
Abstract
The Balearic Islands occupy a central space in the western Mediterranean, at the maritime crossroads between North Africa, the Iberian Peninsula and the rest of southwestern Europe. As such, it is well placed to investigate changes in subsistence practices associated with the major cultural transitions following the arrival of Islamic rule. Stable carbon and nitrogen isotope analysis was carried out on bone collagen from the Islamic cemetery (ca. AD 950–1150) population excavated at 33 Bartomeu Vicent Ramon, Ibiza, including human (n = 42) and faunal remains (n = 3). Stable oxygen and carbon isotope analysis was also undertaken on human tooth enamel carbonate (n = 6), and six humans were directly radiocarbon dated, confirming the presence of two distinct burial phases. The collagen results emphasise a C3-based diet, with variable but generally minor contributions from marine and/or C4 foods. However, the enamel carbonate results indicate a far greater importance of C4 crops than suggested by the collagen results, contributing up to 40% of energy intake. In keeping with previous studies of the region and period, the dietary contribution of marine protein is probably limited. A small number of outliers in both collagen and carbonate isotope results are identified, suggesting the presence of individuals originating elsewhere. The results are compared with those from previous investigations on the Balearics and the Spanish mainland, highlighting the complexity of factors—both cultural and methodological—affecting inter-regional dietary investigation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Located off the eastern shore of the Iberian Peninsula, the Balearic Islands provide a window onto a host of political, cultural, social and economic changes taking place in medieval southwest Europe, primarily as a result of the shifting dominance of Islamic and Christian social, political, and cultural influences. Islamic influence on the Balearics began in the seventh century AD, but was greatly intensified when the islands were formally brought into the Umayyad by ‘Isam al-Khawlâni in AD 902. This was followed by an influx of settlers mainly of Berber origin from the mainland (al-Andalus) who would have encountered a sparsely populated island (Barceló 1997; Ferrer Abárzuza, 2014: 47). This no doubt facilitated the rapid colonisation of the entire island over only a few decades (Barceló 2005: 30), affecting both the countryside (Kirchner 2006: 128–132) and the town or madina. The settlers brought with them new foodstuffs, among which were 13C-enriched C4 plants such as sorghum and possibly sugarcane, as well as advanced irrigation systems that enabled the cultivation of crops on a scale far beyond that seen on the islands previously (Kirchner 2002, 2006, 2009; Kirchner and Retamero 2015; Retamero 2008; Watson 1983). The town of Eivissa at the beginning of the tenth century would have been a simple castle (Ramon 2000: 141; Marlière and Torres Costa in press), which then expanded into a more substantial urban centre in the twelfth century (Ferrer Abárzuza, 2014: 76). This new incoming population brought changes in agricultural technology, cuisine, religion, social structure and language (Glick 2005; Kirchner and Retamero 2015; Waines 1992; Watson 1983; Zaouali 2007). The Islamic period ended with the islands’ conquest under the Catholic monarch James I of Aragón in 1229–1235 (Glick 2005), with Ibiza itself being invaded in 1235 in an expedition led by Guillem de Montgrí, archbishop of Tarragona (Abulafia 1994: 43).
In this paper, light stable isotope analyses of bone collagen (δ13C, δ15N) and tooth enamel (δ13Cap, δ18Oc) are used to investigate diet and mobility at the Islamic necropolis recently excavated at the site 33 Bartomeu Vicent Ramon (33BVR), in the city of Eivissa, Ibiza (Fig. 1). We explore the impact of socio-political, cultural and economic changes on diet in relation to both earlier and later periods on the Balearic Islands as well as on the adjacent Spanish mainland. The excavated area of the necropolis appears to span the late tenth to mid-twelfth centuries, and so relates to one of the most dynamic and critical periods of the Spanish Middle Ages (Glick 2005: 37; Kaegi 2010).

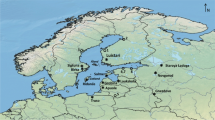
Map of the Balearic Islands and adjacent Spanish mainland with inset view of the Pityusic Islands (Ibiza and Formentera) showing locations of sites mentioned in the text
The two interconnected research questions posed here relate firstly to the relationship between subsistence practices on the Balearic Islands, particularly Ibiza, and those of the mainland, and secondly to diachronic dietary trends in relation to the substantial wider social, cultural and political changes of the period. Given their common origins, it is expected that the medieval Islamic diets of the Balearic Islands were broadly comparable to those of mainland al-Andalus. Nevertheless, the insular position of the Balearic Islands, with easy access to marine resources, suggests the possibility of a distinct subsistence economy from that of the adjacent mainland regardless of period. In addition, the islands’ port towns in particular might be expected to have had higher numbers of immigrants at any given time, some of whom would have died and been buried on the islands. Our study builds on previous work on both themes, which has increased substantially in recent years, allowing these wider comparisons to be made (Alexander et al. 2015; Fuller et al. 2010; Nehlich et al. 2012; Pickard et al. 2017; Salazar-García 2011; Salazar-García et al. 2014; van Strydonck et al. 2002, 2005; Ziriax 2017).
Bartomeu Vicent Ramon, Ibiza
In advance of new construction, an area of 240 m2 was excavated at the former Sindicatos building, 33 Bartomeu Vicent Ramon, Eivissa in 2015 (Marí et al. 2016). In total, 125 burials were excavated from a necropolis that saw use between the tenth and twelfth centuries (Fig. 2). As dictated by Islamic burial practices, bodies were laid on their right sides, facing east, with no accompanying grave goods (Fig. 3). No directly associated settlement has been discovered. The current site was part of a larger cemetery or maqbara, which would also encompass a number of other excavated sites in the vicinity such as those in Avenida España 3 (Kranioti et al. 2015; Girdwood et al. 2015; Martín Parrilla and Graziani Echávarri 2009) and that at the site of en Hort des Pilarets (Roig 2015; Marlière and Torres in press). Full osteological analysis of the skeletal assemblage is still underway.

Plan of the archaeological excavations at 33 Bartomeu Vicent Ramon. Adapted from Marí et al. (2016)

Grave S101 at 33 Bartomeu Vicent Ramon, showing typical Islamic burial position, lying supine on the right side and facing east towards Mecca
Diet in early medieval Spain
Foodways are embedded in culture and in politics and can be used to mark distinct identities (Counihan 1999; Goody 1982; Watson and Caldwell 2004). These can be especially powerful when culture and politics interact with religion, as was the case in medieval Spain (Zaouali 2007). The variety of foodstuffs available in medieval Spain presents a challenge for isotopic studies of diet, which, as discussed below, are limited to distinguishing certain broad food categories, and even then are prone to issues of equifinality in interpretation. Other lines of evidence are therefore important and include archaeobotanical and zooarchaeological findings, as well as early historical documents. These sources are not without their own biases—for example, while providing invaluable detail, the available medieval texts frequently focus on the elite, on urban settings and on special aspects such as religious restrictions rather than on the social, cultural and economic realities of day-to-day subsistence practices (Bolens 1978; Constable 2013; Grewe 1981; Zaouali 2007)—but used in combination with the isotopic evidence they can provide far greater insights than each on its own. The following section very briefly summarises some of this evidence.
Both Muslims and Christians valued wheat (Triticum spp.) highest among the cereals, generally followed by barley (Hordeum vulgare) and rice (Oryza sativa). The importance of wheat and hulled barley is made clear by their dominance in the archaeobotanical record of the period, appearing at numerous sites regardless of their religious or socioeconomic status (Alonso 2005; Alonso et al. 2014; Peña-Chocarro et al. 2018). Less desirable cereals included rye (Secale cereale) and the C4 crops millet (Panicum miliaceum and Setaria italica) and sorghum (Sorghum bicolor) (Alonso 2005; Alòs et al. 2007; de Castro 1993; García Marsilla 1993; García-Sánchez 1995, 1996, 2002, Moreno-Larrazabal et al. 2015; Rubio 1995; Tomás 2009; Watson 1983). These may have seen greater use by the poor, and as livestock fodder and poultry feed. Pulses, including peas (Pisum sativum), broad beans (Vicia faba), lentils (Lens culinaris) and chickpeas (Cicer arietinum) are present but generally do not feature strongly in the medieval period, though they may have been an important source of protein on occasion for the less well off and again were also used as animal fodder (García-Sánchez 2002). Olives (Olea europaea) also made an important dietary contribution, especially in the form of oil, and nuts and fruits were widely consumed, though featuring more strongly in Islamic cuisine. While there is some debate over the details, a number of new crops are thought to have been introduced to the Iberian Peninsula in the Islamic period (Watson 1983). Millets, sometimes considered as among these introductions, were already present since at least the Bronze Age, and feature regularly in Roman period sites (Peña-Chocarro et al. 2018). Sorghum and rice, along with sugarcane (Saccharum officinarum), on the other hand, were introduced in the Islamic period, and certainly feature in the historical documentation (Glick 2005). Perhaps surprisingly, then, no remains of either sorghum or rice have yet been identified in archaeobotantical assemblages from the Iberian Peninsula, though sorghum has been found in medieval sites in southern France (Peña-Chocarro et al. 2018).
