
Abstract
To effectively restore species and habitat diversity in poorly managed degraded grasslands, it is important to understand the relationship between the soil seed bank and aboveground vegetation. Our study aimed to describe changes in soil seed bank composition compared to aboveground vegetation under two ploughing regimes (annual and single ploughing) of narrow strips on the edges of artificially grassed old fields in Podyjí National Park, the Czech Republic. We used the seed germination method, multivariate analyses and Kruskal–Wallis tests to compare differences in the proportion of species and individual numbers in the soil seed bank based on the origin of species (native species, archaeophytes, neophytes) and their habitat preferences (segetal, ruderal, grassland, endangered species) in two types of grasslands with varying soil moisture content. The highest numbers of individuals were present in annually ploughed dry grasslands whereas single ploughing in mesic grasslands resulted in greater species diversity. Ploughing increased the numbers of seedlings of all groups and promoted archaeophytes, grassland specialists, native species and ruderals in dry grasslands whereas mesic grasslands hosted segetal specialists and neophytes. Threatened plants and invasive species rarely emerged from the soil seed bank. The study also revealed the highest similarity between the soil seed bank and aboveground vegetation in annually ploughed plots. It gradually decreased during vegetation succession in the single ploughed variants. Whereas annual ploughing supported segetal specialists, single ploughing promoted grassland species in dry habitats. Neither type of ploughing can be recommended in mesic grasslands because of the risk of inducing the spread of expansive ruderals or invasive neophytes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The European agricultural landscapes have experienced a dramatic loss of biodiversity caused by the intensification of agriculture and the abandonment of historically used traditional farming methods (Poschlod and WallisDeVries 2002; Pärtel et al. 2005; Kleijn et al. 2009; Tsiafouli et al. 2015). The biodiversity decline is particularly evident in the loss of valuable grassland habitats and the subsequent decline of threatened plant species, both at the local (seed limitation; Turnbull et al. 2000; van Klink et al. 2017; Buisson et al. 2019) and landscape (dispersal limitation) scales (Butaye et al. 2005; Tscharntke et al. 2011; Storkey et al. 2012; Meyer et al. 2013; Münzbergová and Herben 2005; Hölzel et al. 2012). However, limited seed supply and dispersal is not the only cause of this decline. In addition, the use of artificially produced seed mixtures for grassland establishment on former arable land has led to overgrowth and the conversion of grasslands into species-poor swards (Dengler et al. 2014; Watson et al. 2014; Buisson et al. 2019). In Czechia, changes in farming practices post-1989 resulted in the widespread abandonment of arable land. While numerous fields were grassed with commercial seed mixtures, some were left to spontaneous succession (Hejcman et al. 2013). The associated negative phenomena of vegetation succession, such as the spread of shrubs into grasslands, the decline of native species and the spread of non-native species, are particularly evident in protected areas where a non-interventional management regime has been applied for a long time. Therefore, the challenge for contemporary conservation management lies in the successful restoration and long-term diversity maintenance of such species-poor grasslands, which have often formed on abandoned old fields (Prach et al. 2013, 2014a; Slodowicz et al. 2023). Abandoned old fields have been identified as suitable habitats for studying spontaneous succession (Rejmánek and van Katwyk 2005; Sojneková and Chytrý 2015; Ryspekov et al. 2021) and are also favourable habitats for the restoration of species-rich meadows with a high conservation value (Bakker et al. 1996; Willems and Bik 1998; Blomqvist et al. 2003; von Blackenhagen and Poschlod 2005; Schmiede et al. 2009). Spontaneous grassland recovery in old fields has also been highlighted as a low-cost restoration method wherever diaspore resources are available in the vicinity of sites undergoing restoration (Prach et al. 2007; Jírová et al. 2012; Prach et al. 2014a, b).
Arable fields have been dependent on long-term ploughing, and small-scale ploughing has proved to be effective at re-opening of dense swards and reducing competition from undesirable species. It also promotes species diversity in old fields (Austrheim and Olsson 1999; Fabšičová et al. 2016; Pereira et al. 2024), or even in heavily disturbed or degraded grasslands (Török et al. 2011; Kiss et al. 2020). However, information on the appropriate intensity and frequency of these interventions for different grassland types is still limited, although Pakeman and Small (2005) suggest that the intensity of disturbance, in terms of depth of disturbed soil, will determine the composition and rate of vegetation recovery. By contrast, Stroh et al. (2012) emphasize the effect of vertical soil mixing.
One of the key roles in the formation of grassland communities is played by the species and trait composition of the soil seed bank, acting as a reservoir of ‘hidden diversity’. This concept is part of the broader ‘dark diversity’ framework, which includes species absent from the aboveground vegetation but capable of establishing themselves at a certain site, effectively constituting the absent portion of a site-specific species pool (Pärtel 2014; Lewis et al. 2016; de Bello et al. 2016; Moeslund et al. 2017; Brown et al. 2019; Carmona and Pärtel 2020). The soil seed bank provides a survival mechanism for plants under unfavourable conditions (Fenner and Thompson 2006; James et al. 2007), buffering unpredictable fluctuations in species abundance in the present vegetation, and should reflect the site’s long-term management, past disturbances and the longevity of the seeds of each species it contains (Waldhardt and Otte 2001; Pakeman and Small 2005). The soil seed bank can thus act as a ‘memory’ of the former plant community with the potential for species to re-appear on the site (Bakker et al. 1996). The species exhibit various strategies, including a transient (up to one or two years), short-term (1–5 years), long-term (5–25 years) persistent, and semi-permanent (> 25 years) soil seed bank (Thompson and Grime 1979; Tyler et al. 2021). The proportion of species with different strategies largely depends on the habitat type, site conditions and the history of the site (Hutchings and Booth 1996; Bekker et al. 1997). In frequently disturbed sites such as arable fields, the vegetation structure is primarily determined by the combination of the soil seed bank composition and seed rain, especially when passive restoration is applied (Luzuriaga et al. 2005). Deep ploughing of species-poor, nutrient-enriched temperate grasslands has been found to reduce residual soil fertility, limit the expansion of tall competitive grasses, and promote the spread of target herbaceous species (Bakker et al. 2012; Lyons et al. 2023). However, the proportion of grassland target species in the soil seed bank of most European degraded grasslands is often low, as many of them form a short-term soil seed bank (< 5 years) largely composed of species occurring in early successional stages or common species (Kiss et al. 2018; de Souza Vieira and Overbeck 2020). Dölle and Schmidt (2009) studied the effect of ploughing on former fields and observed that the amount of seeds in the soil seed bank decreases with decreasing disturbance frequency as the number of species rises. The long-term viability of seeds in the soil is essential for the potential recovery of endangered plant species populations, particularly in disturbed habitats and former arable fields. However, ploughing brings seeds to the soil surface, which stimulates their germination, but can potentially lead to seed bank depletion (Bossuyt and Hermy 2003). As a result, seeds of target species may disappear from the soil seed bank whereas the number of seeds of ruderal and segetal or competitive species may increase. Some authors report a significant enhancement of ruderal species due to ploughing (Wellstein et al. 2007; Lutman et al. 2002). Numerous studies have explored the potential of restoring various habitats using soil seed banks, yielding divergent results (Bakker et al. 1996; Bekker et al. 1997; Klimeš et al. 2010; Kalamees et al. 2012; Stroh et al. 2012), particularly when the target species have disappeared from the aboveground vegetation but persist in the soil seed bank (Warr et al. 2016; Padonou et al. 2022). Their results indicate that attempts at recovering plant communities solely from their soil seed bank are often unsuccessful (Bakker et al. 1996; Dölle and Schmidt 2009; Bossuyt and Honnay 2008; Blomqvist et al. 2003; Smith et al. 2002; Prach and Pyšek 2001; Prach et al. 2014a). Wellstein et al. (2007) observed that only a few grassland species could be recovered from the soil seed banks after their loss from aboveground vegetation, and most studies also report that rare species are often absent from the soil seed bank. By contrast, Kalamees et al. (2012) and Stroh et al. (2012) found unexpectedly high seed density and species richness in the soil seed bank even after prolonged periods of abandonment (25–60 years) in alluvial grasslands and degraded fen meadows. Nevertheless, our knowledge of soil seed bank dynamics in degraded and restored grasslands remains limited or lacking (Engst et al. 2017).
