzki W, Haase J, Kunstler G, Schelhaas M, Trasobares A (2013) Disentangling biodiversity and climatic determinants of wood production. Proceeding of the National Academy of Science, USA (PNAS):e53530
Violita Kotowska MM, Hertel D, Triadiati Miftahuddin, Anas I (2015) Transformation of lowland rainforest into oil palm plantations results in change of leaf litter production and decomposition in Sumatra, Indonesia. Int J Biodivers Environ Sci 6:546–556
Vitousek PM (1984) Litterfall, Nutrient Cycling, and Nuntrient Limitation in Tropical Forests. Ecol Soc Am Ecol 65:285–298
Vitousek PM, Sanford RL (1986) Nutrient cycling in Moist Tropical Forest. Annual Rev Ecol Syst 17:137–167
Wang Q, Wang S, Yu H (2008) Comparison of litter fall, litter decomposition and nutrient return in a monoculture Cunning hamialanceolata and a mixed stand in Southern China. For Ecol Manage 255:1210–1218
Wilcove DS, Giam X, Edwards DP, Fisher B, Koh LP (2013) Navjot’s night mare revisited : logging, agriculture, and biodiversity in Southeast Asia. Trends Ecol Evol 28:531–540
Xiogai G, Zeng L, Xiao W, Zhilin H, Geng X, Tan B (2013) Effect of litter substrate quality and soil nutrients on forest litter decomposition: A review. Acta Ecol Sin 33:102–108
Yang Y-S, Chen G-S, Lin P, Xie J-S, Guo J-F (2004) Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in subtropical China. Annals Forest Sci 61:617–627
Zang T, Luo Y, Chen HYH, Ruan H (2018) Responses of litter decomposition and nutrient release to N addition: A meta-analysis of terrestrial ecosystems. Appl Soil Ecol 128:35–42
Acknowledgements
We are grateful to the DFG (German Research Foundation) for granting this research within the collaborative research center CRC 990 ("efforts"). We thank Dr. Tom Swinfield and Prof. Rhett D. Harrison for the supervision of this research as well as our research assistants who helped us to conduct this research. We further thank PT REKI (HRF) for giving us the research permit for our research activities. We further want to thank the two anonymous reviewers that helped to significantly improve the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL. The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by the authors. The draft of the manuscript was written by all authors and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Declarations
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Elizabeth M Baggs.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Alvafritz, L., Hertel, D. Impacts of land use history on leaf litter input, chemical composition, decomposition and related nutrient cycling in young and old secondary tropical lowland rainforests (Sumatra, Indonesia). Plant Soil 495, 359–370 (2024). https://doi.org/10.1007/s11104-023-06330-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-023-06330-x