Religious and cultural food preferences and prohibitions are often more pronounced in animal than in plant foods. Lamb, kid and chicken were the most highly regarded meats in medieval al-Andalus; wild game was consumed where available, and sheep and goat milk and cheese were common fare (de Castro 1993; Gallart et al. 2005; Garcia et al. 2004; Salas-Salvadó et al. 2006; Waines 1992). Beef, on the other hand, was more strongly associated with Christian cuisine. Fish were not a high status food, but were frequently consumed along the rivers and coasts, as well as being dried and salted for trade inland, though it is difficult to quantify their importance relative to other foods from either textual or archaeological sources. Nevertheless, seafood may have been more important in Mediterranean Spain than in most other parts of the Islamic world (Zaouali 2007). For Christians, fish consumption had religious associations, given the considerable number of holy days such as Lent and frequent ‘fasting’ periods, meaning abstinence from meat (Grumett and Muers 2010; Tomás 2009). Islam had no such religious prohibitions against meat at any time, though of course it prohibited the eating of pork, which was popular among Christian communities (de Castro 1993; García-Sánchez 1996; Waines 1992). However, the extent to which such religious restrictions and admonitions are adhered to can vary considerably over time and by both household and individual (Grumett and Muers 2010; Tomás 2009).
Stable isotope analysis and past diets
All plants exclusively utilise one of three metabolic pathways in order to fix carbon from atmospheric CO2. The two most common pathways are known as C3 and C4, while the third, Crassulacean Acid Metabolism (CAM), is relatively rare and not pertinent to this study (Pearcy and Ehleringer 1984). Both C3 and C4 plants are 13C-depleted relative to the international standard VPDB (defined as 0‰), but C4 plants discriminate less against the heavier 13C isotope and so the two pathways can be easily distinguished. Preindustrial C3 plants in southern Europe will average approximately – 25‰, while C4 plants will average approximately – 11‰. Thus, stable carbon isotope ratios (δ13C) can be used to identify and semi-quantify the dietary contributions of C3 and C4 foods (Lee-Thorp 2008; Schwarz and Schoeninger 1991). In Europe, the vast majority of both domestic and wild plants consumed make use of the C3 photosynthetic pathway. Thus, the introduction of C4 crops, whether consumed directly or indirectly via animal tissues and secondary products (i.e., dairy), will lead to a marked shift to more 13C-enriched values in human consumers. Marine organisms will be intermediate between the two main terrestrial photosynthetic pathways. Thus, it is not always possible to distinguish between marine and C4 contributions to diet using δ13C alone. This is where stable nitrogen isotopes (δ15N) have an important role.
Each trophic level in the food chain is associated with an increase in δ15N ratios of 3–5‰ (Ambrose 1991; Bocherens and Drucker 2003; DeNiro and Epstein 1981; Schoeninger and DeNiro 1984; Sponheimer et al. 2003; O’Connell et al. (2012) have suggested this could be as high as 6‰ in humans). Being longer and more complex than terrestrial food chains, aquatic food chains have the potential for many more trophic levels, and so can impart higher δ15N values to human consumers. This is one means of differentiating C4 and marine protein consumption. Interpretation is complicated, however, by the fact that manuring can increase δ15N values in cereals by 2.6 to 8.0‰, depending on its intensity (Bogaard et al. 2007). Countering this, the consumption of pulses can significantly lower δ15N values. These nitrogen-fixing plants have a symbiotic relationship with denitrifying rhizobacteria, fixing nitrogen (N2) directly from the atmosphere with little or no fractionation so that their δ15N values that closely resemble those of the standard AIR, i.e., near 0‰ (Fraser et al. 2011; Virginia and Delwiche 1982).
As nitrogen is only found in protein, it only informs on that dietary component. Stable carbon isotope measurements on bone collagen are biased towards dietary protein, to a degree that will depend mainly on the quantity and quality of protein in the diet (Ambrose and Norr 1993; Jim et al. 2006). The lower these are, the greater will be the dietary contribution of carbohydrates and lipids to the de novo synthesis of non-essential amino acids. Measurements made on the mineral component, however, reflect the proportional contributions of all three macronutrients (Ambrose and Norr 1993). However, this fraction is less frequently used in archaeological studies because of its greater susceptibility to diagenetic alteration in the burial environment. Enamel is much more resistant than bone or dentine in this respect, and so offers more reliable results in most circumstances (Wang and Cerling 1994). There is some debate concerning whether δ13Cap measurements in enamel and bone carbonate (bioapatite) are directly comparable, with some experiments suggesting an offset of ca. + 2.3‰ for enamel compared to bone (Warinner and Tuross 2009). Other studies, however, have not found this offset (Loftus and Sealy 2012), nor is there any theoretical reason that the two tissues should differ in this respect (Lee-Thorp et al. 1989), and so its applicability remains uncertain.
Stable oxygen isotope (δ18O) analysis has been used extensively to study mobility in both fauna and humans. The lighter isotope (16O) is preferentially evaporated from seawater whilst the heavier isotope (18O) is preferentially removed during precipitation, causing δ18O values to become progressively more negative with distance from the source (Darling et al. 2003; Körner et al. 1991). Because δ18O in drinking water is strongly related to local precipitation, measurements made on humans can be compared against environmental precipitation and drinking water data in order to determine whether or not they are consistent with a local origin (Darling et al. 2003). As well as drinking water, oxygen in food will also contribute to δ18O. For humans, cultural practices such as brewing and stewing can further complicate matters, causing moderate to strong 18O-enrichment (Bretell et al. 2012).
Analysis of δ18O is usually carried out on tooth enamel, again, because of its greater resistance to diagenesis. Enamel hydroxyapatite forms from infancy to adolescence, becoming biologically inert after formation, and so its isotopic composition does not alter once formed. The timing of mineralisation varies depending on the tooth: the first molar crown begins to form in utero and is complete around age 3, the second molar forms between 2.5 and 8 years, and the third molar forms between 8 and 14 years (Hillson 2005).
Materials and methods
Rib samples from 46 individuals from 33BVR were selected for analysis of δ13C and δ15N. Included are adults (age > 20 years), juveniles (age 13–20), children (age 2–13) and infants (age < 2). Infants and young children will be subject to a ‘nursing effect’ resulting in higher δ15N values than the other age categories (Fogel et al. 1989, Fuller et al. 2006). Enamel from the permanent molars of six individuals was analysed for carbonate δ13Cap and δ18Oc. Since the excavated area was a necropolis, faunal remains were scarce, and are limited here to a single cow, bird (probable chicken) and ovicaprid. These samples were combined with faunal data from the wider region to provide a baseline for comparison with the human results. Six individuals were selected for 14C AMS dating, to assess the absolute difference between phase 1 and phase 2 graves as determined stratigraphically, and whether those individuals most enriched in 13C were contemporary with the others.
Collagen extraction from the 46 human and three faunal samples was carried out following a modified Longin method (Richards and Hedges 1999). Briefly, samples were cleaned using a shot blaster before being weighed out (100 mg) and demineralised in 10 ml of 0.5 M HCl at 5 °C over the course of 2 to 5 days. They were then repeatedly rinsed in milliQ water before being transferred into sealed tubes containing a pH 3 HCl solution at 75 °C for 48 h. The gelatinised sample solution was collected using 9-μm EZEE filters (Elkay, UK). Samples were freeze-dried and approximately 1 mg of the resulting ‘collagen’ was placed into tin capsules. Measurements were undertaken in a Europa 20/20 continuous flow isotope ratio mass spectrometer. Alanine was used to control for machine drift, and two internal collagen standards—cow and seal—were used to provide a two-point calibration of the results (see Coplen et al. 2006), referenced to the international standards VPDB and AIR for δ13C and δ15N, respectively. Samples were measured in duplicate and averaged. Measurement precision is on the order of ± 0.2‰ for both δ13C and δ15N. Collagen quality is assessed through collagen yield and C:N ratios (Ambrose 1990, DeNiro 1985; van Klinken 1999). While a C:N range of 2.9–3.6 is often used in stable isotope studies (DeNiro 1985), we use a slightly more restricted range of 2.9–3.5, since it was observed that those samples with ratios of 3.6 or more often differed by > 0.5‰ between duplicate measurements in one or other isotope.