By applying different small-scale ploughing treatments, we studied the importance of the soil seed bank and associated aboveground vegetation for increasing the diversity of degraded species-poor grasslands that were formerly arable fields. We focused on the different functional groups of species present in the soil seed bank of two types of grassland differing in moisture conditions. Using small-scale annual ploughing to restore former segetal vegetation and single ploughing applied for the purpose of supporting the seed germination of grassland specialists, we aimed to answer the following questions: (i) How does the species composition of the soil seed bank vary across different types of grassland and site conditions; (ii) How does the species composition of the soil seed bank differ from that of the aboveground vegetation; (iii) What is the proportion of plant categories in relation to the origin (native species, archaeophytes, neophytes) and habitat preferences (segetal, ruderal, grassland species) of species in the soil seed bank and how can this be influenced by different ploughing treatments; and (iv) What are the benefits and downsides of small-scale ploughing to the facilitate germination of rare plants stored in the soil seed bank.
Material and Methods
Study Site
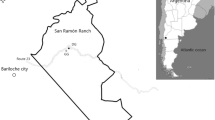
We conducted our research in the area of the Podyjí National Park (PNP) and its protection zone in the south-eastern part of Czechia (Fig. 1). The axis of the region is defined by the deeply incised valley of the river Dyje. The elevation of the PNP varies between 207 and 536 m a. s. l. The climate of the territory is warmer and drier in the southeast (the average annual temperature here is 9.8°C and the annual average sum of precipitation is 500 mm); in the western part it is colder and more humid (average annual temperature 8°C and annual average sum of precipitation 620 mm; averages for the period of 1991–2020, www.chmi.cz). The geology of the PNP is highly diverse, dominated by granites, altered rhyolites and amphibolites weathering into acidic to neutral soils. Due to the location of the territory on the boundary of two phytogeographical units (the Hercynian and the Pannonian region)¸ a significant climatic and vegetation gradient is formed in the area. At present, most of the area is covered by woodlands (oak–hornbeam woods, ravine woods, acidophilous oak woods and thermophilous oak woods). Historically, the landscape has been shaped mainly by traditional farming practices, hay meadows, grazed pastures, viticulture and arable farming, which prevailed here up to the middle of the 20th century. Until then, the landscape was largely treeless. The cessation of traditional management in most of the PNP, the displacement of the inhabitants of some villages, and limited farming in the border zone led to the gradual afforestation of the landscape. For the second half of the 20th century, deforested areas were maintained in several places for military purposes (training grounds, shooting areas, strips of barbed wire along the state border). Nowadays, modern conservation practices promote both no-touch woodland areas and active management that supports dwindling treeless habitats like pastures or hay meadows (eradication of invasive species, removal of woody plants in grasslands, maintenance of treelessness through horse and sheep grazing). Mesophilous grassland types (Arrhenatherion elatioris alliance) of different age and historical management mainly prevailed in the western part of the territory and the Dyje river valley. At some localities, transitions to alluvial meadows, broad-leaved dry grasslands of the alliance Cirsio-Brachypodion pinnati or productive grasslands originating from recently sown seed mixtures were recorded. Dry acidophilous grasslands of the alliance Koelerio-Phleion phleoidis prevailed in the eastern part of the Podyjí National Park. Heterogeneous abiotic conditions combined with a specific varied history have led to the amassment of extremely high plant diversity, and the PNP now hosts an extraordinarily high number of plant species compared even to considerably larger protected areas in Central Europe. Moreover, the PNP hosts a notable proportion of endangered species linked to some form of active management (Němec et al. 2021).

Top – Map of Czechia (CZ) and Austria (AT) marking the position in the Podyjí National Park where study sites were selected for ploughing treatments. Bottom – Design of experimental treatments at one locality as an example (Fládnitzská chata, FLC in the late summer of 2020; A – land subjected to annual ploughing, B – land after single ploughing two years ago; and C – non-ploughed grassland
Experimental Design
The research was performed at twelve experimental grassland sites (see Table S1 in Electronic supplementary material 1) of different species composition and site history. Originally, these were mostly arable fields artificially grassed over using species-poor seed mixtures or left to spontaneous vegetation succession. We observed mesophilous grassland types of the aliance Arrhenatherion elatioris (localities 6, 8, 9, 11, 12), transitions to alluvial meadows (locality 11), broad-leaved dry grasslands of the alliance Cirsio-Brachypodion pinnati (locality 5), species-poor grassed meadows (locality 9), and dry acidophilous grasslands of the alliance Koelerio-Phleion phleoidis (localities 1–4 and 7). Two types of experimental treatments and control plots were studied at all twelve sites, where the area of each variant was typically 0.1–0.2 ha (Fig. 1). Treatment A – ploughing annually, every year in March. Treatment B – single ploughing, plots were ploughed only at the beginning of the study period in March 2019. Treatment C – no ploughing, a control plot with grassland vegetation directly adjacent to ploughed plots. The ploughing depth ranged from 18 to 25 cm, and the soil surface was levelled using a harrow. Single mowing took place annually at the end of August on the same date for all the treatments. The biomass was harvested and then removed from the study site.
Soil Sampling and Cultivation of Soil Seed Bank Samples
Soil samples were collected over a three-year period of 2019–2021 in permanent plots. A design consisting of nine permanent plots was established at each site (three treatments and three replications for each treatment; plot size 4 × 4 m). An Edelman combination auger was used to collect soil samples, which can sample a cylindrical soil monolith 4.5 cm in diameter and 20 cm high. Soil samples were collected at five points within each permanent plot (one sample near its centre and one near each corner). Samples were taken twice a year, in April and November. After the fieldwork was completed, soil samples were taken to the laboratory and stored in a freezer at −18°C until mid-December, when the current year's samples were taken to thaw the soil. Soil from all samples from each plot (5 springs and 5 autumns) was homogenized and all rhizomes and roots were removed. Two litres of soil from each mixed sample were placed in 47 × 20 × 10 cm trays, which were put in 60 × 40 × 10 cm boxes and then transferred to a greenhouse. A dark, non-woven, water-permeable fabric was placed on the bottom of each tray and a layer of sterilized sand about 2 cm thick was spread over it (Fig. 2). The layer of sand was again covered with a strip of dark non-woven textile that allowed us to handle the soil samples better when we needed to turn the samples upside down after about half a year of the first cultivation phase (when no more seedlings emerged) so that the seeds in the deeper layers could germinate. A layer of the homogenized sample was spread on top of the textile and was covered with a white non-woven cloth to prevent seeds from around the greenhouse from entering the trays. The trays were then placed in larger plastic boxes on wooden sticks so that the water from the watering could flow freely to them. Periodic evaluation of the emerging seedlings was then carried out throughout the whole year according to ter Heerdt et al. (1996). Seedlings that could be identified were recorded and removed from the trays (Fig. 2). Poorly developed or unknown individuals (e.g. of biennial species) were cultivated for a longer time in separate pots until they could be determined. Two-week intervals in the spring and monthly intervals from summer till winter were sufficient to record emerging seedlings, and in each case, the species monitoring date was scheduled before the plants began to form fruits.

Cultivation of soil seed bank samples in an unheated greenhouse. 1 – At the beginning of the cultivation, soil samples are arranged in boxes. 2 – Seedlings emerged over the following weeks. 3 – A control visit took place for the purpose of determining plant individuals, counting them and removing them
Vegetation Sampling
In the permanent plots used for soil sampling, plant species composition was recorded twice a year because of the differing phenology of plant species (in early June and late August). Similarly to the soil sampling, plant species data were collected in the field throughout the same study period (2019–2021), with three replicates for each ploughing treatment, including control plots (Fig. 1). The objective was to obtain a list of plant species currently occurring in the aboveground vegetation of each permanent plot from which the soil sample was taken.
Nomenclature and Data Processing
Plant species names were unified following Kaplan et al. (2019). Where possible, identification down to the species or subspecies level was used; only more problematic taxa were combined into aggregates, sections, or genera (see Table S2 in Electronic supplementary material 2). Because most of the taxa were species according to the nomenclature followed, we treat all taxa as species to simplify the text that follows. Unidentified species appearing in the greenhouse (a total of 51 individuals could only be assigned to families) were ultimately excluded from the analyses. The names of syntaxa used in the paper follow Chytrý et al. (2007, 2009). The categories of alien species were unified according to Pyšek et al. (2022), and those of endangered species follow the Red List of Grulich (2017).
