
Abstract
Urbanisation dramatically alters ecosystems, disrupting key processes and threatening species persistence. Despite growing knowledge of the impacts of cities on wildlife, particularly for birds and mammals, there remains a paucity of research on how urbanisation impacts other taxa, including reptiles. We examined how urbanisation affects skink communities across multiple New Zealand cities by undertaking pitfall trapping surveys in a range of habitats. Of the 20 endemic species thought to have been present historically in the cities, only four were captured during surveys, Oligosoma aeneum in Hamilton; O. polychroma, O. aeneum, and O. ornatum in Wellington; O. polychroma in Nelson; and O. aff. polychroma Clade 5 in Dunedin. Our results reveal highly variable patterns of occupancy, abundance, and complex associations with key threats and habitat modifications. Sites in Nelson and Wellington were much more likely to record skinks than Hamilton and Dunedin, and modelling showed abundance of O. polychroma was positively related to rat tracking rates when grass cover was low but showed a negative relationship when grass cover was high. O. polychroma body condition was negatively associated with the proportion of urban land cover (built up areas or transport infrastructure). The low number of captures of several species is concerning and long-term monitoring is needed to assess the trajectories of these vulnerable populations. Effective conservation management of urban wildlife requires knowledge of where populations exist and how urban processes affect persistence. For understudied urban-dwelling taxa such as reptiles, the lack of this information may be putting populations at risk.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The effects of human activities on ecosystems are wide-reaching, long-lasting and seldom more intense than in cities where the majority of the world’s population now lives (McKinney 2002; United Nations 2018). Landscape-scale changes inherent to urbanisation dramatically decrease vegetated land cover, substantially reducing the availability of resources for wildlife (Valiela and Martinetto 2007; Grimm et al. 2008; Dupras and Alam 2015). Additionally, wildlife faces ongoing challenges of anthropogenic disturbance (e.g., traffic, development, etc.), and exotic competitors and predators (McKinney 2006). These urban processes tend to have strong negative effects on native biodiversity (McKinney 2006), and in the United States, urbanisation has been identified as one of the leading causes of species extinction (Czech et al. 2000).
As a result, over the last 20 years there have been numerous calls for conservation in cities (Miller and Hobbs 2002; Dunn et al. 2006), both to reduce negative impacts on the often significant biodiversity that can exist in cities (Ives et al. 2016; Soanes and Lentini 2019) and to ensure that humans continue to experience nature and interact with native species in their daily lives (Miller 2005; Dearborn and Kark 2009). However, conservation in cities requires knowledge of what species are present, how populations are faring and how the urban environment impacts different species (French et al. 2018). Research into urban wildlife has typically been biased towards mammals and birds, with reptiles, amphibians, fish, and arthropods being under-represented (Magle et al. 2012). This is concerning as responses to urbanisation differ considerably among taxa (McKinney 2008).
Globally, reptile populations are in decline and one contributing factor is land cover change resulting from urbanisation (Cox et al. 2022). The few studies examining reptiles in cities illustrate the highly variable nature of their responses to urbanisation – likely due to the highly variable nature of cities, with differing degrees of disturbance and high heterogeneity, particularly at moderate levels of urbanisation (French et al. 2018). In Tucson, Arizona, while reptile abundance and diversity generally decreased across a rural to urban gradient, it peaked at intermediate residential housing density (Germaine and Wakeling 2001). Similar patterns have been observed in other taxa, possibly resulting from the wide range of land uses that take place in the transition zone between a city’s centre and outskirts (McKinney 2006, 2008). It is also inevitable, however, that some ‘urban adaptor’ species are better adapted than others to cope with the challenges of urban life (McKinney 2002; Kark et al. 2007). For example, the eastern blue-tongued skink (Tiliqua scincoides) in suburban Sydney, Australia, has likely been successful in these habitats because it exhibits strong site fidelity, readily makes use of artificial refuges, and has relatively r-selected reproductive traits (i.e., early maturation and large litters; Koenig et al. 2001).
In New Zealand, where ecosystems evolved in the absence of nearly all terrestrial mammals, threats to native reptiles in cities may differ from those in most other parts of the world. Like other modified landscapes across the country, New Zealand’s cities retain only a fraction of the species diversity present in their regions before human colonisation (Woolley et al. 2019). Although the declines of now extirpated species were not documented, patterns of species loss in cities appear similar to those across much of the New Zealand mainland and likely resulted from similar threats: anthropogenic habitat loss and the introduction of mammalian predators (Tingley et al. 2013; Woolley et al. 2019). While the most vulnerable species are probably already lost from cities, of those that remain, seven of the nine skink species and five of the seven gecko species are classified as ‘Declining’, ‘Nationally Vulnerable’, or ‘Relict’ under the New Zealand Threat Classification System (Woolley et al. 2019; Hitchmough et al. 2021).
Despite the challenges of urban living, cities do offer conservation opportunities for lizards (Woolley et al. 2019). Green spaces such as parks and urban bush reserves are a mainstay for biodiversity, including lizard populations (Melzer and Bell 2014; Bell 2019). In addition to reserves, cities also offer valuable habitat in backyards and ‘informal green spaces’, such as the edges of transport infrastructure or urban parks. Although the value of these areas for biodiversity has not been well studied (Rupprecht et al. 2015), some of the only published research about lizards in New Zealand cities comes from these habitats (Barwick 1959; van Heezik and Ludwig 2012; Bell et al. 2018). In general, however, there has been little effort invested in the study of urban lizard populations (Woolley et al. 2019). Few biodiversity surveys for lizards have taken place in cities (but see Melzer and Bell 2014; Bell 2019), and much of the knowledge of species occurrence is anecdotal, often from salvage and biosecurity operations. As a result, there is little information about where lizard populations exist in cities, how populations are faring, and whether species might be vulnerable. Here we illustrate the importance of this information for urban conservation management using New Zealand skinks as a case study. We conducted skink surveys in four New Zealand cities to assess the state of their populations in an urban context. Specifically, we aimed: (1) to compare the current species diversity in New Zealand cities with that thought to be present historically, (2) to compare skink abundance among cities and between urban and non-urban habitats, and (3) to explore relationships between population parameters and variation in the urban environment.
