
Abstract
In this study, we examine the role of foxes in Palaeolithic economies, focusing on sites of the Middle Palaeolithic, Aurignacian, Gravettian and Magdalenian of the Swabian Jura. For this purpose, we used published faunal data from 26 assemblages from the region, including new information from the Magdalenian layers of Langmahdhalde. We explore how the abundance of foxes changes over time, how they were used by humans, and how they were deposited at the sites, with a special focus on fox hunting methods. To evaluate these hunting methods, we use the prey choice model of optimal foraging theory (OFT) and simulate possible hunting scenarios, which we test based on the published faunal assemblages. Our research indicates that foxes were hunted since the early Upper Palaeolithic for their meat, fur and teeth, possibly with traps. We find that the abundance of fox remains in the archaeological record of the region increased continuously starting in the Aurignacian, which cannot be explained by taphonomic factors. The trend of foxes to adapt to human-influenced environments with commensal behavior may also have contributed to them being hunted more often.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The transition from the Middle Palaeolithic, which was dominated by Neanderthals, to the Upper Palaeolithic, which is associated with the appearance of anatomically modern humans, is an important and very active field of research in archaeology (Arrighi et al. 2019; Benazzi et al. 2011; Fa et al. 2016; Jones et al. 2019; Jones et al. 2018; Morales et al. 2016; Peresani et al. 2016; Pirson et al. 2012; Pleurdeau et al. 2016; Richard et al. 2019; Romandini et al. 2019). In zooarchaeological studies, this transition is often interpreted as a change in human hunting behavior or an expansion of the food spectrum (Romandini et al. 2019; Starkovich 2012; Starkovich 2014; Stiner 2009; Stiner et al. 2000; Stiner et al. 1999). Although recent studies suggest that Neanderthals had a broad diet in some regions (Droke et al. 2020; Wißing et al. 2019; Wißing et al. 2016; Wroth et al. 2019; Yravedra et al. 2019), many sites in Europe and the Middle East have rapid increases in faunal diversity from their Middle to Upper Palaeolithic layers (Conard et al. 2013; Starkovich 2012; Starkovich 2014; Stiner 2009; Stiner et al. 2000), which can be interpreted as a result of a higher population density of modern humans and a resulting over-hunting of certain food resources (Stiner et al. 1999). In this context, representation of primarily small prey animals, such as hares, birds, fish and foxes, increases. While archaeological evidence strongly supports the use of hares, fish and birds as additional components of human diets (Conard et al. 2013; Stiner 2009; Stiner et al. 2000; Stiner et al. 1999), the use of foxes for both fur and meat is less strongly supported but is likely to have occurred (Camarós et al. 2016; Conard et al. 2013; Yeshurun et al. 2009). In modern hunter-gatherer cultures, such as some Inuit tribes of the eastern Arctic (Eber 1989), it is common to consume foxes regularly only in times when no other food is available. This makes the presence of fox remains an important indicator for over-hunting of main prey and human population growth, following the hypothesis of Stiner et al. (1999). Furthermore, in some contexts, it is also possible that humans were not responsible for depositing fox remains at the sites. Our study focuses on the human use of foxes from the Middle Palaeolithic to the Magdalenian in the Swabian Jura of southwestern Germany, a region with some of the most well-researched Palaeolithic sites in Europe.
Researchers have studied the archaeofaunal remains from the Swabian Jura for decades. The sites are particularly famous for their rich Middle and Upper Palaeolithic layers. While the Middle Palaeolithic settlement of the region appears to have been sparse, most archaeological sites dating to the Upper Palaeolithic seem to have been used more intensively (Conard et al. 2012). In the Swabian Jura, the Upper Palaeolithic is represented by the Aurignacian (42,000–34,000 cal BP (Conard and Bolus 2003; Conard and Bolus 2008; Hahn 1982; Higham et al. 2012)), Gravettian (34,000–24,000 cal BP (Conard and Bolus 2008; Housley et al. 1997; Taller and Conard 2019)), and after the Last Glacial Maximum (LGM), the Magdalenian (16,300 to approximately 12,700 cal BP (Gaudzinski and Street 2003; Hahn 1995; Housley et al. 1997; Kind 2003; Taller et al. 2014)).
In general, archaeological sites in the Ach and Lone valleys of the Swabian Jura are dominated by remains of cave bear and ungulates throughout the Palaeolithic (e.g. Bertacchi (2017); Camarós et al. (2016); Conard et al. (2013); Kitagawa et al. (2012); Krönneck (2012); Lykoudi (2017); Münzel (2019); Münzel and Conard (2004a); Münzel and Conard (2004b); Napierala et al. (2014); Wong et al. (2017)). However, there are also small numbers of carnivore taxa in almost all of the sites, including red fox (Vulpes vulpes) and arctic fox (Vulpes lagopus) (Bertacchi 2017; Camarós et al. 2016; Conard et al. 2013; Kitagawa et al. 2012; Krönneck 2012; Lykoudi 2017; Münzel 2019; Münzel and Conard 2004a; Münzel and Conard 2004b; Napierala et al. 2014; Wong et al. 2017). We know that carnivores must have played an important role in the lives of Palaeolithic peoples, based on, among other things, the presence of perforated carnivore teeth in the archaeological record, which were most likely used as pendants (Camarós et al. 2016; Conard 2003; Hahn 1992; Langguth and Malina 2003; Münzel 2019). The earliest evidence of these pendants in the Swabian Jura comes from the Aurignacian layers of Hohle Fels and Geißenklösterle. In the Gravettian, perforated teeth of carnivores are found even more frequently (Camarós et al. 2016; Conard 2003). In addition to pendants made of carnivore teeth, felids and bears were illustrated in mobile art in hunting poses (Hahn 1986). Although no Palaeolithic figurines have been interpreted as depicting foxes, the regular presence of foxes in the faunal record, as well as the use of their teeth as pendants, indicates that these animals were important to humans during these periods. However, the nature of this relationship has not yet been studied in detail.
A particular behavior in foxes could be related to their occurrence in archaeological sites: commensalism. Commensal behavior is a symbiotic behavior between two species, which benefits one species and does not affect the other (Hulme-Beaman et al. 2016; Yeshurun et al. 2009). Modern foxes show two types of commensalism. In one type, they are commensal to large predators (Pulliaines 1993; Wandeler and Lüps 1993), such as wolves or polar bears and, and in the other type, they are commensal to humans (Jędrzejewski and Jędrzejewska 1992; Kidawa and Kowalczyk 2011; MacDonnald 1977; Panek and Budny 2017; Pulliaines 1993; Savory et al. 2014; Sidorovich et al. 2006; Soe et al. 2017; Wandeler and Lüps 1993). In both cases, the foxes benefit by obtaining food more easily without positively or negatively affecting the large predators or humans (Hulme-Beaman et al. 2016; Kays and Feranec 2011; Merkle et al. 2011; Murray et al. 2015; Newsome et al. 2010; Newsome et al. 2015; Warsen et al. 2014). Recent research has indicated that this phenomenon is not restricted to modern foxes (West and Yeshurun 2019; Yeshurun et al. 2009), but also likely occurred during the Magdalenian (Baumann et al. 2020).
This paper has three primary goals. First, we seek to determine how the abundance of foxes has changed from the Middle Palaeolithic to the Magdalenian in the Swabian Jura. Second, we explore how foxes were used during these different periods. Finally, we discuss the circumstances under which fox remains were likely deposited in the Palaeolithic sites of the Swabian Jura, including possible methods of fox hunting by humans. We address these goals by conducting a meta-analysis of previously published studies and presenting new data from recent excavations at Langmahdhalde. Furthermore, we interpret our results through the paradigm of human behavioral ecology and the prey choice model (Charnov 1976) in order to explore possible hunting scenarios Palaeolithic people might have used to procure foxes.
Material and methods
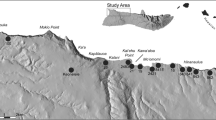
Our studied material consists of 26 assemblages from twelve different cave and rock shelter sites from the Ach and Lone valleys in the Swabian Jura that date to the Middle Palaeolithic (MP), Aurignacian (A), Gravettian/Aurignacian transition (G/A), Gravettian (G) and Magdalenian (M) (Fig. 1, Table 1). From the Ach Valley, we have included Kogelstein, Hohle Fels, Helga Abri, Sirgenstein, Geißenklösterle, Brillenhöhle and Große Grotte, and the Lone Valley is represented by Hohlenstein-Stadel, Fetzershaldenhöhle, Vogelherd, Langmahdhalde and Bockstein. We report the publications we used as sources for the NISP values (number of identified specimens, Grayson (2014); Lyman (2008)) in Table 1.

Map of sites included in this study. 1 = Langmahdhalde; 2 = Fetzershaldenhöhle; 3 = Vogelherd; 4 = Bockstein; 5 = Hohlenstein-Stadel; 6 = Große Grotte; 7 = Brillenhöhle; 8 = Geißenklösterle; 9 = Sirgenstein; 10 = Hohle Fels and Helga Abri; 11 = Kogelstein. Map made in QGIS version 3.4 with topographic data from SRTM NASA version 3, hydrology data from the Landesanstalt für Umwelt Baden-Württemberg, administrative boundaries from ©EuroGeographics, and ocean data from Natural Earth
The data we present in this paper are based on both piece-plotted finds from excavations of the 1970s onwards and individual collected finds from excavations before this date. In the following, we will refer to them as “single finds”. Even if many excavation sites were wet-sieved, the majority of these wet-sieved remains have not yet been published. The published NISP values for the Middle Palaeolithic assemblages of Kogelstein group the wet-sieved and single finds together; in order to address any biases this may introduce, we analysed Kogelstein separately. Wong et al. (2017) calculated the NISP values for the faunal remains from the Magdalenian site of Langmahdhalde using both single finds and wet-sieved material, but, as this site is still under excavation, these NISP values do not reflect the most current values. Therefore, supplementary Table S1 presents the most up-to-date NISP values from the Langmahdhalde Magdalenian assemblage for the taxonomic groups in this study and we use the values based only on single finds.