Data Analysis
Species lists for both plants emerged from the soil samples in the greenhouse and plants found in the aboveground vegetation were digitized and imported into JUICE (Tichý 2002). Both datasets were merged together and the nomenclature of plants was unified to avoid misinterpretations. For analyses concerning only the dataset for seedlings germinated in the greenhouse experiment, the data originating from phytosociological relevés were removed. To compare datasets of different abundance types (numbers of seedlings in the greenhouse experiment vs Braun–Blanquet scale values from phytosociological relevés), we worked with analyses comparing the datasets, utilizing presence–absence scaling. The numbers of individuals were logarithmically transformed and used only for data pertaining to the seed bank. Calculation of the phi coefficient and Fisher’s exact test were applied to detect species significantly related to ploughing treatments or years. Differences in the numbers of species and the numbers of seedlings from germinated seeds between years, between different moisture levels of graslands and between treatments were tested by Kruskal–Wallis tests with the following non-parametric post-hoc comparisons (StatSoft, Inc 2012). To better understand the effect of ploughing on the soil seed bank composition, the seedlings of plant species were classified and analysed separately for two ecological groups according to the moisture parameters of the grasslands (1 – Dry grasslands category: dry acidophilous grassland vegetation of Koelerio-Phleion phleoidis and broad-leaved semi-dry grasslands of Bromion erecti alliance; 2 – Mesic grasslands category: mesic or mesic–moist grasslands of the Arrhenatherion alliance, including transitions to alluvial grasslands of the river Dyje). Kruskal–Wallis tests with post-hoc comparisons were applied to show whether the soil seed bank had the potential to restore annual weed vegetation (Segetal specialists – species with optima in Annual vegetation of arable land (= habitat 13B) in Sádlo et al. 2007), grassland vegetation (Grassland specialists – grassland habitats: Mesic Arrhenatherum meadows (6A), Montane mesic meadows (6B), Pastures and park grasslands (6C), Alluvial meadows of lowland rivers (6D), Wet Cirsium meadows (6E), Intermittently wet Molinia meadows (6F), Vegetation of wet disturbed soils (6G), Hercynian dry grasslands on rock outcrops (8A), Submediterranean dry grasslands on rockoutcrops (8B), Narow-leaved sub-continental steppes (8C), Broad-leaved dry grasslands (8D), Acidophilous dry grasslands (8E) and Thermophilous forest fringe vegetation (8F) in Sádlo et al. 2007) or whether it will support undesirable ruderal species (Ruderal specialists – habitats with annual or perennial ruderal vegetation types: Annual vegetation of ruderal habitats (13A), Annual vegetation of trampled habitats (13C), Perennial thermophilous ruderal vegetation (13D), Perennial nitrophilous herbaceousvegetation of mesic sites (13E) in Sádlo et al. 2007). Finally, the numbers of species and the numbers of seedlings were categorized into three categories according to the origin of the species in Czechia as native species, archaeophytes and neophytes (Pyšek et al. 2022).
Detrended correspondence analysis (DCA) and canonical correspondence analysis (CCA) were used to explain the differences and variability in species composition of samples derived from different experiments (field survey of phytosociological relevés vs greenhouse experiment with seed germination), from different years (2019–2021), from different ploughing treatments in the field (annual ploughing, single ploughing, no ploughing) and among the twelve sites selected for the study (ter Braak and Šmilauer 2012). Diagrams obtained by unrestricted ordination (DCA) methods were used to visualize the data in graphs, and forward selection and a Monte Carlo permutation test (999 permutations) in CCA were used to detect the significance of variables. The unimodal method was applied because we used presence–absence data, and the length of the first axis in the DCA varied between 2.4–3.8 SD units for each ordinal analysis. Species lists relating to the same locality, year and treatment were merged together to avoid pseudoreplicates. Jaccard’s dissimilarity index was calculated to compare similarities between species-list pairs of different origins (vegetation samples vs soil seed bank samples). ANOVA and Tukey’s HSD post-hoc test were used to prove differences in Jaccard’s dissimilarity statistically.
Results
We found a total of 179 plant species in the soil samples cultivated in the greenhouse, with the most species (147) emerging from samples collected in plots subjected to single ploughing, 136 species growing in pots originating from samples from non-ploughed plots, and 119 species identified among seedlings growing from seed banks sampled in annually ploughed plots. In total, we recorded 43,734 seedlings during the greenhouse cultivation of the soil seed bank samples. Most of the seedlings were recorded in samples from annually ploughed plots (treatment A; 18,369), slightly fewer emerged in samples from single ploughed plots (treatment B; 17,554), and the fewest were observed in samples from non-ploughed plots (treatment C; 7,811 individuals). The highest average number of species per plot was also found for plots ploughed once (15.08) whereas annually and non-ploughed plots contained fewer plant species (13.29 and 13.03, respectively). Significant differences were evident between the years, with the number of species gradually increasing from 2019 to 2020 and 2021, but the seedlings counts did not differ between the years (Fig. 3).

Numbers of species and logarithmized numbers of seedlings emerged from soil seed bank samples in the greenhouse. Different letters (a–c) above boxes indicate statistically significant differences between treatmsnts (A – annual ploughing, B – single ploughing at the beginning of the first year, C – non-ploughed grassland) and years (Kruskal–Wallis test, multiple comparisons of mean ranks for all groups, P < 0.05)
The numbers of species differed depending on the moisture conditions of the habitat, where significantly greater numbers of species were found in habitats with higher moisture (mesic grasslands) in contrast to drier habitats (dry grasslands) in the case of no ploughing (H [K–W test] = −4.916, P = −0.027) and that of single ploughing (H = 7.842, P = 0.005). By contrast, there was no significant result for the annual ploughing treatment (H = 0.57, P = 0.449). Focusing on the numbers of seedlings, we found greater numbers in annually ploughed drier habitats in contrast to habitats with higher moisture (H = 17.25, P < 0.001), and no differences were found for the single ploughing treatment and the no ploughing treatment (H = 0.018, P = 0.893 and H = 0.327, P = 0.568, respectively). By classifying the species into functional groups according to their origin in the Czech flora (native species, archaeophytes, neophytes), we revealed that ploughing generally increased the numbers of seedlings for all groups, native species, archeophytes and neophytes. In regard of the number of species, ploughing led to an increase in non-native species, archaeophytes and neophytes whereas the numbers of native species were found to be lower in both ploughed treatments compared to non-ploughed grasslands (Fig. 4). A higher proportion of grassland specialists was found in non-ploughed plots. For annually ploughed plots, the number of segetal specialists increased over time. Ploughing promoted the occurrence of archaeophytes, grassland specialists and also ruderals on dry grassland sites whereas greater numbers of neophytes were recorded in ploughed plots in mesic grasslands. Mesic grasslands also show a significant increase in native species and a slight decrease in ruderals in the first two years after ploughing. The number of seedlings of grassland specialists mirrored the number of their species and was therefore the greatest in non-ploughed plots. More seedlings of segetal specialists were recorded in ploughed plots in mesic grasslands whereas significantly greater numbers of seedlings of native species and also some ruderals were found in ploughed plots at dry grassland sites. This was not the case with neophytes, where ploughing supported greater seedling numbers only in mesic grasslands; this trend was not observed at dry grassland sites (see Results of the Kruskal-Wallis test in Electronic supplementary material 3). Only six rare and threatened plants of Czechia according to the IUCN and Czech Red List criteria were recorded in the soil seed bank: Centaurium erythraea, Filago arvensis, Myosotis discolor, Myosurus minimus, Verbascum phoeniceum and V. chaixii subsp. austriacum. We found seedlings of the rare species in all treatments, but only in small numbers (from one to several individuals) except for Verbascum phoeniceum (more individuals were present in non-ploughed plots). On the other hand, we recorded only few invasive species, such as Conyza canadensis, Erigeron annuus or Solidago gigantea, emerging from the soil seed bank (see Table S2 in Electronic supplementary material 2 for details). The soil seed bank contained a number of species that were not recorded in the vegetation at the sites (e.g. Gnaphalium uliginosum, Juncus bufonius agg., Lychnis flos-cuculi, Myosurus minimus, Typha sp.). Abundant species both in the soil seed bank and in the aboveground vegetation included, for example, Potentilla argentea, Rumex acetosella, Setaria viridis and Thlaspi arvense. In addition, we should note that 168 species were recorded only in the field and never found emerging in the greenhouse, which is a rather high number. Crepis biennis, Equisetum arvense, Euphorbia helioscopia, Falcaria vulgaris, Galium verum agg., Knautia arvensis and Rosa canina agg. were the most frequent in the aboveground vegetation but were not found emerging from the soil seed bank. Disturbance by ploughing strongly affected the structure of the soil seed bank. The variant of ploughing treatment (three categories according to the ploughing treatment) was even the most significant explanatory variable (F = 7.5, P = 0.001) in the CCA (forward selection, Monte Carlo permutation test). All other tested explanatory variables also appeared to be significant (site, i.e. variability among the 12 localities: F = 6.63, P = 0.001; origin of the data in terms of 1 – the seed bank or 2 – aboveground vegetation: F = 1.66, P = 0.001; and, finally also variation over time – difference between the years 2019, 2020 and 2021: F = 1.33, P = 0.045). Differences between the ploughing treatments are markedly evident from ordination diagrams (DCA, Fig. 5), where this variable was separated along the first axis. The second axis was determined by differences between sites (Figs. 5 and 6), especially the strong moisture gradient (Echium vulgare, Potentilla argentea and Trifolium arvense characterize drier soils in the upper part of the plot whereas Mentha arvensis, Equisetum arvense or Stellaria graminea are typical of soils with higher moisture; see the lower part of the plot); see also Fig. S4 – DCA diagrams for species in Electronic supplementary material 4. In annually ploughed plots, annual, weedy and ruderal species, such as Viola arvensis, Thlaspi arvense or Tripleurospermum inodorum, were more strongly represented in the soil seed bank whereas typical grassland species, such as Hypericum perforatum, Lotus corniculatus, Potentilla argentea, Rumex acetosa, Trifolium repens or Veronica chamaedrys, were frequent both in the non-ploughed treatment and in the variant ploughed only once; see Fig. 6, and Table S3 in Electronic supplementary material 5 for details.