Methods
Study sites
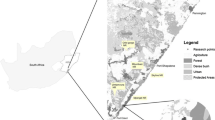
Twelve study sites were selected in each of four New Zealand cities: Hamilton, Wellington, Nelson and Dunedin (n = 47 in total, as one Dunedin backyard was withdrawn from the study; Fig. 1). To ensure sites were representative of the diversity of habitats available in urban environments, in each city the sites were evenly divided among three strata: bush reserve, amenity area and backyard. Within strata, site characteristics differed widely, however, strata did have internal similarities and defining features. Bush reserves comprised areas of remnant or replanted native woody vegetation that were often actively managed and valued as ‘green spaces’ (Fig. 2a). Amenity areas were less densely vegetated and usually comprised more grassland or scrubland, compared with taller vegetation in reserves. Amenity areas were often located at the margins of sports fields or transport infrastructure (Fig. 2b). Backyards consisted of privately-owned residential gardens, with participants recruited by door-knocking. Backyards were highly variable: lawns and cultivated exotic species were common components, though many had native plantings. The degree of management ranged from untidy (Fig. 2c) to highly maintained (Fig. 2d).


Examples of the three habitat strata from Wellington City. (a) a bush reserve site that receives community planting and weeding and is adjacent to a native forest; (b) an amenity area site of scrub/grassland habitat next to the coast and a major road; (c) an unmanaged backyard with a mixture of exotic and native vegetation; (d) a more highly managed backyard with mainly native plantings. Photos: C. Woolley
Skink surveys
Pitfall trapping was used to sample skinks at each site, as all skink species thought to be present in the cities are ground-active (rather than arboreal) and thus able to be captured in ground-based traps. In bush reserves and amenity areas, trapping grids consisted of 25 traps in a 5 × 5 arrangement with 2 m spacing between traps as recommended by other studies (Lettink et al. 2011; Wilson et al. 2017). In backyards, grids of either 6 or 10 traps were used, depending on space available. These grids were installed in 2 × 3 or 2 × 5 arrangements comparable with the other grids or, where space did not allow, traps were distributed around the garden at distances greater than 2 m.
Traps were 4 L plastic buckets dug into the ground so that their openings were flush with the substrate. A lid was pegged 10–20 mm above each trap to provide shade. Traps had 6–8 holes (approx. 4 mm in diameter) drilled in the bottom to allow water drainage, and contained a layer of soil and foliage (approx. 30 mm deep) to provide cover, a moistened sponge (40 mm × 60 mm × 10 mm) to prevent dehydration, and a square of wire mesh (100 mm2 with ~ 10 mm mesh) curved over the floor of the trap to help prevent predation by rodents (Hare 2012). Traps were baited with canned pear (approx. 2 cm3) and checked daily for 7–10 days. Two trapping sessions (early and late summer, i.e., November – December and February – March) took place in each of 2017-18 and 2018-19 (four sessions in total). Trapping was done in all four sessions in Wellington, but in only three sessions in Hamilton, Nelson and Dunedin (see S1 for details of trapping effort).
Captured animals were identified to species level, weighed and measured (snout-vent length, total length, length of tail regeneration). In Wellington and Nelson, skinks were made individually identifiable by clipping one toe on each foot (natural toe loss was incorporated into combinations). Skinks were not marked in Hamilton and Dunedin as prior knowledge suggested that the number of captures would be insufficient to perform mark-recapture analyses. Tail loss and natural toe loss were recorded, and pregnant females were identified visually by the shape of their abdomen.
Habitat- and landscape-scale covariates
Environmental covariates were collected to characterise the study sites, both at the scale of the local habitat and the wider landscape. A convex densiometer (Model A, Forest Suppliers) was used to measure summer canopy cover at each site by recording the proportion of mirrored squares in which the sky was obscured by foliage. Measurements were taken at heights of 1.2 m at the four cardinal compass points at each corner of the 25-trap grids or both ends of the smaller backyard grids. These measurements from each site were then averaged to calculate a mean canopy cover. Proportion of grass cover within pitfall grids was estimated visually on a 1–5 scale (1 = 0–19%, 2 = 20–39%, 3 = 40–59%, 4 = 60–79%, 5 = 80–100%).
Hourly temperature at the substrate was recorded at each site using iButton data loggers (DS1921G-F5#, Maxim Integrated) covered in 400 mm2 squares of Parafilm (Parafilm M, Bemis Company Inc.) to increase weather resistance. Two data loggers were deployed at each site (n = 94) in November–December 2018 and were collected in January–February 2019, yielding a period of 34 days when all loggers were in the field at the same time (19/12/18–21/1/19). Data loggers were tethered with nylon monofilament to traps at opposite corners of the grid, positioned 200 mm from the trap, and oriented north. They were nestled into soil, vegetation or leaf litter to simulate realistic locations of skinks and to avoid direct sunlight.
Spatial analysis was used to calculate the proportion of urban land cover within a 500 m radius of each site. The Land Cover Database (LCDB v.4.1, https://lris.Scinfo.org.nz/layer/48423-lcdb-v41-land-cover-database-version-41- mainland-new-zealand/, accessed: 12/3/19) was clipped within 500 m radii of each of the site locations using the buffer and clip tools in Arcmap v.10.5.1 (ESRI 2017). The resulting polygons were exported into their own layer and the dissolve tool used to calculate single values for the proportional area of each land cover type. Urban cover consisted of ‘Built-up Area (settlement)’, ‘Urban Parkland/Open Space’ and ‘Transport Infrastructure’.
Pest mammal surveys
Concurrent with lizard surveys, pest mammal surveys took place in each city. In Hamilton, Wellington, and Dunedin, mammal surveys were carried out in spring (late October – early December) 2017 and 2018, and autumn (late May – June) 2018 and 2019. In Nelson, we collected data from the public sites only, in early December 2018.
At each site, a line of ten stations of monitoring equipment was deployed at 50 m spacing. Each station consisted of a plastic footprint tracking tunnel (Black Trakka, NZ) with pre-inked card (Gotchatraps, NZ) and a plastic chew card pre-baited with peanut-butter-flavoured possum dough (traps.co.nz). Tracking tunnels were pegged into the ground and baited at both ends of the tunnel with rabbit paste (Erayz). Chew cards were folded and nailed to a tree or other suitable object approximately 300 mm from the ground and 2–5 m from the tunnel. Both tracking tunnels and chew cards were deployed for 6 nights.