In general, fox remains are not well-represented at these sites compared to other mammalian taxa, except in Kogelstein, probably as a result of including screened materials. Calculating the relative abundance of foxes using percent NISP (%NISP) does not provide reliable results as the total number of foxes is very small in relation to the total NISP. Therefore, we use the ratio of the NISP of foxes to the NISP of large herbivores (fox/LH index), following Tchernov (1994) and Yeshurun et al. (2009). We only use faunal remains that are identified to the genus (i.e. Vulpes sp.) or species level (i.e. arctic fox: Vulpes lagopus and red fox: Vulpes vulpes). The category of large herbivore (LH) includes mammoth (Mammuthus primigenius), horse (Equus ferus), wooly rhinoceros (Coelodonta antiquitatis), giant deer (Megaloceros giganteus), red deer (Cervus elaphus), reindeer (Rangifer tarandus), bison (Bison sp.), aurochs (Bos primigenius), ibex (Capra ibex), chamois (Rupicapra rupicapra) and the size classes “mammoth/rhino”, “horse” and “reindeer.” Similarly, we calculated the ratio of hare (specimens identified to European hare, Lepus europaeus, mountain hare, Lepus timidus and Lepus sp.) to large herbivores (LH). It is possible that fox teeth are overrepresented in the single find material since they are easily identified during excavation. However, we have decided to include both postcranial and cranial elements, including teeth, in our analysis, because an overrepresentation of teeth is only present in the Magdalenian layers of Geißenklösterle (see Fig. S1), and because at some sites (e.g. Brillenhöhle and Große Grotte), no distinction was made between cranial and postcranial remains for fox or hare.
Another method we used to calculate the relative abundance of foxes is the ratio of foxes within the carnivores (NISP fox/NISP all carnivores, including foxes), or %fox of Carnivora. The carnivore category includes foxes, cave lion (Panthera leo spelaea), lynx (Lynx lynx), wild cat (Felis silvestris), wolf (Canis lupus), wolverine (Gulo gulo), martens (Martes sp.), weasels (Mustela sp.), otter (Lutra lutra) and cave hyena (Crocuta crocuta spelaea). We excluded the European badger (Meles meles) from our analysis, because we could not exclude the possibility that modern badgers invaded the archaeological layers of some of the sites, such as Vogelherd (Niven 2006).
Moreover, we have excluded the cave bear (Ursus spelaeus), which belongs to the order Carnivora, but did not have a carnivorous diet (Bocherens et al. 1994; Münzel et al. 2014; Naito et al. 2016; Nelson et al. 1998; Pacher and Stuart 2009). Furthermore, due to their hibernation behavior, it is possible that cave bears died in the cave sites without human influence (Baca et al. 2016; Kitagawa et al. 2012; Münzel and Conard 2004a; Nelson et al. 1998; Pacher and Stuart 2009; Stiner 1999). To get an impression of whether the number of foxes is related to the human occupation density, we use fox density (NISP of fox/m3 excavated sediment) and lithic artefact density (number of lithic artefacts/m3 excavated sediment, from Conard et al. (2012)). The sites and values we include in this analysis are given in supplementary Table S2.
Since the sample size within archaeological assemblages is very small, we applied Bayesian statistics to measure fox and hare abundance. Bayesian statistics have the advantage that they provide usable probability and likelihood information even with small sample sizes (n > 5, Gelman et al. (2014)). Meanwhile, Bayesian statistics are increasingly used in archaeological studies (Borradaile 2003; Gearey et al. 2009; Halekoh and Vach 1999; Halekoh and Vach 2004; Otárola-Castillo and Torquato 2018). One way to evaluate the reliability of the statistical analysis is by using the credibility interval (CI; 95% by default) of the regression line. The narrower the CI, the more certain is the given regression trend.
For the correlation between fox and hare indices (fox/LH and hare/LH, respectively), the fox abundance within the carnivore sub-assemblages (%fox of Carnivora) between periods and the analysis of the relationship between fox density and lithic artefact density, we use a normal linear model (lm) analysis in a Bayesian framework (R package arm; Gelman and Hill (2006)), following the protocol of Korner-Nievergelt et al. (2015). The lm analysis gives an adjusted (adj.) r2 value between 0 and 1, which can be used for the interpretation of the regression line (Korner-Nievergelt et al. 2015): positive values indicate a positive correlation and negative values indicate a negative correlation. The use of Bayesian statistics eliminates the need for the p value (probability value in frequency analyses), as this is generally too strongly biased by the sample size (Gelman et al. 2014; Halekoh and Vach 1999; Halekoh and Vach 2004; Korner-Nievergelt et al. 2015; Otárola-Castillo and Torquato 2018). All Bayesian analyses were done with R Version 3.6.1.
Results
In this section, we present the relative abundance and frequency distributions of foxes from the Swabian Jura Middle Palaeolithic to Magdalenian. We also summarize incidences of human-made modifications on fox remains, such as cut marks and perforated tooth pendants, and the abundance of hares and foxes.
Fox representation from the Middle Palaeolithic to the Upper Palaeolithic
The majority of fox bones in all sites are from adult animals. We have recorded only 28 bones and teeth from foxes younger than 1 year. Seventeen juvenile fox bones/teeth come from the Middle Palaeolithic layers at Hohlenstein-Stadel (Kitagawa 2014), one from the Aurignacian of Vogelherd (Niven 2006), four from the Gravettian layer at Geißenklösterle (Münzel 2019) and six from the Magdalenien layers of Vogelherd and Langmahdhalde (1 and 5, respectively; Niven (2006)).
In most assemblages, fox remains make up only 1 to 5% of the faunal material (Bertacchi (2017); Boessneck et al. (1973); Boger et al. (2014); Böttcher et al. (2000); Conard et al. (2013); Kitagawa (2014); Kitagawa et al. (2012); Krönneck (2012); Lykoudi (2017); Münzel (2019); Napierala et al. (2014); Niven (2006); Weinstock (1999); Wong et al. (2017); Table S1). Only in the Middle Palaeolithic layers of Kogelstein Cave are foxes the dominant taxa by NISP count, with 418 specimens (37.5% of the assemblage) (Böttcher et al. 2000). Nevertheless, in all of the other assemblages, we see an increase in the relative abundance of foxes within the carnivore sub-assemblages (%fox of Carnivora) from the Middle Palaeolithic to the Magdalenian (Fig. 2a, Table 2). Similar to the increasing abundance of foxes within the carnivores, the fox/LH index increases from the Middle Palaeolithic to the Magdalenian layers, as does the hare/LH index (Fig. 2b, Table 2).

a Relative abundance of foxes in relation to other carnivores. Red solid line shows the calculated mean regression based on Bayesian linear model, red shaded area shows 95% credibility interval. b Index of the relationship between foxes and large herbivores (LH) and between hares and large herbivores (LH). Solid lines show the calculated mean regressions based on Bayesian linear model, shaded areas show 95% credibility interval. All values are based on NISP data. MP = Middle Palaeolithic, G/A = Gravettian/Aurignacian transition layer
In five assemblages (Fig. S2, Table 2) from the Middle Palaeolithic layers (excluding Kogelstein), the index of both foxes and hares is very low (on average 0.06 and 0.03, respectively). In the Aurignacian layers, the average frequency of both taxa remained low, with the exception of the assemblages from Brillenhöhle, Hohlenstein-Stadel and Sirgenstein. In the faunal material of Hohlestein-Stadel, both indices clearly increased (on average 0.34 and 0.29, respectively). In the Brillenhöhle and Sirgenstein material, only the hare/LH index increased (0.25 and 0.29, respectively). With the transition to the Gravettian (i.e. the G/A layers from Hohle Fels and Fetzershaldenhöhle), as well as throughout the Gravettian, there is an increase in the two indices (G/A: on average 0.18 and 0.21, respectively; G: on average 0.14 and 0.59, respectively). The average frequency of hares in the faunal material was higher than that of foxes, both in the G/A transition and in the Gravettian. This was particularly evident in Brillenhöhle (hare/LH = 2.07) and Sirgenstein (hare/LH = 1.74), where hares outnumber large herbivores by NISP count.
When we look at the Magdalenian, we see that the average frequency of foxes continued to increase, while the average frequency of the hares decreased slightly. For the foxes, the highest value is from Geißenklösterle (fox/LH = 0.72). However, the number of faunal remains from the Magdalenian in Geißenklösterle is generally low (NISP = 229) compared to the other archaeological layers.
Previous studies have found that including fox teeth in fox abundance data leads to an overrepresentation of foxes in the assemblage of Geißenklösterle (Camarós et al. 2016; Conard et al. 2012; Conard et al. 2013; Hahn 1988; Münzel and Conard 2004b). For the Magdalenian in Geißenklösterle, for example, 20 out of 28 fox specimens are teeth. To test whether this is also true of the other assemblages, we calculated two fox/LH indices, one with and one without isolated teeth, and plotted them against each other (Fig. S1). We found that the abundance of fox remains from the Magdalenian of Geißenklösterle is particularly affected by tooth representation, but with all other assemblages, the difference between the two indices was minor. Based on these results and the fact that the published faunal results for the sites in this study do not all indicate how many isolated fox teeth are in the assemblages (e.g. in Hohlenstein-Stadel and Kogelstein), we consider only the fox/LH index that includes teeth in discussions below. In addition, we tested the inclusion of teeth (of foxes and hares) on the relationship between foxes and hares (Fig. S4). We found that the linear correlation for both taxa is lower when teeth are not included (all remains: adj. r2 = 0.13, only postcranial: r2 = 0.05). There are two reasons for this. Firstly, teeth and bone were not reported separately for all sites, which leads to a smaller sample size and thus to a higher uncertainty. Secondly, because of its high mineral content, enamel generally preserves better than bone and is thus represented in higher proportions. However, we consider including teeth to be useful because it enables us to compare even the sites that do not provide separate data on teeth and bone.
