
Abstract
Traumatic brain injuries (TBI) commonly occur following head trauma. TBI may result in short- and long-term complications which may lead to neurodegenerative consequences, including cognitive impairment post-TBI. When investigating the neurodegeneration following TBI, studies have highlighted the role reactive astrocytes have in the neuroinflammation and degeneration process. This review showcases a variety of markers that show reactive astrocyte presence under pathological conditions, including glial fibrillary acidic protein (GFAP), Crystallin Alpha-B (CRYA-B), Complement Component 3 (C3) and S100A10. Astrocyte activation may lead to white-matter inflammation, expressed as white-matter hyperintensities. Other white-matter changes in the brain following TBI include increased cortical thickness in the white matter. This review addresses the gaps in the literature regarding post-mortem human studies focussing on reactive astrocytes, alongside the potential uses of these proteins as markers in the future studies that investigate the proportions of astrocytes in the post-TBI brain has been discussed. This research may benefit future studies that focus on the role reactive astrocytes play in the post-TBI brain and may assist clinicians in managing patients who have suffered TBI.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Traumatic brain injuries (TBI) occur in up to 70 million individuals per year (Dewan et al. 2019), and may cause many debilitating consequences. The brain is commonly injured following fractures to the facial region due to the thinness of many of the bones in this area (Kühnel and Reichert 2015; Murray et al. 2017). The most common causes of facial fractures include motor vehicle accidents, assaults and falling injuries (Kühnel and Reichert 2015; Murray et al. 2017; Povolotskiy et al. 2019; Fernandes et al. 2022; Grill et al. 2022), with recent studies showing that from 30 to 55% of facial fractures had resulted in a TBI, with a significantly larger number of males sustaining these facial fractures compared to females (Rajandram et al. 2014; Fernandes et al. 2022; Tsur et al. 2023). When bones in the facial region are fractured, the common areas of the brain to sustain damage are the temporal and frontal lobes (Yang et al. 2021; Tsur et al. 2023).
TBI in the frontal lobes are associated with a variety of complications ranging from short to long-term, with short-term complications including cerebrospinal fluid (CSF) leaks when the skull base is damaged, increased intracranial pressure and risk of haemorrhage (Gatzinsky et al. 2020), and death (Tavarez et al. 2012; Shah et al. 2017; Nasi et al. 2018; Burns et al. 2019; Varshneya et al. 2019; Adatia et al. 2021; Choi and Kim 2022; Pellot and De Jesus 2023). Long-term side effects of TBI may include cognitive impairments (Faulkner et al. 2020; Sullivan et al. 2020; Rostowsky and Irimia 2021; Cao et al. 2022; Zhang et al. 2022), alongside a growing base of literature discussing the impact TBI may have on the development of neurodegenerative diseases such as Alzheimer’s Disease (AD), Parkinson’s, and Amyotrophic Lateral Sclerosis (Gu et al. 2022). The link between AD and certain TBI conditions is much more clearly defined, with Chronic Traumatic Encephalopathy and severe TBI having a large amount of literature showing links with AD pathology and neurodegeneration (Washington et al. 2016; Shao et al. 2022). However, for single mild TBI (mTBI) the link is yet to be completely established, with one case–control study with a sample of 65 finding no links between mTBI and dementia or AD risk (Tsolaki et al. 1997), alongside a more recent 12-year follow-up study with 4144 participants finding no links between TBI with loss of consciousness and dementia or AD (Grasset et al. 2023). However, several recent reviews have concluded there may be links between mTBI and neurodegenerative pathology, with mTBI resulting in AD pathology, including increased amyloid-beta plaques, neurofibrillary tangles, Tau and Lewy pathology (Washington et al. 2016; Wilson et al. 2017; Ramos-Cejudo et al. 2018).
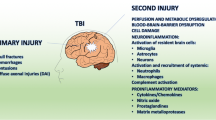
Severe TBI commonly results in neuroinflammation, which activates a variety of immune cells in the brain, including microglia, oligodendrocytes and astrocytes (Nikam et al. 2021). Microglia and astrocytes upregulate and downregulate a variety of proteins to perform various functions (Yuan and Wu 2022). Under pathological conditions, they undergo a shift in morphology and gene expression regulation patterns to perform neuroprotective and neuroinflammatory function in the brain, including the formation of a glial scar to section off injuries, and repair of synapses and the blood–brain barrier (Yuan and Wu 2022).
Recent studies have uncovered different types of reactive astrocytes that are activated following TBI and in neurodegenerative diseases, studied as neurotoxic astrocytes and neuroprotective astrocytes (Liddelow et al. 2017). Astrocytes are initially resting, however, transition into reactive astrocytes following a pathological event; Glial fibrillary acidic protein (GFAP) gene expression is higher in reactive astrocytes, and is commonly used as a marker for reactive astrocytes (Yuan and Wu 2022). Reactive astrocytes upregulate specific proteins to increase or decrease inflammation; neurotoxic astrocytes that increase inflammation have been linked to a compound called Complement Component 3 (C3) which causes apoptosis in surrounding cells (Clark et al. 2019), while other compounds such as Crystallin Alpha-B and S100A100 are neuroprotective in nature and have been linked to neuroprotective astrocytes (King et al. 2020; Hou et al. 2022). Whilst astrocytes themselves may not be visible using neuroimaging techniques, the accompanying neuroinflammation can be seen using a variety of imaging techniques.
Radiographic studies have investigated the changes in the frontal white matter following a TBI using Magnetic Resonance Imaging (MRI) and found hyperintensities in the white matter following TBI, which is associated with impaired cognitive abilities (Zhang et al. 2022). White-matter hyperintensities (WMHs) reveal areas of higher blood flow and are common under pathological conditions due to neuroinflammation (Zhang et al. 2022). Other changes in cognitive abilities which may result from damaged white matter of the brain include acute cognitive impairment and attention deficits in young adults (Cao et al. 2022).