Enamel from first (BVR162), second (BVR104, BVR155, BVR157, BVR161) and third (BVR150) molars was sampled for δ13Cap and δ18Oc analysis. Second molars were preferred, though one third and one first molar were also included, the latter of which could be subject to a small nursing enrichment (White et al. 2004; Wright and Schwarcz 1998; though see Murphy et al. 2007). Enamel was removed using a tungsten drill bit across the entire length of each sample (from crown to root) to provide a long-term average signal. The samples were treated with 2–3% NaOH for 24 h to remove contaminants, before the addition of 0.1 M acetic acid for 4 h to remove exogenous carbonate (cf. Balasse 2002). Powder weighing 600–750 μg was then reacted with 100% phosphoric acid at 90 °C in a VG Isogas Prism II mass spectrometer. Both δ18Oc and δ13Cap are reported relative to the international VPDB standard.
Radiocarbon dating was carried out at the Oxford Radiocarbon Accelerator Unit (ORAU), following the protocols in Brock et al. (2010), using the same collagen prepared for stable isotope analysis (note that this did not include the use of an ultrafiltration step). A Bayesian model is employed to interpret the results making use of the stratigraphic relationships found between some of the graves (Bronk Ramsey 2009).
Data were assessed for normality using Shapiro-Wilk tests and then subjected to parametric (Student’s t test) or non-parametric (Mann-Whitney U test) statistical tests as appropriate. Tests were undertaken in SPSS and StatPlus. A Bonferroni post-hoc correction to the standard critical p level (α = 0.05) is applied in cases where multiple simultaneous comparisons are made (α′ = α/k, where k = the number of groups being compared).
Results
Stable isotopes
Four human samples failed to meet the C:N criterion (2.9–3.5) for well-preserved collagen, leaving 42 individuals (Table 1; Fig. 4). The three faunal samples yielded acceptable collagen. The overall human results exhibit mean values of − 18.6 ± 0.8 and 10.8 ± 0.8‰ for δ13C and δ15N, respectively (Table 2). There are no significant differences between the adult/juvenile males and females in either δ13C (Mann-Whitney U test, Z = 1.64, p = 0.244), or δ15N (Student’s t test, t = 0.202; p = 0.842). While there is an impression that phase 2 contains more individuals slightly enriched in 13C than phase 1, this is not statistically significant (δ13C: Mann-Whitney, Z = 1.25; p = 0.126; δ15N: heteroscedastic Student’s t test, t = 0.912; p = 0.404). As would be expected as a result of the nursing effect, infants and young children (ranging from 1 to 2 to 3.5–4.5 years of age) exhibit significantly higher δ15N values than the other combined ages (Mann-Whitney U test, Z = 2.10; p = 0.036). This is even clearer if sample BVR116 is omitted, with its anomalously low result of 9.8‰, compared to the mean of 12.3 ± 1.0‰ for the remaining four infants. There is no corresponding difference in δ13C.

Bone collagen δ13C and δ15N results from 33BVR by phase. The regression line (r2 = 0.198, p = 0.006, n = 37) refers to adult/juveniles only and so excludes two infants with high δ15N values as well as three other children age ca. 3 years
The δ13C ratios of the single cattle (− 21.7‰) and ovicaprid (− 20.4‰) samples indicate entirely C3 diets, while that of the single bird (chicken?) is slightly elevated (− 18.9‰), suggest some contribution of C4 grain and/or marine foods. Its relatively high δ15N value (9.7‰) might be interpreted as supporting the latter, but birds can have a highly varied diet including 15N-enriched insects, so that this cannot be assumed.
The δ18OcVPBD results average − 5.7 ± 1.3‰. Measurements on carbonate relate in a predictable way to those made on phosphate and can be converted to facilitate comparison with other studies (e.g., Guede et al. 2017), and both can be converted to the expected values for ingested water (Chenery et al. 2012), although each conversion introduces additional uncertainty (Pollard et al. 2011) (Table 3). The δ13Cap carbonate results average − 9.6 ± 1.6‰. Using a diet-consumer fractionation of ca. 9.5‰ for bone (Ambrose and Norr 1993) would equate with an average bulk dietary intake (i.e., including protein, carbohydrate and lipid) value of ca. − 19.1‰, significantly elevated above the expected value of ca. − 25‰ for a purely C3-based diet. An adjustment of ca. 2.3‰ for the enamel-bone apatite difference suggested by Warinner and Tuross (2009) would reduce this estimated diet value to − 21.4‰, though as noted above the need for this step is not yet clear.
Radiocarbon dating
The six radiocarbon dates fall as expected within the tenth to twelfth centuries (Table 4; Fig. 5). As there is limited evidence for the contribution of marine foods (see below), it is unlikely that there is a significant marine reservoir effect to take into account. A Bayesian model placing the four dated graves assigned to phases 1 and 2 in a contiguous sequence suggests that the phases are separated by an interval of 0 to 80 years (95.4% confidence). Phase 1 may include individuals from the first or second generation after the conquest of 902. The remaining two 14C determinations cannot be assigned stratigraphically, but their results are more consistent with Phase 2. There are two few dates to usefully discuss the overall modelled start and end date ranges.

Modelled radiocarbon dates (OxCal v4.3.2)
Discussion
Diet and mobility at 33 Bartomeu Vicent Ramon
The δ13Ccoll mean of − 18.6 ± 0.8 at 33BVR suggests a diet dominated by terrestrial C3 protein with a variable but generally minor contribution of C4 and/or marine foods. A common means of deciding between the latter two alternatives is to determine whether or not there is a significant positive correlation between δ13C and δ15N, since, as noted above, trophic levels will be substantially higher in most marine systems compared to terrestrial systems (Richards and Hedges 1999). This is particularly so, since the isotopic data for fauna from the region (see below) suggest that domestic stock generally were not being fed C4 crops or waste, implying that any C4 crops would have been directly consumed by humans. There is a weak but significant positive correlation (r2 = 0.198; p = 0.006) between δ13C and δ15N, suggesting that marine foods may have featured in the diets of some individuals (Fig. 3). The weakness of this relationship itself suggests that C4 crops are a likely confounding factor.
To explore diets at 33BVR further, it is necessary to compare the human results with a faunal baseline. Given the small number of faunal samples available from the site itself, we draw upon previously published faunal data from the wider region (Table 5). We recognise that this is not ideal, as both spatial and temporal differences in environments and in animal management practices can affect stable isotope results. Nevertheless, this does at least provide a starting point for comparison. The mean δ13C value of − 19.6 ± 1.3‰ for the combined herbivores clearly indicates that these animals were predominately C3 consumers. Given that a significant contribution of C4 plants is only evident in two cattle and two poultry (Gallus) samples (Alexander et al. 2015), we can safely assume that any elevated 13C ratios measured in human tissues indicate either direct consumption of C4 foods or marine foods.
The apparently minimal contribution of marine foods raises a question regarding the mean δ15N value of 10.7 ± 0.6‰ for post-weaning age individuals at 33BVR, as this is ca. 5‰ higher than ovicaprids, averaging 5.5 ± 2.0‰. While falling within the upper range of the expected trophic level shift of 3–5‰, this would require that all protein consumed was animal-based, which is problematic given the known contribution of both cereals and 15N-depleted legumes to medieval Mediterranean diets. A number of possible explanations can be considered. Firstly, high-trophic-level foods may not be adequately represented in the limited faunal database. The minor contribution of marine protein could be emphasised in δ15N over δ13Ccoll, recalling that the latter will to some degree reflect carbohydrates and lipids as well as proteins (cf. Craig et al. 2009). Ovicaprids are the most important animals in Islamic period diets, but they show a wide range of δ15N values, and would be highest for the culturally favoured suckling lamb and kid due to the nursing effect. Surprisingly, pigs are only slightly higher in δ15N than ovicaprids, and in any case would not be expected to feature significantly in Islamic communities due to strong religious proscriptions, whether on Ibiza or on the mainland (Alexander et al. 2015; Antunes 1996; Benito 2006; Davis 2006; Lentacker and Ervynck 1999; Morales et al. 1988). Another potentially important food source is poultry (both meat and eggs), which exhibit substantially higher δ15N values that all the herbivores, averaging 8.7 ± 0.9‰ (Table 5). Secondly, it may be that the trophic level shift for humans is nearer to 6‰ (O’Connell et al. 2012), though even this would reduce rather than eliminate the problem. Finally, the manuring of arable fields would lead to 15N- enriched crops that might be indistinguishable from animal protein, obviating the problem of the unreasonably high contribution of animal protein required to account for the human δ15N results (Alexander et al. 2015; Bogaard et al. 2007). Any combination of these factors could account for the moderately high elevation in human δ15N values relative to the available faunal data.