Numbers of native, archaeophyte and neophyte species numbers and logarithmized numbers of seedlings emerged from soil seed bank samples in the greenhouse. Different letters (a–f) above/near boxes indicate statistically significant differences between treatments (A – annual ploughing, B – single ploughing at the beginning of the first year, C – non-ploughed grassland; Kruskal–Wallis test, multiple comparisons of mean ranks for all groups, P < 0.05)

Diagram of DCA analysis showing the positions of soil samples in ordination space. Soil samples were taken in the field and used for germination of plant seedlings in a greenhouse. Symbols refer to the three years of the research (transparent – 2019, light solid fill – 2020, dark solid fill – 2021), data sources and treatments (red – annual ploughing, blue – single ploughing at the beginning of the first year, green – non-ploughed grassland). Total variation: 6.131, the eigenvalues of the first two axes being 0.39 and 0.28

Diagram of DCA analysis showing the positions of plant species germinated in the greenhouse in ordination space based on the same dataset as in the Fig. 5. To avoid overlap, only sixty species with the highest weight are presented (see Table S3 in Electronic supplementary material 5 for full plant names). Colours to differentiate the species optima within the habitats defined for Czechia (Sádlo et al. 2007)
Differences arising over time are clearly visible in the case of the single ploughing treatment (B, Fig. 7). The plots present the results of partial analyses for the individual ploughing treatments (A, B, C), where the differences between the years (especially for variant B) and between the soil seed bank and the aboveground vegetation (especially for variant C) can be seen in detail (Fig. 7). We found that dissimilarities in plant species composition between the aboveground vegetation sampled in the field and the species recorded from germinating soil seed banks varied for all ploughing treatments (F = 18.35, P < 0.001) and over time (F = 4.82, P < 0.05; Fig. 8). When testing for differences between the years for all treatments separately, we found a significant result only for the annual ploughing treatment, where the dissimilarity decreased over time (A; this differed between 2019 and 2021, F = 4.02, P < 0.05).

DCA ordinations presenting similarities between plots based on the plant species data. The first figure (upper left) shows all samples together (triangles – germinating seedlings from soil samples; squares – aboveground vegetation) whereas the other three figures are for samples from plots subjected to particular treatment variants (A – annual ploughing, B – single ploughing and C – no ploughing). Symbols refer to the three years of the research (transparent – 2019, light solid fill – 2020, dark solid fill – 2021). Total variation counted for all samples: 7.396, the same for subset A: 5.202, the same for subset B: 5.321, and the same for subset C: 5.148. Eigenvalues counted for first and second axes are as follows: Axis 1: 0.385, Axis 2: 0.277 for all samples; Axis 1: 0.322, Axis 2: 0.261 for variant A; Axis 1: 0.326, Axis 2: 0.254 for variant B; and Axis 1: 0.420, Axis 2: 0.275 for variant C. Values of variation explained by the DCA axes are indicated in the figure itself

Jaccard dissimilarity between pairs of samples, where one list of species comes from our field survey (aboveground vegetation sampled in the phytosociological relevés) and the other from our experiment with seed cultivation in a greenhouse (the results of Tukey post-hoc test are represented using letters above the boxes)
Discussion
Effect of Different Ploughing Treatments on the Species and Functional Composition of the Soil Seed Bank of Two Moisture-Distinct Grassland Types
Our study of soil seed banks revealed that they generally contain a large number of viable seeds, a typical characteristic of previously managed arable land (Thompson et al. 1997). Whereas non-ploughed plots had the lowest number of individuals in their soil seed banks (about half the number of seedlings from ploughed plots), the single ploughing treatment resulted in the greatest average number of species, probably due to the high species turnover during three years of succession (Schnoor and Olsson 2010). A functional group analysis of species in the soil seed banks of two distinct grassland types differing in moisture showed that ploughing promoted a greater number of seedlings across all functional groups, indicating a tendency towards generative reproduction. This trend varied between different habitat types in relation to site moisture conditions. In dry grassland sites, ploughing generated more species (qualitative changes), while in mesic grassland habitats, seedling numbers were more supported (quantitative changes); this is consistent with the different successional trajectories in dry and mesic grasslands (Prach et al. 2007). Ploughing of dry grassland sites supported archaeophytes and grassland specialists, as well as some ruderals, to a greater extent (Sojneková and Chytrý 2015). By contrast, ploughing in mesic grasslands promoted both native species (e.g. Elymus repens, Artemisia vulgaris) and neophytes (e.g. Erigeron annuus, Solidago gigantea). The large number of seedlings of segetal specialists and neophytes confined to mesic grasslands was likely directly related to soil moisture and a higher content of available nutrients. The large amounts of nutrients, especially nitrogen, released during ploughing contribute to the dominance of ruderal species in early stages of vegetation succession (Prach 2003; Wellstein et al. 2007). Annual ploughing resulted in a slight increase in the number of species over time, especially segetal specialists, although a small quantity of perennial ruderal species also appeared in the soil seed bank, besides being present in the aboveground vegetation. This is in line with the fact that annuals and biennials almost always have more persistent seeds than related perennials (Thompson et al. 1997). The difference in the number of individuals between the treatments was mainly due to the high abundance of seeds of a few species, mostly annuals, weeds or ruderals that occurred in high frequencies in ploughed plots. The highest seedling numbers were reached by species such as Chenopodium album agg. (6,711 seedlings), Artemisia vulgaris (5,984 seedlings), Polygonum aviculare agg. (4,338 seedlings) and Juncus bufonius agg. (4,130 seedlings). Despite the high seed production, increased numbers of seeds of ruderal species in the soil in ploughed sites do not always mean increased numbers of their seedlings (Rahman et al. 2000, 2006). Large annual losses of viable seeds of Chenopodium album agg. have been found by Lutman et al. (2002) as a consequence of the depletion of the soil seed bank, a phenomenon observed in abandoned fields (Davies and Waite 1998; Dölle and Schmidt 2009). In principle, the rate of loss depends on a number of factors such as the farming system, the soil type, size and shape of seeds, and the seed longevity in the soil. The seed longevity in the soil seed banks is supposed to be one of the key factors for successful habitat restoration (Bakker et al. 1996; Guo et al. 2023; Suleiman et al. 2023). Seeds of short-lived species (typically annuals) are generally more persistent than seeds of long-lived species, and perennial seeds are in turn typical of species living in disturbed habitats (Wang et al. 2013). In dry grasslands in unploughed plots, former arable fields grassed over decades ago, we recorded large numbers of seedlings of segetal specialists (Arabidopsis thaliana, Viola arvensis), which implies that segetal specialists and also some grassland species can survive in the soil seed bank for several decades after the abandonment of the former field (Sojneková and Chytrý 2015) or probably even centuries. Disturbance in the form of annually ploughing tends to increase the abundance of weeds over time and thus select for species with long-lasting seeds (Kleyer 1999; Thompson et al. 1997). By contrast, single ploughing in dry grassland types led to an increase in species numbers in the soil seed bank. However, typical grassland species, such as Achillea millefolium agg., Hypericum perforatum, Lotus corniculatus, Plantago lanceolata, Potentilla argentea, Rumex acetosa, Trifolium spp. or Veronica chamaedrys, increased their abundances in the soil seed bank only slowly over time. On the other hand, the soil seed bank of grassland vegetation tends to be poorer in the number of individuals but not in species composition (Bossuyt and Honnay 2008; Schellenberger et al. 2022). This is due to the presence of species with perennial life forms in the vegetation, especially grasses, which are often closely linked to predominantly clonal dispersal, a longer time required for seed production, fewer seeds on plants, or seed predation.