Studies on the brain using Angio-Computed Tomography (Angio-CT) have found a variety of cerebrovascular injuries, including cerebral contusions, cerebral venous sinus injuries and cerebral haemorrhages (Haddad et al. 2022; Chtara et al. 2023). These cerebrovascular injuries have also been discussed to contribute to AD pathology and neurodegeneration in the brain following the TBI (Ramos-Cejudo et al. 2018). Cerebrovascular consequences and reactive astrocytes may all play their own part in the neurodegeneration of the brain and complications which follow TBI, and are all of importance when considering treatments (Zhou et al. 2020).
This review aims to identify the gaps in the current literature regarding TBI effect on the immunohistochemical profile of the brain, alongside establishing a link between TBI, neuroinflammation, reactive astrocyte activation and changes in white matter. Furthermore, future areas of research are discussed regarding reactive astrocytes and areas which are yet to be investigated regarding TBI, neuroinflammation and astrocytes.
Traumatic brain injuries and head fractures
Aetiology and specific areas
TBI commonly result from fractures in the neurocranium and frequently occurs following blunt-force trauma to the head (Kühnel and Reichert 2015). Maxillofacial fractures are one of the most common consequences of blunt-force trauma to the head (Kühnel and Reichert 2015). Approximately 30–55% of maxillofacial fractures will also result in TBI (Rajandram et al. 2014; Fernandes et al. 2022; Tsur et al. 2023). The most common causes of TBI are motor vehicle accidents, however, other notable causes include falls, assault, sport-related injuries, industry-related injuries and pedestrian injuries (Rajandram et al. 2014; You et al. 2018). Researchers have also investigated how the severity of facial fractures was related to TBI and found that as the severity of the facial fractures increased, so did the severity of TBI and less favourable TBI outcomes 3 months following TBI (You et al. 2018).
Whilst a variety of bones are in the maxillofacial region, one bone of interest that has been discussed in previous literature is the ethmoid bone, which forms the roof of the nasal cavity with the cribriform plate portion (Kühnel and Reichert 2015; Murray et al. 2017). These fractures which are commonly associated with TBI may have a variety of both short and long-term complications which are of interest to clinicians managing their patients.
Complications of TBI
Short-term complications
TBI may be associated with various short-term complications depending on the severity of the fracture and the immune response, such as CSF leaks resulting from a damaged meningeal layer in the skull, and may also result in cerebral haemorrhaging (Shah et al. 2017). For both of these complications increased intracranial pressure may occur, which can cause headaches, blurred vision, stroke and death (Shah et al. 2017). As previously mentioned, depending on the severity of the brain injuries, this may lead to an immediate immune response driven by astrocytes and other glial cells in the brain (Clark et al. 2019). This response occurs as a result of complicated fractures, such as frontal nasal-sinus fractures, which may contribute to CSF leaks and transmission of pathogens into the neurocranium through the nasal cavity (Zele and Dewaele 2016; Alhusain and Aladwani 2019; Rotter et al. 2020). Such pathogens may result in pneumocephalus, a condition where there is increased air in the neurocranium, which commonly occurs in the frontal region following nasal injuries (Yoneoka et al. 2020). When the frontal lobe is damaged due to a TBI, complications may include cerebral abscesses in the frontal lobe (Chtara et al. 2023), pulmonary embolism in the pulmonary arteries (Haddad et al. 2022), or seizures (Son Nguyen et al. 2016), alongside psychological symptoms such as loss of consciousness and confusion which may last from short to long term (Son Nguyen et al. 2016; Thuy-My et al. 2019).
Long-term complications
Studies have discussed the long-term impact of TBI on the brain, and generally, the symptoms differ based on a variety of factors, including the severity of the injury and the immune response following the incident, which both may play a role in the potential long-term neurodegenerative changes in the brain (Rostowsky and Irimia 2021).
Long-term neurological complications following a TBI have been studied, and long-lasting cognitive impairments such as attention problems post-TBI have been found (Cao et al. 2022), alongside slowed information processing speeds (Rostowsky and Irimia 2021; Zhang et al. 2022). Studies have also found persisting intelligence impairments following TBI in both adults and children, with more severe TBI resulting in more severe cognitive impairments (Königs et al. 2016; Ko et al. 2022). Some studies have tested hyperbaric oxygen treatment on IQ changes and cognitive symptoms following TBI and discovered significant improvements in IQ, memory motor speed and attention (Harch et al. 2017). Furthermore, a variety of larger randomised studies have investigated this relationship and found cognitive and symptomatic improvements following the application of hyperbaric oxygen treatment (Harch 2022). Other researchers have also investigated oestrogen as a potential treatment to reduce TBI-induced inflammation in the brain, specifically finding that oestrogen treatment inhibited neuroinflammation by preventing neurotoxic astrocyte pathways in the brain, however further research is required to deduce the neuroprotective effects that oestrogen may have on the brain and glial cells (Wang et al. 2021).
TBI may result in several further long-term complications to brain health and overall wellbeing (Wilson et al. 2017), with a variety of studies suggesting that neurodegeneration and AD-like pathology resulting from a severe TBI are quite common (Giunta et al. 2012; Heppner et al. 2015; Washington et al. 2016; Wilson et al. 2017; Ramos-Cejudo et al. 2018; Agrawal et al. 2022). Studies have found links between TBI and amyloid-beta plaques, neurofibrillary tangles, Tau and Lewy body pathology (Washington et al. 2016; Wilson et al. 2017; Agrawal et al. 2022). Furthermore, some studies specifically found loss of consciousness (LOC) post-TBI to be a determining factor for neurodegeneration, specifically finding TBI with LOC was associated with increased amyloid-β load (Agrawal et al. 2022) and hippocampal pTau (Postupna et al. 2021). Another study investigated how acute cognitive impairment following TBI may be linked with neurodegeneration, and found that as a degree of cognitive impairment post-TBI increases, so does the likelihood of neurodegenerative brain atrophy (Rostowsky and Irimia 2021). Whilst the specific factors that cause neurodegeneration following TBI are still under debate, understanding the functional changes that occur in the brain after a TBI may provide knowledge on how to best treat the neurodegenerative effects on the brain.
