
Abstract
Despite fertilization efforts, phosphorus (P) availability in soils remains a major constraint to global plant productivity. Soil incorporation of biochar could promote soil P availability but its effects remain uncertain. To attain further improvements in soil P availability with biochar, we developed, characterized, and evaluated magnesium-oxide (MgO) and sepiolite (Mg4Si6O15(OH)2·6H2O)-functionalized biochars with optimized P retention/release capacity. Field-based application of these biochars for improving P availability and their mechanisms during three growth stages of maize was investigated. We further leveraged next-generation sequencing to unravel their impacts on the plant growth-stage shifts in soil functional genes regulating P availability. Results showed insignificant variation in P availability between single super phosphate fertilization (F) and its combination with raw biochar (BF). However, the occurrence of Mg-bound minerals on the optimized biochars’ surface adjusted its surface charges and properties and improved the retention and slow release of inorganic P. Compared to BF, available P (AP) was 26.5% and 19.1% higher during the 12-leaf stage and blister stage, respectively, under MgO-optimized biochar + F treatment (MgOBF), and 15.5% higher under sepiolite-biochar + F (SBF) during maize physiological maturity. Cumulatively, AP was 15.6% and 13.2% higher in MgOBF and SBF relative to BF. Hence, plant biomass, grain yield, and P uptake were highest in MgOBF and SBF, respectively at harvest. Optimized-biochar amendment stimulated microbial 16SrRNA gene diversity and suppressed the expression of P starvation response and P uptake and transport-related genes while stimulating P solubilization and mineralization genes. Thus, the optimized biochars promoted P availability via the combined processes of slow-release of retained phosphates, while inducing the microbial solubilization and mineralization of inorganic and organic P, respectively. Our study advances strategies for reducing cropland P limitation and reveals the potential of optimized biochars for improving P availability on the field scale.
Graphical Abstract

Article Highlights
-
MgO and sepiolite doping optimized biochar’s surface properties for phosphorus (P) retention and slow release
-
The potential formation of Mg-PO4 phases on the optimized biochar surface regulated P retention and release
-
MgO and sepiolite-ptimized biochars increased soil available P by promoting microbial P mineralization and solubilization.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Phosphorus (P) is an essential element required to optimize plant productivity across terrestrial ecosystems and it has no substitute (Glaser and Lehr 2019; Hou et al. 2021). It is sparingly mobile, and its highly reactive inorganic or available forms are fixed in acidic soils by aluminium and iron, and in alkaline soils by calcium, magnesium, potassium, and sodium, or immobilized by microorganisms (Khan et al. 2021; Zhu et al. 2018). Consequently, a significant quantity (75–90%) of soil-applied P fertilizers are fixed and unavailable for plant use in acidic and alkaline conditions (Vitousek et al. 2009). Continuous P addition without corresponding uptake results in low P use efficiency, P runoff, and water body pollution (Andersson et al. 2013). The loss of P from soils is influenced by various factors including soil type, land management practices, and hydrological processes. Hence, the extent of P limitation on the above-ground productivity of plants remains pervasive across global ecosystems (Hou et al. 2020). As a major staple food cultivated in most parts of the world, the productivity of maize has been greatly impacted by P limitation due to its high P requirement, particularly in tropical soils (Arif et al. 2017; Rafique et al. 2020). Due to the low availability of added P fertilizer for crop use, P fertilization is repeatedly carried out to meet plants’ P demand. Unfortunately, P fertilizers are mainly produced from a limited source derived from phosphate rocks, whose reserves are diminishing (Chowdhury et al. 2017).
The utilization of biochar (derived from biomass pyrolysis under limited oxygen) has gained increasing attention due to its positive agronomic and environmental impacts. Its soil incorporation can improve the soil’s pH, nutrient retention and supply, fertility, fertilizer use efficiency, and crop productivity, as well as physical and biological properties (Agegnehu et al. 2016; Ibrahim et al. 2020b; Lehmann et al. 2011; Song et al. 2018). Unlike for nitrogen and carbon, utilizing biochar to promote P cycling in soils, either alone or combined with organic, inorganic or biological sources of P has gained less attention, while its reported effects have been largely inconsistent (Deinert et al. 2024; Hu et al. 2023; Jin et al. 2023a, 2023b; Liu et al. 2023; Qayyum et al. 2021; Rafique et al. 2020). When P-rich biomasses are pyrolyzed, the derived biochar could serve as a P source for plants and microbial use (Sun et al. 2018; Zhang et al. 2016). Also, the phosphates adsorbed by biochar from soil solutions could be subsequently released to increase soil P availability (Gao et al. 2019; Penn and Camberato 2019; Streubel et al. 2012). Biochar could also increase P availability by promoting soil microbial activity and mycorrhizal colonization (Bornø et al. 2018a; Kahura et al. 2018; Sarfraz et al. 2020). However, there are reports of a reduction in P availability after biochar amendment in different soil types with varying pH (Bornø et al. 2018b; Chintala et al. 2014; Xu et al. 2016). Meta-analysis has indicated biochar’s effect on P availability varies among soil conditions and cannot be generalized (Glaser and Lehr 2019). Despite the promising roles of biochar in promoting plant productivity and sustainable agriculture, its inconsistent effect on P availability requires techniques targeted at improving its surface and chemical properties to optimize P retention and release from soils to meet plant needs.