Given its central importance to our study, the potential contribution of C4 foods requires additional discussion. Using human consumer endpoints of − 19.5 and − 6‰ for C3 and C4 consumers, respectively (based on faunal data from Table 5, and values of ca. 11‰ for millet from Tafuri et al. (2009), plus 5‰ for diet to bone collagen fractionation) and given the overall human mean δ13C value of − 18.8‰, a simple linear extrapolation estimates the average C4 protein contribution to be minimal, on the order of ca. 5%. Some individuals, however, may have made greater use of C4 foods, most notably BVR104, with a δ13Ccoll value of − 15.3‰, more than four standard deviations above the site mean, and suggesting a C4 contribution of ca. 30‰ (if marine foods are not taken into account, which is a plausible scenario given that the δ15N value for this individual is raised above the mean by only 1 SD). Given the limited evidence for 13C-enriched fauna in the study region, we can infer that humans were mainly consuming millet and/or sorghum directly, though generally only in small quantities. It is perhaps surprising that the impact of millet and sorghum is not greater, but this has been noted in other studies focussing on collagen (see below). However, their contribution may be underestimated if the diet contained adequate animal protein, since plant foods contain relatively less and lower quality protein, and so may be underrepresented in isotopic measurements on bone collagen (this is particularly so for sugarcane, with very low crude protein content, though whether this crop was cultivated on Ibiza specifically is not known).
Measurements on enamel carbonate, however, reflect whole diet (Ambrose and Norr 1993), and the average δ13Cap value of − 9.6‰ is consistent with a C4 intake of approximately 42% (based on a simple linear extrapolation between C3 and C4 plant endpoints of − 25 and − 11‰, respectively, and a diet-bioapatite spacing of 9.5‰). Any contribution of fish and shellfish would mean that this estimate is itself too high. Nevertheless, it seems clear that there was in fact a significant component of C4 cereals in the diets of medieval Islamic population on Ibiza. Plotting the δ13Ccoll and δ13Cap results against the model developed by Kellner and Schoeninger (2007) places the 33BVR results near the C3 protein line and intermediate between C3 and C4 energy endpoints, though nearer the former (Fig. 6). The dominance of C3 protein is clear, and will derive from a combination of meat and dairy, as well as eggs, along with C3 cereals. Given the expected predominance of C3 cereals, especially wheat, and the presumed secondary but important contribution of olives, nuts and fruits—all C3 plants— in medieval Islamic diets, the high level of C4 consumption inferred from the enamel measurements is perhaps surprising. Even assuming that there is indeed a bone-enamel offset of + 2.3‰ as proposed by Warinner and Tuross (2009), the C4 energy contribution would still be significant, although it would be reduced to ca. 25%. Further research on this assemblage, with additional enamel samples in particular, may help resolve the matter.

The 33BVR δ13Ccoll and δ13Cap results plotted against Kellner and Schoeninger’s (2007) model to distinguish C3, C4 and marine sources of protein and energy
The average δ18Oc value of − 7.3 ± 1.3‰ can be converted into an estimated δ18OVSMOW value of − 8.9 ± 2.2‰ (Chenery et al. 2012). For comparison, the δ18OVSMOW value for annual precipitation at Ibiza is estimated at ca. − 5.3 ± 1.5‰, a range that is widely shared along the eastern coast of Spain and the north coast of North Africa (http://waterisotopes.org; see also Araguas-Araguasa and Diaz Teijeiro 2005; Evans et al. 2012) and so is of limited resolution in terms of investigating an individual’s origins. Taken at face value, only one individual at 33BVR (BVR155) is consistent with a local origin, with the others all being 18O-depleted, raising the possibility of origins in the mountains of southeast (cf. Guede et al. 2017: Fig. 8) or northern Spain (cf. Prevedorou et al. 2010). A similarly high proportion of immigrants was also inferred from stable sulphur isotope (δ34S) data in a study by Nehlich et al. (2012), in which the authors concluded that 18 of 20 Islamic period individuals analysed from Es Soto were immigrants to Ibiza. This could perhaps be seen in the context of Eivissa’s position as the major port of the island. But there are numerous complications with making direct comparisons between δ18O in enamel and precipitation, leading many researchers to avoid this approach altogether (Bell et al. 2010; Pellegrini et al. 2016; Pollard et al. 2011). Alternatives, however, rely on the presence of larger datasets than we have at present for the region.
A comparison of the δ18Oc and δ13Cap results offers another way forward. Interestingly, the two isotopes are strongly correlated (r2 = 0.844, p = 0.01, df = 5) (Fig. 7), suggesting the possibility of varying childhood origins. Those enriched in both 18O and 13C (BVR155, BVR104) would be consistent with the greater consumption of C4 foods in more arid environments, such as those of the arid coastal region of southeast Spain. Individual BVR155’s second molar yielded a δ18Oc value of − 3.7‰. While their δ13Ccoll value of − 18.9‰ is not significantly raised above the 33BVR site average, the δ13Cap result of − 7.3‰ is the highest of the six individuals sampled, suggesting a more substantial dietary contribution of C4 foods in childhood, with the subsequent adoption of the Balearic Island diet being reflected in the bone collagen values.

Plot of δ18Oc and δ13Cap results on human tooth enamel from 33BVR, showing a strong positive correlation between the two isotopes (r2 = 0.844, p = 0.01, df = 5)
Bartomeu Vicent Ramon in its temporal and regional context
One of the questions posed in this paper is the impact of Islamic religion, cultural preferences and agricultural practices on Balearic foodways. In order to address this, 33BVR needs to be seen in the context of what went before and what came after (Table 6; Fig. 8). In addition, there is the matter of whether or not diets differed on the islands versus the adjacent mainland, either because of easier access to marine foods, and/or because of the differing possibilities for farming and herding. A caveat when comparing isotopic data produced in various laboratories with varying sample preparation protocols and instrumentation is that they may be subject to small inter-laboratory differences (Pestle et al. 2014). Any larger differences observed, however, should be robust, as should even small differences found within single studies.

Human δ13C and δ15N values (\( \overline{\mathrm{X}} \) ± 2SE) for comparative medieval and Islamic sites (infants excluded)
The differences between Late Antiquity/Early Byzantine and the medieval Islamic period on the Balearic Islands are relatively minor for both isotopes and tend to be site-based rather than temporal: Can Fonoll, for example, stands out from the other sites in having significantly lower means for both isotopes (p < 0.01 in all comparisons, with the exception of no significant difference in δ13C compared to S′Hort des Llimoners). On the face of it, this would suggest that there was no marked increased emphasis on C4 crops following the early tenth century Islamic colonisation of the Balearics. However, as discussed above, δ13C measurements on collagen can underrepresent the contribution of lower protein plant foods, particularly when there is sufficient animal protein in the diet. To resolve this matter will require δ13Cap analysis of enamel from the earlier periods. There is a small but significant average decrease of ca. 0.5‰ in δ15N between Late Antiquity/Early Byzantine (Sanisera and S′Hort des Llimoners, n = 61) and the Islamic period (33BVR, Es Soto and Can Fonoll, n = 170) on the Balearics (Mann-Whitney U test, Z = 3.141, p = 0.002), which could relate to a decline in the consumption of marine foods and/or animal products more generally, to an increase in the consumption of pulses or indeed to a combination of all three. This decrease in the medieval period can be taken as robust, since δ15N values are on average also higher in the Punic period sites on Ibiza (Can Marines, Puig des Molins and Ses Païsses de Cala d’Hort), on par with those in Late Antiquity (Fuller et al. 2010; Salazar-García 2011).