Interestingly, several species that were not present in the vegetation at all (e.g. Gnaphalium uliginosum, Juncus bufonius agg., Lychnis flos-cuculi, Myosotis discolor, Myosurus minimus, Typha sp.) were recorded in the soil seed bank samples. These are mostly hygrophilous species of gap vegetation whose seed germination was probably promoted by regular watering during the greenhouse cultivation. Juncus bufonius agg. was one of the most abundant species ‘hidden’ in the soil seed bank, as also reported by Berge and Hestmark (1997), and grew mostly in samples from non-ploughed plots. The occurrence of this species in the soil seed bank was rather high even though the study sites were mostly xeric to mesophilic and this species did not occur in the present-day aboveground vegetation. This seems to be a typical strategy of Juncus species, whose seeds are often found in large numbers in the seed bank even in places where Juncus is absent from the vegetation, as also reported by Stroh et al. (2012). By contrast, a remarkable number of species were recorded only in the aboveground vegetation and did not emerge from the soil seed bank (e.g. Crepis biennis, Falcaria vulgaris, Galium verum agg., Knautia arvensis and others). This may be down to a variety of reasons: It may be related to the management history of the site, low seed production or persistence of only some species, or because the collected soil samples covered a much smaller area than the vegetation records, etc. (DeMalach et al. 2023; Hopfensperger 2007; Karlík and Poschlod 2014; White et al. 2012). A significant proportion of such non-recruiting species is composed of representatives of the Asteraceae family, whose seedlings form small leaf rosettes and may have been difficult to identify in cultivation, also due to their long-term persistence in the vegetative phase and losses caused by seed predation. This may also be the case with seeds of Rosa canina agg. and other tree and shrub species that occurred in the vegetation or the permanent plots. As for other species, it was quite surprising that one common annual weed species, Euphorbia helioscopia, was not found emerging from the soil seed bank. This could be due to a very low density of its plants at the experimental sites that complicated the capturing of its seeds in soil samples and by suboptimal conditions for the growth of the species during the greenhouse cultivation. There were only two seeds left after the determination of ungerminated in all samples taken in 2019 from the Mašovice site (Zdražílková, pers. comm.). Another common species, Equisetum arvense, a sporogenous horsetail, was a special case. Its spores have a short viability of about only two weeks and the species is dispersed on arable land vegetatively by rhizome segments (Lebkuecher 1997). Despite E. arvense being abundant in the aboveground vegetation, its spores probably did not survive storage in the freezer, or the plants did not produce sporophytes and therefore could not be recorded. Some seedlings were left unidentified to the species level and were therefore excluded from the analyses.
Impact of Ploughing Disturbance on the Relationship Between the Soil Seed Bank and Aboveground Vegetation
To understand how community composition is influenced by factors such as disturbance and succession, the similarity between the soil seed bank and aboveground vegetation is frequently studied (Hopfensperger 2007; White et al. 2012). The degree of similarity may indicate the potential of the soil seed bank to maintain and restore aboveground vegetation following a disturbance, suggesting that the soil seed bank is more reflective of the vegetation (Valkó et al. 2011). The relationship between the soil seed bank and vegetation can also provide insight into community resilience to disturbance. Generally, ploughing is considered a relatively strong type of disturbance that significantly affects the structure of the soil seed bank and subsequently that of the aboveground vegetation (Jongepierová 2008; Klaus et al. 2017). In our analyses, it was the ploughing method that emerged as the most important variable explaining differences in the species composition of the soil seed bank and aboveground vegetation. In grasslands, the similarity between the soil seed bank and the aboveground vegetation sometimes increases after a sudden and intense disturbance, which includes ploughing (Hopfensperger 2007). We recorded increasing similarity between the soil seed bank and the current vegetation in the annually ploughed plots, but the similarity decreased in the single ploughed plots over time. The same result was emphasized also by Dessaint et al (1997) and Bekker et al (1999), who found that only communities dominated by annual plants or communities with frequent disturbance showed a marked similarity in species composition between the soil seed bank and the vegetation. Soil seed banks that consist mainly of perennial species often have a lower degree of similarity in terms of species composition to the aboveground vegetation (Touzard et al. 2002; Blood et al. 2010; Loydi et al. 2012). Touzard et al. (2002) also noted that the majority of seedlings emerging after experimental disturbances were mainly recruited from the soil seed bank. This suggests that ploughing activates the seeds stored in the soil seed bank, contributing to the restoration of diversity in species-poor grasslands and potentially promoting the establishment of rare plant seedlings. We expected the species composition of vegetation in the ploughed treatments to be more similar to that of the soil seed bank, as it is assumed that species from the soil seed bank that were previously suppressed by dense vegetation would germinate after ploughing (Klaus et al. 2017). Instead, we have confirmed the low similarity between the soil seed bank and aboveground vegetation in grasslands, as reported by Janicka (2016) or Skuodienė and Matyžiūtė (2022). Furthermore, the dissimilarity did not change significantly over time. Completely the opposite situation was observed in the once ploughed plots, which in the third year of succession after ploughing began to resemble grasslands, at least in physiognomy. A general trend reported in various grassland studies is that the similarity in species composition between the seed bank and the vegetation increases with the age of the system (Hopfensperger 2007), although some studies report a decrease in similarity during succession in grasslands (Kalamees and Zobel 1998; Wagner et al. 2003). The effect of different types of ploughing treatments on aboveground vegetation in the Podyjí National Park is discussed by Vymyslický et al. (2024).
Can the Composition of the Soil Seed Bank Support the Restoration of Plant Diversity?
The study of soil seed banks provides information about past disturbances (Hong et al. 2012), management practices and land use history (Karlík and Poschlod 2014; Valkó et al. 2011; Stroh et al. 2012) and plays an important role in the spontaneous restoration of grasslands (Jongepierová et al. 2004; Wagner et al. 2018). The results of our present study confirm that small-scale disturbances, such as annual ploughing of grassland margins in old fields, can support segetal specialists emerging from the soil seed bank. Ödman et al. (2012) also indicated that in order to promote diversity at a landscape scale, present-day disturbance management should be based on the historical disturbance regime, habitat productivity and propagule supply. Such a disturbance could support some endangered species, locally rare species or even species absent from the aboveground vegetation for a long time (Prach et al. 2007; Sojneková and Chytrý 2015). However, it is important to note that a significant proportion of the land remains untouched by ploughing, providing both anti-erosion protection and long-term continuity of the grassland ecosystem. On the other hand, diversification of sites through different small-scale ploughing regimes and cutting or pastoral management can create heterogeneous mosaics and thereby increase the local biodiversity of habitats with herbaceous vegetation. Our research has recorded several threatened plants, for example Pseudognaphalium luteoalbum, a critically endangered annual species, which emerged from soil seed bank samples (Entová 2011). However, during subsequent cultivation (2012–2022), we observed only six endangered species emerging from the soil samples, and with the exception of Myosurus minimus, they were also found in the aboveground vegetation. In this context, it should be mentioned that rare species were scarce even before the old fields were grassed over (Němec et al. 2021), so the chance of them being captured by the soil sampling was relatively small. On the other hand, we found that the soil seed bank contained seeds of species that have declined significantly in the region but are not included in the national Red List, for example Spergula arvensis and Chenopodium glaucum (Němec et al. 2021). In general, dry grassland soil seed banks are characterized by transient seeds and contain only few persistent seeds of typical grassland species (Bossuyt and Honnay 2008; Kiss et al. 2016). We found that our cultivation method has not been very successful in detecting rare species that were present in the soil seed bank but not in the aboveground vegetation. This could be due to the fact that the conditions used for the cultivation in the greenhouse stimulated the emergence of hygrophilous species rather than that of mesophilous or xerophilous ones. To get a comprehensive picture of the amount of viable seeds in the collected samples, it will be necessary to supplement cultivation with further determination of ungerminated seeds (Fabšičová et al. 2023). This is the only way to answer the question of the suitability of ploughing as a measure for supporting rare species of segetal specialists, which have practically disappeared from the aboveground vegetation of the study sites. However, in degraded grasslands that were regularly managed as arable land in the past, the potential for restoration from the soil seed bank is rather limited (Török et al. 2018; Valkó et al. 2021).