Immunohistochemical changes in the brain following TBI
Role of microglial cells and astrocytes post-TBI
The immune response following a TBI has been investigated with a focus on the microglial changes in a post-TBI brain (Clark et al. 2019), with studies discussing two types of microglial cells which have been labelled as M1 and M2 (Clark et al. 2019; Guo et al. 2022). M1 microglia have been reported to increase inflammation and are neurotoxic, whilst M2 microglia are considered anti-inflammatory and neuroprotective (Guo et al. 2022). These categorisations however have been disused by a variety of researchers, as they do not reflect the complexity of gene expression upregulation that microglia facilitate (Escartin et al. 2021). Whilst microglia perform a variety of functions in the brain following TBI, importantly they activate astrocytes in response to inflammation and pathology, which may drastically alter the immune response that occurs in the brain (Liddelow et al. 2017; Xu et al. 2019; Guo et al. 2022).
The response of astrocytes has similarly been categorised into A1 reactive astrocytes which promote pro-inflammatory responses, are neurotoxic and release a variety of chemicals that increase inflammation in the brain, while A2 reactive astrocytes promote anti-inflammatory responses and elicit neuroprotective effects on the brain (Liddelow et al. 2017; Fan and Huo 2021). However, similar to microglia, this categorisation system may not account for the diverse phenotypes that astrocytes possess, with a variety of upregulation patterns which does not conform to only two phenotypes (Escartin et al. 2021). In this paper and similar to previous studies, reactive astrocytes which under pathological conditions such as TBI or AD, upregulate inflammatory gene expressions will be called neurotoxic astrocytes, and those which upregulate anti-inflammatory gene expressions will be called neuroprotective astrocytes (Liddelow et al. 2017; Zhou et al. 2020).
Many authors have discussed these reactive astrocytes and their connexions with AD. A recent review suggested astrocytes form an important role in synapse recovery and loss following a TBI; releasing proteins such as Hevin which may assist in circuit re-organisation following TBI, and enzymes such as matrix metalloproteinase 3 which breaks down extracellular matrixes and inhibits synapse recovery (Jamjoom et al. 2021). They suggested that reactive astrocytes may be associated with the spread of tau through the brain via trans-synaptic transmission, through the systematic transfer of tau pathology throughout the brain via impaired synapses (Jamjoom et al. 2021). This suggests that the role of reactive astrocytes in breaking down synapses via the upregulation of Matrix metalloproteinase 3 and other pro-inflammatory proteins could assist in the spread of tau, which could increase the likelihood of AD pathology (Jamjoom et al. 2021). These upregulation patterns suggest that astrocytes may play a dual role in neuroinflammation and neurodegeneration following TBI, regulating both the inflammatory and anti-inflammatory responses (Liddelow et al. 2017).
Some studies have found increases in neurotoxic astrocytes in individuals with AD (Liddelow et al. 2017), and similarly have found increases in neurotoxic astrocytes in individuals who have suffered from TBI (Clark et al. 2019). Other studies have discussed a more complex relationship between neurotoxic and neuroprotective astrocytes during AD, due to differing densities of these reactive astrocytes’ gene expressions in different regions of the AD brain (King et al. 2020). Their study found that following a TBI, there were increased neurotoxic astrocytes in the upper frontal cortex, and increased neuroprotective astrocytes in the frontal upper and lower cortexes, alongside the white matter of the brain when compared with controls (King et al. 2020). This shows that the distribution of reactive astrocytes is complex, and further research into the distributional activation of reactive astrocytes in the brain following pathology is required (King et al. 2020). Investigating the variety of genes which may be upregulated by reactive astrocytes under pathological conditions such as AD or post-TBI could provide additional insight into the localised functions of astrocytes following trauma and may advance our knowledge of the complex gene upregulation patterns of reactive astrocytes following trauma to the brain.
Gene expression changes resulting from TBI
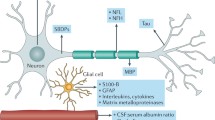
Following TBI, (GFAP is commonly released by glial cells (Becerra-Hernández et al. 2022). GFAP is an intermediate filament protein that is released in higher amounts under pathological conditions and has been discussed to assist in the proliferation of astrocytes from resting to reactive (Yuan and Wu 2022), and thus GFAP is commonly used in studies as a marker for reactive astrocytes (Clark et al. 2019; Escartin et al. 2021; Wang et al. 2021; Becerra-Hernández et al. 2022). A list including many gene expressions that are changed following TBI and under neurodegenerative conditions can be seen in Table 1.
Neurotoxic reactive astrocytes gene expression
Studies have focussed on the secretions of neurotoxic astrocytes and their implications following a TBI and during AD, such as nuclear factor Kappa-B (NF-κB), a protein that is activated following damage to the central nervous system and promotes a variety of pro-inflammatory pathways in the brain (Lawrence et al. 2023). Some researchers have suggested that NF-κB may be upregulated by reactive astrocytes (Kang and Hébert 2011); activating the NF-κB pathway which causes neuroinflammation via the increased transcription of C3 (Lian et al. 2015).
C3 is a protein that is part of the complement system and elicits apoptosis in affected cells (Clark et al. 2019). Researchers have investigated the relationship between Amyloid-β, the NF-κB pathway and C3 production using human control samples with post-mortem AD brains and found higher NF-κB and C3 following exposure to Amyloid-β (Lian et al. 2015). Using lab mice to investigate this further, the researchers concluded that after exposure to Aβ, astrocytes upregulated NF-κB and C3 which caused further increases in Amyloid-β deposits (Lian et al. 2015). Post-mortem human studies have investigated the amount of C3 in the brain of individuals with AD and found 60% of GFAP-positive astrocytes in the pre-frontal cortex of individuals with AD were expressing C3 as well (Liddelow et al. 2017). This fits with previous research regarding GFAP as a marker for reactive astrocytes (Clark et al. 2019; Escartin et al. 2021; Wang et al. 2021; Becerra-Hernández et al. 2022), and signifies that both neurotoxic and neuroprotective astrocytes have their roles in neuroinflammation as previously reported (King et al. 2020). Other studies have supported these findings of increased amounts of C3 in the brains of AD patients, with one study comparing the distributions of neurotoxic and neuroprotective gene expressions in AD patients with reference to controls and found significantly higher C3 in the pre-frontal cortex (King et al. 2020). Studies have also investigated the amount of C3 in the brains of rats following a TBI using a sample of 5 controls with sham injury, and 5 experimental rats with lateral fluid percussion injury (Clark et al. 2019). Their study found increases in the amount of C3 and GFAP in the injured cortex of the brain (Clark et al. 2019). This suggests that astrocytes in the injured cortex following TBI show similar neurotoxic patterns to AD, however, the current literature is yet to investigate the proportion of reactive astrocytes and C3 expression in the post-mortem brain of humans following a TBI.