We analysed data from spring (when surveys were conducted in all cities) using a subset of the dataset focussing on three key predators of lizards: rats (Rattus rattus, R. norvegicus), mice (Mus musculus), and European hedgehogs (Erinaceus europaeus). To make the mammal indices more relevant to the immediate area of the trapping grid, only the five closest stations were used in analyses. The maximum distance between any grid of pitfall traps and the furthest mammal station used in analyses was c. 200 m.
As tracking tunnel and chew card indices were correlated for the three mammal types (rats, r = 0.63, p < 0.001; mice, r = 0.35, p < 0.044; and hedgehogs, r = 0.37 p = 0.032), only the tracking tunnel indices were used in analyses as these indices were more sensitive to detections of the targeted species (especially hedgehogs; Balls 2019).
Statistical analyses
Site characteristics (Table 1) were compared using one-way anova, and where significant effects were found, pairwise comparisons were made using Tukey’s HSD post-hoc tests.
For each site, an index of abundance for each skink species was made by calculating catch per unit effort (CPUE; number of skinks captured per 100 trap days) across all sessions.
At sites with grids of 25 traps where O. polychroma was recaptured, we estimated population density during each of three trapping sessions, using spatially explicit capture-recapture (SECR) models (Efford and Fewster 2013). Populations were assumed to be closed for the duration of each session. We fitted spatial detection models to the capture data, combining data from all sites and trapping sessions, by maximising full likelihood with the ‘secr’ package v. 3.2.0 (Efford 2019) in R v. 3.5.1 (R Core Team 2018). SECR uses mark-recapture data to estimate density along with two spatial detection parameters: g0, the probability of capture at a trap located at the centre of an animal’s home range, and σ, the width of the detection function, which was assumed to be half-normal. σ describes the spatial scale over which an animal is detected and can be interpreted as implying a bivariate normal model for movement, where 2.45σ is the 95% home range radius (Wilson et al. 2007; Jennrich and Turner 1969). Skink home range centres were assumed to be distributed according to a Poisson point process, with the density parameter estimated from the capture data. Model selection was used to compare models with different covariates for the capture probability (g0) term. The parameter g0 was either constant (i.e., the null model) or with one of four behavioural responses: b, bk, B, Bk; where b is a permanent behavioural response to capture (either trap-happiness or trap-shyness), bk is a permanent behavioural response to capture in relation to a particular trap (i.e., trap-specific), B is a transient response of trap-happiness or -shyness only if it had been captured on the previous capture occasion, and Bk is a transient, trap-specific response (Efford 2019). For all models, density was modelled as a function of trapping session (a trapping period at a particular site) and σ was fitted as a constant. Models were compared using Akaike’s Information Criterion adjusted for small sample sizes (AICc; Burnham and Anderson 2002). When more than one model was supported (ΔAICc <8), we used model-averaging (Burnham and Anderson 2002).
A body condition index (BCI) was calculated using the ratio of log10-transformed mass to log10-transformed SVL for all non-pregnant individuals of a species (as used in other studies, e.g., Grayson et al. 2014; Batson et al. 2015). Linear mixed effects models were used to assess the effect of different environmental factors on abundance and body condition of the most abundant skink, O. polychroma, in Nelson and Wellington. Before running models, multicollinearity was assessed by testing for correlations among predictors using Spearman’s rank correlation analyses. Where correlations were strong (|rs|>0.6; Hosmer and Lemeshow 2005), the predictor that correlated most strongly with other predictors was dropped from the model. CPUE and BCI were fitted as dependent variables in separate models. CPUE was log10-transformed to meet the assumptions of the model (normally distributed residuals, homogeneity of variance). Canopy cover, grass cover, mammal tracking indices (rat, hedgehog and mouse), substrate temperature and proportion of urban cover within 500 m of the sites were fixed effects, and site nested within city and session were random effects (Table 1). Interactions between grass cover and the three mammal tracking indices were tested but were removed from the model if non-significant. Model goodness-of-fit was determined by calculating marginal R2 (variance explained by fixed effects only) and conditional R2 (variance explained by both fixed and random effects; Nakagawa & Schielzeth 2012). All analyses were carried out using R version 3.5.1 (R Core Team 2018).
Results
Site variation
The study sites varied substantially in their environmental characteristics within and among the cities and habitat types. The average substrate temperature during the 34-day summer period ranged from 24.6℃ at one backyard in Nelson to 12.9℃ in a bush reserve in Dunedin. Temperature varied significantly between the cities (F3,43= 43.35, p < 0.001); temperatures at sites in Dunedin were significantly lower than at all other cities (Tukey HSD, p < 0.001; Fig. 3a), and temperatures at sites in Nelson were warmer than those in Wellington (Tukey HSD, p < 0.001; Fig. 3a). Proportion of canopy cover also differed between cities (F3,43 = 9.833, p < 0.001), with sites in Dunedin having more canopy cover than Nelson and Wellington (Tukey HSD, p < 0.001; Fig. 3b). Higher rat tracking indices were recorded at sites in Hamilton than all other cities (F3,37 = 7.514; Tukey HSD, Dunedin and Nelson p < 0.01, Wellington p < 0.05; Fig. 3e). Hedgehog tracking rates also varied significantly between cities (F3,37 = 11.92), with higher rates detected in Dunedin and Nelson compared with Hamilton and Wellington (Tukey HSD, p < 0.001 and p < 0.05 repectively; Fig. 3f). No differences were detected between cities in terms of proportion of grass cover, proportion of urban land cover or mouse tracking rates (Fig. 3c, d and g).
The proportion of urban land cover did differ among the three habitat types (F2,40 = 6.18, p < 0.01). Sites in residential backyards on average had 76% more urban cover within a 500 m radius compared with bush reserve sites. No other variables differed significantly among site types.