The increasing frequency of foxes through time seems to coincide with an increase in hare frequency. Examining fox frequency in relation to hare may yield some important insights, as these taxa are similar in size, share the same habitats and are nocturnal (Averianov et al. 2003; Pulliaines 1993; Thulin and Flux 2003; Wandeler and Lüps 1993). Figure 3 shows a comparison of the fox/LH index and the hare/LH index. In order to examine whether foxes and hares were captured in a similar manner, we analysed the correlation between both indices for each period. If the correlation yields a positive linear relationship, it would mean that both taxa were caught in similar proportions to one another (Yeshurun et al. 2009). This could then indicate that humans used an unselective hunting strategy, such as snares on an open landscape, to catch these animals. For the Middle Palaeolithic (n = 5), we found a very low negative correlation (adj. r2 = −0.26), while there is positive correlation for the Aurignacian (n = 7, adj. r2 = 0.50) and a much higher positive correlation in the Gravettian (n = 5, adj. r2 = 0.88). We also see a strong positive correlation for the Magdalenian (n = 5, adj. r2 = 0.99); however, we had to exclude the data from Geißenklösterle, again because of the overrepresentation of fox teeth and the resulting bias to the index (see Fig. S1).

Plot of the fox/large herbivore (LH) and hare/large herbivore (LH) indices. The cluster in the lower left corner indicates a balanced relationship between hare and fox remains in the assemblages. The Gravettian layer of Sirgenstein and both layers from Brillenhöhle (BH) show an overrepresentation in hares, while the two assemblages in the lower middle part indicate an overrepresentation in foxes. All values are based on NISP data (including teeth). GK = Geißenklösterle, Si = Sirgenstein, KS = Kogelstein, BH = Brillenhöhle, MP = Middle Palaeolithic, G/A = Gravettian/Aurignacian transition layer
In order to investigate whether foxes became more frequent when humans were using the sites, we compared lithic artefact density to the density of fox remains in the Middle Palaeolithic and early Upper Palaeolithic (Fig. 4). The assemblages from the early Upper Palaeolithic (n = 6) show a strong positive correlation between lithic artefact density and fox density (adj. r2 = 0.72). In contrast, the correlation in the Middle Palaeolithic sites (n = 5) is much lower (adj. r2 = 0.37).

Relationship between lithic artefact density (published in Conard et al. (2012)) and the density of foxes. Orange points = Middle Palaeolithic sites. Blue points = Aurignacian sites (Exception: G/A = Gravettian/Aurignacian transition layer). Blue solid line shows the calculated mean regression based on Bayesian linear model, blue shaded area shows 95% credibility interval. GK = Geißenklösterle, BS = Bockstein, HF = Hohle Fels, VH = Vogelherd, HS = Hohlenstein-Stadel, Si = Sirgenstein, GG = Große Grotte, MP = Middle Palaeolithic
Human modification of fox remains in the Middle and Upper Palaeolithic
Cut marks on fox bones are generally rare in the Palaeolithic, even if foxes are well-represented in assemblages (Lipecki and Wojtal 2015; Street and Turner 2013; Wilczyński et al. 2015; Wojtal and Wilczyński 2015; Wojtal et al. 2012), and the sites of the Swabian Jura are no exception. Cut marks were found on only ten fox bones in our study. From the Middle Palaeolithic layer of Bockstein, Krönneck (2012) observed a phalanx with a circular cutting line. In the Aurignacian, there are five bones with cut marks: two mandibles from Vogelherd, one with cuts on the buccal side and the other with cuts on the lingual side (Niven 2006), a fifth metacarpal from Geißenklösterle (Hahn 1988; Münzel 2019) and a rib and an ulna from Hohle Fels (Conard et al. 2013). Additionally, the only two bones with butchering marks from the Gravettian come from Hohle Fels: a pelvis and a femur (Conard et al. 2013). There are two modified bones from the Magdalenian: an ulna from Hohle Fels (Napierala et al. 2014) and a mandible with parallel cuts from Langmahdhalde (Wong et al. 2017). While cut marks on the pelvis and femur can come from meat removal, cut marks on the outside of the mandible and on phalanges tend to come from skinning (Andersson and Paulsson 1993; Binford 1981; Valensi 1991).
The fox remains from Kogelstein Cave
The Middle Palaeolithic layer of Kogelstein is a special case in our analysed assemblages. It stands out from the other sites in our study because it has 418 fox specimens, a fox abundance in relation to other carnivores of approximately 70% and a fox/LH index of 1.05. As mentioned above, Böttcher et al. (2000) included wet-sieved material in the NISP counts of the faunal remains. Since small elements and fragments, such as phalanges and epiphysis fragments, are often not recovered without screening, this could explain why Kogelstein has such high NISP values for fox compared to the other sites. We would therefore assume that the hares and other small taxa would be represented in higher numbers in the Kogelstein faunal assemblage. However, this is not the case; Böttcher et al. (2000) report only 65 hare remains from the Middle Palaeolithic horizons at the site. This site is considered to have been a hyena den that had only a short human occupation during the Middle Palaeolithic (Böttcher et al. 2000), meaning that the fox remains were not necessarily deposited as the result of human activity. Therefore, factors other than the inclusion of wet-sieved material likely influenced the high representation of fox remains in the Middle Palaeolithic horizons of Kogelstein.
Discussion
We now consider Palaeolithic foxes from the Swabian Jura in a broader context and focus on three topics: the extent to which foxes were used as food or raw material sources by humans, the population structure of foxes and finally human hunting methods used in the Upper Palaeolithic for foxes. For the first topic, we will compare our results with those of other studies in the Swabian Jura (Camarós et al. 2016; Conard et al. 2013; Kitagawa et al. 2012), Gönnersdorf (Street and Turner 2013) and the Gravettian sites of the Czech Republic (Wilczyński et al. 2015; Wojtal et al. 2012). In the second and third topics that deal with population structures and hunting methods, we will compare our results with modern behavioral studies of foxes and consider the prey choice model from optimal foraging theory (OFT) to explain human hunting behavior.
Fox use in the Central European Upper Palaeolithic
As we have shown in our study, humans living in the Swabian Jura during the Palaeolithic likely considered foxes to be both food resources and sources of raw material, such as fur and teeth. Other studies have come to similar conclusions, with Camarós et al. 2016 focusing on their use for food and Conard et al. (2013) highlighting their use for fur.
In our sample, we find only three specimens that suggest the use of foxes for their fur: one specimen from the Middle Palaeolithic at Bockstein, one from the Aurignacian at Vogelherd and one from the Magdalenian at Langmahdhalde. However, not every cut leaves a mark and analysts have shown that experienced hunters or butchers leave hardly any marks on bone during the butchering process (Andersson and Paulsson 1993; Binford 1981; Charles 1997). Furthermore, cut marks are often found on small bones, such as phalanges, which, due to their small size, are usually only recovered using finer methods of excavation, such as wet-sieving (Charles 1997). As this study does not consider wet-sieved finds, this may affect our results.
There have been several studies demonstrating that carnivore teeth were used as raw material for ornaments in the Swabian Jura during the Aurignacian and Gravettian (Camarós et al. (2016); Conard (2003); Hahn (1988); Hahn (1989); Hahn (1992); Kitagawa (2014); Kitagawa et al. (2012); Langguth and Malina (2003); Münzel (2019); Wehrberger (2007); Table 3, Fig. 5). This includes not only fox teeth, but also perforated cave lion, wolf and other carnivore teeth (Camarós et al. 2016; Conard 2003; Kitagawa et al. 2012; Langguth and Malina 2003; Pacher 2005). Together, the number of perforated teeth from the Aurignacian and Gravettian increases through time in archaeological assemblages (Camarós et al. 2016; Conard et al. 2013; Kitagawa et al. 2012). If only foxes are considered, however, more ornaments are found in the Aurignacian than Gravettian layers. However, the number of fox tooth ornaments and ornament fragments may increase if the finds from wet-sieving are eventually included. Based on the current evidence, fox tooth ornaments might have been more important in the Aurignacian than later in the Gravettian, where ornaments of other carnivores became more abundant.

a Nine fox tooth ornaments from the Aurignacian layers of Geißenklösterle (GK) and Hohle Fels (HF). Photos/drawings taken from (1) Langguth and Malina (2003), (2) Conard (2003) and (3) Camarós et al. (2016). Photos taken by Hilde Jenssen. AH = archaeological horizon. b Eight fox tooth ornaments from the Gravettian layers of Geißenklösterle (GK) and Hohle Fels (HF). Photos/drawings taken from (1) Langguth and Malina (2003), (2) Conard (2003) and (3) Camarós et al. (2016). Photos taken by Hilde Jenssen. AH = archaeological horizon
In order to understand whether the Swabian Jura is an exception in the case of fox use, we will examine other European Gravettian sites. The most suitable sites are Dolní Vĕstonice I and Pavlov I of the Czech Republic, because they are well-researched and contain relatively high numbers of fox remains (NISP = 766 and 5460, respectively). Since for Pavlov I only the south-eastern area was evaluated, it is likely that the number of fox remains is even higher. Both sites are open-air sites. While Dolní Vĕstonice I has been described as a specialized mammoth hunting camp, Pavlov I has been described as a long-term base camp (Wilczyński et al. 2015; Wojtal et al. 2012). Only adult foxes, which are represented by all skeletal parts, have been found at Dolni Vestonice I (Wilczyński et al. 2015). No cut marks were found on fox bones at the site, making it unclear what purpose, if any, the humans that used the site had for these animals (Wilczyński et al. 2015).
Pavlov I shows a similar scenario, though the number of fox bones is much higher. Archaeologists recovered several fox bones with cut marks, which indicate that the animals were skinned and butchered. Furthermore, there are also very deep cuts on fox long bones from this assemblage, which could not be assigned to specific human activities (Wojtal et al. 2012). Both sites demonstrate that foxes were common faunal elements in Czech Gravettian open-air sites and, at least in Pavlov I, foxes were exploited by humans for both their fur and meat.