Neuroprotective reactive astrocyte gene expression
S100A10 is a calcium-binding protein that promotes neuronal survival and growth (Arranz and Strooper 2019) and assists in intracellular neurotransmitter systems (Rescher and Gerke 2008). Various studies have found that S100A10 is upregulated by neuroprotective astrocytes due to its neuroprotective functions, and it is commonly used as a marker to determine quantities of astrocytes upregulating anti-inflammatory gene expressions (Liddelow et al. 2017; Arranz and Strooper 2019; King et al. 2020). A recent study that examined the distribution of neuroprotective astrocytes in the human brain by immunostaining for S100A10 found low immunopositivity in their control cases for both the white matter and the cortex (King et al. 2020). However, S100A10 expression was increased in the white matter in AD compared to controls, which led researchers to believe that neuroprotective astrocytes were also activated in these areas for individuals with AD (King et al. 2020). Whilst research has investigated S100A10 under neurodegenerative conditions such as AD, we were unable to find any studies investigating the release of S100A10 by reactive astrocytes following TBI. Due to the prior use of S100A10 as a marker for neuroprotective astrocytes (Liddelow et al. 2017; Arranz and Strooper 2019; King et al. 2020), future studies could investigate this protein as a marker for neuroprotective astrocytes following TBI.
Another protein that is upregulated by neuroprotective reactive astrocytes is CRYA-B, which assists in maintaining homeostasis in the cell, alongside protecting the cell from oxidative stress, promoting cell survival pathways, and preventing apoptosis via inhibition of caspase activity (Kuipers et al. 2017; Becerra-Hernández et al. 2022). CRYA-B has been suggested to exhibit neuroprotective effects on the brain following trauma (Shao et al. 2013; Guo et al. 2019; Saglam et al. 2021), and thus has been suggested to be upregulated by neuroprotective reactive astrocytes (Hou et al. 2022) and may be useful for the identification of neuroprotective reactive astrocytes.
Changes in neuronal cells following TBI
One recent study found that following a TBI, CRYA-B and GFAP were overexpressed in contused tissue which was collected from the frontal and temporal lobes (Becerra-Hernández et al. 2022), with the researchers suggesting this as a marker for reactive astrogliosis, and neuroprotective astrocyte activation in contused tissue. Another study investigated cortical and white-matter changes in the brain following semi-recent severe TBI and found “ballooned” neurons in the cortex and white matter which also expressed CRYA-B, which likely signified the presence of CRYA-B upregulating neuroprotective astrocytes in these damaged neurons following severe TBI (Michaud et al. 2023).
Interestingly, a recent study investigated the morphology of neurotoxic and neuroprotective astrocytes and found neurotoxic astrocytes which express C3 exhibited elongated dendrites, however, neuroprotective astrocytes which express S100A10 showed fewer dendrites and increased cell hypertrophy (Zong et al. 2020; Fan and Huo 2021). This morphological finding of neuroprotective astrocytes exhibiting hypertrophy may be supported by the previous study which found “ballooned” neurons in the presence of CRYA-B, an upregulated protein via neuroprotective astrocytes (Zong et al. 2020; Fan and Huo 2021; Hou et al. 2022; Michaud et al. 2023). Perhaps the neuroprotective astrocytes exhibit effects on the surrounding neurons to change their structure and size, and CRYA-B may perform a role in these changes (Becerra-Hernández et al. 2022). These “ballooned” neurons may be related to MRI studies which have found increases in cortical thickness in the white matter following TBI (Mazaharally et al. 2022), however additional studies that use both histology and neuroimaging are required to verify any relationships.
Diagnostic methods and neuronal cell inflammation following TBI
The changes in the brain following a TBI are complex and have been investigated using various radiographical studies over the recent years, this includes investigating the impact that TBI may have on brain function using MRI, functional MRI (fMRI) and Angio-CT (Ramos-Cejudo et al. 2018). Mazaharally et al. (2022) investigated cortical changes in the brain following TBI using MRI scans from 67 individuals with clinically confirmed TBI taken at 4-time points post-TBI ranging from 5 months to 7 years, compared with 18 controls whose scans were taken at 2-time points. Their study found that the post-TBI individuals showed increased cortical thickness in the frontal regions however cortical thickness reductions in the posterior cingulate/precuneus regions which can be seen in Fig. 1 (Mazaharally et al. 2022). This shows regional differences in response to TBI, and further investigation into this topic is required alongside histological studies to investigate the role that astrocytes may play in changing the cortical morphometry post-TBI.

Effect of time on cortical thickness in TBI group. Coloured areas represent the location of vertices that demonstrate a significant change in cortical thickness over time (10% false discovery rate [FDR] corrected), where darker shades reflect the most substantial changes (see colour bar). Cortical thickness at single vertices indicated by crosshairs on the cortical surfaces above are plotted to illustrate cortical thickness changes over time. a Regions of the cortex decrease in thickness over time. b Regions of the cortex increase in thickness over time (Mazaharally et al. 2022)
Another study used MRI and fMRI on a sample of 46 patients who suffered mTBI matched with 46 healthy controls to investigate changes in white-matter neurons which may be caused by TBI (Zhang et al. 2022). Their study found increased WMH volume in the frontal and parietal lobes, which correlated with poorer information processing speed via tests such as the digital symbol coding test and trail-making test (Zhang et al. 2022). Similarly, white WMHs in the frontal lobe were associated with significantly lower functional connectivity in areas such as the right middle temporal gyrus, left middle frontal gyrus, left anterior cingulate cortex, and the right superior frontal gyrus (Zhang et al. 2022). The researchers postulated that the frontal white-matter damage, impaired the functional connectivity of the default mode network as seen in Fig. 2, resulting in poorer information processing speed and cognitive impairment (Zhang et al. 2022).

