
Abstract
Purpose
The aim of the study was to compare the temperature sensitivity of soil respiration rate in two soil horizons of seven types of temperate forests.
Materials and methods
Soil samples were collected in O and A horizons in seven types of temperate forests, each one represented by five independent stands distributed throughout Poland. Soil respiration rates were measured at standard moisture in five temperatures (4 °C, 10 °C, 16 °C, 22 °C and 28 °C), and the first-order Q10 values were calculated for each stand. General linear models (GLM) were fitted for respiration rate and for Q10 values separately using selected soil physical-chemical properties: C:N ratio, dissolved organic carbon (DOC) content and soil pH.
Results and discussion
The soil respiration rate increased with temperature was the highest in O horizon of fresh mixed forest dominated by hornbeam and increased with C:N ratio, DOC content and soil pH (model p < 0.0001). In turn, model for Q10 was not significant meaning none of tested variables affected soil temperature sensitivity (p = 0.2886).
Conclusions
Despite studied forest types exhibit substantial distinctness in many soil properties including respiration rate, they showed similar susceptibility to temperature increase (roughly to climate warming).
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Soil respiration is one of the most important processes of the global carbon cycling. Soil organic matter (SOM) decomposition processes are temperature dependent and may accelerate global climate warming through positive feedback (Kirschbaum 2000; Karhu et al. 2010; You et al. 2019). The temperature sensitivity of soil respiration can be regulated by a range of biotic and abiotic factors (Klimek et al. 2016a, 2020). However, our knowledge in this matter is still far from satisfactory.
Traditionally, temperate forests are known as ecosystems characterized by near-to-zero carbon balance, that is, emitting and absorbing (accumulating) similar amounts of carbon. However, last reports showed that they are an important carbon sink (Jílková et al. 2019; Ma et al. 2020). We can observe increasing interest in temperate forest studying in context of global climate warming (Huang et al. 2020). Temperate forests cover 1038 Mh and the amount of C stored in soils of this biome is estimated on 100 Pg (Lal 2005), nearly twice as much is stored in vegetation. Temperate forests are characterized by high variability vegetation diversity and composition across climatic regions and gradient of soil bedrock properties (Giliam 2016; Dukunde et al. 2019). The basic division of temperate forest is coniferous vs deciduous (mixed) forests; more recalcitrant coniferous needles might play a different role than more labile deciduous leaves (Berg and McClaugherty 2008). Coniferous tree species litter is characterized typically by a higher C:N ratio than deciduous tree species litter (Demoling et al. 2007). However, during decomposition the C:N ratio of litter decrease (Gallardo and Merino 1993), therefore soil C:N ratio cannot serve as a proxy for soil carbon decomposability. Dissolved organic carbon (DOC) is the most labile soil C fraction, the direct precursor of microbial growth and activity (Andreasson et al. 2009), thus driving the microbial response to temperature increase. DOC concentration was found to be higher in deciduous temperate forests compared with coniferous (Klimek et al. 2016b). Coniferous tree species litter reduces soil pH comparing deciduous ones (Chodak et al. 2016) which affect soil microbial community diversity and structure, for example, bacteria to fungi ratio and microbial community physiological capabilities (Chodak et al. 2015). All these soil factors may alter temperature-dependent soil processes (Xu et al. 2019). However, some temperate forest types do not follow simple division on coniferous vs deciduous/mixed forests. For example, soil properties of an acid variant of beech forest are closer to features of coniferous tree species stands, that is, low soil pH, low DOC and elements concentration (Chodak et al. 2016). Therefore, both forest type and soil properties have to be analysed simultaneously when looking for some general trends in temperature sensitivity of soil respiration rate process.