Pavlov I also provides evidence that carnivore teeth were a preferred material for making personal ornaments. In total, 284 perforated fox teeth and 65 perforated wolf teeth were found at this site. Dolni Vestonice I and Pavlov I include wet-sieved material (Wilczyński et al. 2015; Wojtal et al. 2012), which may account for the large number of small carnivore remains at these sites compared to those in the Swabian Jura.
Consider now the situation in the Magdalenian. For the sites of the Swabian Jura, the calculated indices (%fox of Carnivora and fox/LH) indicate a high abundance of foxes. However, only two cut marks indicate the use of these animals by humans. There is also no evidence for the use of fox teeth as ornaments or pendants. In order to contextualize these results, we compare the data of the Swabian Jura with the Magdalenian of Gönnersdorf, in western Germany. A large number of fox remains were found at this open-air site (Street and Turner 2013). These are mainly arctic fox (NISP = 2810), but 12 specimens are red fox. About half of the finds are cranial elements, including isolated teeth which dominate the fox remains from the site (NISP = 907). Similar to the Swabian Jura sites, in Gönnersdorf there are rarely cut marks indicating the use of these animals for fur. Despite the high representation of cranial elements, only five bones show cut marks which were clearly the result of skinning (Street and Turner 2013). A few more cut marks (n = 30), which indicate butchering, were found on long bones, the pelvis, ribs, vertebrae and scapulae. These modifications allowed Street and Turner (2013) to argue that hunter-gatherers at Gönnersdorf used the open skinning technique for the production of flat pelts. Furthermore, they suggest that different methods were used for butchering the foxes. As in the Swabian sites, it seems likely that foxes were also used as food at Gönnerdorf (Street and Turner 2013). Unlike in the Swabian Jura, perforated fox teeth occur in the Magdalenian layers of Gönnersdorf. While the fox ornaments of Geißenklösterle, Hohle Fels and Hohlenstein-Stadel were mainly limited to canines, both canines and other perforated teeth (n = 45), most frequently premolars, were found in the Gönnersdorf assemblage (Álvarez-Fernández 2000; Álvarez-Fernández 1999; Street and Turner 2013).
Population structure and the prey choice model
As discussed above, foxes were exploited by humans throughout the Upper Palaeolithic, which leads us to the question, what methods did humans use to hunt foxes? Fox hunting probably differed from large mammal hunting due to differences in social structure, diurnal vs. nocturnal behavior, and meat yield, among other factors. We explore this topic using optimal foraging theory (OFT) and, specifically, the prey choice model (Charnov 1976) which has been used to examine hunter-gatherer subsistence in many different contexts (e.g. Jones (2004); Nagaoka (2019); Starkovich (2014); Stephens and Krebs (1986); Stiner (2009)).
Following the prey choice model, we assume that foragers take into account the time and energy required to search for and handle a food resource when making decisions about which resources to pursue. Foxes and hares have approximately the same body proportions (Averianov et al. 2003; Pulliaines 1993; Thulin and Flux 2003; Wandeler and Lüps 1993), so we assume they have similar net energetic returns and are, therefore, ranked similarly in terms of caloric return (Broughton et al. 2011). Furthermore, we assume that the handling costs (i.e. the killing and processing of the prey) are similar for both taxa. The main difference between the two taxa would therefore be search time. To quantify search time for fox and hare, it is necessary to have a look at the ecology and population structure of these animals.
The number of red or arctic foxes living in an area and their reproductive rate is mainly determined by the food supply (Pulliaines 1993; Wandeler and Lüps 1993). Studies of the average population density of modern foxes find 4 to 18 red foxes and 0.03 to 2.5 arctic foxes per 10 km2 (Pulliaines 1993; Wandeler and Lüps 1993). Within modern red fox populations, randomly taken samples have shown that 49 to 77% of the animals were younger than 1 year of age (Bögel et al. 1974; Jensen and Nielsen 1968; Lloyd et al. 1976; Van Haaften 1970; Wandeler and Lüps 1993). Modern red and arctic foxes in the wild have a maximum lifespan of 10 years (Pulliaines 1993; Wandeler and Lüps 1993).
Modern hares, in comparison, generally have much higher population densities and a shorter lifespan (maximum age of four years, Averianov et al. (2003); Thulin and Flux (2003)). On average, 200 to 300 European hares occur per 10 km2 (Averianov et al. 2003) and mountain hare numbers vary strongly from 10 animals per 10 km2 in Sweden (Bergengren 1969) to up to 2450 animals per 10 km2 in some parts of Scotland (Watson and Hewson 1973; Watson et al. 1973). In both hare species, population densities can increase dramatically if conditions are favorable, such as mild winters and more food availability in the spring (Averianov et al. 2003; Thulin and Flux 2003). Based on this information, the average ratio of modern foxes to hares in the environment (Europe) is approximately one fox to 20 hares (Averianov et al. 2003; Pulliaines 1993; Wandeler and Lüps 1993; Watson and Hewson 1973; Watson et al. 1973).
In most of the Swabian Jura sites, we find higher NISP values for hare than fox in the assemblages, but not in all. In the Aurignacian layers of Hohlenstein-Stadel and the Magdalenian layers of Geißenklösterle and Vogelherd, as well as in all Middle Palaeolithic layers (except Große Grotte), foxes are more abundant than hares. In the case of the Middle Palaeolithic, this difference in hare and fox representation is probably due to the low number of small mammal remains overall and higher likelihood that foxes would enter the sites naturally. The values of the two Magdalenian layers of Geißenklösterle and Vogelherd can also be explained by the low number of hare and fox remains. At both sites, Magdalenian humans occupied the caves only for a short time (Münzel 2019; Niven 2006). With regard to the ratio of foxes to hares in the assemblages, we find a ratio of 1:7 in the Gravettian of Hohle Fels, which is the layer with the highest difference between foxes and hares. Some of the other layers in the Swabian Jura sites even have more fox than hare remains (Table 2). We hypothesize that there is a discrepancy between the relative abundance of fox and hare remains in the archaeological record and in the natural environment.
We do not assume that taphonomic processes and excavation methods provide a suitable explanation for this discrepancy because, as already mentioned, foxes and hares have similar body sizes and their bones are similarly proportioned. This means that post-depositional taphonomic processes and the time depth of the assemblage should affect both taxa similarly.
All the sites considered in our study show this discrepancy between the relative abundance of fox and hare remains in the faunal assemblages and in the natural environment, even those with very low human occupation densities. As we found at Kogelstein Cave, humans do not have to be involved in the accumulation of many foxes remains (Böttcher et al. 2000). Since this cave was a hyena den, the foxes could have been hunted by hyenas or could have lived commensal to them. Alternatively, foxes might have used the cave as a den behavior hyenas were not around. Similar commensal behavior to large predators has been demonstrated among modern foxes (Hartová-Nentvichová et al. 2010; Kidawa and Kowalczyk 2011; Murdoch et al. 2010; Pulliaines 1993; Roth 2003; Wandeler and Lüps 1993; Wikenros et al. 2017).
Another important point is that hares may also have been brought to the sites by carnivores, as described, for example, by Krajcarz and Krajcarz (2014). However, these bones should then show evidence of bite marks. This is the case in Kogelstein, Langmahdhalde, and in the archaeologically sterile layers of the other examined sites. In the layers of most of the sites we studied that showed considerable human activity, more cuts were found on the bones than bite marks. We can, therefore, assume that hares were introduced and used by humans rather than carnivores.
Kogelstein Cave and Langmahdhalde are exceptions for the sites examined in this study in this respect. At Kogelstein, fox and hare remains were deposited naturally. Further, at Langmahdhalde, there is evidence that some of the hare remains were deposited or scavenged by non-human predators in the form of puncture marks, bite marks and gnawing (Wong et al. in review). Therefore, we argue that fox behavior and human hunting methods can provide the best explanations for the large differences between the expected and observed ratios of hare and fox, assuming our expected ratio estimates are correct.
Palaeolithic fox hunting methods
As discussed above, we assume that the decision to hunt foxes vs. hares was made due to differences in the search time associated with each taxon. Therefore, we will now discuss four scenarios with different hunting methods that have a direct influence on search time. Scenario 1 is a hunt for individual animals with long-range weapons such as a throwing spear, throwing stick, sling or bow and arrow. In this case, the strongest influence on search time is the population size of the taxa, assuming these taxa are distributed similarly. Since hares occur in much higher population densities than foxes, hares would be taken more often than foxes because they would have been encountered more often. We would expect the ratio of hares to foxes in the archaeological assemblage be about the same as the natural population density ratio. Furthermore, we assume that mainly adult foxes (and hares) would be hunted, because adults are ranked higher than juveniles, based on their body size/weight (Broughton et al. 2011).
In the second scenario, traps are used to hunt both taxa. These are set up at game passes in an open landscape, which are used by small game. Since trap hunting only requires time to find the right place and set up the traps, there is no search time, unlike in scenario 1. Hares, which have higher population densities, should be more likely to be caught than foxes. We would therefore expect that in scenario 2, like scenario 1, the ratio between hare and fox remains in the archaeological assemblage would be similar to the population density ratio. Furthermore, we assume that mainly adult foxes (and hares) would be hunted, because adults are more mobile than juveniles (Averianov et al. 2003; Pulliaines 1993; Thulin and Flux 2003; Wandeler and Lüps 1993).
Scenario 3 involves setting traps near fox dens. Similar to scenario 2, this would mean that search time is low. It would also mean that hares would be less likely to be caught in these traps. We would therefore expect foxes to be more common than hares in the faunal assemblages. Furthermore, trapping near fox dens would sample a fox population randomly and should result in trapping approximately 49 to 77% young animals (Bögel et al. 1974; Jensen and Nielsen 1968; Lloyd et al. 1976; Van Haaften 1970; Wandeler and Lüps 1993).