Conclusion
The soil seed bank plays a crucial role in vegetation recovery after ploughing, but its effectiveness depends on the type of treatment. Annual ploughing tends to promote segetal specialists and ruderals, particularly on moist and nutrient-rich soils. On the other hand, single ploughing supports archaeophytes and grassland species, enhancing the overall diversity of grasslands on drier and nutrient-poor soils. Ploughing can support the emergence of rare species, especially annuals, weeds and hygrophilous species that rely on frequent disturbances. However, due to their low abundance in the soil seed bank, such species successfully germinate and establish themselves in the vegetation rather sporadically, depending on whether the environmental conditions are suitable. The management impact of ploughing activates the soil seed bank, but this effect persists only for two years after the initial ploughing. Therefore, it must be repeated subsequently to support the emergence of annual and competitively weak species. Our findings also confirm that the soil seed banks at our study sites do not serve as a reservoir for invasive species. The presence of invasive species in aboveground vegetation has more to do with repeated colonization from nearby sites than with the accumulation of seeds in the soil. Such species were present in low numbers mainly in mesic grassland sites. For this reason, we cannot recommend any type of ploughing in mesic grasslands with a higher soil nutrient content. In mesic grasslands, we suggest ploughing management only at sites without expansive ruderals or invasive neophytes in the vicinity because of the risk that these species might spread into newly established niches. In summary, detailed analyses of soil seed banks and their relationship to aboveground vegetation contribute to our understanding of how to successfully restore degraded species-poor grassland habitats and allow us to select appropriate management interventions for different vegetation types and habitats.
Data Availability
All datasets generated and analysed during the current study are available from the corresponding author on request.
References
Austrheim G, Olsson EGA (1999) How does continuity in grassland management after ploughing affect plant community patterns? Pl Ecol 145:59–74
Bakker JP, Poschlod P, Strykstra RJ, Bekker RM, Thompson K (1996) Seed banks and seed dispersal: important topics in restoration ecology. Acta Bot Neerl 45:461–490
Bakker JP, Van Diggelen R, Bekker RM, Marrs RH (2012) Restoration of dry grasslands and heathlands. In van Andel J, Aronson J (eds) Restoration ecology: the new frontier. Blackwell Publishing, pp 173–188
Bekker RM, Verweij GL, Smith REN, Reine R, Bakker JP, Schneider S (1997) Soil seed banks in European grasslands: does land use affect regeneration perspectives? J Appl Ecol 34:1293–1310
Bekker RM, Lammerts EJ, Schutter A, Grootjans AP (1999) Vegetation development in dune slacks: the role of persistent seed banks. J Veg Sci 10:745–754
Berge G, Hestmark G (1997) Composition of seed banks of roadsides, stream verges and agricultural fields in southern Norway. Ann Bot Fenn 34:77–90
Blomqvist MM, Bekker RM, Vos P (2003) Restoration of ditch bank plant species richness: the potential of the soil seed bank. Appl Veg Sci 6:179–188
Blood LE, Pitoniak HJ, Titus JH (2010) Seed bank of a bottomland swamp in western New York. Castanea 75:19–38
Bossuyt B, Hermy M (2003) The potential of soil seed banks in the ecological restoration of grassland and heathland communities. Belg J Bot 136:23–34
Bossuyt B, Honnay O (2008) Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J Veg Sci 19:875–884
Brown JJ, Mennicken S, Massante JC, Dijoux S, Telea A, Benedek AM, Götzenberger L, Májeková M, Lepš J, Šmilauer P, Hrček J, de Bello F (2019) A novel method to predict dark diversity using unconstrained ordination analysis. J Veg Sci 30:610–619
Buisson E, Le Stradic S, Silveira FA, Durigan G, Overbeck GE, Fidelis A, Fernandes GW, Bond WJ, Hermann J-M, Mahy G, Alvarado ST, Zaloumis NP, Veldman JW (2019) Resilience and restoration of tropical and subtropical grasslands, savannas, and grassy woodlands. Biol Rev 94:590–609
Butaye J, Adriaens D, Honnay O (2005) Conservation and restoration of calcareous grasslands: a concise review of the effects of fragmentation and management on plant species. Biotechnol Agron Soc Environm 9:111–118
Carmona CP, Pärtel M (2020) Estimating probabilistic site‐specific species pools and dark diversity from co‐occurrence data. Global Ecol Biogeogr 30:316–326
Chytrý M (ed) (2007) Vegetation of the Czech Republic. 1, Grassland and heathland vegetation. Academia, Praha
Chytrý M (ed) (2009) Vegetation of the Czech Republic. 2, Ruderal, weed, rock and scree vegetation. Academia, Praha
Davies A, Waite S (1998) The persistence of calcareous grassland species in the soil seed bank under developing and established scrub. Pl Ecol 136:27–39
de Souza Vieira M, Overbeck GE (2020) Small seed bank in grasslands and tree plantations in former grassland sites in the South Brazilian highlands. Biotropica 52:775–782
de Bello F, Fibich P, Zelený D, Kopecký M, Mudrák O, Chytrý M, Pyšek P, Wild J, Michalcová D, Sádlo J, Šmilauer P, Lepš J, Pärtel M (2016) Measuring size and composition of species pools: a comparison of dark diversity estimates. Ecol Evol 6:4088–4101
DeMalach N, Kigel J, Sternberg M (2023) Contrasting dynamics of seed banks and standing vegetation of annuals and perennials along a rainfall gradient. Perspect Pl Ecol Evol Syst 58:125718
Dengler J, Janišová M, Török P, Wellstein C (2014) Biodiversity of Palaearctic grasslands: a synthesis. Agric Ecosys Environm 182:1–14
Dessaint F, Chadoeuf R, Barralis G (1997) Nine years’ soil seed bank and weed vegetation relationships in an arable field without weed control. J Appl Ecol 34:123–130
Dölle M, Schmidt W (2009) The relationship between soil seed bank, above‐ground vegetation and disturbance intensity on old‐field successional permanent plots. Appl Veg Sci 12:415–428
Engst K, Baasch A, Bruelheide H (2017) Predicting the establishment success of introduced target species in grassland restoration by functional traits. Ecol Evol 7:7442–7453
Entová M (2011) Úhory v Národním parku Podyjí jako možná lokalita pro protěž žlutobílou (Pseudognaphalium luteoalbum). Thayensia 8:311–314
Fabšičová M, Vymyslický T, Entová M, Kůrová J (2016) Mohou být úhory refugii vzácných a ohrožených druhů rostlin? Thayensia 13:89–102
Fabšičová M, Frei I, Jiroušek M, Smetanová S, Šipoš J, Trnka F, Vymyslický T, Winkler J, Zdražílková M (2023) Podpora biodiverzity travních porostů pomocí maloplošných úhorů. Uplatněná certifikovaná metodika 62/23, Baštan, Olomouc, 61 pp
Fenner M, Thompson K (2006) The Ecology of Seeds. Cambridge University Press, New York
Grulich V (2017) Červený seznam ohrožených druhů České republiky: cévnaté rostliny. Příroda 35:75–132
Guo N, Sang C, Huang M, Zhang R, Degen AA, Ma L, Bai Y, Zhang T, Wang W, Niu N, Li S, Long R, Shang Z (2023) Long-term active restoration of degraded grasslands enhances vegetation resilience by altering the soil seed bank. Agron Sustain Developm 43:6
Hejcman M, Hejcmanová P, Pavlů V, Beneš J (2013) Origin and history of grasslands in central Europe—a review. Grass Forage Sci 68:345–363
Hölzel N, Buisson E, Dutoit T (2012) Species introduction – a major topic in vegetation restoration. Appl Veg Sci 15:161–165
Hong J, Liu S, Shi G, Zhang Y (2012) Soil seed bank techniques for restoring wetland vegetation diversity in Yeyahu Wetland, Beijing. Ecol Engin 42:192–202
Hopfensperger KN (2007) A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 116:1438–1448
Hutchings MJ, Booth KD (1996) Studies on the feasibility of re-creating chalk grassland vegetation on ex-arable land. I. The potential roles of the seed bank and the seed rain. J Appl Ecol 33:1171–1181
James CS, Capon SJ, White MG, Rayburg SC, Thoms MC (2007) Spatial variability of the soil seed bank in a heterogeneous ephemeral wetland system in semi-arid Australia. Pl Ecol 190:205–217
Janicka M (2016) The evaluation of soil seed bank in two Arrhenatherion meadow habitats in Central Poland. Acta Sci Polon Agric 15:25–38
Jírová A, Klaudisová A, Prach K (2012) Spontaneous restoration of target vegetation in old‐fields in a central European landscape: a repeated analysis after three decades. App Veg Sci 15:245–252