Environmental covariates at study sites in four cities: (a) substrate temperature; (b) percentage canopy cover; (c) grass cover; (d) percentage urban cover; (e) rat tracking index; (f) hedgehog tracking index; (g) mouse tracking index. Symbols indicate site habitat type: circles = amenity, crosses = backyard, triangles = bush reserve. Points are jittered horizontally to improve clarity. Non-identitical letters above boxplots indicate statistically significant differences (p < 0.05)
Skink species diversity and abundance in the four cities
Across the four cities, a total of five species of skinks were captured (four endemic and one introduced), though species diversity and site occupancy varied considerably among the cities with high spatial turnover (Table 2).
Catch per unit effort of the four endemic species varied considerably both by city and by site. The largest CPUEs of any species were for O. polychroma in Nelson and Wellington. In Nelson, mean CPUE across all sessions (± standard error of mean; SEM) was 6.47 ± 2.19 captures per 100 trap days, ranging from 0.17 to 28.00 (Fig. 4a). In Wellington, similarly high CPUEs were recorded at some sites (mean = 3.56 ± 1.2, range = 0.00–15.25). In Dunedin, CPUE of O. aff. polychroma Clade 5 at one coastal site (4.32 captures per 100 trap days) was comparable with those of O. polychroma in Wellington and Nelson (Fig. 4a). However, the species was only caught once at one other Dunedin site during the study (mean = 0.41 ± 0.3). In Wellington, mean CPUE of O. aeneum was 0.92 ± 0.74 captures per 100 trap days ranging from 0.00 to 9.03 and in Hamilton mean CPUE of O. aeneum was 0.12 ± 0.08 (range = 0.00–1.04) (Fig. 4b). O. ornatum was captured only twice in the course of this study and only at one site, a backyard in Wellington (1.04 captures per 100 trap days). Mean CPUE for this species in Wellington was 0.09 ± 0.09.

Captures per 100 trap days (CPUE) for a. O. aff. polychroma Clade 5 in Dunedin, b. O. polychroma in Wellington and Nelson, and c. O. aeneum in Hamilton and Wellington. Points are jittered horizontally for clarity. Box plots shown only where medians and quartiles were not zero. Note different scales of y-axes
We caught 148 individual O. polychroma (180 total captures) across all sites (all in Wellington and Nelson) in February 2018, 68 (80 total captures) in November 2018, and 190 (247 total captures) in February 2019. None of the four SECR models which modelled g0 by the different behavioural responses were substantially better supported than each other, although all had more support than the null model (Table 3). Density estimates calculated by averaging these models ranged from 1.04 (95% CI 0.13–8.33) to 61.55 (33.29–113.82) skinks per 100 m2 (Fig. 5).

Density estimates of O. polychroma at sites with 25-trap grids where more than one animal was captured in (a) Wellington and (b) Nelson. (c) shows estimates for Reserve 2 in Nelson with an extended y-axis to accommodate the high estimate in February 2019. Density estimates are averaged from the four supported spatially explicit capture-recapture (SECR) models (ΔAICc < 3.2; Table 3). All models assumed that the parameter σ was constant across sites. Error bars are 95% confidence intervals
Effect of environmental factors on Oligosoma polychroma
Only O. polychroma was captured in high enough numbers to allow the assessment of environmental factors on its populations. At sites where this species was captured, mouse tracking rate was negatively correlated with grass cover (Spearman’s rs = -0.75, p < 0.001), and substrate temperature (rs = -0.47, p < 0.001) and positively corelated with rat tracking rate (r = 0.34, p = 0.004; S2). As these relationships were moderate to strong, mouse tracking rate was dropped from the models to prevent multicollinearity. All other correlations were either non-significant or had |rs| < 0.3 (S2). In linear mixed effects models, although rat tracking rate and grass cover were both positively related to O. polychroma CPUE, these main effects interacted; rat tracking rate was positively related to O. polychroma CPUE at low proportions of grass cover, and negatively at high (Table 4; Fig. 6a). Proportion of urban cover had a negative relationship with O. polychroma BCI (Table 5; Fig. 6b). No other factors were significant in either model. A summary of CPUE and BCI is available in S3.

(a) Relationship between rat tracking index and catch per unit effort of O. polychroma at sites of high and low grass cover. Points are jittered horizontally for clarity. (b) Body condition index (BCI) of O. polychroma declined with increasing proportion of urban cover in both Nelson and Wellington. Trend lines are fitted to raw data points. Grey shading indicates 95% confidence intervals
Discussion
Diversity and abundance of skinks in New Zealand cities
Of the 20 skink species thought to have been present in the regions of our study cities prior to human colonisation (Woolley et al. 2019), only four (20%) were detected in this study. Furthermore, in only one city (Wellington) was more than one skink species detected. These results are not altogether unexpected but are highly concerning and illustrate the dramatic reduction in biodiversity that has taken place in New Zealand’s urban environments.
The four urban-dwelling skink species share characteristics that may make them more adept at surviving in the altered habitats that cities present. Compared with other New Zealand lizards, they have relatively high reproductive outputs (c. five offspring per year) and fast maturation (less than 2 years to sexual maturity; Cree and Hare 2016; summarised in Table 6). They are also all habitat generalists and exist in a wide variety of environments (van Winkel et al. 2018). Behavioural and life history traits such as these have been identified in other urban-dwelling reptiles and it is possible that these traits allow these species to survive in urban environments where others cannot (Koenig et al. 2001). Notwithstanding these similarities, capture rates and site occupancy of the four species varied considerably and while some may be secure in cities, it is possible that others are vulnerable.
The highest site occupancy and capture rates were of Oligosoma polychroma in both Wellington and Nelson, where their estimated densities ranged widely from a few hundred per hectare (e.g., Reserve 2, Wellington) to several thousand per hectare (e.g., Reserve 2, Nelson; Table 7). In general, these densities were comparable with those of other studies of O. polychroma and the related O. aff. polychroma Clade 5 in a range of habitats with varying predator control regimes (Table 7).