The fourth scenario is trapping with meat as bait. Similar to the third scenario, this is a specialized hunt for foxes (Monchot and Gendron 2011). Despite the higher population density of hares, they would be unlikely to be trapped because they would not be attracted to the bait. In this scenario, we would expect there to be more foxes in zooarchaeological assemblages than hares. Furthermore, the trapped foxes would be mostly adult individuals, because adults are more mobile than juveniles (Pulliaines 1993; Wandeler and Lüps 1993).
Looking at all the sites from the Aurignacian in the Swabian Jura, there is an almost balanced 1:1 ratio between fox and hare remains (Table 2). In the Gravettian, there is an average ratio of 1:4, in some sites even 1:6 (Sirgenstein) or 1:7 (Hohle Fels). This shows that hares have gained in importance as a food and/or fur source from the Aurignacian to the Gravettian. Nevertheless, foxes were also important prey animals, as their NISP values also rose from the Aurignacian to the Gravettian. In the Magdalenian, the ratio between these two taxa is approximately 1:2 on average. None of the time/cultural periods studied, therefore, have a fox to hare ratio corresponding to the natural population density of both taxa (approximately 1:20), indicating that scenarios 1 and 2 probably do not alone describe how foxes were hunted during the Palaeolithic in the Swabian Jura. In scenarios 3 and 4, the number of fox remains is higher than the number of hare remains, a situation that does not fit with our results from the Aurignacian or Gravettian. Furthermore, scenario 3 predicts more young foxes than adults, which we are unable to evaluate in this study. Currently, analysts have identified 28 bones and teeth from foxes younger than 1 year in the Swabian Jura, though it is likely this number will increase when wet-sieved finds are evaluated. However, the Czech sites reviewed above show that young animals are also underrepresented there, despite the analysis of wet-sieved materials (Wilczyński et al. 2015; Wojtal et al. 2012). Another reason may be the poorer preservation of juvenile bones and teeth as opposed to adult bones and teeth, as argued by Street and Turner (2013). For our discussion of hunting methods, we will therefore not look at juveniles any further.
If we now consider the data for the individual periods, it becomes apparent that there can be no general statement as to the agent of accumulation for the fox remains across all periods. During the Middle Palaeolithic, which has a large depth of time compared to the other periods studied here, most of the sites contained more fox remains than hare remains (Table 2). Additionally, human occupation at this time was less intense in the Swabian Jura (Conard et al. 2012), indicating that the majority of the fox remains were likely deposited in the cave sites naturally and that humans were only sporadically involved.
The data from the Aurignaican and Gravettian show an increase in fox and hare remains, and the importance of fox teeth as ornaments demonstrates clearly that foxes were used by humans. Therefore, we hypothesize that foxes, as well as hares, were caught primarily with traps and snares at game passes in the early Upper Palaeolithic of the Swabian Jura. Further, the higher number of foxes in the find material could reflect a partial baiting of the traps with food remains. However, we cannot exclude the targeted hunting of foxes at their dens because of the limited information about juveniles in the record.
With the exception of Geißenklösterle, the trend continues in the two indices fox/LH and hare/LH with a lower number of finds in the Magdalenian of the Swabian Jura. We assume that some foxes were also hunted by humans, but due to the low occupation density in some of the studied sites compared to the early Upper Palaeolithic (Conard 2019; Münzel 2019; Niven 2006; Taller et al. 2014; Weniger 1987a; Weniger 1987b), they may have been introduced naturally. For this period, we also hypothesize that humans used traps and snares to hunt foxes, although to a lesser extent than in the Aurignacian and Gravettian.
It is also possible that there was a behavioral change, such as the development of a commensal relationship between foxes and humans, during the Upper Palaeolithic that influenced how often humans encountered foxes versus hares in their territories.
Commensalism between foxes and humans could explain the number of fox remains in the Swabian Jura during some periods of the Palaeolithic and has been discussed to explain trends in Levantine sites (Yeshurun et al. 2009) and in the Late Holocene of Alaska (West and Yeshurun 2019). Modern commensal animals appear in larger groups than their non-commensal relatives and are therefore more tolerant of other species, including humans (Dell'Arte et al. 2007; Hulme-Beaman et al. 2016; Panek and Budny 2017; Reshamwala et al. 2018; Wandeler and Lüps 1993). Especially for foxes, this behavior is an adaptation to a human-influenced environment (Lord et al. 2019) and has been shown to have occurred at Kesslerloch, a Magdalenian site in Switzerland (Baumann et al. 2020).
If we refer to the prey choice model, commensal behavior in foxes changes the search time for foxes because commensal foxes would have higher population densities within the vicinity of human settlement areas. This decrease in search time associated with commensal foxes would increase the success rate of hunting these animals compared to non-commensal foxes, making them more likely to have been included as prey for Palaeolithic hunter-gatherers. The positive correlation of Aurignacian foxes with occupation density (Fig. 4) could also be an indication of commensal behavior during the Upper Palaeolithic in the Swabian Jura. It is likely that not only one factor was decisive, but several. Thus, commensal, food-habituated foxes may have been hunted more often with traps, alongside non-commensal animals that were trapped or occasionally hunted as single prey items.
Conclusion
In this paper, we have established that the abundance of foxes at sites in the Swabian Jura changed from the Middle Palaeolithic to the Magdalenian. While only a few foxes were found in most sites of the Middle Palaeolithic, NISP values of these animals increased steadily beginning in the Aurignacian. A similar phenomenon was observed with the hare remains. Based on the assumptions of the prey choice model, this indicates that the suppression of higher-ranked ungulate taxa in the Upper Palaeolithic led foragers to turn to lower-ranked prey or, alternatively, that the introduction of different hunting methods (i.e. snares or traps) or desire for different currencies (i.e. fur or fox teeth) influenced the abundance of different taxa at the sites.
Fox teeth, which were used as a raw material for the production of pendants and ornaments (Camarós et al. 2016; Conard 2003; Conard et al. 2013; Langguth and Malina 2003; Münzel 2019), appear to be an important aspect of the human use of foxes in the Swabian Jura during the Palaeolithic. However, we also found some evidence for the use of foxes as a source of fur and meat. Finally, we discussed the circumstances under which fox remains were likely deposited in the Palaeolithic sites of the Swabian Jura: while we found no clear evidence of humans hunting foxes during the Middle Palaeolithic, the data from the Aurignacian and Gravettian show a pattern that suggests primarily trap hunting. It is also possible that foxes became commensal to humans and were therefore hunted more often during these periods. In the Magdalenian of the Swabian Jura, the human occupation of the region appears to have been smaller than in the early Upper Palaeolithic, which is also reflected in the lower density of faunal remains at the sites. We hypothesize that foxes were hunted by humans during this period and were also introduced naturally to the sites.
With our study, we have shown that, since the early Upper Palaeolithic, the fox gained in importance as a resource, similar to hares, fish and birds. If we apply these results to the hypothesis of Stiner et al. (1999), we can predict an increase in the population density of humans from the Aurignacian onwards. This result is consistent with previous studies in the region (Conard et al. 2012; Schmidt and Zimmermann 2019).
Availability of data and material
All data used are accessible via the tables in this study or from the cited publications.