Jongepierová I, Jongepier JW Klimeš L (2004) Restoring grassland on arable land: an example of a fast spontaneous succession without weed-dominated stages. Preslia 76:361–369
Jongepierová I (Ed) (2008) Louky Bílých Karpat [Grasslands of the White Carpathian Mountains]. ZO ČSOP Bílé Karpaty, Veselí nad Moravou
Kalamees R, Zobel M (1998) Soil seed bank composition in different successional stages of a species rich wooded meadow in Laelatu, western Estonia. Acta Oecol 19:175–180
Kalamees R, Püssa K, Zobel K, Zobel M (2012) Restoration potential of the persistent soil seed bank in successional calcareous (alvar) grasslands in Estonia. Appl Veg Sci 15:208–218
Kaplan Z, Danihelka J, Chrtek J, Kirschner J, Kubát K, Štech M, Štěpánek J (2019) Klíč ke květeně České republiky. Ed. 2. Academia, Praha
Karlík P, Poschlod P (2014) Soil seed-bank composition reveals the land-use history of calcareous grasslands. Acta Oecol 58:22–34
Kiss R, Deák B, Török P, Tóthmérész B, Valkó O (2018) Grassland seed bank and community resilience in a changing climate. Restorat Ecol 26:S141–S150
Kiss R, Deák B, Tóthmérész B, Migléc, T, Tóth K, Török P, Lukács K, Godó L, Körmöczi Z, Radócz S, Kelemen A, Sonkoly J, Kirmer A, Tischew S, Švamberková E, Valkó O (2020) Establishment gaps in species-poor grasslands: artificial biodiversity hotspots to support the colonization of target species. Restorat Ecol 29:e13135
Kiss R, Valkó O, Tóthmérész B, Török P (2016) Seed bank research in Central-European grasslands – an overview. In Murphy J (ed.) Seed banks: types, roles and research. Nova Science Publishers, pp 1–34
Klaus VH, Schäfer D, Kleinebecker T, Fischer M, Prati D, Hölzel N (2017) Enriching plant diversity in grasslands by large-scale experimental sward disturbance and seed addition along gradients of land-use intensity. J Pl Ecol 10:581–591
Kleijn D, Kohler F, Báldi A, Batáry P, Concepción ED, Clough Y, Díaz M, Gabriel D, Holzschuh A, Knop E, Kovács A, Marshall EJP, Tscharntke T, Verhulst J (2009) On the relationship between farmland biodiversity and land-use intensity in Europe. Proc Roy Soc London, Ser B, Biol Sci 276:903–909
Kleyer M (1999) Distribution of plant functional types along gradients of disturbance intensity and resource supply in an agricultural landscape. J Veg Sci 10:697–708
Klimeš L, Jongepierová I, Doležal J, Klimešová J (2010) Restoration of a species‐rich meadow on arable land by transferring meadow blocks. Appl Veg Sci 13:403–411
Lebkuecher JG (1997) Desiccation‐time limits of photosynthetic recovery in Equisetum hyemale (Equisetaceae) spores. Amer J Bot 84:792–797
Lewis RJ, Szava-Kovats R, Pärtel M (2016) Estimating dark diversity and species pools: an empirical evaluation of two methods. Meth Ecol Evol 7:104–113
Loydi A, Zalba SM, Distel RA (2012) Viable seed banks under grazing and exclosure conditions in montane mesic grasslands of Argentina. Acta Oecol 43:8–15
Lutman PJW, Cussans GW, Wright KJ, Wilson BJ, Wright GM, Lawson HM (2002) The persistence of seeds of 16 weed species over six years in two arable fields. Weed Res 42:231–241
Luzuriaga AL, Escudero A, Olano JM, Loidi J (2005) Regenerative role of seed banks following an intense soil disturbance. Acta Oecol 27:57–66
Lyons KG, Török P, Hermann JM, Kiehl K, Kirmer A, Kollmann J, Overbeck GE, Tischew S, Allen EB, Bakker JD, Brigham C, Buisson E, Crawford K, Dunwiddie P, Firn J, Grobert D, Hickman K, Stradic SLE, Temperton VM (2023) Challenges and opportunities for grassland restoration: A global perspective of best practices in the era of climate change. Global Ecol Conservation 46:e02612
Meyer S, Wesche K, Krause B, Leuschner C (2013) Dramatic losses of specialist arable plants in Central Germany since the 1950s/60s - a cross-regional analysis. Diversity & Distrib 19:1175–1187
Moeslund JE, Brunbjerg AK, Clausen KK, Dalby L, Fløjgaard C, Juel A, Lenoir J (2017) Using dark diversity and plant characteristics to guide conservation and restoration. J Appl Ecol 54:1730–1741
Münzbergová Z, Herben T (2005) Seed, dispersal, microsite, habitat and recruitment limitation: identification of terms and concepts in studies of limitations. Oecologia 145:1–8
Němec R, Filippov P, Grulich V, Hrouda L, Hummel J, Musil Z, Němcová Z, Reiterová L, Řepka R, Vymyslický T (2021) Rozšíření cévnatých rostlin národních parků Podyjí a Thayatal. Správa NP Podyjí, Znojmo
Ödman AM, Schnoor TK, Ripa J, Olsson PA (2012) Soil disturbance as a restoration measure in dry sandy grasslands. Biodivers & Conservation 21:1921–1935
Padonou EA, Akakpo BA, Tchigossou B, Djossa B (2022) Methods of soil seed bank estimation: a literature review proposing further work in Africa. iForest 15:121–127
Pakeman RJ, Small JL (2005) The role of the seed bank, seed rain and the timing of disturbance in gap regeneration. J Veg Sci 16:121–130
Pärtel M (2014) Community ecology of absent species: hidden and dark diversity. J Veg Sci 25:1154–1159
Pärtel M, Bruun HH, Sammul M (2005) Biodiversity in temperate European grasslands: origin and conservation. Grassland Sci Eur 10:1–14
Pereira AJ, Porto M, Correia O, Beja P (2024) Traditional ploughing is critical to the conservation of threatened plants in Mediterranean olive groves. Agric Eco-Syst Environm 359:108775
Poschlod P, WallisDeVries MF (2002) The historical and socioeconomic perspective of calcareous grasslands—lessons from the distant and recent past. Biol Conservation 104:361–376
Prach K (2003) Spontaneous succession in Central-European man-made habitats: What information can be used in restoration practice? Appl Veg Sci 2:125–129
Prach K, Pyšek P (2001) Using spontaneous succession for restoration of human-disturbed habitats: experience from Central Europe. Ecol Engin 17:55–62
Prach K, Lepš J, Rejmánek M (2007) Old field succession in Central Europe: local and regional patterns. In Cramer VA, Hobbs RJ (eds) Old fields: dynamics and restoration of abandoned farmland. Island Press, Washington, DC, US, pp 180–201
Prach K, Jongepierová I, Řehounková K (2013) Large‐scale restoration of dry grasslands on ex‐arable land using a regional seed mixture: establishment of target species. Restorat Ecol 21:33–39
Prach K, Jongepierová I, Řehounková K, Fajmon K (2014a) Restoration of grasslands on ex-arable land using regional and commercial seed mixtures and spontaneous succession: successional trajectories and changes in species richness. Agric Eco-Syst Environm 182:131–136
Prach K, Řehounková K, Lencová K, Jírová A, Konvalinková P, Mudrák O, Študent V, Vaněček Z, Tichý L, Petřík P, Šmilauer P, Pyšek P (2014b) Vegetation succession in restoration of disturbed sites in Central Europe: the direction of succession and species richness across 19 seres. Appl Veg Sci 17:193–200
Pyšek P, Sádlo J, Chrtek J Jr, Chytrý M, Kaplan Z, Pergl J, Pokorná A, Axmanová I, Čuda J, Doležal J, Dřevojan P, Hejda M, Kočár P, Kortz A, Lososová Z, Lustyk P, Skálová H, Štajerová K, Večeřa M, Vítková M, Wild J, Danihelka J (2022) Catalogue of alien plants of the Czech Republic (3rd edition): species richness, status, distributions, habitats, regional invasion levels, introduction pathways and impacts. Preslia 94:447–577
Rahman A, James TK, Mellsop J, Grbavac N (2000) Effect of cultivation methods on weed seed distribution and seedling emergence. New Zealand Pl Protect 53:28–33
Rahman A, James TK, Grbavac NIK (2006) Correlation between the soil seed bank and weed populations in maize fields. Weed Biol Managem 6:228–234
Rejmánek M, van Katwyk KP (2005) Old field succession: a bibliographic review 1901–1991. Section of evolution and ecology, University of California, Davis. Available at http://botanika.prf.jcu.cz/suspa/pdf/BiblioOF.pdf. Accessed 28 June 2024
Ryspekov T, Jandák J, Balkozha M, Winkler J (2021) Vegetation of abandoned fields on soil types of kastanozems in Northern Kazakhstan (Kostanay Region). J Ecol Engin 22: 176–184
Sádlo J, Chytrý M, Pyšek P (2007) Regional species pools of vascular plants in habitats of the Czech Republic. Preslia 79:303–321
Schellenberger J, Csontos P, Csorba Á, Lengyel A, Málnási Csizmadia G, Szirmai O, Penksza K, Czóbel S (2022) Soil seed banks of continental grasslands with different water regimes—a comparative study from the aspect of recovery potential. Agronomy 12:2830