Conversely, the low site occupancy and capture rates of Oligosoma aeneum and O. ornatum are cause for concern. O. aeneum was captured at only three sites in each of Hamilton and Wellington, and generally only in low numbers. Mark-recapture studies of this species are rare, but one study in the early 1970s in suburban Wellington found that O. aeneum populations can reach densities greater than 2000 skinks per hectare in suitable habitat (Bell et al. 2018). We were unable to estimate density for O. aeneum in this study, and capture rates were very low at most sites where they were captured (mean = 2.02 captures per 100 trap days, range = 0.15–9.03; Fig. 4). These results suggest that present urban populations may be smaller than they were 50 years ago. However, long-term monitoring is necessary to ascertain if populations are declining. Recent skink surveys for Wellington City Council recorded the species at fewer sites than expected across the city and described this low site occupancy as a cause for concern (Melzer and Bell 2014; Bell 2019). O. ornatum was captured only in a single backyard in Wellington and was not observed in Hamilton (despite its presence being recorded subsequently in a reserve in Hamilton in February 2020; pers. comm. M. Nelson-Tunley, Waikato Regional Council, 2020). This result too is a cause for concern. Other recent skink surveys of parks and reserves in Wellington have identified very few individuals of this species, and at few sites (Melzer and Bell 2014; Bell 2019). In contrast, the species is frequently captured in pitfall trapping surveys at the predator-exclusion-fenced Zealandia ecosanctuary (Nelson et al. 2016), suggesting that predation by introduced mammals may be an important factor driving these patterns. It is likely that populations of O. ornatum in Hamilton and Wellington are sparse, in low numbers, and declining.
Our surveys failed to detect other species thought to be present in cities (Table 2). In particular, O. zelandicum was not captured at any site in Wellington despite being regularly recorded in surveys in the fenced Zealandia ecosanctuary (Romijn 2013). This species has also not been captured in other recent surveys of Wellington reserves (Melzer and Bell 2014; Bell 2019). It has a Threat Status Classification of ‘Declining’ (Hitchmough et al. 2021).
Impact of urban environments on skink populations
We found no clear evidence for a negative impact of rats or hedgehogs on CPUE of O. polychroma. While many mammalian predators consume lizards (Newman 1994; Gillies and Clout 2003; Jones et al. 2013) and the impact of rodents on some lizard populations has been inferred from patterns of loss on New Zealand’s mainland and population recovery on islands following pest mammal eradication (Towns 1991; Towns and Daugherty 1994; Norbury et al. 2014), evidence for the negative impacts of pest mammals on O. polychroma populations is mixed. As discussed in Wilson et al. (2017; and adapted in Table 7), there is no clear pattern indicating that populations of this species respond numerically to predator control. While locations with predator control have recorded some of the highest densities of O. aff. polychroma Clade 5 (e.g., Eglinton Valley; Table 7), some populations of O. polychroma achieved similarly high densities in the absence of predator control (e.g., Kelburn Cemetery and Pukerua Bay; Table 7).
The effect of rat tracking rate on CPUE was, however, dependent upon the proportion of grass cover, with a positive relationship when grass cover was low and a negative relationship when high (Table 4; Fig. 6a). This result suggests that grass cover may benefit populations of O. polychroma when rat abundance is low but not when it is high. It is possible that, in cities, grassland may act as an ecological trap for skinks, being preferentially selected as habitat but not providing adequate protection from rodent predation (Rotem et al. 2013; Battin 2004). Interactions between vegetation, herbivores and predators can result in complex and sometimes unexpected impacts for lizard populations (Norbury 2001; Hoare et al. 2007; Knox et al. 2012; Norbury et al. 2013). Densities of jewelled geckos (Naultinus gemmeus) in Coprosma spp. shrubland on the Otago Peninsula were four times higher in grazed sites than ungrazed, likely due to the higher activity of rodents at ungrazed sites (Knox et al. 2012).
Body condition of O. polychroma was negatively related to urban land cover (Table 5), in both Wellington and Nelson (Fig. 6b). Possibly, as the proportion of urban land cover increases, patch size of suitable habitat decreases making habitat and food resources more limited. Several studies document a decline in invertebrate abundance and diversity with increasing urbanisation (Niemelä et al. 2002; Ishitani et al. 2003; McKinney 2008). Alternatively, increased levels of disturbance may cause changes in foraging behaviours. Disturbances could be in the form of human activity (e.g., green space maintenance) or increased predator activity in more urban environments (e.g., domestic cats, Felis catus; Woolley and Hartley 2019). For example, several studies report differences in flight initiation distances and sprint speeds between lizards living in modified urban habitats and those in more natural environments (Prosser et al. 2006; Chejanovski et al. 2017).
Conservation of urban lizards – future directions
Urban wildlife research is especially challenging when target species have small home ranges or patchy distributions due to the difficulty of sampling across a highly heterogeneous environment. Additionally, as cities have not typically been considered habitat for threatened species, until recently conservation has not been a strong motivation for urban research (McDonnell and Hahs 2013; Ives et al. 2016; Soanes and Lentini 2019). These factors have no doubt contributed to the underrepresentation of reptiles and several other taxa in urban research globally (Magle et al. 2012). Yet, in order to conserve these taxa in cities, ecological knowledge about their urban populations is imperative. Indeed, the present lack of research into how the urban environment affects these groups may be putting some species at risk.
Our study provides some of this essential information for New Zealand skinks and, by doing so, establishes a platform for their management in cities. The future of some lizard species in these environments is far from secure and it seems likely that population monitoring and conservation action will be required to ensure their longevity.
While the range of species captured in our sampling was generally consistent with existing knowledge of lizards in New Zealand cities, the failure to detect O. ornatum in Hamilton in a reserve where it has subsequently been found, illustrates a key limitation of this study: that detection of some species, particularly those that are secretive, at low densities, or have patchy distributions, requires sustained effort and a range of detection tools. Further surveys in cities are needed to identify the locations of populations of skink species for which records are scarce (in particular, O. ornatum, O. aeneum and O. zelandicum). Additionally, future surveys in cities should target the full range of species that may be present, including geckos and arboreal skinks not surveyed in this study, using the full gamut of lizard sampling tools (Hare 2012a, 2012b, 2012c; Lettink 2012). Due to the cryptic, secretive nature of many of New Zealand’s lizards, it remains possible that species thought to have been lost from the mainland could yet be discovered (as was the case for Hoplodactylus duvaucelii at Maungatautari Mountain Sanctuary; Morgan-Richards et al. 2016 ). For this reason, managers should be open minded in their use of survey techniques.