The blue represents regions of reduced resting-stated functional connectivity in the patients with high frontal WMH volume (Patient-A) compared with patients with low frontal WMH volume (Patient-B) and the control. Cluster extent threshold of P value of 0.01 using an Family-Wise Error (FWE) correction for multiple comparisons (Zhang et al. 2022)
Researchers have used fMRI to compare brain patterns and cognitive impairments between three groups, one mTBI group with 33 patients, one AD group with 66 patients, and healthy controls with 81 patients (Rostowsky and Irimia 2021). Their study found that the degree of acute cognitive impairment 48 h post-injury (measured using the Montréal Cognitive Assessment), was correlated with the severity of brain atrophy patterns found in AD (Rostowsky and Irimia 2021). Contrarily, a recent article investigated the long-term impact of TBI with loss of consciousness, and while an association was found between TBI with loss of consciousness and lower white-matter volumes, no association was found between TBI history and white-matter WMHs and other brain volumes (Grasset et al. 2023).
Other studies have investigated the relationship between young adults having a TBI and hyperactive/impulsive behaviours (Cao et al. 2022). In their study, 89 participants aged between 18 to 26 years were included, 44 participants had suffered from TBI and 45 were group-matched healthy controls (Cao et al. 2022). The participants who had suffered TBI showed significantly altered neurite densities in the bilateral parietal and frontal lobes of the grey matter, significantly decreased neurite densities in the left precentral gyrus and superior longitudinal fasciculus, alongside changes in the white matter which is connected to these areas (Cao et al. 2022). Their study also found a strong relationship between decreases in neurite densities in the left precentral gyrus and superior longitudinal fasciculus and hyperactive/impulsive behaviours, as measured by Conner’s Adult ADHD Self-Reporting Scale (Cao et al. 2022).
MRI and Angio-CT have been utilised to view the cerebrovascular impact of TBI. One study found that a variety of cerebrovascular impairments following TBI such as post-traumatic cerebral venous thrombosis, lateral and sigmoid sinus impairments and thrombosis in the jugular vein (Chtara et al. 2023), while another study found TBI commonly leads to post-traumatic pulmonary embolism, cerebral contusions, and subdural/extradural haemorrhages (Haddad et al. 2022). Some post-TBI studies have implicated cerebrovascular injuries as having a role in expediting AD pathology (Ramos-Cejudo et al. 2018). This pathology is suggested to increase due to endothelial cells and blood-–brain barrier damage which may inhibit the clearance of waste in the brain and increase tau pathology, alongside cerebrovascular inflammation which may cause neurotoxic glial activation such as neurotoxic astrocytes (Ramos-Cejudo et al. 2018). The immune response, initiated by reactive astrocytes following cerebrovascular inflammation plays an important role in the neurodegeneration which may result from TBI, and these different areas should all be investigated due to the potential therapeutic benefits that may be gained (Zhou et al. 2020).
Conclusion
Whilst a variety of studies have investigated TBI and neurodegeneration, whether a single mTBI results in an increased risk of AD is still in debate, a variety of studies suggest that other factors may be at play such as loss of consciousness at the time of TBI, and cognitive impairment following a TBI. The microglial and astrocyte changes in the brain during AD and following TBI are of increasing interest in recent years, and while studies have investigated the impact of neurotoxic astrocytes post-TBI by analysing C3 in post-TBI mice, no studies yet have investigated C3 in post-mortem human samples. Similarly, the exact distribution of neurotoxic and neuroprotective astrocytes following TBI is unknown. Due to the variety of proteins released by neurotoxic and neuroprotective astrocytes, further studies investigating the variety of gene expressions may assist in discovering the roles of neurotoxic and neuroprotective astrocytes post-TBI, alongside seeing the distributions of these reactive astrocytes in different portions of the brain. This could gain insight into the immunological changes in the brain that occur as a result of TBI. Furthermore, additional studies could explore potential sub-groups of the neurotoxic and neuroprotective reactive astrocytes, with different upregulation and downregulation patterns. This review may provide insight into the functions of neurotoxic and neuroprotective astrocytes following TBI, and in the future may build upon the knowledge that results in improved outcomes for patients with short-term and long-term complications following TBI.
References
Adatia K, Newcombe VFJ, Menon DK (2021) Contusion progression following traumatic brain injury: a review of clinical and radiological predictors, and influence on outcome. Neurocrit Care 34(1):312–324
Agrawal S, Leurgans SE, James BD, Barnes LL, Mehta RI, Dams-O’Connor K et al (2022) Association of traumatic brain injury with and without loss of consciousness with neuropathologic outcomes in community-dwelling older persons. JAMA Netw Open 5(4):e229311
Alhusain M, Aladwani N (2019) Incidental spontaneous CSF fistula with pterygoid process meningoencephalocele. Neurosci off J Pan Arab Union Neurol Sci 24(3):231–235
Arranz AM, De Strooper B (2019) The role of astroglia in Alzheimer’s disease: pathophysiology and clinical implications. Lancet Neurol 18(4):406–414