Schmiede R, Donath TW, Otte A (2009) Seed bank development after the restoration of alluvial grassland via transfer of seed-containing plant material. Biol Conservation 142:404–413
Schnoor TK, Olsson PA (2010) Effects of soil disturbance on plant diversity of calcareous grasslands. Agric Eco-Syst Environm 139:714–719
Skuodiené R, Matyžiuté V (2022) Assessment of an abandoned grassland community and the soil seed bank of a hilly relief. Zemdirbyste Agric 109:3–10
Slodowicz D, Durbecq A, Ladouceur E, Eschen R, Humbert JY, Arlettaz R (2023) The relative effectiveness of different grassland restoration methods: a systematic literature search and meta‐analysis. Ecol Solut Evidence 4:e12221
Smith RS, Shiel RS, Millward D, Corkhill P, Sanderson RA (2002) Soil seed banks and the effects of meadow management on vegetation change in a 10‐year meadow field trial. J Appl Ecol 39:279–293
Sojneková M, Chytrý M (2015) From arable land to species-rich semi-natural grasslands: succession in abandoned fields in a dry region of central Europe. Ecol Engin 77:373–381
StatSoft, Inc (2012) Statistica for Windows. StatSoft, Tulsa, USA
Storkey J, Meyer S, Still KS, Leuschner C (2012) The impact of agricultural intensification and land‐use change on the European arable flora. Proc Roy Soc London, Ser B, Biol Sci 279:1–9
Stroh PA, Hughes FM, Sparks TH, Mountford JO (2012) The influence of time on the soil seed bank and vegetation across a landscape‐scale wetland restoration project. Restorat Ecol 20:103–112
Suleiman MK, Bhatt A, Jacob S, Thomas RR, Sivadasan MT (2023) Seed longevity in desert species and the possibility of forming a persistent soil seed bank. Sustainability 15:15904
ter Braak CJF, Šmilauer P (2012) Canoco reference manual and users guide: software for ordination (version 5.0). Microcomputer Power, Ithaca
ter Heerdt GNJ, Verweij GL, Bekker RM, Bakker JP (1996) An improved method for seed-bank analysis: seedling emergence after removing the soil by sieving. Funct Ecol 10:144–151
Thompson K, Bakker JP, Bekker RM (1997) The soil seed banks of North West Europe: methodology, density and longevity. Cambridge University Press, Cambridge, UK
Thompson K, Grime JP (1979) Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. J Ecol 893–921
Tichý L (2002) JUICE, software for vegetation classification. J Veg Sci 13:451–453
Török P, Vida E, Deák B, Lengyel S, Tóthmérész B (2011) Grassland restoration on former croplands in Europe: an assessment of applicability of techniques and costs. Biodivers & Conservation 20:2311–2332
Török P, Helm A, Kiehl K, Buisson E, Valkó O (2018) Beyond the species pool: Modification of species dispersal, establishment and assembly by habitat restoration. Restorat Ecol 26(Suppl. 2):S65–S72
Touzard B, Amiaud B, Langlois E, Lemauviel S, Clément B (2002) The relationships between soil seed bank, aboveground vegetation and disturbances in an eutrophic alluvial wetland of Western France. Flora-Morphology, Distribution. Funct Ecol Pl 197:175–185
Tscharntke T, Batáry P, Dormann CF (2011) Set-aside management: How do succession, sowing patterns and landscape context affect biodiversity? Agric Eco-Syst Environm 143:37–44
Tsiafouli MA, Thébault E, Sgardelis SP, Ruiter PC, Putten WH, Birkhofer K, Hemerik L, Vries FT, Bardgett RD, Brady MV, Bjornlund L, Jørgensen HB, Christensen S, Hertefeldt TD, Hotes S, Gera Hol W, Frouz J, Liiri M, Mortimer SR, Setälä H, Tzanopoulos J, Uteseny K, Pižl V, Stary J, Wolters V, Hedlund K (2015) Intensive agriculture reduces soil biodiversity across Europe. Global Change Biol 21:973–985
Turnbull L A, Crawley M J, Rees M (2000). Are plant populations seed‐limited? A review of seed sowing experiments. Oikos, 88:225–238
Tyler T, Herbertsson L, Olofsson J, Olsson PA (2021) Ecological indicator and traits values for Swedish vascular plants. Ecol Indicators 120:106923
Valkó O, Török P, Tóthmérész B, Matus G (2011) Restoration potential in seed banks of acidic fen and dry-mesophilous meadows: Can restoration be based on local seed banks? Restorat Ecol 19:9–15
Valkó O, Deák B, Török P, Tóth K, Kiss R, Kelemen A, Miglécz T, Sonkoly J, Tóthmérész B (2021) Dynamics in vegetation and seed bank composition highlight the importance of post‐restoration management in sown grasslands. Restorat Ecol 29:e13192
van Klink R, Boch S, Buri P, Rieder NS, Humbert JY, Arlettaz R (2017) No detrimental effects of delayed mowing or uncut grass refuges on plant and bryophyte community structure and phytomass production in low-intensity hay meadows. Basic Appl Ecol 20:1–9
von Blanckenhagen B, Poschlod P (2005) Restoration of calcareous grasslands: the role of the soil seed bank and seed dispersal for recolonisation processes. Biotechnol Agron Soc Environm 9:143–149
Vymyslický T, Jiroušek M, Frei I, Smetanová S, Winkler J, Zdražílková M, Fabšičová M (2024) Will small-scale ploughing increase the plant diversity of the species-poor grasslands? Folia Geobot Available at https://doi.org/10.1007/s12224-024-09448-7
Wagner M, Poschlod P, Setchfield RP (2003) Soil seed bank in managed and abandoned semi-natural meadows in Soomaa National Park, Estonia. Ann Bot Fenn 40:87–100
Wagner M, Walker KJ, Pywell RF (2018) Seed bank dynamics in restored grassland following the sowing of high‐and low‐diversity seed mixtures. Restorat Ecol 26:S189–S199
Waldhardt K, Otte A (2001) The significance of the seed bank as a potential for the reestablishment of arable-land vegetation in a marginal agricultural landscape. Web Ecol 2:83–87
Wang Y, Jiang D, Toshio O, Zhou Q (2013) Recent advances in soil seed bank research. Contemp Probl Ecol 6:520–524
Warr SJ, Thompson K, Kent M (2016) Seed banks as a neglected area of biogeographic research: a review of literature and sampling techniques. Progr Phys Geogr 17:329–347
Watson JE, Dudley N, Segan DB, Hockings M (2014) The performance and potential of protected areas. Nature 515:67–73
Wellstein C, Otte A, Waldhardt R (2007) Seed bank diversity in mesic grasslands in relation to vegetation type, management and site conditions. J Veg Sci 18:153–162
White S, Bork E, Karst J, Cahill J (2012) Similarity between grassland vegetation and seed bank shifts with altered precipitation and clipping, but not warming. Community Ecol 13:129–136
Willems JH, Bik LPM (1998) Restoration of high species density in calcareous grassland: the role of seed rain and soil seed bank. Appl Veg Sci 1:91–100
Acknowledgements
We thank Dušan Lekeš (Institute of Botany of the Czech Academy of Sciences) for field assistance, for arranging and conducting laboratory analyses, and for his technical support. Our thanks belong also to Lenka Reiterová from the Podyjí National Park administration for her support and administrative assistance. Many thanks go to everyone that helped us collect and process field data and recorded and identified plant seedlings during greenhouse cultivation – technical assistants from the company Agricultural Research in Troubsko, Martina Entová, Jana Posslová, Pavlína Kadaňková and many others. The research was supported by the Technological Agency of the Czech Republic (project TH04030244) and was also supported by the Ministry of Agriculture of the Czech Republic within the framework of the Long-Term Conception of the Development of the Research Organization Agricultural Research, Ltd. Troubsko and the long-term research development project of the Institute of Botany of the Czech Academy of Sciences RVO 67985939.
Funding
Open access publishing supported by the National Technical Library in Prague.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
All authors of this paper declare that they do not have any competing interests that could be directly or indirectly related to the work submitted for publication. The authors have no relevant financial or non-financial interests to disclose. MF is an employee of the Institute of Botany of the Czech Academy of Sciences, which is one of the co-publishers of the journal Folia Geobotanica.
All the research and the experiments were performed in compliance with the applicable laws of the Czech Republic.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fabšičová, M., Vymyslický, T., Frei, I. et al. The importance of soil seed banks for biodiversity restoration in degraded grasslands. Folia Geobot 59, 17–37 (2024). https://doi.org/10.1007/s12224-024-09452-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12224-024-09452-x