The three medieval Islamic sites on Ibiza are broadly contemporary in the tenth to twelfth/thirteenth centuries and are all located in the southeast of the island. Indeed, Es Soto and 33BVR are both in Eivissa, while Can Fonoll is ca. 5 km to the southwest (Fig. 1). Despite their spatial and temporal proximity, the site means differ slightly but significantly in both δ13C and δ15N for all pairwise comparisons, with the exception of δ15N between 33BVR and Es Soto (Table 7). The implication is one of very small-scale local differences in diets between contemporaneous populations. This is perhaps easiest to understand when comparing the rural site of Can Fonoll with the two Eivissa sites, with the latter having greater access to marine foods and to imported foodstuffs (Pickard et al. 2017), though it was not until the twelfth century that the Balearic Islands became more thoroughly enmeshed in international Mediterranean trade (Doxey 1994). The explanation for this subtle, small-scale patterning may instead relate to the strong, clan-based territorial divisions on Ibiza set out during its re-settlement in 902 (Kirchner 2002, 2006, 2009; Kirchner and Retamero 2015: Fig. 4.7). A number of such territories ran radially through Eivissa and its hinterland, providing access to the port to different clans (Kirchner 2002: 173). It is possible that the various clans emphasised slightly different combinations of crops and animals in their diets, depending on the local soils and access to irrigation, differences in their labour pools and wealth etc. Further exploration of this idea would firstly need to confirm that the small isotopic differences observed are ‘real’ and not an artefact of inter-laboratory measurement variability (Pestle et al. 2014).
Much stronger, and therefore more robust, differences are seen between the Balearic Islands and the adjacent mainland during the medieval period. The mainland sites of El Raval, Gandía-Benipeixcar and Gandía-Colegiata de Santa María are all significantly higher in δ13C, suggesting greater consumption of C4 foods. The contribution of marine foods is made improbable for these sites, despite their coastal locations, by the absence of any positive correlations between δ13C and δ15N results (Table 5). This includes the late medieval Islamic site of El Raval (fourteenth to sixteenth centuries), which exhibits a significantly higher average δ15N value than any of the other sites considered here, but shows no correlation with δ13C values (Salazar-García et al. 2014). Gandía presents an interesting case, as both Islamic (Benipeixcar) and Christian (Colegiata de Santa María) communities were present during the late medieval period (thirteenth to sixteenth centuries), though Benipeixcar was a Muslim enclave on the outskirts of the larger town of Gandía (Alexander et al. 2015). Significantly higher δ13C values from Benipeixcar suggest a greater contribution of C4 foods for the Muslim mudéjares than for the Christian community at Colegiata. Interestingly, this difference appears to have been driven largely or entirely by enriched 13C in Muslim females, with no clear differences seen between Muslim males and Christian males and females (Alexander et al. 2015: Fig. 4). While δ15N values are on average significantly higher at Benipeixcar (10.7 ± 0.6‰) than at Colegiata (10.2 ± 0.8‰), the lack of any correlation with δ13C values again provides no clear support for greater consumption of marine fish by either the Islamic or the Christian communities, despite expectations arising from religious prohibitions for the latter (Tomás 2009).
In contrast to the mainland, some sites on the Balearic Islands show low but statistically significant positive correlations between δ13C and δ15N (Table 5), consistent with some contribution of marine foods in island diets. The two exceptions are the Late Antiquity/Byzantine cemetery at S′Hort des Llimoners and the large Islamic cemetery of Can Fonoll, both on Ibiza (Fuller et al. 2010; Pickard et al. 2017). The site means at Can Fonoll of − 19.0 ± 1.3 and 10.3 ± 0.8‰ for δ13C and δ15N (n = 112), respectively, indicate predominantly C3 diets. Five individuals are identified as outliers in δ13C, with values above − 18‰ (but no corresponding increase in δ15N), the highest being − 8.6‰, suggesting a diet almost entirely comprised of C4 foods and so undoubtedly a recent migrant to Ibiza (Pickard et al. 2017). Removing these outliers, together with an infant, does result in a significant positive correlation between δ13C and δ15N values in the remaining 106 individuals, though it is very low and arguably not meaningful (r2 = 0.056, p = 0.014, df = 105). It should be noted here that the suggestion in Pickard et al. (2017: 4) that Mediterranean fish have similar δ13C and δ15N values to terrestrial fauna, and so cannot be easily distinguished in isotopic studies, is, in the case of the former, based on an inappropriate comparison of modern fish flesh values to archaeological fish bone. Thus, for example, the average δ13C value of − 17.8‰ in the cited study of modern fish off the coast of Ibiza (Polunin et al. 2001; cf. Garcia-Guixé et al. 2010) needs to be adjusted by + 2.9‰ for tissue fractionation (Robson et al. 2012), and by a further + 1.1‰ to account for 13C-depletion of the modern atmosphere and oceans by the burning of fossil fuels, thus raising the expected analogous archaeological bone collagen value to − 13.8‰, entirely compatible with marine bone collagen measurements obtained along the Atlantic façade (cf. Schulting 2018; see also Alexander et al. 2015: Table 2). Stable nitrogen isotope values in the Mediterranean, however, are relatively low.
An issue when investigating the role of marine foods, as discussed above and as also noted by Pickard et al. (2017; cf. Craig et al. 2009), is that their contribution to δ13C in bone collagen can be masked in diets dominated by C3 carbohydrates (wheat, barley, rice) and lipids (e.g. olive oil). The fact that we see positive correlations at some sites suggests that it may still be possible to detect low-level consumption of marine foods in bulk stable isotope measurements of collagen. A question that arises, then, is whether similar levels of marine resource utilisation on mainland sites would be rendered isotopically invisible in terms of δ13C by being swamped by the greater dietary contribution of C4 plants there, which would have the effect of obscuring any correlation with δ15N values. Nevertheless, based on the presently available evidence, the results from 33BVR suggest that marine foods constituted only a minor component to Balearic diets, consistent with the findings of previous isotopic studies (Davis 2002; Fuller et al. 2010; Nehlich et al. 2012; Pickard et al. 2017; Salazar-García 2011; Van Strydonck et al. 2002, 2005; Ziriax 2017).
The three mainland sites all exhibit a higher reliance on C4 crops than the Balearic Islands, though the explanation for this is not yet clear. The sites differ in age, with those on the mainland all being late medieval (thirteenth/fourteenth to sixteenth centuries) while the Balearic sites are earlier (tenth to thirteenth centuries). The decrease in rainfall in the western Mediterranean during the Medieval Climatic Anomaly might be expected to favour millet and sorghum, both drought resistant and with short growing seasons, but this event dates to the eleventh to thirteenth centuries (Roberts et al. 2012), and so largely overlaps both the mainland and insular sites.
To return to the two questions posed at the beginning of this paper, we do not see any clear isotopic impacts following from the Islamic conquest on the Balearic Islands. It may be that this expectation was itself flawed, given that C4 crops were present on the Iberian Peninsula since at least the Middle Bronze Age and would already have been integrated into the farming economy to some extent (Alonso 2008; Buxó and Piqué 2008). If anything, the greatly improved irrigation systems introduced into al-Andalus by Arab and Berber settlers (Kirchner 2002, 2006, 2009; Kirchner and Retamero 2015; Retamero 2008; Watson 1983) may have facilitated a greater emphasis on more highly culturally valued C3 crops, especially wheat. The frequent focus in stable isotope studies on the contributions of C4 (and marine) foods is driven in part by the analytical technique itself, since there is such a clear distinction between the 13C values of C3 and C4 plants, ostensibly making this a very amenable research question. At the same time, it must be recognised that the analysis of δ13C in bone collagen will be biased against C4 crops when there is sufficient C3-based animal protein in the diet. Of course, wheat, barley, rice and legumes are also C3 crops, with the latter higher in protein than cereals, again acting against the detection of low to moderate C4 consumption. That this can be partly overcome by the analysis of δ13Cap in enamel carbonate has been suggested with the preliminary results presented here. This approach needs to be applied more widely, to determine the extent of and variability in the impact of C4 crops across medieval Iberia.
Conclusions
The stable isotope and 14C dating results presented here from the Islamic necropolis of 33 Bartomeu Vicent Ramon add to the growing body of data on diet and mobility on Ibiza and the Balearic Islands, as well as contributing to studies of the wider region. The bone collagen δ13C and δ15N results offer further support for the dominance of C3 plant and animal foods, with a substantial contribution from C4 crops and a minor contribution from marine resources both likely. That the contribution of C4 foods may be underestimated more generally in medieval Spain is suggested by the enamel carbonate δ13C data for six individuals, which to our knowledge represents the first such results for this region. Similar problems may arise in the underestimation of the use of marine foods, when consumed together with large amounts of carbohydrates and/or lipids from cereals and olive oil, respectively. Future research on these matters might make additional use of other isotopes (sulphur, hydrogen, strontium), single amino acid analysis and high-resolution sequential sampling of enamel and dentine in order to obtain more detailed life histories.