Baecker J, Wartchow K, Sehm T, Ghoochani A, Buchfelder M, Kleindienst A (2020) Treatment with the neurotrophic protein S100B increases synaptogenesis after traumatic brain injury. J Neurotrauma 37(8):1097–1107
Becerra-Hernández LV, Escobar-Betancourt MI, Pimienta-Jiménez HJ, Buriticá E (2022) Crystallin alpha-B overexpression as a possible marker of reactive astrogliosis in human cerebral contusions. Front Cell Neurosci 16:838551
Burns EC, Burns B, Newgard CD, Laurie A, Fu R, Graif T et al (2019) Pediatric minor traumatic brain injury with intracranial hemorrhage: identifying low-risk patients who may not benefit from ICU admission. Pediatr Emerg Care 35(3):161–169
Cao M, Luo Y, Wu Z, Wu K, Li X (2022) Abnormal neurite density and orientation dispersion in frontal lobe link to elevated hyperactive/impulsive behaviours in young adults with traumatic brain injury. Brain Commun. 4(1):fcac011
Chen K, Gu H, Zhu L, Feng D-F (2020) A new model of repetitive traumatic brain injury in mice. Front Neurosci 13:1417
Choi JI, Kim SD (2022) Pediatric minor traumatic brain injury: growing skull fracture, traumatic cerebrospinal fluid leakage. Concussion J Korean Neurosurg Soc 65(3):348–353
Chtara K, Bradai S, Baccouche N, Toumi N, Amar WB, Chelly H et al (2023) Post-traumatic cerebral venous sinus thrombosis in an intensive care unit: A case series of ten patients. J Méd Vasc 48(2):62–68
Clark DPQ, Perreau VM, Shultz SR, Brady RD, Lei E, Dixit S et al (2019) Inflammation in traumatic brain injury: roles for toxic A1 astrocytes and microglial-astrocytic crosstalk. Neurochem Res 44(6):1410–1424
Dewan MC, Rattani A, Gupta S, Baticulon RE, Hung YC, Punchak M et al (2019) Estimating the global incidence of traumatic brain injury. J Neurosurg 130(4):1080–1097
Diniz LP, Almeida JC, Tortelli V, Vargas Lopes C, Setti-Perdigão P, Stipursky J et al (2012) Astrocyte-induced synaptogenesis is mediated by transforming growth factor β signaling through modulation of d-serine levels in cerebral cortex neurons. J Biol Chem 287(49):41432–41445
Escartin C, Galea E, Lakatos A, O’Callaghan JP, Petzold GC, Serrano-Pozo A et al (2021) Reactive astrocyte nomenclature, definitions, and future directions. Nat Neurosci 24(3):312–325
Falo MC, Fillmore HL, Reeves TM, Phillips LL (2006) Matrix metalloproteinase-3 expression profile differentiates adaptive and maladaptive synaptic plasticity induced by traumatic brain injury. J Neurosci Res 84(4):768–781
Fan Y-Y, Huo J (2021) A1/A2 astrocytes in central nervous system injuries and diseases: angels or devils? Neurochem Int 148:105080
Faulkner JW, Theadom A, Mahon S, Snell DL, Barker-Collo S, Cunningham K (2020) Psychological flexibility: a psychological mechanism that contributes to persistent symptoms following mild traumatic brain injury? Med Hypotheses 143:110141
Fernandes TB, Mandrekar PN, Visen A, Sinai Khandeparker PV, Dhupar V, Akkara F (2022) Pattern of associated brain injury in maxillofacial trauma: a retrospective study from a high-volume centre. Br J Oral Maxillofac Surg 60(10):1373–1378
Furman JL, Sompol P, Kraner SD, Pleiss MM, Putman EJ, Dunkerson J et al (2016) Blockade of astrocytic calcineurin/NFAT signaling helps to normalize hippocampal synaptic function and plasticity in a rat model of traumatic brain injury. J Neurosci 36(5):1502–1515
Gatzinsky K, Johansson E, Jennische E, Oshalim M, Lange S (2020) Elevated intracranial pressure after head trauma can be suppressed by antisecretory factor-a pilot study. Acta Neurochir (wien) 162(7):1629–1637
Giunta B, Obregon D, Velisetty R, Sanberg PR, Borlongan CV, Tan J (2012) The immunology of traumatic brain injury: a prime target for Alzheimer’s disease prevention. J Neuroinflamm 9(1):185
Grasset L, Power MC, Crivello F, Tzourio C, Chêne G, Dufouil C (2023) How Traumatic brain injury history relates to brain health MRI markers and dementia risk: findings from the 3C dijon cohort. J Alzheimers Dis 92(1):183–193
Grill FD, Roth C, Zyskowski M, Fichter A, Kollmuss M, Stimmer H et al (2022) E-scooter-related craniomaxillofacial injuries compared with bicycle-related injuries—a retrospective study. J Cranio Maxillofacial Surg 50(9):738–744
Gu D, Ou S, Liu G (2022) Traumatic brain injury and risk of dementia and Alzheimer’s disease: a systematic review and meta-analysis. Neuroepidemiology 56(1):4–16
Guo Y-S, Liang P-Z, Lu S-Z, Chen R, Yin Y-Q, Zhou J-W (2019) Extracellular αB-crystallin modulates the inflammatory responses. Biochem Biophys Res Commun 508(1):282–288
Guo S, Wang H, Yin Y (2022) Microglia Polarization from M1 to M2 in neurodegenerative diseases. Front Aging Neurosci. 14:815347
Haddad IAE, Mouhib AE, Hattab O, Assamti M, Mojahid A, Bkiyer H et al (2022) Early bilateral pulmonary embolism in a polytrauma patient: about a case report. Ann Med Surg 78:103868
Halford J, Shen S, Itamura K, Levine J, Chong AC, Czerwieniec G et al (2017) New astroglial injury-defined biomarkers for neurotrauma assessment. J Cereb Blood Flow Metab 37(10):3278–3299
Harch PG (2022) Systematic Review and dosage analysis: hyperbaric oxygen therapy efficacy in mild traumatic brain injury persistent postconcussion syndrome. Front Neurol 13:815056
Harch P, Andrews S, Fogarty E, Lucarini J, Van Meter K (2017) Case control study: Hyperbaric oxygen treatment of mild traumatic brain injury persistent post-concussion syndrome and post-traumatic stress disorder. Med Gas Res 7(3):156–174
Harris NG, Carmichael ST, Hovda DA, Sutton RL (2009) Traumatic brain injury results in disparate regions of chondroitin sulfate proteoglycan expression that are temporally limited. J Neurosci Res 87(13):2937–2950