References
Álvarez-Fernández E (2000) Las perlas de madera fósil del terciario y los objetos de adorno-colgantes sobre dientes de zorro y ciervo del Magdaleniense de Gónnersdorf y de Andernach-Martinsberg-2,(Neuwied, Rheinland Pfalz, Alemania) Zephyus 52:79-106
Álvarez-Fernández (1999) Arte mueble renano: Gönnersdorf y Andernach-Martinsberg-2, Neuwied, Alemania. Unpublished Doctoral Dissertation: Universidad de Salamanca
Andersson A, Paulsson T (1993) Jägarens skinn och hudar. Settern
Arrighi S, Moroni A, Tassoni L, Boschin F, Badino F, Bortolini E, Boscato P, Crezzini J, Figus C, Forte M, and Lugli F (2019) Bone tools, ornaments and other unusual objects during the Middle to Upper Palaeolithic transition in Italy. Quaternary International https://doi.org/10.1016/j.quaint.2019.11.016
Averianov A, Niethammer J, Pegel M (2003) Lepus europaeus Pallas, 1778—Feldhase. In: Krapp F, Niethammer J (eds) Handbuch der Säugetiere Europas, vol 3. AULA, pp 35-104
Baca M, Popović D, Stefaniak K, Marciszak A, Urbanowski M, Nadachowski A, Mackiewicz P (2016) Retreat and extinction of the Late Pleistocene cave bear (Ursus spelaeus sensu lato) The Science of. Nature 103:92
Baumann C, Starkovich BM, Drucker DG, Münzel SC, Conard NJ, Bocherens H (2020) Dietary niche partitioning among Magdalenian canids in southwestern Germany and Switzerland Quaternary Science Reviews 227:106032
Benazzi S et al (2011) Early dispersal of modern humans in Europe and implications for Neanderthal behaviour ature 479:525-528. https://doi.org/10.1038/nature10617
Bergengren A (1969) On genetics, evolution and history of distribution of the heath-hare, a distinct population of the Arctic hare, Lepus timidus Lin. Viltrevy 6:381–460
Bertacchi A (2017) Subsistence strategies and environmental change during the middle and upper Palaeolithic in the Swabian Jura (SW Germany): insights from Sirgenstein cave. Universität Tübingen, Master's Thesis
Binford LR (1981) Bones: ancient men and modern myths. Academic press
Bocherens H, Fizet M, Mariotti A (1994) Diet, physiology and ecology of fossil mammals asinferred from stable carbon and nitrogen isotope biogeochemistry: implications for Pleistocene bears paleogeography. Paleoclimatology, Paleoecology 107:213–225
Boessneck J, von den Driesch A, Lepiksaar J, Riek G, Storch G (1973) Das Paläolithikum der Brillenhöhle bei Blaubeuren (Schwäbische Alb) II: die jungpleistozänen Tierknochenfunde aus der Brillenhöhle. Verlag Müller & Gräff, Stuttgart
Bögel K, Arata A, Moegle H, Knorpp F (1974) Recovery of reduced fox populations in rabies control 1 Zentralblatt für Veterinärmedizin Reihe B 21:401-412
Boger U, Starkovich BM, Conard NJ (2014) New insights gained from the faunal material recovered during the latest excavations at Vogelherd Cave Mitteilungen der Gesellschaft für Urgeschichte 23:57–81
Borradaile GJ (2003) Viscous magnetization, archaeology and Bayesian statistics of small samples from Israel and England Geophysical research letters 30
Böttcher R et al. (2000) Kogelstein–eine mittelpaläolithische Fundstelle bei Schelklingen-Schmiechen Fundberichte aus Baden-Württemberg 24:7–176
Broughton JM, Cannon MD, Bayham FE, Byers DA (2011) Prey body size and ranking in zooarchaeology: theory, empirical evidence, and applications from the northern Great Basin. Am Antiq 76:403–428
Camarós E, Münzel SC, Cueto M, Rivals F, Conard NJ (2016) The evolution of Paleolithic hominin–carnivore interaction written in teeth: Stories from the Swabian Jura (Germany) J Archaeol Sci Rep 6:798–809. https://doi.org/10.1016/j.jasrep.2015.11.010
Charles R (1997) The exploitation of carnivores and other fur-bearing mammals during the North-western European ate and pper Paleolithic and Mesolithic. Oxf J Archaeol 16:253–277
Charnov EL (1976) Optimal foraging: attack strategy of a mantid. Am Nat 110:141–151
Conard NJ (2003) Eiszeitlicher schmuck auf der Schwäbischen Alb. In: Köbl S, Conard NJ (eds) Eiszeitschmuck - status und Schönheit, vol Museumsheft 6. Urgeschichtliches Museum, Blaubeuren, pp 15–50
Conard NJ (2019) Excavations at Geißenklösterle. In: Conard NJ, Bolus M, Münzel SC (eds) Geißenklösterle: Chronostratigraphie. Paläoumwelt und Subsistenz im Mittel- und Jungpaläolithikum der Schwäbischen Alb. Kerns Verlag, Tübingen, pp 9–21
Conard NJ, Bolus M (2003) Radiocarbon dating the appearance of modern humans and timing of cultural innovations in Europe: new results and new challenges. J Hum Evol 44:331–371
Conard NJ, Bolus M (2008) Radiocarbon dating the late Middle Paleolithic and the Aurignacian of the Swabian Jura Journal of Human Evolution 55:886–897
Conard NJ, Bolus M, Münzel SC (2012) Middle Paleolithic land use, spatial organization and settlement intensity in the Swabian Jura, southwestern Germany Quaternary International 247:236–245
Conard NJ, Kitagawa K, Krönneck P, Böhme M, Münzel SC (2013) The importance of fish, fowl and small mammals in the Paleolithic diet of the Swabian Jura, southwestern Germany. Zooarchaeology and Modern Human Origins. Vertebrate Paleobiology and Paleoanthropology, In, pp 173–190. https://doi.org/10.1007/978-94-007-6766-9_11
Dell'Arte GL, Laaksonen T, Norrdahl K, Korpimäki E (2007) Variation in the diet composition of a generalist predator, the red fox, in relation to season and density of main prey Acta Oecologica 31:276–281. https://doi.org/10.1016/j.actao.2006.12.007
Droke JL, Schmidt CW, Williams FLE, Karriger WM, Smith FH, Becam G, de Lumley M-A (2020) Regional variability in diet between Northern European and Mediterranean Neandertals: evidence from dental microwear texture analysis. Dental Wear in Evolutionary and Biocultural Contexts. Elsevier, In, pp 225–241
Eber DH (1989) When the whalers were up north: Inuit memories from the eastern Arctic vol 1. McGill-Queen's Press-MQUP
Fa DA, Finlayson JC, Finlayson G, Giles-Pacheco F, Rodríguez-Vidal J, Gutiérrez-López JM (2016) Marine mollusc exploitation as evidenced by the Gorham's Cave (Gibraltar) excavations 1998–2005: The Middle–Upper Palaeolithic transition Quaternary international 407:16–28
Gaudzinski S, Street M (2003) Reconsidering hunting specialisation in the German Magdalenian faunal record BAR International Series 1144:11–22
Gearey BR, Marshall P, Hamilton D (2009) Correlating archaeological and palaeoenvironmental records using a Bayesian approach: a case study from Sutton Common, South Yorkshire. England J Archaeol Sci 36:1477–1487
Gelman A, Carlin J, Stern H, Dunson D, Vehtari A, Rubin D (2014) Bayesian data analysis. Taylor & Francis, Boca Raton
Gelman A, Hill J (2006) Data analysis using regression and multilevel/hierarchical models. Cambridge university press
Grayson DK (2014) Quantitative Zooarchaeology: topics in the analysis of archaelogical faunas. Elsevier
Hahn J (1982) Eine menschliche Halbreliefdarstellung aus der Geißenklösterle-Höhle bei Blaubeuren Fundberichte aus Baden-Württemberg 7:1–12
Hahn J (1986) Kraft und Aggression: die Botschaft der Eiszeitkunst im Aurignacien Süddeutschlands? Vol 7. Archaeologica Venatoria, Tübingen
Hahn J (1988) Die Geißenklösterle-Höhle im Achtal bei Blaubeuren. Forschung und Berichte zur Vor- und Frühgeschichte Baden-Württemberg, vol 26. Theiss Verlag, Stuttgart
Hahn J (1989) Zur Funktion einer Aurignacien-Feuerstelle aus dem Geißenklösterle bei Blaubeuren Fundberichte aus Baden-Württemberg 14:1–22
Hahn J (1992) Eiszeitschmuck auf der Schwäbischen Alb. Süddeutsche Verlag-Gesellschaft
Hahn J (1995) Neue Beschleuniger-14C-Daten zum Jungpaläolithikum in Südwestdeutschland Eiszeitalter und Gegenwart 45:86–92
Halekoh U, Vach W (1999) Bayesian seriation as a tool in archaeology. Bar Int Series 750:107–107
Halekoh U, Vach W (2004) A Bayesian approach to seriation problems in archaeology Computational statistics & data analysis 45:651–673
Hartová-Nentvichová M, Šálek M, Červený J, Koubek P (2010) Variation in the diet of the red fox (Vulpes vulpes) in mountain habitats: effects of altitude and season mammalian biology - Zeitschrift für Säugetierkunde 75:334–340. https://doi.org/10.1016/j.mambio.2009.09.003
Higham T, Basell L, Jacobi R, Wood R, Ramsey CB, Conard NJ (2012) Τesting models for the beginnings of the Aurignacian and the advent of figurative art and music: The radiocarbon chronology of Geißenklösterle Journal of human evolution 62:664–676
Housley RA, Gamble CS, Street M, Pettitt P Radiocarbon evidence for the Lateglacial human recolonisation of Northern Europe. In: Proceedings of the Prehistoric Society, 1997. Cambridge University Press, pp 25–54
Hulme-Beaman A, Dobney K, Cucchi T, Searle JB (2016) An ecological and evolutionary framework for commensalism in anthropogenic environments. Trends Ecol Evol 31:633–645. https://doi.org/10.1016/j.tree.2016.05.001
Jędrzejewski W, Jędrzejewska B (1992) Foraging and diet of the red fox Vulpes vulpes in relation to variable food resources in Biatowieza National Park, Poland Ecography 15:212–220. https://doi.org/10.1111/j.1600-0587.1992.tb00027.x
Jensen B, Nielsen LB (1968) Age determination in the red fox (Vulpes vulpes L.) from canine tooth sections. Danish Rev. Game Biol. 5(6):1–16
Jones EL (2004) Dietary evenness, prey choice, and human–environment interactions. J Archaeol Sci 31:307–317
Jones JR, Richards MP, Reade H, de Quirós FB, Marín-Arroyo AB (2019) Multi-isotope investigations of ungulate bones and teeth from El Castillo and Covalejos caves (Cantabria, Spain): implications for paleoenvironment reconstructions across the Middle-Upper Palaeolithic transition. J Archaeol Sci Rep 23:1029–1042