While not showing the presence of immigrants as clearly as some previous studies, the collagen δ13C and the enamel δ18Oc and δ13Cap results from 33BVR suggest the presence of both recent and earlier arrivals to the island from regions with a greater emphasis on C4 foods. Based on previously published δ13C results from the mainland around Valencia, and the historically attested origins of the Berber clans as coming primarily from al-Andalus, this region seems plausible as their source. While complications in the interpretation of δ18Oc results were highlighted, specifically in terms of direct comparisons with δ18O in meteoric waters, this avenue of investigation is promising. Despite the comparative homogeneity of precipitation values across much of the coastal western Mediterranean, the variation seen in even the limited number of individuals analysed here suggests that they originated from a number of different locations. This was further confirmed by the strong positive correlation seen with enamel δ13Cap values. The availability of larger datasets both from individual sites and from the wider region would facilitate an alternative approach focused directly on variability in the distribution of results, circumventing problems with comparing δ18O values in enamel and in precipitation.
While the Balearic sites exhibit broadly similar δ13C and δ15N results, suggesting similar diets, there are some significant differences between roughly contemporaneous sites on Ibiza, between pre-Islamic and Islamic sites on Ibiza and between pre-Islamic sites on Minorca and Ibiza. What is less clear is the extent to which these are entirely site-based (or indeed lab-based differences in isotopic measurements) or whether there is a more consistent underlying temporal and/or spatial pattern. A clearer pattern does emerge in a comparison of medieval Islamic sites on Ibiza with those on the adjacent Andalusi mainland, where consistently greater use was made of C4 crops. Why this should be so remains unclear, and could be the focus of a concerted multi-disciplinary study combining isotopic approaches with specialists in the analysis of irrigation systems, medieval farming practices and the socio-political milieu of this dynamic period of Iberian history.
References
Abulafia D (1994) A mediterranean emporium: the catalan kingdom of majorca. Cambridge University Press, Cambridge
Alexander MM, Gerrard CM, Gutiérrez A, Millard A (2015) Diet, society, and economy in late medieval Spain: stable isotope evidence from Muslims and Christians from Gandía, Valencia. Am J Phys Anthropol 156(2):263–273
Alonso N (2005) Agriculture and food from the Roman to the Islamic period in the north-east of the Iberian Peninsula: archaeobotanical studies in the city of Lérida (Catalonia, Spain). Veg Hist Archaeobotany 14(4):341–361
Alonso N (2008) Crops and agriculture during the Iron Age and late antiquity in Cerdanyola del Vallès (Catalonia, Spain). Veg Hist Archaeobotany 17(1):75–84
Alonso N, Antolín F, Kirchner H (2014) Novelties and legacies in crops of the Islamic period in the northeast Iberian peninsula: the archaeobotanical evidence in Madîna Balagî, Madîna Lârida, and Madîna Turṭûsa. Quat Int 346:149–161
Alòs C, Camats A, Monjo M, Solanes E, Alonso N, Martínez J (2007) El Pla d’Almatà (Balaguer, la Noguera): primers aportacions interdisciplinàries a l’estudi de les sitges i els pous negres de lo Zona 5. Revista d’Arqueologia de Ponent 16(17):145–168
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451
Ambrose SH (1991) Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. J Archaeol Sci 18:293–317
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert JB, Grupe G (eds) Prehistoric human bone: archaeology at the molecular level. Springer-Verlag, New York, pp 1–37
Antunes MT (1996) Alimentação de origem animal em regime islâmico: Alcaris longa e casa il de Alcáçova de Mértola. Arqueologia Medieval 4:267–227
Araguas-Araguasa LJ, Diaz Teijeiro MF (2005) Isotope composition of precipitation and water vapour in the Iberian peninsula. First results of the Spanish network of isotopes in precipitation. Isotopic composition of precipitation in the Mediterranean Basin in relation to air circulation patterns and climate. IAEA, Vienna, pp 173–190
Balasse M (2002) Reconstructing dietary and environmental history from enamel isotopic analysis: time resolution of intra-tooth sequential sampling. Int J Osteoarchaeol 12:155–165
Barceló M (1997) El curs de les aigües. Treballs sobre els pagesos de Yabisa (290-633H/902–1235 dC), Eivissa: Qüaderns d’Arqueologia Pitiüsa, no. 3
Barceló M (2005) Negre i roig. Els contextos historiogràfics per a l’estudi de la societat andalusina de Menorca i la seva destrucció. In: Barceló M, Retamero F (eds) Els Barrancs Tancats. L’Ordre Pagès al Sud de Menorca en Época Andalusina (segles X-XIII). Maó: Institut Menorquí d’Estudis
Bell LS, Lee-Thorp JA, Elkerton A (2010) Sailing against the wind: reply to Millard and Schroeder: ‘true British sailors’: a comment on the origin of the men of the Mary Rose. J Archaeol Sci 37(4):683–686
Benito M (2006) Arqueozoología del castillo de Ambra (Pego, Alicante). MARQ Arqueol Museos 1:85–118
Bocherens H, Drucker D (2003) Trophic level isotopic enrichments for carbon and nitrogen in collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13(1–2):46–53
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343
Bolens L (1978) La Révolution agricole andalouse du XIe siècle. Stud Islam (47):121–141
Brettell R, Montgomery J, Evans J (2012) Brewing and stewing: the effect of culturally mediated behaviour on the oxygen isotope composition of ingested fluids and the implications for human provenance. J Anal At Spectrom 27(5):778–785
Brock F, Higham TFG, Ditchfield P, Bronk Ramsey C (2010) Current pretreatment methods for AMS radiocarbon dating at the Oxford Radiocarbon Accelerator Unit (ORAU). Radiocarbon 52(1):102–112
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51(1):337–360
Buxó R, Piqué R (2008) Arqueobotánica. Los Usos de las Plantas en la Península Ibérica. Ariel, Barcelona
de Castro T (1993) La Alimentación en las Crónicas Castellanas Bajomedievales. Universidad de Granada, Granada
Chenery CA, Pashley V, Lamb AL, Sloane HJ, Evans JA (2012) The oxygen isotope relationship between the phosphate and structural carbonate fractions of human bioapatite. Rapid Commun Mass Spectrom 26(3):309–319
Constable OR (2013) Food and meaning: Christian understandings of Muslim food and food ways in Spain, 1250-1550. Viator 44:199–236
Coplen TB, Brand WA, Gehre M, Gröning M, Meijer HJ, Toman B, Verkouteren RM (2006) New guidelines for δ13C measurements. Anal Chem 78:2439–2441
Counihan C (1999) The anthropology of food and body: gender, meaning and power. Routledge, London
Craig OE, Biazzo M, O'Connell TC, Garnsey P, Martinez-Labarga C, Lelli R, Salvadei L, Tartaglia G, Nava A, Renò L, Fiammenghi A, Rickards O, Bondioli L (2009) Stable isotopic evidence for diet at the imperial roman coastal site of Velia (1st and 2nd centuries AD) in southern Italy. Am J Phys Anthropol 139(4):572–583
Darling WG, Bath A, Talbot J (2003) The O and H stable isotope composition of fresh waters in the British Isles. 2. Surface waters and groundwater. Hydrol Earth Syst Sci 7:183–195
Daux V, Lecuyer C, Heran M-A, Amiot R, Simon L, François F, Martineau F, Lynnerup N, Reychler H, Escarguel G (2008) Oxygen isotope fractionation between human phosphate and water revisited. J Hum Evol 55(6): 1138–1147