The aim of the study was to compare the temperature sensitivity of soil respiration rate in seven types of temperate forest in a selected region of temperate zone (Poland), which are expected to be continuously changing in terms of vegetation and basic soil properties, that is, C:N ratio, soil pH and DOC concentration. We aimed to identify factors responsible the most for soil respiration rate temperature sensitivity in two soil organic horizons and which forest type soils could be especially susceptible to climate warming. We expect that we will detect at least the difference in soil respiration temperature sensitivity between coniferous and deciduous forest and that soils of coniferous forests will be characterized by a higher soil respiration rate temperature sensitivity due to its specific soil properties. As a quantifier of soil respiration rate temperature sensitivity, the commonly used Q10 coefficients were chosen, describing the how many times soil respiration rate increase with temperature increase with every 10 °C. A value of Q10 coefficient the most commonly reported is 2.0–2.5; however, some studies reported much higher values for some soils, meaning much higher risk for soil carbon depletion rate to atmosphere (Klimek et al. 2020).
2 Materials and methods
2.1 Soils collection and vegetation analysis
Soils were collected in seven types of temperate forest, occurring typically in Poland:
-
Dry pine forest (dry pine)—Cladonio-Pinetum and Vaccinio-Pinetum, with typical poor forest floor with Cladonia lichens
-
Mesic pine forest (mesic pine)—Peucedano-Pinetum and Leucobryo-Pinetum
-
Acidophilous beech forest (acid beech)—Luzulo pilosae-Fagetum and Luzulo luzuloides-Fagetum
-
Fertile beech forest (fertile beech)—Dentario glandulosae-Fagetum
-
Mixed deciduous forests with domination of hornbeam (mixed hornbeam)—Carpinion betuli
-
Mixed deciduous forests with domination of oak (mixed oak)—Potentillo albae-Quercetum
-
Riparian forest with domination of ash (riparian ash)—Ficario-Ulmetum and Fraxino-Alnetum
Each forest type was represented by five independent stands, giving altogether 35 stands.
Mixed soil samples were taken in June 2014 from O and A horizons (FAO 1998) from 100 m2 plots (10 m × 10 m, 5 subsamples). The O horizon was not found on some stands as it is absent in more fertile and humic forest types (most of mixed deciduous and riparian forest stands). Finally, 35 samples of A horizon and 24 samples of O horizon were collected. Directly after collection, the samples were sieved (A horizon through 2 mm mesh, O horizon through 1 cm mesh) to remove visible plant debris and stones and to homogenize the samples, packed into plastic boxes and transported field moist to the laboratory.
2.2 Soil physical-chemical analysis
The dry weights (DW) of the soil samples were determined by drying them at 105 °C for 24 h. Next, the organic matter content (OM) in dry weight was determined as the loss on ignition at 550 °C for 24 h. The water holding capacity (WHC) was measured by a standard gravimetric method after soil soaking for 24 h in net-ended plastic pipes immersed in water (Schlichting and Blume 1966). The organic C and total N were analysed by dry combustion with an elemental analyser (Vario El III, Elementar Analysensysteme GmbH), and the C:N ratios were calculated for each soil sample. Total contents of P were extracted by wet digestion of 0.5 g of soil in 10 ml of concentrated HNO3 and HClO4 (Sigma-Aldrich) mixture (7:1 v/v), and the concentration of P in the digests was measured on flow injection analyser (FIA compact, MLE). The concentration of dissolved organic carbon (DOC) was measured in water extracts obtained from 3 g of soil dry mass equivalent shaken for 1 h at 10:1 water-to-soil ratio at 200 rpm (TOC-VCPN, Shimadzu) (Zhang 2010). The soil pH was measured in air-dried subsamples (2 g) shaken in 1 M KCl solution for 1 h at 200 rpm. The soil texture of mineral soils (A horizon) was determined hydrometrically. Each analysis was performed in three subsamples from every soil sample, and the data were averaged and expressed based on the dry weight of the soil. For more detailed information about studied stands, see article of Chodak et al. (2016).
2.3 Soil respiration rate measurements and Q 10 calculations
Separate soil samples were prepared for each of five temperatures, dependently on organic matter content and expected respiration rate: equivalents of 50 g DW for A horizon of pine forest, equivalents of 25 g DW for A horizon for other forest types and equivalents of 5 g DW for O horizon. The soil samples were placed in glass vials, adjusted to 60% of their maximum water holding capacity (WHC) and acclimated at 4 °C, 10 °C, 16 °C, 22 °C and 28 °C (± 0.5 °C) for 5 days before respiration rate measurements. The samples’ moisture was adjusted daily with deionized water.