However, due to the heterogeneity of the urban landscape and costs associated with formal lizard surveys, sampling all potential habitats at the necessary intensity is likely not possible. Backyards, though highly variable in the habitat they offer, often constitute large proportions of urban green space in a city (e.g., 36% in Dunedin; Mathieu et al. 2007). Furthermore, the backyards surveyed in this study were among the most important sites for lizards in Wellington City, supporting high diversity (three species in one backyard), high abundance (highest CPUE for O. aeneum), and the only records of O. ornatum. While more formal surveys of backyards are recommended, these may be supplemented using citizen science. Records of lizard observations by members of the public could provide valuable information about species’ distributions in backyards and other habitats that may allow the targeting of more intensive surveys. Such a citizen science project may also serve to increase public awareness for lizards and threats to them in cities (Ives et al. 2018).
Identifying the presence and location of lizard populations in cities, however, is only the first step towards their management and protection. It is possible that populations of some species are in unnoticed decline. To provide the information needed to identify population trends, as well as to demonstrate the efficacy of strategies proposed to benefit lizard populations (e.g., predator control, including cat management and habitat manipulation; Woolley et al. 2019), there is a need to establish long-term monitoring sites where surveys are repeated at regular intervals. Such long-term studies are rare in the urban context (Magle et al. 2012) but may be critical to ensure the persistence of some species in cities, particularly those that are cryptic and secretive.
Finally, it is possible that some of New Zealand’s urban-dwelling lizards may be well adapted to survive in urban environments, in particular O. polychroma. Internationally, other studies have identified apparently urban-adapted reptiles (Germaine and Wakeling 2001; Koenig et al. 2001) and in New Zealand, endemic birds vary in their resilience to introduced predators depending on their body size and degree of endemicity (Fea et al. 2020). Future research should identify what life history or behavioural traits facilitate the survival of these species in New Zealand cities. Understanding the mechanisms by which some species are able to survive in these highly modified environments will allow these species to be managed through further environmental change (Shine and Fitzgerald 1996; Koenig et al. 2001).
Data availability
Not applicable.
Code availability
Not applicable.
References
Balls CS (2019) Understanding the distribution of introduced mammalian predators in urban environments using monitoring tools and community trapping. Unpublished MSc Thesis. Victoria University of Wellington 132pp
Barwick RE (1959) The life history of the common New Zealand skink Leiolopisma zelandica (Gray, 1843). Trans Royal Soc New Z 86(3):331–380
Batson WG, O’Donnell CFJ, Nelson NJ, Monks JM (2015) Placement period of artificial retreats affects the number and demographic composition but not the body condition of skinks. New Z J Ecol 39:273–279
Battin J (2004) When good animals love bad habitats: ecological traps and the conservation of animal populations. Conserv Biol 18(6):1482–1491
Bell T (2019) Wellington City Council lizard surveys: 2016–2018. Technical report prepared for Wellington City Council by EcoGecko Consultants Limited 40pp
Bell BD, Hare KM, Pledger SA (2018) Lizards in the suburbs: a single-garden study of a small endemic New Zealand skink (Oligosoma aeneum). New Z J Zool 0:1–18
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer-Verlag, New York
Chejanovski ZA, Avilés-Rodríguez KJ, Lapiedra O, Preisser EL, Kolbe JJ (2017) An experimental evaluation of foraging decisions in urban and natural forest populations of Anolis lizards. Urban Ecosyst 20:1011–1018
Cox N, Young BE, Bowles P, Fernandez M, Marin J, Rapacciuolo G, Böhm M, Brooks TM, Hedges SB, Hilton-Taylor C, Hoffman M, Jenkins RKB, Tognelli M, Alexander GJ, Allison A, Ananjeva NB, Auliya M, Avila LJ, Chapple DG, Cisneros-Heredia DF, Cogger HG, Colli GR, de Silva A, Eisemberg CC, Els J, Fong AG, Grant TD, Hitchmough RA, Hoffmann M, Iskander DT, Kidera N, Martins M, Meiri S, Mitchell NJ, Molur S, Nogueria CC, Oritz JC, Penner J, Rhodin AGJ, Rivas G, Rodel MO, Roll U, Sanders KL, Santos-Barrera G, Shea GM, Spawls S, Stuart BL, Tolley KA, Trape JF, Vidal MA, Wagner P, Wallance BP, Xie Y (2022) A global reptile assessment highlights shared conservation needs of tetrapods. Nature 605:285–290
Cree A, Hare KM (2016) Reproduction and life history of New Zealand lizards. In: Chapple DG (ed) New Zealand Lizards. Springer, pp 169–206
Czech B, Krausman PR, Devers PK (2000) Economic associations among causes of species endangerment in the United States. Bioscience 50:593–601
Dearborn DC, Kark S (2009) Motivations for conserving urban biodiversity. Conserv Biol 24:432–440
Dixon KM (2004) Biodiversity along a gradient of modification: plant, invertebrate and reptile diversity in mid-altitude tall tussock (Chionochloa rigida) grasslands, eastern Central Otago, New Zealand. Unpublished PhD thesis, University of Otago, Dunedin, New Zealand. 266pp
Dunn RR, Gavin MC, Sanchez MC, Solomon JN (2006) The pigeon paradox: dependence of global conservation on urban nature. Conserv Biol 20:1814–1816
Dupras J, Alam M (2015) Urban sprawl and ecosystem services: a half century perspective in the Montreal area (Quebec, Canada). J Environ Policy Plann 17:180–200
Efford MG (2019) secr: Spatially explicit capture-recapture models. R package version 4.3.1. https://CRAN.R-project.org/package=secr
Efford MG, Fewster RM (2013) Estimating population size by spatially explicit capture – recapture. Oikos 122:918–928