Davis MHLA (2002) Putting meat on the bone: an investigation into palaeodiet in the Balearic Islands using carbon and nitrogen stable isotope analysis. In: Waldren WH, Ensenyat J (eds) World islands in prehistory: international insular investigations. Archaeopress, British Archaeological Reports, International Series 1095, Oxford, pp 198–216
Davis SJM (2006) Faunal Remains from Alcáçova de Santarém, Portugal. Lisbon: Instituto Português de Arqueologia, Trabalhos De Arqueologia 43
DeNiro MJ (1985) Post-mortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341–351
Doxey G (1994) Diplomacy, trade and war: Muslim Majorca in international politics. J Mediev Hist 20(1):39–61
Evans JA, Chenery C, Montgomery J (2012) A summary of strontium and oxygen isotope variation in archaeological human tooth enamel excavated from Britain. J Anal At Spectrom 27(5):754–764
Ferrer Abárzuza A (2014) Yabisa, Faramantira (s.VIII-1235). Aportació a un estat de la qüestió, Sol erat incipiens per iter conscendere caeli… Jornades d’estudi conmemoratives del 900è aniversari de l’atac pisano-català a Yabisa i Faramantira, Eivissa. Treballs del Museu Arqueològic d’Eivissa i Formentera 71:41–89
Fogel ML, Tuross N, Owsley DW (1989) Nitrogen isotope tracers of human lactation in modern and archeological populations. Annual Report of the Director, Geophysical Laboratory, Carnegie Institute of Washington 89:111–117
Fraser RA, Bogaard A, Heaton THE, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton PR, Sparkes D, Styring AK (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38(10):2790–2804
Fuller BT, Fuller JL, Harris DA, Hedges REM (2006) Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am J Phys Anthropol 129:279–293
Fuller BT, Márquez-Grant N, Richards MP (2010) Investigation of diachronic dietary patterns on the islands of Ibiza and Formentera, Spain: evidence from carbon and nitrogen stable isotope ratio analysis. Am J Phys Anthropol 143(4):512–522
Gallart L, Roberto IE, Maupoei PF (2005) La Salazón de Pescado: una Tradición en la Dieta Mediterránea. Universidad Politécnica de Valencia, Valencia
García Marsilla JV (1993) La Jerarquía de la Mesa. Centre d’Estudis d’Història Local, Valencia
Garcia E, Subirà ME, Richards MP (2004) Régime et société d’après l’analyse des isotopes stables : l’exemple de la population de Can Reinés (Mallorca, Espagne, 600 ap. J.C.). Antropo 7:171–176
Garcia-Guixé E, Eulàlia Subirà M, Marlasca R, Richards M (2010) δ13C and δ15N on recent and ancient fish bones from the Mediterranean Sea. J Nordic Archaeol Sci 17:83–92
García-Sánchez E (1995) La gastronomía andalusí, El zoco: Vida Económica y Artes Tradicionales en al-Andalus y Marruecos. Lunwerg Editores, Barcelona, pp 49–57
García-Sánchez E (1996) La alimentación popular urbana en al-Andalus. Arqueología Medieval 4:219–236
García-Sánchez E (2002) Dietetic aspects of food in al-Andalus. In: Waines D (ed) Patterns of Everyday Life. Routledge, London, pp 275–288
Girdwood LK, Langstaff H, Kranioti EF (2015) Settlement of Ibiza, Spain: From Rome to Islam. The biological evidence from dentition, VI Jornades d’Arqueologia de les Illes Balears. Formentera, pp 323–330
Glick TF (2005) Islamic and Christian Spain in the early middle ages. Brill, Leiden
Goody J (1982) Cooking, cuisine and class: a study in comparative sociology. Cambridge University Press, Cambridge
Grewe R (1981) Catalan cuisine, in an historical perspective. In: Davidson A (ed) National and regional styles of cookery. Oxford University Press, Oxford, pp 170–178
Grumett D, Muers R (2010) Theology on the menu: asceticism, meat and Christian diet. Routledge, London
Guede I, Ortega LA, Zuluaga MC, Alonso-Olazabal A, Murelaga X, Pina M, Gutierrez FJ, Iacumin P (2017) Isotope analyses to explore diet and mobility in a medieval Muslim population at Tauste (NE Spain). PLoS One 12(5):e0176572
Hillson S (2005) Teeth. Cambridge University Press, Cambridge
Jim S, Jones V, Ambrose SH, Evershed RP (2006) Quantifying dietary macronutrient sources of carbon for bone collagen biosynthesis using natural abundance stable carbon isotope analysis. Br J Nutr 95:1055–1062
Kaegi WE (2010) Muslim expansion and byzantine collapse in North Africa. Cambridge University Press, Cambridge
Kellner CM, Schoeninger MJ (2007) A simple carbon isotope model for reconstructing prehistoric human diet. Am J Phys Anthropol 133(4):1112–1127
Kirchner H (2002) El mapa de los asentamientos rurales Andalusíes de la isla de Ibiza. In: Trillo C (ed) Asentamientos Rurales y Territorio en el Mundo Mediterráneo en Época Medieval. Athos Pergamos, Berja, pp 120–186
Kirchner H (2006) Yabisa i l’estudi sobre la migració i colonització berber i àrab de les Illes Balears. Fites 6:39–47
Kirchner H (2009) Original design, tribal management and modifications in medieval hydraulic systems in the Balearic Islands (Spain). World Archaeol 41(1):151–168
Kirchner H, Retamero F (2015) Becoming islanders. Migration and settlement in the Balearic Islands (10th – 13th centuries). In: Retamero F, Schjellerup I, Davies A (eds) Agricultural and pastoral landscapes in pre-industrial society: choices, stability and change. Oxbow, Oxford, pp 57–78
Körner C, Farquhar GD, Wong SC (1991) Carbon isotope discrimination by plants follows latitudinal and altitudinal trends. Oecologia 88(1):30–40
Kranioti EF, García-Donas JG, Langstaff H (2015) Cranial variation of the Ibiza populations, VI Jornades d’Arqueologia de les Illes Balears. Formentera, pp 273-284
Lee-Thorp JA (2008) On isotopes and old bones. Archaeometry 50:925–950
Lee-Thorp JA, Sealy J, van der Merwe NJ (1989) Stable carbon isotope ratio differences between bone collagen and bone apatite, and their relationship to diet. J Archaeol Sci 16:585–599
Lentacker A, Ervynck A (1999) The archaeofauna of the late medieval, Islamic harbour town of Saltés (Huelva, Spain). Archaeofauna 8:141–157
Loftus E, Sealy J (2012) Technical note: interpreting stable carbon isotopes in human tooth enamel: an examination of tissue spacings from South Africa. Am J Phys Anthropol 147:499–507
Marí JJ, Graziani G, García-Rubio A (2016) La maqbara de Yabisa. Fites 16:29–40
Marlière, E. and Torres Costa, J. in press. L’àrea portuària d’Eivissa durant l’antiguitat i l’època medieval. Noves dades arqueològiques de s’Hort des Pilarets i del passeig de Vara de Rey. Eivissa: Quaderns d’Arqueologia Ebusitana, n°4
Martín Parrilla A, Graziani Echávarri G (2009) Un tram de la muralla medieval, el llenç intermedi documentat al carrer Major 16. Quaderns d’Arqueologia Ebusitana 1:39–43
Morales A, Moreno R, Cereijo MA (1988) Calatrava la Vieja: primer informe sobre la fauna de vertebrados recuperada en el yacimiento Almohad. Primera parte: mamíferos Boletín de Arqueología Medieval 2:7–48
Moreno-Larrazabal A, Teira-Brión A, Sopelana-Salcedo I, Arranz-Otaegui A, Zapata L (2015) Ethnobotany of millet cultivation in the north of the Iberian peninsula. Veg Hist Archaeobotany 24:541–554
Murphy BP, Bowman D, Gagan MK (2007) The interactive effect of temperature and humidity on the oxygen isotope composition of kangaroos. Funct Ecol 21(4):757–776
Nehlich O, Fuller BT, Márquez-Grant N, Richards MP (2012) Investigation of diachronic dietary patterns on the islands of Ibiza and Formentera, Spain: evidence from sulfur stable isotope ratio analysis. Am J Phys Anthropol 149(1):115–124
O'Connell TC, Kneale CJ, Tasevska N, Kuhnle GGC (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149(3):426–434
Pearcy RW, Ehleringer JR (1984) Comparative ecophysiology of C3 and C4 plants. Plant Cell Environ 7(1):1–13
Pellegrini M, Pouncett J, Jay M, Parker Pearson M, Richards MP (2016) Tooth enamel oxygen “isoscapes” show a high degree of human mobility in prehistoric Britain. Nature: Sci Rep 6:e34986