The respiration rate was measured by CO2 trapping: each soil sample was placed in an airtight jar with a beaker of 5 ml 0.2 M NaOH. Closed jars were incubated for 4 days for 4 °C and 10 °C and 2 days for 16 °C, 22 °C and 28 °C (the incubation time was recorded to the nearest minute). After incubation jars were opened and 2 ml BaCl2 was added to the NaOH solution; the excess sodium hydroxide was titrated using a digital Jencons burette with 0.1 M HCl (0.01 ml precision) in the presence of phenolphthalein as a colour indicator. Several empty jars (with only NaOH) were placed among the other samples as blanks. The soil respiration rate was expressed as mM CO2 kg SOM−1 24 h−1. The respiration rate of each sample was measured twice (A horizon) or four times (O horizon) for each sample and averaged.
The respiration rate (R) results were fitted into exponential Q10 model defined as:
where R1 > 0 and b > 0. This model is traditionally used to determine the Q10 value, a factor that represents the relative growth of respiration as the temperature increases by 10 °C:
The Q10 calculated with that model is constant for a temperature range, which makes it convenient to further processing. The usefulness of this model is restricted to some intermediate temperature range, with excluding very low (close to 0 °C, water freezing point) and high temperatures (close to 40 °C, peptides/enzymes denaturation).
2.4 Statistical analysis
Differences in soil physicochemical characteristics (OM, WHC, concentrations of C, N, P and C:N ratio, DOC concentrations and pH) between seven forest types and two soil organic horizons were compared with two-way ANOVA, and differences in soil texture (sand, silt and clay content) were analysed with one-way ANOVA. Tukey’s honestly significant difference (HSD) test for multiple comparisons was run if significant differences were found (p < 0.05). Right- or left-skewed data were transformed to fulfil the normality criterion. The Shapiro-Wilk test was used to confirm the normality of the data distribution within groups. Non-significant interactions were removed from a model.
Soil respiration rate results were analysed with general linear model (GLM) where forest type, soil horizon and temperature were categorical factors and C:N ratio, soil pH and DOC concentrations were quantitative factors. The categorical factors were tested for differences in means between groups with Tukey’s test, and interactions were tested also and consider to be significant when p < 0.05. Q10 values were analysed with general linear model (GLM) where forest type and soil horizon were categorical factors, whereas C:N ratio, soil pH and DOC concentrations were quantitative factors. The categorical factors were tested for differences in means between groups with Tukey’s test, and interaction between forest type and soil horizon was tested also and consider to be significant when p < 0.05.
3 Results
With a gradient of water availability, soil texture and element content, soil horizons in temperate forest soils tend to be unified and were found less often. We collected 35 samples of A horizon and 24 samples of O horizon. We found O soil horizon in all pine stands, in eight beech stands, in three mixed hornbeam stands, in two mixed oak stands and in just one riparian ash forest (therefore, no standard deviation was given for that group in Table 1). Soil properties of two soil horizons in seven forest types were variable according to the general characteristics of temperate forests (Table 1). Upper O soil horizon was characterized by a higher organic matter content, water holding capacity and C content than A horizon (p < 0.0004), but with exclusion of mixed oak and riparian ash forests, as indicated by a significant interactions between forest type and soil horizon (Table 1). In turn, N and P content was higher in O horizon than in A horizon in each forest type (p < 0.0001), as the interaction between forest type and soil horizon was not significant (Table 1). Forest soils could be divided on two groups along to N and P content in soil: less affluent (both pine forest and mixed oak forest) and more affluent (both beech forest, mixed hornbeam forest and riparian ash forest) (p < 0.0001) (Table 1).
Both forest type and soil horizon affected soil C:N ratio (both p < 0.0001) as well as their interaction (p < 0.02), meaning that soil C:N ratio was higher in O than in A soil horizon only in some forest types, that is, dry pine, mesic pine and fertile beech (Fig. 1). The difference in DOC concentration between soil horizons was observed (p < 0.0001), but the higher DOC concentration was found only in both pine forests (p < 0.0001) (Fig. 2). Soil pH was the lowest in dry pine and the highest in mixed hornbeam and mixed oak forest (p < 0.0002), whereas there was no difference between soil horizons (p = 0.8031) (Fig. 3). Results of pH measurements were in line with soil texture measurements; sand mineral fraction was the highest in pine forest and the lowest in mixed hornbeam and oak forests (Table 2).