Heppner FL, Ransohoff RM, Becher B (2015) Immune attack: the role of inflammation in Alzheimer disease. Nat Rev Neurosci 16(6):358–372
Hindinger C, Bergmann CC, Hinton DR, Phares TW, Parra GI, Hussain S et al (2012) IFN-γ signaling to astrocytes protects from autoimmune mediated neurological disability. PLoS ONE 7(7):e42088
Hou J, Bi H, Ge Q, Teng H, Wan G, Yu B et al (2022) Heterogeneity analysis of astrocytes following spinal cord injury at single-cell resolution. FASEB J 36(8):e22442
Jamjoom AAB, Rhodes J, Andrews PJD, Grant SGN (2021) The synapse in traumatic brain injury. Brain (london, England: 1878) 144(1):18–31
Jing R, Wilhelmsson U, Goodwill W, Li L, Pan Y, Pekny M et al (2007) Synemin is expressed in reactive astrocytes in neurotrauma and interacts differentially with vimentin and GFAP intermediate filament networks. J Cell Sci 120(7):1267–1277
Kang W, Hébert JM (2011) Signaling pathways in reactive astrocytes, a genetic perspective. Mol Neurobiol 43(3):147–154
King A, Szekely B, Calapkulu E, Ali H, Rios F, Jones S et al (2020) The Increased densities, but different distributions, of both C3 and S100A10 immunopositive astrocyte-like cells in Alzheimer’s disease brains suggest possible roles for both A1 and A2 astrocytes in the disease pathogenesis. Brain Sci 10(8):503–516
Ko MSM, Poh P-F, Heng KYC, Sultana R, Murphy B, Ng RWL et al (2022) Assessment of long-term psychological outcomes after pediatric intensive care unit admission. JAMA Pediatr 176(3):e215767
Königs M, Engenhorst PJ, Oosterlaan J (2016) Intelligence after traumatic brain injury: meta-analysis of outcomes and prognosis. Eur J Neurol 23(1):21–29
Kühnel TS, Reichert TE (2015) Trauma of the midface. Laryngorhinootologie 94:S206–S247
Kuipers HF, Yoon J, van Horssen J, Han MH, Bollyky PL, Palmer TD et al (2017) Phosphorylation of αB-crystallin supports reactive astrogliosis in demyelination. Proc Nat Acad Sci PNAS 114(9):E1745–E1754
Lawrence JM, Schardien K, Wigdahl B, Nonnemacher MR (2023) Roles of neuropathology-associated reactive astrocytes: a systematic review. Acta Neuropathol Commun 11(1):42
Lian H, Yang L, Cole A, Sun L, Chiang ACA, Fowler SW et al (2015) NFκB-activated astroglial release of complement C3 compromises neuronal morphology and function associated with Alzheimer’s disease. Neuron (cambridge, Mass) 85(1):101–115
Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L et al (2017) Neurotoxic reactive astrocytes are induced by activated microglia. Nature (london) 541(7638):481–487
Mazaharally M, Stojanovski S, Trossman R, Szulc-Lerch K, Chakravarty MM, Colella B et al (2022) Patterns of change in cortical morphometry following traumatic brain injury in adults. Hum Brain Mapp 43(6):1882–1894
Michaud J, Plu I, Parai J, Bourgault A, Tanguay C, Seilhean D et al (2023) Ballooned neurons in semi-recent severe traumatic brain injury. Acta Neuropathol Commun 11(1):37
Michetti F, D’Ambrosi N, Toesca A, Puglisi MA, Serrano A, Marchese E et al (2019) The S100B story: from biomarker to active factor in neural injury. J Neurochem 148(2):168–187
Moreels M, Vandenabeele F, Dumont D, Robben J, Lambrichts I (2008) Alpha-smooth muscle actin (α-SMA) and nestin expression in reactive astrocytes in multiple sclerosis lesions: potential regulatory role of transforming growth factor-beta 1 (TGF-β1). Neuropathol Appl Neurobiol 34(5):532–546
Murray RD, Friedlander R, Hanz S, Singh H, Anand VK, Schwartz TH (2017) Nonrandom spatial clustering of spontaneous anterior fossa cerebrospinal fluid fistulas and predilection for the posterior cribriform plate. J Neurosurg 126(5):1720–1724
Nasi D, di Somma L, Gladi M, Moriconi E, Scerrati M, Iacoangeli M et al (2018) New or blossoming hemorrhagic contusions after decompressive craniectomy in traumatic brain injury: analysis of risk factors. Front Neurol 9:1186
Nikam RM, Yue X, Kandula VV, Paudyal B, Langhans SA, Averill LW et al (2021) Unravelling neuroinflammation in abusive head trauma with radiotracer imaging. Pediatr Radiol 51(6):966–970
Pellot JE, De Jesus O (2023) Cerebral contusion. StatPearls, Treasure Island
Postupna N, Rose SE, Gibbons LE, Coleman NM, Hellstern LL, Ritchie K et al (2021) The delayed neuropathological consequences of traumatic brain injury in a community-based sample. Front Neurol 12:624696
Povolotskiy R, Youssef P, Kaye R, Paskhover B (2019) Facial fractures in young adults: a national retrospective study. Ann Otol Rhinol Laryngol 128(6):516–523
Rajandram RK, Syed Omar SN, Rashdi MFN, Abdul Jabar MN (2014) Maxillofacial injuries and traumatic brain injury–a pilot study. Dent Traumatol 30(2):128–132
Ramos-Cejudo J, Wisniewski T, Marmar C, Zetterberg H, Blennow K, de Leon MJ et al (2018) Traumatic brain Injury and Alzheimer’s disease: the cerebrovascular link. EBioMedicine 28:21–30
Rescher U, Gerke V (2008) S100A10/p11: family, friends and functions. Pflugers Arch 455(4):575–582
Rostowsky KA, Irimia A (2021) Acute cognitive impairment after traumatic brain injury predicts the occurrence of brain atrophy patterns similar to those observed in Alzheimer’s disease. GeroScience 43(4):2015–2039
Rotter J, Graffeo CS, Perry A, Gilder HE, Wilson JW, Link MJ (2020) Polymicrobial intracerebral abscess growing mycobacterium avium complex and achromobacter xylosoxidans: case report and literature review. World Neurosurg 141:441–7.e1
Saglam A, Calof AL, Wray S (2021) Novel factor in olfactory ensheathing cell-astrocyte crosstalk: anti-inflammatory protein α-crystallin B. Glia 69(4):1022–1036