Jones JR, Richards MP, Straus LG, Reade H, Altuna J, Mariezkurrena K, Marín-Arroyo AB (2018) Changing environments during the middle-upper Palaeolithic transition in the eastern Cantabrian region (Spain): direct evidence from stable isotope studies on ungulate bones Scientific reports 8:1–20
Kays R, Feranec RS (2011) Using Stable Carbon Isotopes to Distinguish Wild from Captive Wolves Northeastern Naturalist 18:253–264. https://doi.org/10.1656/045.018.0301
Kidawa D, Kowalczyk R (2011) The effects of sex, age, season and habitat on diet of the red fox Vulpes vulpes in northeastern Poland Acta Theriol (Warsz) 56:209–218. https://doi.org/10.1007/s13364-011-0031-3
Kind C-J (2003) Die absolute Datierung des Magdaléniens und des Mesolithikums in Süddeutschland Erkenntnisjäger Kultur und Umwelt des Frühen Menschen Festschrift D Mania Veröffentlichungen des Landesamtes für Archäologie Sachsen-Anhalt Landesmuseum für Vorgeschichte 57:303–219
Kitagawa K (2014) Exploring hominins and animals in the Swabian Jura: study of the Paleolithic fauna from Hohlenstein-Stadel. Unpublished Doctoral Dissertation: University of Tübingen
Kitagawa K, Krönneck P, Conard NJ, Münzel SC (2012) Exploring cave use and exploitation among cave bears, carnivores and hominins in the Swabian Jura, Germany. J Taphonomy 10:439–461
Korner-Nievergelt F, Roth T, Von Felten S, Guélat J, Almasi B, Korner-Nievergelt P (2015) Bayesian data analysis in ecology using linear models with R, BUGS, and Stan. Academic Press
Krajcarz M, Krajcarz MT (2014) The red fox (Vulpes vulpes) as an accumulator of bones in cave-like environments International. J Osteoarchaeol 24:459–475
Krönneck P (2012) Die pleistozäne Makrofauna des Bocksteins (Lonetal–Schwäbische Alb). Ein neuer Ansatz zur Rekonstruktion der Paläoumwelt. Ph. D. Dissertation, University of Tübingen
Langguth K, Malina M (2003) Katalog der Ausstellung Eiszeitschmuck - status und Schönheit. In: Köbl S, Conard NJ (eds) Eiszeitschmuck - status und Schönheit, vol Museumsheft 6. Urgeschichtliches Museum, Blaubeuren, pp 93–128
Lipecki G, Wojtal P (2015) Carnivores from the open-air Gravettian site Kraków Spadzista. In: Wojtal P, Wilczyński J, Haynes G (eds) A Gravettian site in Southern Poland: Kraków Spadzista. ISEA PAS, Krakow, pp 117–157
Lloyd HG, Jensen B, Van Haaften J, Niewold F, Wandeler A, Bögel K, Arata A (1976) Annual turnover of fox populations in Europe Zentralblatt für Veterinärmedizin Reihe B 23:580–589
Lord KA, Larson G, Coppinger RP, Karlsson EK (2019) The history of farm foxes undermines the animal domestication syndrome Trends Ecol Evol:1–12. https://doi.org/10.1016/j.tree.2019.10.011
Lykoudi M (2017) Fetzershaldenhöhle faunal analysis. Universität Tübingen, Master's Thesis
Lyman RL (2008) Quantitative paleozoology. Cambridge University Press
MacDonnald DW (1977) On food preference in the red fox mammal review 7:7–23. https://doi.org/10.1111/j.1365-2907.1977.tb00359.x
Merkle JA, Derbridge JJ, Krausman PR (2011) Using stable isotope analysis to quantify anthropogenic foraging in black bears Human–Wildlife Interactions 5:16
Monchot H, Gendron D (2011) Fox exploitation by the paleoeskimo at the tayara site. Nunavik Arctic Anthropology 48:15–32
Morales JI et al (2016) Expanding the geography of the Middle to Upper Palaeolithic transition: Foradada Cave (Calafell, Spain), a new site on the Iberian Mediterranean coastline. Antiquity 351:1–4
Münzel SC (2019) Die jungpleistozäne Großsäugerfauna aus dem Geißenklösterle. In: Conard NJ, Bolus M, Münzel SC (eds) Geißenklösterle: Chronostratigraphie. Paläoumwelt und Subsistenz im Mittel- und Jungpaläolithikum der Schwäbischen Alb. Kerns Verlag, Tübingen, pp 147–327
Münzel SC, Conard NJ (2004a) Cave bear hunting in the Hohle Fels, a cave site in the Ach Valley. Swabian Jura Revue de Paléobiologie 23:877–885
Münzel SC, Conard NJ (2004b) Change and continuity in subsistence during the Middle and Upper Palaeolithic in the Ach Valley of Swabia(south-west Germany) International Journal of Osteoarchaeology 14:225–243. https://doi.org/10.1002/oa.758
Münzel SC, Rivals F, Pacher M, Döppes D, Rabeder G, Conard NJ, Bocherens H (2014) Behavioural ecology of Late Pleistocene bears (Ursus spelaeus, Ursus ingressus): insight from stable isotopes (C, N, O) and tooth microwear Quaternary International 339:148–163
Murdoch JD, Munkhzul T, Buyandelger S, Reading RP, Sillero-Zubiri C (2010) Seasonal food habits of corsac and red foxes in Mongolia and the potential for competition Mammalian Biology - Zeitschrift für Säugetierkunde 75:36–44. https://doi.org/10.1016/j.mambio.2008.12.003
Murray M, Edwards MA, Abercrombie B, St. Clair CC (2015) Poor health is associated with use of anthropogenic resources in an urban carnivore Proceedings of the Royal Society B: Biological Sciences 282
Nagaoka L (2019) Human behavioral ecology and Zooarchaeology. Handbook of Evolutionary Research in Archaeology. Springer, In, pp 231–253
Naito YI, Germonpré M, Chikaraishi Y, Ohkouchi N, Drucker DG, Hobson KA, Edwards MA, Wißing C, Bocherens H (2016) Evidence for herbivorous cave bears (Ursus spelaeus) in Goyet Cave, Belgium: implications for palaeodietary reconstruction of fossil bears using amino acid δ15N approaches. J Quat Sci 31:598–606
Napierala H, Münzel SC, Conard NJ (2014) Die Fauna des Magdalénien vom Hohle Fels. In: Taller A (ed) Das Magdalénien des Hohle Fels. Chronologische Stellung, Lithische Technologie und Funktion der Rückenmesser. Kerns Verlag, Tübingen, pp 275–311
Nelson DE, Angerbjörn A, Lidén K, Turk I (1998) Stable isotopes and the metabolism of the European cave bear Oecologia 116:177–181
Newsome SD, Ralls K, Van Horn JC, Fogel ML, Cypher BL (2010) Stable isotopes evaluate exploitation of anthropogenic foods by the endangered San Joaquin kit fox (Vulpes macrotis mutica). J Mammal 91:1313–1321. https://doi.org/10.1644/09-MAMM-A-362.1
Newsome TM, Dellinger JA, Pavey CR, Ripple WJ, Shores CR, Wirsing AJ, Dickman CR (2015) The ecological effects of providing resource subsidies to predators. Glob Ecol Biogeogr 24:1–11. https://doi.org/10.1111/geb.12236
Niven L (2006) The palaeolithic occupation of Vogelherd cave: Tübingen publications in prehistory
Otárola-Castillo E, Torquato MG (2018) Bayesian statistics in archaeology. Annu Rev Anthropol 47:435–453
Pacher M (2005) Die Verwendung von Bärenzähnen als Schmuck im Paläolithikum Mitteilungen der Kommission für Quartärforschung der Östereichischen Akademie der Wissenschaften 14:135–151
Pacher M, Stuart AJ (2009) Extinction chronology and palaeobiology of the cave bear (Ursus spelaeus) Boreas 38:189–206
Panek M, Budny M (2017) Variation in the feeding pattern of red foxes in relation to changes in anthropogenic resource availability in a rural habitat of western Poland. Mamm Biol 82:1–7. https://doi.org/10.1016/j.mambio.2016.09.002
Peresani M, Cristiani E, Romandini M (2016) The Uluzzian technology of Grotta di Fumane and its implication for reconstructing cultural dynamics in the Middle–Upper Palaeolithic transition of Western Eurasia Journal of human evolution 91:36–56
Pirson S et al (2012) Chronostratigraphic context of the Middle to Upper Palaeolithic transition: recent data from Belgium Quaternary International 259:78–94. https://doi.org/10.1016/j.quaint.2011.03.035
Pleurdeau D, Moncel MH, Pinhasi R, Yeshurun R, Higham T, Agapishvili T, Bokeria M, Muskhelishvili A, le Bourdonnec FX, Nomade S, Poupeau G, Bocherens H, Frouin M, Genty D, Pierre M, Pons-Branchu E, Lordkipanidze D, Tushabramishvili N (2016) Bondi cave and the middle-upper Palaeolithic transition in western Georgia (South Caucasus). Quat Sci Rev 146:77–98
Pulliaines E (1993) Alopex lagopus (Linnaeus, 1758) - Eisfuchs. In: Stubbe M, Krapp F (eds) Handbuch der Säugetiere Europas. Raubsäuger (Teil 1). AULA-Verlag, Wiesbaden, pp 195–214
Reshamwala HS, Shrotriya S, Bora B, Lyngdoh S, Dirzo R, Habib B (2018) Anthropogenic food subsidies change the pattern of red fox diet and occurrence across trans-Himalayas, India. J Arid Environ 150:15–20. https://doi.org/10.1016/j.jaridenv.2017.12.011
Richard M, Falguères C, Pons-Branchu E, Richter D, Beutelspacher T, Conard N, Kind C-J (2019) The Middle to Upper Palaeolithic transition in Hohlenstein-Stadel cave (Swabian Jura, Germany): A comparison between ESR, U-series and radiocarbon dating,Quaternary International https://doi.org/10.1016/j.quaint.2019.04.009
Riek G (1973) Das Paläolithikum der Brillenhöhle bei Blaubeuren (Schwäbische Alb) I. Verlag Müller & Gräff, Stuttgart
Romandini M, Crezzini J, Bortolini E, Boscato P, Boschin F, Carrera L, Nannini N, Tagliacozzo A, Terlato G, Arrighi S, Badino F, Figus C, Lugli F, Marciani G, Oxilia G, Moroni A, Negrino F, Peresani M, Riel-Salvatore J, Ronchitelli A, Elena Spinapolice E, Benazzi S (2019) Macromammal and bird assemblages across the late Middle to Upper Palaeolithic transition in Italy: an extended zooarchaeological review, Quaternary International, https://doi.org/10.1016/j.quaint.2019.11.008
Roth JD (2003) Variability in marine resources affects arctic fox population dynamics. J Anim Ecol 72:668–676. https://doi.org/10.1046/j.1365-2656.2003.00739.x
Savory G, Hunter C, Wooller M, O’Brien D (2014) Anthropogenic food use and diet overlap between red foxes (Vulpes vulpes) and arctic foxes (Vulpes lagopus) in Prudhoe Bay, Alaska. Can J Zool 92:657–663
Schmidt I, Zimmermann A (2019) Population dynamics and socio-spatial organization of the Aurignacian: Scalable quantitative demographic data for western and central Europe. PLoS One 14:e0211562