C:N ratio in seven forest types and two soil horizons and interaction between forest type and soil horizon. Central points indicate the sample means, and error bars indicate 95% Tukey honestly significant difference intervals. Two-way ANOVA test results were presented here; p values for each factor and significant differences (if existed) between groups were indicated by small letters (a, b, c)

DOC concentration in soil in seven forest types and two soil horizons and interaction between forest type and soil horizon. Central points indicate the sample means, and error bars indicate 95% Tukey honestly significant difference intervals. Two-way ANOVA test results were presented here; p values for each factor and significant differences (if existed) between groups were indicated by small letters (a, b)

Soil pH in seven forest types and two soil horizons. Central points indicate the sample means, and error bars indicate 95% Tukey honestly significant difference intervals. Two-way ANOVA test results were presented here; p values for each factor and significant differences (if existed) between groups were indicated by small letters (a, b). Non-significant interaction between forest type and soil horizon was removed from a model
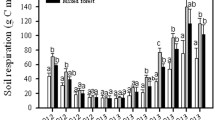
GLM yielded significant model for soil respiration rate (p < 0.0001). The model explained 85.5% in data variability, and all factors tested were significant: forest type (p < 0.0001), soil horizon (p < 0.0001), temperature (p < 0.0001), C:N ratio (p < 0.001), soil pH (p < 0.0005) and DOC concentration (p < 0.0001) as well as interaction between forest type and soil horizon (p < 0.0003) (Fig. 4).

GLM for soil respiration rate. Central points indicate the sample means, and error bars indicate 95% Tukey honestly significant difference intervals. Effects for categorical factors (forest type, soil horizon and temperature), effects for linear factors (C:N ratio, DOC concentration and soil pH) as well as interaction between forest type and soil horizon were presented here; p values for each factor and significant differences (if existed) between groups were indicated by small letters (a, b, c, d, e)
In turn, GLM model for Q10 was not significant (p = 0.2886, R2adj = 0.04), also after subsequent removing non-significant variables. The mean Q10 value for all forest types was 2.31 for soil A horizon and 2.25 for soil O horizon (Table 3).
4 Discussion
Despite substantial differences in soil properties and respiration rate between forest types, they did not differ in Q10 values, that is, temperature sensitivity of soil respiration process. Our short-term incubation experiment was designed to avoid typical harms in such type of experiments related to prolonged soil incubation. Short incubation time prevent for adaptation of soil microbial communities to increased temperatures (Hui et al. 2014) and indirect effects from changes in soil properties, especially depletion in labile carbon (DOC) (Fissore et al. 2013; Liu 2013; You et al. 2019). Short-term soil incubation allow to catch the soil reaction to temperature to being as close as possible to its field response, although it cannot serve to predict ecosystem response in a longer time scale (Smith et al. 2007).
Similar temperature sensitivity of soil respiration in various types of temperate forests is, anyway, an intriguing observation. Studied forest soil represented gradients in soil properties, which resulted in unification of final soil biological activity (respiration rate). For example, both types of pine forest studied, that is, dry pine and mesic pine, were characterized by a relatively high DOC concentration (O soil horizon), which is known to favour for high microbial activity, especially in higher temperatures (Gielen et al. 2011; Liu 2013), but were characterized by a low soil pH, which is not suitable for part of important soil microbial community, namely, bacteria (Rousk et al. 2010). In turn, fertile beech and mixed hornbeam forests soil were characterized by the higher pH but lower DOC concentration than pine forests. DOC is possibly intensively used by microorganisms during respiration rate, as shown in our results for soil respiration rate. Anyway, these opposite effects should be possible to be included into our GLM for Q10 values consisting of all these variables.