Peña-Chocarro L, Perez Jorda G, Alonso N, Antolín F, Teira Brion A, Tereso JP, Montes Moya EM, Reyes DL (2018) Roman and medieval crops in the Iberian Peninsula: a first overview of seeds and fruits from archaeological sites. Quat Int. https://doi.org/10.1016/j.quaint.2017.09.037
Pestle WJ, Crowley BE, Weirauch MT (2014) Quantifying inter-laboratory variability in stable isotope analysis of ancient skeletal remains. PLoS One 9(7):e102844
Pickard C, Girdwood L-K, Kranioti E, Márquez-Grant N, Richards MP, Fuller BT (2017) Isotopic evidence for dietary diversity at the mediaeval Islamic necropolis of can Fonoll (10th to 13th centuries CE), Ibiza, Spain. J Archaeol Sci Rep 13:1–10
Pollard AM, Pellegrini M, Lee-Thorp JA (2011) Some observations on the conversion of dental enamel δ18O(p) values to δ18O(w) to determine human mobility. Am J Phys Anthropol 145:499–504
Polunin NVC, Morales-Nin B, Pawsey WE, Cartes JE, Pinnegar JK, Moranta J (2001) Feeding relationships in Mediterranean bathyal assemblages elucidated by stable nitrogen and carbon isotope data. Mar Ecol Prog Ser 220:13–23
Prevedorou E, Bonilla MD-Z, Romero A, Buikstra JE, Ibáñez MPdM, Knudson KJ (2010) Residential mobility and dental decoration in early medieval Spain: results from the eighth century site of Plaza del Castillo, Pamplona. Dent Anthrop 23(2):42–52
Ramon J (2000) Estudi Arqueològic i Històric del Castell d’Eivissa. I. Estructures i Elements Arquitectònics. Eivissa: Quaderns d’Arqueologia Pitiusa, 6
Reimer PJ, Bard E, Bayliss A, Beck JW, Blackwell PG, Bronk Ramsey C, Buck CE, Cheng H, Edwards RL, Friedrich M, Grootes PM, Guilderson TP, Haflidason H, Hajdas I, Hatté C, Heaton TJ, Hoffmann DL, Hogg AG, Hughen KA, Kaiser KF, Kromer B, Manning SW, Niu M, Reimer RW, Richards DA, Scott EM, Southon JR, Staff, R.A, Turney CSM, van der Plicht J (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55(4):1869–1887
Retamero F (2008) Irrigated agriculture, risk and population. The Andalusi hydraulic systems of the Balearic Islands as a case study (Xth-XIIIth century). In: Compatangelo-Soussignan R, Bertrand J-R, Chapman J, Laffont P-Y (eds) Marqueurs des Paysages et Systèmes Socio-Économiques. Presses Universitaires de Rennes, Renne, pp 135–147
Richards MP, Hedges REM (1999) Stable isotope evidence for similarities in the types of marine foods used by Late Mesolithic humans on the Atlantic coast of Europe. J Archaeol Sci 26:717–722
Roberts N, Moreno A, Valero-Garcés BL, Corella JP, Jones M, Allcock S, Woodbridge J, Morellón M, Luterbacher J, Xoplaki E, Türkeş M (2012) Palaeolimnological evidence for an east–west climate see-saw in the Mediterranean since AD 900. Glob Planet Chang 84–85:23–34
Robson H, Andersen S, Craig OE, Fischer A, Glykou A, Hartz S, Lübke H, Schmölcke U, Heron C (2012) Carbon and nitrogen isotope signals in eel bone collagen from Mesolithic and Neolithic sites in northern Europe. J Archaeol Sci 39:2003–2011
Roig J (2015) Jaciment de l’Hort des Pilarets (Municipi d’Eivissa). La Linde 5
Rubio A (1995) El consumo de pan en la Valencia bajomedieval. Col.loqui d’Història de l’Alimentació a la Corona d’Aragó 1:153–184
Salas-Salvadó J, Huetos-Solano MD, García-Lorda P, Bulló M (2006) Diet and dietetics in al-Andalus. Br J Nutr 96(Suppl 1):S100–S104
Salazar-García DC (2011) Patrón de dieta en la población púnica de Can Marines (Ibiza) a través del análisis de isótopos estables (C y N) en colágeno óseo. Saguntum 40:95–102
Salazar-García DC, Richards MP, Nehlich O, Henry AG (2014) Dental calculus is not equivalent to bone collagen for isotope analysis: a comparison between carbon and nitrogen stable isotope analysis of bulk dental calculus, bone and dentine collagen from same individuals from the medieval site of El Raval (Alicante, Spain). J Archaeol Sci 47:70–77
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639
Schulting RJ (2018) How ‘marine’ were coastal Mesolithic diets? In: Borić D, Antonović D, Mihailovic B, Stefanovic S (eds) Holocene foragers in Europe and beyond. Serbian Archaeological Society and Oxbow Books, Oxford
Schwarcz HP, Schoeninger MJ (1991) Stable isotope analyses in human nutritional ecology. Yearb Phys Anthropol 34:283–321
Sponheimer M, Robinson T, Ayliffe L, Roeder B, Hammer J, Passey B, West AT, Cerling T, Dearing D, Ehleringer J (2003) Nitrogen isotopes in mammalian herbivores: hair δ15N values from a controlled feeding study. Int J Osteoarchaeol 13:80–87
van Strydonck M, Boudin M, Ervynck A (2002) Stable isotopes (13C and 15N) and diet: animal and human bone collagen from prehistoric sites on Mallorca, Menorca and Formentera. In: Waldren WH, Ensenyat JA (eds) World Islands in Prehistory, vol 1095. Archaeopress, BAR International Series, Oxford, pp 189–197
van Strydonck M, Boudini M, Ervynck A, Orvay J, Borms H (2005) Spatial and temporal variation of dietary habits during the prehistory of the Balearic Islands as reflected by 14C, δ15N and δ13C analyses on human and animal bones. Mayurqa 30:523–541
Tafuri MA, Craig OE, Canci A (2009) Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Am J Phys Anthropol 139(2):146–153
Tomás MS (2009) El uso terapéutico de la alimentación en la Baja Eded Media. In: Arízaga B, Solórzano JA (eds) Alimentar la Ciudad en la Edad Media. Instituto de Estudios Riojanos, Logroño, pp 459–490
Van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26(6): 687–695
Virginia R, Delwiche C (1982) Natural 15N abundance of presumed N2-fixing and non-N2-fixing plants from selected ecosystems. Oecologia 54:317–325
Waines D (1992) The culinary culture of al-Andalus. In: Jayyusi SK (ed) The legacy of Muslim Spain. Brill, Lieden, pp 725–740
Wang Y, Cerling TE (1994) A model of fossil tooth and bone diagenesis: implications for paleodiet reconstruction from stable isotopes. Palaeogeogr Palaeoclimatol Palaeoecol 107:281–289
Warinner C, Tuross N (2009) Alkaline cooking and stable isotope tissue-diet spacing in swine: archaeological implications. J Archaeol Sci 36:1690–1167
Watson AM (1983) Agricultural innovation in the early Islamic world. The diffusion of crops and farming techniques, 700–1000. Cambridge University Press, Cambridge
Watson JL, Caldwell ML (eds) (2004) The cultural politics of food and eating. Blackwell, Oxford
White CD, Longstaffe FJ, Law KR (2004) Exploring the effects of environment, physiology and diet on oxygen isotope ratios in ancient Nubian bones and teeth. J Archaeol Sci 31:233–250
Wright LE, Schwarcz HP (1998) Stable carbon and oxygen isotopes in human tooth enamel: identifying breastfeeding and weaning in prehistory. Am J Phys Anthropol 106(1):1–18
Zaouali L (2007) Medieval cuisine of the Islamic world. University of California Press, Berkeley
Ziriax M (2017) The effects of climate and geography on Palaeodietary reconstruction with a focus on late antique Spain. Unpublished DPhil thesis, School of Archaeology, University of Oxford
Acknowledgments
The authors would like to thank Julia Lee-Thorp and Maura Pellegrini for discussions concerning the interpretation of enamel carbonate δ18O results, and to two anonymous reviewers and the guest editors of this issue. Thanks also to Pete Ditchfield for mass spectrometry support. The stable isotope and AMS radiocarbon measurements were supported through the School of Archaeology, University of Oxford.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Dury, G., Lythe, A., Marquez-Grant, N. et al. The Islamic cemetery at 33 Bartomeu Vicent Ramon, Ibiza: investigating diet and mobility through light stable isotopes in bone collagen and tooth enamel. Archaeol Anthropol Sci 11, 3913–3930 (2019). https://doi.org/10.1007/s12520-018-0644-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-018-0644-4