ESRI 2017. ArcGIS Desktop: Release 10.3.1. Environmental Systems Research Institute, Redlands, California
Fea N, Linklater W, Hartley S (2020) Responses of New Zealand forest birds to management of introduced mammals. Conserv Biol 35:35–49
Freeman AB (1997) Comparative ecology of two Oligosoma skinks in coastal Canterbury: a contrast with Central Otago. New Z J Ecol 21:153–160
French SS, Webb AC, Hudson SB, Virgin EE (2018) Town and country reptiles: a review of reptilian responses to urbanization. Integr Comp Biol 58:948–966
Germaine SS, Wakeling BF (2001) Lizard species distributions and habitat occupation along an urban gradient in Tucson, Arizona, USA. Biol Conserv 97:229–237
Gillies C, Clout M (2003) The prey of domestic cats (Felis catus) in two suburbs of Auckland City, New Zealand. J Zool 259:309–315
Gillies C, Williams D (2013) Using tracking tunnels to monitor rodents and mustelids. Tracking tunnel guide v2.5.2. Department of Conservation, New Zealand, p 14
Grayson KL, Mitchell NJ, Monks JM, Keall SN, Wilson JN, Nelson NJ (2014) Sex ratio bias and extinction risk in an isolated population of tuatara (Sphenodon punctatus). PLoS ONE 9
Grimm NB, Faeth SH, Golubiewski NE, Redman CL, Wu J, Bai X, Briggs JM (2008) Global change and the ecology of cities. Science 319:756–760
Hare KM (2012a) In: Herpetofauna (ed) Pitfall trapping. Inventory and monitoring toolbox: Herpetofauna. Department of Conservation 22pp
Hare KM (2012b) In: Herpetofauna (ed) Funnel trapping. Inventory and monitoring toolbox: Herpetofauna. Department of Conservation 24pp
Hare KM (2012c) In: Herpetofauna (ed) Systematic searches. Inventory and monitoring toolbox: Herpetofauna. Department of Conservation 29pp
Hitchmough R, Barr B, Knox C, Lettink M, Monks JM, Patterson GB, Reardon JT, van Winkel D, Rolfe J, Michel P (2021) Conservation status of New Zealand reptiles, 2021. New Zealand Threat Classification Series
Hoare JM, Adams LK, Bull LS, Towns DR (2007) Attempting to manage complex predator–prey interactions fails to avert imminent extinction of a threatened New Zealand skink population. J Wildl Manage 71:1576–1584
Hosmer DW, Lemeshow S (2005) Introduction to the logistic regression model. Applied Logistic Regression. John Wiley & Sons, Hoboken, NJ, pp 1–30
Ishitani M, Kotze DJ, Niemelä J (2003) Changes in carabid beetle assemblages across an urban-rural gradient in Japan. Ecography 26:481–489
Ives CD et al (2016) Cities are hotspots for threatened species. Glob Ecol Biogeogr 25:117–126
Ives CD, Abson DJ, von Wehrden H, Dorninger C, Klaniecki K, Fischer J (2018) Reconnecting with nature for sustainability. Sustain Sci 13:1389–1397
Jennrich RI, Turner FB (1969) Measurement of non-circular home range. J Theor Biol 22:227–237
Jones C, Norbury G, Bell T (2013) Impacts of introduced european hedgehogs on endemic skinks and weta in tussock grassland. Wildl Res 40:36–44
Kark S, Iwaniuk A, Schalimtzek A, Banker E (2007) Living in the city: can anyone become an “urban exploiter”? J Biogeogr 34:638–651
Knox CD, Cree A, Seddon PJ (2012) Direct and indirect effects of grazing by introduced mammals on a native, arboreal gecko (Naultinus gemmeus). J Herpetology 46:145–152
Koenig J, Shine R, Shea G (2001) The ecology of an australian reptile icon: how do blue tongue lizards (Tiliqua scincoides) survive in suburbia? Wildl Res 28:215–227
Lettink M (2012) Herpetofauna: Artificial retreats. Inventory and monitoring toolbox: Herpetofauna. Department of Conservation 26pp
Lettink M, Cree A (2016) Reproduction and Life History of New Zealand Lizards. pp 169–206 in D. G. Chapple, editor. New Zealand Lizards. Springer
Lettink M, O’Donnell CFJ, Hoare JM (2011) Accuracy and precision of skink counts from artificial retreats. New Z J Ecol 35:236–246
LINZ (2015) Land Cover Database version 4.1, https://lris.scinfo.org.nz/layer/48423-lcdb-v41-land-cover-database-version-41-mainland-new-zealand/, accessed: 12/3/19
Magle SB, Hunt VM, Vernon M, Crooks KR (2012) Urban wildlife research: past, present, and future. Biol Conserv 155:23–32
Mathieu R, Freeman C, Aryal J (2007) Mapping private gardens in urban areas using object-oriented techniques and very high-resolution satellite imagery. Landsc Urban Plann 81:179–192
McDonnell MJ, Hahs AK (2013) The future of urban biodiversity research: moving beyond the “low-hanging fruit. Urban Ecosyst 16:397–409
McKinney ML (2002) Urbanization, Biodiversity and Conservation. Bioscience 52:883–890
McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Conserv 127:247–260
McKinney ML (2008) Effects of urbanisation on species richness: a review of plants and animals. Urban Ecosyst 11:161–176
Melzer S, Bell T (2014) Lizard survey of Wellington City Council administered parks & reserves: final report. Unpublished EcoGecko Consultants Ltd report prepared for the Wellington Council, p 46
Miller JR (2005) Biodiversity conservation and the extinction of experience. Trends in Ecology and Evolution 20(8):430–434
Miller JR, Hobbs RJ (2002) Conservation where people live and work. Conserv Biol 16:330–337
Morgan-Richards M, Hinlo AR, Smuts-Kennedy C, Innes J, Ji W, Barry M, Brunton D, Hitchmough RA (2016) Identification of a rare gecko from North Island New Zealand, and genetic assessment of its probable origin: a novel mainland conservation priority? J Herpetology 50:77
Nakagawa S, Schielzeth H (2012) A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol Evol 4:133–142
Nelson N, Romijn R, Dumont T, Reardon JT, Monks J, Hitchmough RA, Empson R, Briskie J (2016) Lizard conservation in mainland sanctuaries. In: Chapple DG (ed) New Zealand Lizards. Springer, pp 321–339
Newman DG (1994) Effects of a mouse, Mus musculus, eradication programme and habitat change on lizard populations of Mana Island, New Zealand, with special reference to McGregor’s skink, Cyclodina macgregori. New Z J Zool 21:443–456