Soil respiration rate is regulated by a variety of biotic and abiotic factors (You et al. 2019). Respiration rate is the highest in the range 50–70% of maximal water holding capacity (WHC) of particular soils, which in turn is related mainly to SOM content (Howard and Howard 1993; Ilsteld et al. 2000). In our experiment, soil moisture was set on constant level of 60% of maximal WHC for each of soils, which should allow for achieving the maximal respiration rate. However, moisture may modify the temperature effect on the microbial respiration rate and thus the soil temperature sensitivity (Kechavarzi et al. 2010; Lellei-Kovács et al. 2011). Moreover, the moisture effect on soil respiration rate may be dependent on soil texture as shown for mixed pine forest by Dillustro et al. (2005). Moisture effect may also be stand-dependent; Swallow and Quideau (2013) showed that soil moisture affects soil microbial community structure under aspen stands whereas did not affect spruce stands. In other words, measuring of soil temperature sensitivity in a single soil moisture level is suitable only when studied soils did not differ strongly in their physicochemical properties. Therefore, studies on soil temperature sensitivity of diverse temperate forests should be completed at least by applying different moisture levels and possibly by analysing SOM fractions. The other solutions are studies on common garden experiments, with controlling bedrock properties.
Lack of effect of forest type on Q10 can be, anyway, an optimistic information, meaning that none of temperate forest type is especially susceptible to climate warming through higher soil temperature sensitivity. This could lead (in a long time) to potential disturbances in organic matter cycling in an ecosystem scale and for loss of some environments, being habitats for some specific or endangered species.
Conclusions
Our study highlights the effects of soil physicochemical properties on temperature sensitivity of soil respiration rate. Despite soil respiration rate was strongly differed between soil horizons and forest types and was affected by soil C:N ratio, DOC content and soil pH, we do not observed differences in temperature sensitivity of soil respiration rate. We conclude that other soil properties may affect it, possibly biological not physicochemical soil properties.
References
Andreasson F, Bergkvist B, Bååth E (2009) Bioavailability of DOC in leachates, soil matrix solutions and soil water extracts from beech forest floors. Soil Biol Biochem 41:1652–1658
Berg B, McClaugherty C (2008) Plant litter. Decomposition, humus formation. Springer-Verlag, Berlin Heidelberg Carbon Sequestration
Chodak M, Klimek B, Azarbad H, Jaźwa M (2015) Functional diversity of soil microbial communities under Scots pine, Norway spruce, silver birch and mixed boreal forests. Pedobiologia 58:81–88
Chodak M, Klimek B, Niklińska M (2016) Composition and activity of soil microbial communities in different types of temperate forests. Biol Fertil Soils 52:1093–1104
Demoling F, Figueroa D, Bååth E (2007) Comparison of factors limiting bacterial growth in different soils. Soil Biol Biochem 39:2485–2495
Dillustro JJ, Collins B, Duncan L, Crawford C (2005) Moisture and soil texture effects on soil CO2 efflux components in southeastern mixed pine forests. For Ecol Manag 204:85–95
Dukunde A, Schneider D, Schmidt M, Veldkamp E, Daniel R (2019) Tree species shape soil bacterial community structure and function in temperate deciduous forests. Front Microbiol 10:15–19
FAO (1998) World reference base for soil resources. Appendix 1. FAO, ISRIC and ISSS, Rome, Italy
Fissore C, Giardina CP, Kolka RK (2013) Reduced substrate supply limits the temperature response of soil organic carbon decomposition. Soil Biol Biochem 67:306–311
Gallardo A, Merino J (1993) Leaf decomposition in two Mediterranean ecosystems of southwest Spain: influence of substrate quality. Ecology 74:152–161
Gielen B, Neirynck J, Luyssaert S, Janssens IA (2011) The importance of dissolved organic carbon fluxes for the carbon balance of a temperate Scots pine forest. Agric For Meteorol 151:270–278
Giliam FS (2016) Forest ecosystems of temperate climatic regions: from ancient use to climate change. New Phytol 212:871–887