Shah N, Deopujari CE, Chhapola SS (2017) Spontaneous recurrent CSF rhinorrhoea: a rare case and review of literature. Indian J Otolaryngol Head Neck Surg 69(3):420–424
Shao W, Zhang S-Z, Tang M, Zhang X-H, Zhou Z, Yin Y-Q et al (2013) Suppression of neuroinflammation by astrocytic dopamine D2 receptors via αB-crystallin. Nature (london) 494(7435):90–94
Shao F, Wang X, Wu H, Wu Q, Zhang J (2022) Microglia and neuroinflammation: crucial pathological mechanisms in traumatic brain injury-induced neurodegeneration. Front Aging Neurosci 14:825086
Son Nguyen H, Oni-Orisan A, Doan N, Mueller W (2016) Transnasal penetration of a ballpoint pen: case report and review of literature. World Neurosurg 96:611.e1-.e10
Stym-Popper G, Matta K, Chaigneau T, Rupra R, Demetriou A, Fouquet S et al (2023) Regulatory T cells decrease C3-positive reactive astrocytes in Alzheimer-like pathology. J Neuroinflamm 20(1):64
Sullivan KA, Kaye SA, Blaine H, Edmed SL, Meares S, Rossa K et al (2020) Psychological approaches for the management of persistent postconcussion symptoms after mild traumatic brain injury: a systematic review. Disabil Rehabil 42(16):2243–2251
Tavarez MM, Atabaki SM, Teach SJ (2012) Acute evaluation of pediatric patients with minor traumatic brain injury. Curr Opin Pediatr 24(3):307–313
Thuy-My T, Le RSB, Oleck NC, Dobitsch AA, Halsey JN, Hoppe IC, Lee ES, Granick MS (2019) Patterns of nasoorbitalethmoid fractures in the pediatric population. Am Surg 85(7):730–732
Tsolaki M, Fountoulakis K, Chantzi E, Kazis A (1997) Risk factors for clinically diagnosed Alzheimer’s disease: a case-control study of a greek population. Int Psychogeriatr 9(3):327–341
Tsur N, Talmy T, Radomislensky I, Almog O, Gendler S (2023) Traumatic maxillofacial injuries: patterns, outcomes, and long-term follow-up of a military cohort. Dent Traumatol 39(2):147–156
Tyzack GE, Sitnikov S, Barson D, Adams-Carr KL, Lau NK, Kwok JC et al (2014) Astrocyte response to motor neuron injury promotes structural synaptic plasticity via STAT3-regulated TSP-1 expression. Nat Commun 5(1):4294
Van Zele T, Dewaele F (2016) Traumatic CSF leaks of the anterior skull base. B-ENT 2:19–27
Varshneya K, Rodrigues AJ, Medress ZA, Stienen MN, Grant GA, Ratliff JK et al (2019) Risks, costs, and outcomes of cerebrospinal fluid leaks after pediatric skull fractures: a MarketScan analysis between 2007 and 2015. Neurosurg Focus 47(5):E10
Wang Y, Cheng X, He Q, Zheng Y, Kim DH, Whittemore SR et al (2011) Astrocytes from the contused spinal cord inhibit oligodendrocyte differentiation of adult oligodendrocyte precursor cells by increasing the expression of bone morphogenetic proteins. J Neurosci 31(16):6053–6058
Wang Z-G, Cheng Y, Yu X-C, Ye L-B, Xia Q-H, Johnson NR et al (2016) bFGF protects against blood-brain barrier damage through junction protein regulation via PI3K-Akt-Rac1 pathway following traumatic brain injury. Mol Neurobiol 53(10):7298–7311
Wang J, Hou Y, Zhang L, Liu M, Zhao J, Zhang Z et al (2021) Estrogen attenuates traumatic brain injury by inhibiting the activation of microglia and astrocyte-mediated neuroinflammatory responses. Mol Neurobiol 58(3):1052–1061
Washington PM, Villapol S, Burns MP (2016) Polypathology and dementia after brain trauma: does brain injury trigger distinct neurodegenerative diseases, or should they be classified together as traumatic encephalopathy? Exp Neurol 275:381–388
Wilson L, Stewart W, Dams-O’Connor K, Diaz-Arrastia R, Horton L, Menon DK et al (2017) The chronic and evolving neurological consequences of traumatic brain injury. Lancet Neurol 16(10):813–825
Xu X, Zhang A, Zhu Y, He W, Di W, Fang Y et al (2019) MFG-E8 reverses microglial-induced neurotoxic astrocyte (A1) via NF-κB and PI3K-Akt pathways. J Cell Physiol 234(1):904–914
Yang M, Wu Z, Yu H, Cheng J (2021) Reconstruction for diverse fronto-orbital defects with computer-assisted designed and computer-assisted manufactured PEEK implants in one-stage operation. Medicine 100(40):e27452
Yoneoka Y, Aizawa N, Nonomura Y, Ogi M, Seki Y, Akiyama K (2020) Traumatic nonmissile penetrating transnasal anterior skull base fracture and brain injury with cerebrospinal fluid leak: intraoperative leak detection and an effective reconstruction procedure for a localized skull base defect especially after coronavirus disease 2019 outbreak. World Neurosurgery 140:166–172
You N, Choi MS, Roh TH, Jeong D, Kim S-H (2018) Severe facial fracture is related to severe traumatic brain injury. World Neurosurgery 111:e47–e52
Yuan M, Wu H (2022) Astrocytes in the traumatic brain injury: the good and the bad. Exp Neurol 348:113943
Zhang G, Ma P, Wan S, Xu J, Yang M, Qiu G et al (2019) Dystroglycan is involved in the activation of ERK pathway inducing the change of AQP4 expression in scratch-injured astrocytes. Brain Res 1721:146347
Zhang D, Zhu P, Yin B, Zhao P, Wang S, Ye L et al (2022) Frontal White Matter Hyperintensities Effect on Default Mode Network Connectivity in Acute Mild Traumatic Brain Injury. Front Aging Neurosci. 13:793491
Zhou Y, Shao A, Yao Y, Tu S, Deng Y, Zhang J (2020) Dual roles of astrocytes in plasticity and reconstruction after traumatic brain injury. Cell Commun Signal 18(1):62–16
Zong X, Li Y, Liu C, Qi W, Han D, Tucker L et al (2020) Theta-burst transcranial magnetic stimulation promotes stroke recovery by vascular protection and neovascularization. Theranostics 10(26):12090–12110
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Boulton, M., Al-Rubaie, A. Neuroinflammation and neurodegeneration following traumatic brain injuries. Anat Sci Int (2024). https://doi.org/10.1007/s12565-024-00778-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12565-024-00778-2