Niemelä J, Kotze DJ, Venn S, Penev L, Stoyanov I, Spence J, Hartley D, Montes de Oca E (2002) Carabid beetle assemblages (Coleoptera, Carabidae) across urban-rural gradients: an international comparison. Landscape Ecol 17:387–401
Norbury G (2001) Conserving dryland lizards by reducing predator-mediated apparent competition and direct competition with introduced rabbits. J Appl Ecol 38:1350–1361
Norbury G, Byrom A, Pech R, Smith J, Clarke D, Anderson D, Forrester G (2013) Invasive mammals and habitat modification interact to generate unforeseen outcomes for indigenous fauna. Ecol Appl 23:1707–1721
Norbury G, Van Den Munckhof M, Neitzel S, Hutcheon A, Reardon J, Ludwig K (2014) Impacts of invasive house mice on post-release survival of translocated lizards. New Z J Ecol 38(2):322–327
Patterson GB (1985) The ecology and taxonomy of the common skink, Leiolopisma nigriplantare maccanni, in tussock grasslands in Otago. Unpublished PhD thesis, University of Otago, Dunedin, New Zealand. 217pp
Prosser C, Hudson S, Thompson MB (2006) Effects of Urbanization on Behavior, Performance, and morphology of the Garden Skink, Lampropholis guichenoti. J Herpetology 40:151–159
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Romijn RL (2013) Can skinks recover in the presence of mice? Unpublished Honours Thesis. Victoria University of Wellington 57pp
Rotem G, Ziv Y, Giladi I, Bouskila A (2013) Wheat fields as an ecological trap for reptiles in a semiarid agroecosystem. Biol Conserv 167:349–353
Rupprecht CDDD, Byrne JA, Garden JG, Hero JM (2015) Informal urban green space: a trilingual systematic review of its role for biodiversity and trends in the literature. Urban Forestry and Urban Greening 14:883–908
Shine R, Fitzgerald M (1996) Large snakes in a mosaic rural landscape: the ecology of carpet pythons Morelia spilota (Serpentes: pythonidae) in Coastal Eastern Australia. Biol Conserv 76:113–122
Soanes K, Lentini PE (2019) When cities are the last chance for saving species. Front Ecol Environ 17:225–231
Tingley R, Hitchmough RA, Chapple DG (2013) Life-history traits and extrinsic threats determine extinction risk in New Zealand lizards. Biol Conserv 165:62–68
Towns DR (1991) Response of lizard assemblages in the Mercury Islands, New Zealand, to removal of an introduced rodent: the kiore (Rattus exulans). J Royal Soc New Z 21:119–136
Towns DR, Daugherty CH (1994) Patterns of range contractions and extinctions in the New Zealand herpetofauna following human colonisation. New Z J Zool 21:325–339
Towns DR, Elliott GP (1996) Effects of habitat structure on distribution and abundance of lizards at Pukerua Bay, Wellington, New Zealand. New Z J Ecol 20:191–206
United Nations (2018) World Urbanization Prospects: The 2018 Revision, Highlights (ST/ESA/SER.A/420) 103pp
Valiela I, Martinetto P (2007) Changes in bird abundance in eastern North America: urban sprawl and global footprint? Bioscience 57:360–370
van Heezik Y, Ludwig K (2012) Proximity to source populations and untidy gardens predict occurrence of a small lizard in an urban area. Landsc Urban Plann 104:253–259
van Winkel D, Baling M, Hitchmough R (2018) Reptiles and amphibians of New Zealand. Auckland University Press, Auckland, New Zealand
Wilson DJ, Efford MG, Brown SJ, Williamson JF, McElrea GJ (2007) Estimating density of ship rats in New Zealand forests by capture-mark-recapture trapping 31(1): 47–59
Wilson DJ, Mulvey RL, Clarke DA, Reardon JT (2017) Assessing and comparing population densities and indices of skinks under three predator management regimes. New Z J Ecol 41:84–97
Woolley CK, Hartley S (2019) Activity of free-roaming domestic cats in an urban reserve and public perception of pet-related threats to wildlife in New Zealand. Urban Ecosyst 22:1123–1137 Urban Ecosystems
Woolley CK, Hartley S, Hitchmough RA, Innes JG, van Heezik Y, Wilson DJ, Nelson NJ (2019) Reviewing the past, present and potential lizard faunas of New Zealand cities. Landsc Urban Plann 192:103647
Acknowledgements
The authors wish to thank the landowners and managers who permitted us to carry out this work on their property. We wish to acknowledge E Webb, J Burgoyne-Smith, N Stanton, T Peters, JC Agulló Brotons, E. Irwin, CJ Macfarlane, R Shaw, R MacKinlay, N Higgens, J Kim, L Gaiduch, B Florence-Bennett, J Aguilar, C Bannon, B Panzarella, G Tocker, S van Wyk, B Woolley, J Woolley, F Grealish, F Perez, M Harvey, K Miller, L Dutoit, H Ricardo, N Fitzgerald, V Elangovan, and X Chai for their assistance with fieldwork. We thank the Nelson lab group ‘Fixit’, David Chapple and an anonymous reviewer for their helpful comments on earlier versions of this manuscript. We thank the PCaN Mammal Team for providing mammal index data for Hamilton, Wellington, and Dunedin. This research was funded through a Victoria University of Wellington Doctoral Scholarship, the Ministry of Business Innovation and Employment grant: UOWX1601 (People, Cities and Nature), and a Wellington City Council Our Natural Capital Grant. Approval was granted for this research by the Victoria University of Wellington Animal Ethics Committee (AEC 27041) and the Department of Conservation (Wildlife Act approval: 50568-FAU).
Funding
This research was funded through a Victoria University of Wellington Doctoral Scholarship, the Ministry of Business Innovation and Employment grant: UOWX1601 (People, Cities and Nature), and a Wellington City Council Our Natural Capital Grant.
Open Access funding enabled and organized by CAUL and its Member Institutions
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval
Approval was granted for this research by the Victoria University of Wellington Animal Ethics Committee (AEC 27,041) and the Department of Conservation (Wildlife Act approval: 50,568-FAU).
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Woolley, C.K., Hartley, S., Innes, J.G. et al. Conservation of skinks in New Zealand cities. Urban Ecosyst 26, 1493–1508 (2023). https://doi.org/10.1007/s11252-023-01398-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01398-w