Howard DM, Howard PJA (1993) Relationships between CO2 evolution, moisture content and temperature for a range of soil types. Soil Biol Biochem 25:1537–1547
Huang L, Zhou M, Lv J, Chen K (2020) Trends in global research in forest carbon sequestration: a bibliometric analysis. J Clean Prod 252:119908
Hui W, Guenet B, Vicca S, Nunan N, Abdelgawad H, Pouteau V, Shen W, Janssens IA (2014) Thermal acclimation of organic matter decomposition in an artificial forest soil is related to shifts in microbial community structure. Soil Biol Biochem 71:1–12
Ilsteld U, Nordgren A, Malmer M (2000) Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer. Soil Biol Biochem 32:1591–1599
Jílková V, Jandová K, Sim A, Thornton B, Paterson E (2019) Soil organic matter decomposition and carbon sequestration in temperate coniferous forest soils affected by soluble and insoluble spruce needle fractions. Soil Biol Biochem 138:107595
Karhu K, Fritze H, Tuomi M, Vanhala P, Spetz P, Kitunen V, Liski J (2010) Temperature sensitivity of organic matter decomposition in tow boreal forest soil profiles. Soil Biol Biochem 42:72–82
Kechavarzi C, Dawson Q, Bartlett M, Leeds-Harrison PB (2010) The role of soil moisture, temperature and nutrient amendment on CO2 efflux from agricultural peat soil microcosms. Geoderma 154:203–210
Kirschbaum MUF (2000) Will changes in soil organic carbon act as a positive or negative feedback on global warming? Biogeochemistry 48:21–51
Klimek B, Jelonkiewicz Ł, Niklińska M (2016a) Drivers of temperature sensitivity of decomposition of soil organic matter along a mountain altitudinal gradient in the Western Carpathians. Ecol Res 31:609–615
Klimek B, Chodak M, Jaźwa M, Solak A, Tarasek A, Niklińska M (2016b) The relationship between soil bacteria substrate utilization patterns and the vegetation structure in temperate forests. Eur J For2 Res 135:179–189
Klimek B, Chodak M, Jaźwa M, Azarbad H, Niklińska M (2020) Soil physicochemical and microbial drivers of boreal forest soils temperature sensitivity. Pedosphere 30:528–534
Lal R (2005) Forest soils and carbon sequestration. For Ecol Manag 220:242–258
Lellei-Kovács E, Kovács-Láng E, Botta-Dukát Z, Kalapos T, Emmett B, Beieret C (2011) Thresholds and interactive effects of soil moisture on the temperature response of soil respiration. Eur J Soil Biol 47:247–255
Liu H (2013) Thermal response of soil microbial respiration is positively associated with labile carbon content and soil microbial activity. Geoderma 193–194:275–281
Ma J, Xiao X, Li R, Zhao B, Myint SW (2020) Enhanced spring phenological temperature sensitivity explains the extension of carbon uptake period in temperate forest protected areas. For Ecol Manag 455:117679
Rousk J, Bååth E, Brookes PC, Lauber CL, Lozupone C, Caporaso JG, Knight R, Fierer N (2010) Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J 4:1340–1351
Schlichting E, Blume HO (1966) Bodenkundliches Praktikum. Paul Parey, Hamburg
Smith JL, Bell JM, Bolton H, Bailey VL (2007) The initial rate of C substrate utilization and longer-term soil C storage. Biol Fertil Soils 44:315–320
Swallow M, Quideau SA (2013) Moisture effects on microbial communities in boreal forest floors are stand-dependent. Appl Soil Ecol 63:120–126
Xu X, Yang B, Wang H, Cao Y, Li K, Gao S (2019) Temperature sensitivity of soil heterotrophic respiration is altered by carbon substrate along the development of Quercus Mongolica forest in northeast China. Appl Soil Ecol 133:52–61
You G, Zhang Z, Zhang R (2019) Temperature adaptability of soil respiration in short-term incubation experiments. J Soils Sediments 19:557–565
Zhang G (2010) Changes of oil labile organic carbon in different land uses in Sanjiang Plain, Heilongjiang Province. Chin Geogr Sci 20:139–143
Funding
This study was financially supported through a grant from the Polish National Science Centre (00421/NZ8/2012/29) and Jagiellonian University (DS 759).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Zhenli He
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Klimek, B., Chodak, M. & Niklińska, M. Soil respiration in seven types of temperate forests exhibits similar temperature sensitivity. J Soils Sediments 21, 338–345 (2021). https://doi.org/10.1007/s11368-020-02785-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-020-02785-y