Sidorovich VE, Sidorovich AA, Izotova IV (2006) Variations in the diet and population density of the red fox Vulpes vulpes in the mixed woodlands of northern Belarus Mammalian Biology - Zeitschrift für Säugetierkunde 71:74–89. https://doi.org/10.1016/j.mambio.2005.12.001
Soe E, Davison J, Süld K, Valdmann H, Laurimaa L, Saarma U (2017) Europe-wide biogeographical patterns in the diet of an ecologically and epidemiologically important mesopredator, the red fox Vulpes vulpes: a quantitative review. Mammal Rev 47:198–211. https://doi.org/10.1111/mam.12092
Starkovich BM (2012) Intensification of small game resources at Klissoura Cave 1 (Peloponnese, Greece) from the Middle Paleolithic to Mesolithic Quaternary international 264:17–31
Starkovich BM (2014) Optimal foraging, dietary change, and site use during the Paleolithic at Klissoura Cave 1 (southern Greece). J Archaeol Sci 52:39–55
Stephens DW, Krebs JR (1986) Foraging theory. Princeton University Press
Stiner M (2009) Prey choice, site occupation intensity & economic diversity in the Middle–early Upper Palaeolithic at the Üçağizli Caves, Turkey. Before Farming 2009:1–20
Stiner MC (1999) Cave bear ecology and interactions with Pleistocene humans Ursus:41–58
Stiner MC, Munro ND, Surovell TA (2000) The tortoise and the hare: small-game use, the broad-spectrum revolution, and Paleolithic demography. Curr Anthropol 41:39–79
Stiner MC, Munro ND, Surovell TA, Tchernov E, Bar-Yosef O (1999) Paleolithic population growth pulses evidenced by small animal exploitation. Science 283:190–194
Street M, Turner E (2013) The faunal remains from Gönnersdorf. Verlag des Römisch-Germanischen Zentralmuseums Mainz, Mainz
Taller A, Bolus M, Conard N (2014) The Magdalenian of Hohle Fels Cave and the Resettlement of the Swabian Jura after the LGM Modes de contacts et de déplacements au Paléolithique eurasiatique/Modes of contact and mobility during the Eurasian Palaeolithic ERAUL 140:383–399
Taller A, Conard NJ (2019) Transition or replacement? Radiocarbon Dates from Hohle Fels Cave (Alb-Donau-Kreis/D) and the Passage from Aurignacian to Gravettian Archäologisches Korrespondenzblatt 49:165–181
Tchernov E (1994) An early Neolithic Village in the Jordan Valley part II: the Fauna of Netiv Hagdud vol bulletin 44. Peabody Museum of Archaeology and Ethnology, American School of Prehistoric Research Cambridge
Thulin C, Flux J (2003) Lepus timidus Linnaeus, 1758–Schneehase. In: Krapp F, Niethammer J (eds) Handbuch der Säugetiere Europas Band, vol 3. AULA, pp 155–185
Valensi P (1991) Étude des stries de boucherie sur les ossements de cerf élaphe des niveaux supérieurs de la grotte du Lazaret (Nice, Alpes-Maritimes). L'anthropologie 85:797–829
Van Haaften J Fox ecology studies in the Netherlands. In: Trans International Congress of Game Biologists, 1970. pp. 539–543
Wandeler AI, Lüps P (1993) Vulpes vulpes (Linnaeus, 1758) - Rotfuchs. In: Stubbe M, Krapp F (eds) Handbuch der Säugetiere Europas. Raubsäuger (Teil 1). AULA-Verlag, Wiesbaden, pp 139–193
Warsen SA, Frair JL, Teece MA (2014) Isotopic investigation of niche partitioning among native carnivores and the non-native coyote (Canis latrans). Isot Environ Health Stud 50:414–424. https://doi.org/10.1080/10256016.2014.897946
Watson A, Hewson R (1973) Population densities of mountain hares (Lepus timidus) on western Scottish and Irish moors and on Scottish hills. J Zool 170:151–159
Watson A, Hewson R, Jenkins D, Parr R (1973) Population densities of mountain hares compared with red grouse on Scottish moors Oikos:225–230
Wehrberger K (2007) Der Löwenmensch vom Hohlenstein-Stadel. In: floss H, Rouquerol N (eds) Das Aurignacien und die Anfänge der Kunst in Europa, Internationale Fachtagung Aurignac, 16-18 September 2005. Editions Musee-forum Aurignac, pp 331–344
Weinstock J (1999) The upper Pleistocene mammalian fauna from the Große Grotte near Blaubeuren (southwestern Germany) Stuttgarter Beiträge zur Naturkunde Ser B 277:1–50
Weniger G-C (1987a) Magdalenian settlement and subsistence in South-west Germany Proceedings of the Prehistoric Society 53:293–307. https://doi.org/10.1017/S0079497X0000623X
Weniger G-C (1987b) Magdalenian settlement pattern and subsistence in Central Europe. In: Soffer O (ed) The Pleistocene Old World: regional perspectives. Springer US, Boston, MA, pp 201–215. https://doi.org/10.1007/978-1-4613-1817-0_13
West CF, Yeshurun R (2019) Foxes and humans at the Late Holocene Uyak site, Kodiak. Alaska Arctic Anthropology 56:39–51
Wikenros C, Aronsson M, Liberg O, Jarnemo A, Hansson J, Wallgren M, Sand H, Bergström R (2017) Fear or food – abundance of red fox in relation to occurrence of lynx and wolf Scientific Reports 7:9059. https://doi.org/10.1038/s41598-017-08927-6
Wilczyński J, Wojtal P, Robličková M, Oliva M (2015) Dolní Věstonice I (Pavlovian, the Czech Republic) – results of zooarchaeological studies of the animal remains discovered on the campsite (excavation 1924–52) Quaternary International 379:58–70. https://doi.org/10.1016/j.quaint.2015.05.059
Wißing C, Rougier H, Baumann C, Comeyne A, Crevecoeur I, Drucker DG, Gaudzinski-Windheuser S, Germonpré M, Gómez-Olivencia A, Krause J, Matthies T, Naito YI, Posth C, Semal P, Street M, Bocherens H (2019) Stable isotopes reveal patterns of diet and mobility in the last Neandertals and first modern humans in Europe Scientific Reports 9. https://doi.org/10.1038/s41598-019-41033-3
Wißing C, Rougier H, Crevecoeur I, Germonpré M, Naito YI, Semal P, Bocherens H (2016) Isotopic evidence for dietary ecology of late Neandertals in North-Western Europe Quaternary International 411:327–345. https://doi.org/10.1016/j.quaint.2015.09.091
Wojtal P, Wilczyński J (2015) Zooarchaeological studies of large mammal remains from Kraków Spadzista site-trench C2 and trech E1 (2011-2012 excatations). In: Wilczyński J, Haynes G (eds) Wojtal P. A Gravettian site in Southern Poland, Kraków Spadzista ISEA PAS, Krakow, pp 93–111
Wojtal P, Wilczyński J, Bocheński ZM, Svoboda JA (2012) The scene of spectacular feasts: animal remains from Pavlov I south-east, the Czech Republic Quaternary International 252:122–141
Wong GL, Starkovich BM, Conard NJ (2017) Human subsistence and environment during the Magdalenian at Langmahdhalde: evidence from a new rock shelter in the Lone Valley, Southwest Germany Mitteilungen der Gesellschaft für Urgeschichte 26:103
Wong GL, Starkovich BM, Drucker DG, Conard NJ (in press) New perspectives on human subsistence during the Magdalenian in the Swabian Jura, Germany. Archaeological and Anthropological Sciences, 10.1007/s12520-020-01119-w.
Wroth K, Cabanes D, Marston JM, Aldeias V, Sandgathe D, Turq A, Goldberg P, Dibble HL (2019) Neanderthal plant use and pyrotechnology: phytolith analysis from roc de Marsal. France Archaeol Anthrop Sci 11:4325–4346
Yeshurun R, Bar-Oz G, Weinstein-Evron M (2009) The role of foxes in the Natufian economy: a view from Mount Carmel, Israel Before Farming 2009/1:1–15
Yravedra J, Rubio-Jara S, Panera J, van der Made J, Pérez-González A (2019) Neanderthal diet in fluvial environments at the end of the middle Pleistocene/early Late Pleistocene of PRERESA site in the Manzanares Valley (Madrid, Spain). Quat Int 520:72–83
Acknowledgements
We would like to thank Maria Malina and Alexander Janas for providing the excavation databases and data analysis support as well as the two anonymous reviewers. This research was supported by a UNESCO World Cultural Heritage doctoral fellowship funded by Alb-Donau County and the Heritage Authority of Baden-Württemberg, the Senckenberg Nature Research Society and the University of Tübingen. G. Wong’s research on the faunal remains from Langmahdhalde is funded by the Ministerium für Wissenschaft, Forschung und Kunst Baden-Württemberg as part of the Evolution of Cultural Modernity Project at the University of Tübingen.
Funding
Open Access funding provided by Projekt DEAL. This research was supported by a UNESCO World Cultural Heritage doctoral fellowship (CB) funded by Alb-Donau County and the Heritage Authority of Baden-Württemberg, the Senckenberg Nature Research Society and the University of Tübingen. GLW’s research on the faunal remains from Langmahdhalde is funded by the Ministerium für Wissenschaft, Forschung und Kunst Baden-Württemberg as part of the Evolution of Cultural Modernity Project at the University of Tübingen.
Author information
Authors and Affiliations
Contributions
The idea for this study originated from BMS, NJC and CB. The design of the study was discussed jointly by CB, BMS, SCM, GLW and NJC. Research and statistical analyses were conducted by CB. GLW analysed the new data for Langmahdhalde and SCM for Helga Abri. CB wrote the manuscript with the help of GLW. BMS, SCM and NJC provided helpful comments in the discussion. All authors contributed improvements and comments for the text flow. Language corrections were made by GLW and BMS.
Corresponding author
Ethics declarations
Conflicts of interest/competing interests
The authors declare that they have no competing interests.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable
Code availability
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the topical collection on Do good things come in small packages?
Electronic supplementary material
ESM 1
(PDF 1222 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Baumann, C., Wong, G.L., Starkovich, B.M. et al. The role of foxes in the Palaeolithic economies of the Swabian Jura (Germany). Archaeol Anthropol Sci 12, 208 (2020). https://doi.org/10.1007/s12520-020-01173-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-020-01173-4