The characteristic negative charges dominant on the surfaces of raw biochar (Tan et al. 2016) provide limited ability for phosphate (H2PO4−, HPO42−, and PO43−) retention and subsequent slow release. When raw biochar (with a high pH value) is applied in acidic soils, it increases the soil’s pH, resulting in the deprotonation of its surface phenolic and lactonic functional groups, thus increasing its negative charges (Xiong et al. 2021) and reducing its potential for phosphate retention. Some post-pyrolysis optimization of biochar could increase its adsorption capability relative to the non-optimized forms (Ibrahim et al. 2020a; Lu et al. 2023; Tan et al. 2016). Integrating magnesium (Mg) nanocomposites on biochars’ surface can improve its potential for adsorbing ions by integrating more functional groups, expanding its specific surface area, and altering its surface ions (Ibrahim et al. 2023b; Tao et al. 2019; Yang et al. 2019; Zheng et al. 2020). Particularly, modifying biochar with Mg-oxide (which has high phosphate affinity) can increase binding sites for phosphates (Li et al. 2017). While available utilizations of MgO-functionalized biochar nanocomposites are largely limited to P removal from wastewater, there remains a significant dearth of information on its soil application or field-scale impacts on growth-stage-related P availability and its regulatory mechanisms in the soil–plant system. Sepiolite (Mg4Si6O15(OH)2·6H2O), a Mg-rich silicate clay mineral is an important binder used in the production of fertilizers due to its effective adsorption of a wide range of ions (Frost et al. 1998). Sepiolite is effective for the retention and slow release of nitrogen (nitrate) (Mohammadi et al. 2020) and in remediating contaminated soils (Yin et al. 2017). As a biochar composite, sepiolite could facilitate its particles’ aggregation to biochar to increase the biochar’s binding capacity for the adsorption of ions, thus useful for nutrient retention and soil remediation (Ibrahim et al. 2023a; Zhan et al. 2019). The abundant Mg ions in sepiolite could facilitate the adsorption of phosphates and their timely release for plant use. However, it remains unknown if, and how sepiolite-biochar composites would regulate soil P availability on a field scale across different plant growth stages.
Furthermore, the soils’ properties could be modified after adding biochar, and the extent of this modification is dependent on the source of biochar, its method of production, and other soil and environmental conditions. Therefore, biochar could induce positive, adverse, or nonsignificant influence on soil microbial properties (Luo et al. 2017; Palansooriya et al. 2019; Tian et al. 2016). While these inconsistent effects of biochar have been documented for some major microbial-mediated nutrient cycling processes (nitrogen and carbon cycling), there are currently no available reports of how Mg-functionalized biochar regulates P availability by impacting bacterial 16S rRNA and functional genes regulating P cycling across different key maize growth stages under field conditions.
To this end, this study was designed to explore the basic chemical and biological mechanisms regulating P availability in the soil-maize system at varying maize growth stages in soils amended with functionalized MgO- and sepiolite-biochar composites. We hypothesized that relative to pristine biochar, the abundant Mg ions in MgO- and sepiolite-functionalized biochars would improve their chemical and surface characteristics for optimized phosphate retention and their release for plant use, with variations at different maize growth stages ii. The impact of the modified biochars on P availability would be connected to their effect on microbial functional genes regulating P availability.
2 Materials and methods
2.1 Production of biochar/optimized biochar
Bamboo shoot shavings were utilized as the feedstock for biochar production. Details of its production conditions are outlined in a sister paper (Ibrahim et al. 2022). Briefly, the feedstock was dried and pyrolyzed at a heating rate of 10 °C min−1 on an electricity-powered pyrolysis unit at 500 °C ± 10 °C and held for 2 h. To achieve nanoscale Mg incorporation into the pristine biochar, 5:1 (v/w) of 500 mL 1 M MgCl2⋅6H2O solution (pH = 7.5) was mixed with biochar and stirred for 1 h and kept for 24 h. The mixture was then drained and passed through a 0.45 µm membrane and oven-dried for 12 h at a constant temperature of 105 °C (Wu et al. 2019). The obtained engineered biochar was sieved using a 1 mm sieve and denoted as MgOB. Sepiolite modification of the pristine biochar was carried out by evenly mixing 2:1 (m/m) of the pristine biochar and factory-grade sepiolite. To obtain a uniform material, ultrapure water was added to the mixture and thoroughly mixed. (Shi et al. 2020). The obtained biochar-sepiolite composite was dried in an oven at 60 °C for 12 h and the cooled product was crushed, passed through a 1 mm sieve, and denoted as SB. Table S1 shows the characterization of the derived biochars.
2.1.1 Analyses of biochars’ properties
The derived biochar’s pH assay was done in a 1:10 (w:v) mixture of biochar to water (deionized). The samples were digested in H2O2 and H2SO4 at 120 °C to determine their total P and K contents (Enders and Lehmann 2012). The concentration of total P in the filtered digest was assayed by the indophenol blue technique, while total K concentration was assayed by flame photometry. The amount of Na+ displaced by NH4+ from biochar’s exchange sites was used to measure its CEC after leaching with 1 M ammonium acetate (Zhao et al. 2019). An elemental analyzer (TruMac, MI, USA) measured the total carbon (C), nitrogen (N), and hydrogen (H), while the oxygen (O) content was measured using:
To evaluate the deposition of Mg nanocomposites on the surface and in the matrix of the optimized biochar, X-ray diffractometry (Ultima IV, MA, USA) was carried out. Scanning electron microscopy (SEM) was conducted to visualize the surface characteristics of the biochar and its modified forms (NanoSEM; OR, USA). The biochars’ surface area was estimated by the Brunauer-Emmet-Teller (BET) gas autosorp system (Trister II 3020, Shanghai, China).
2.2 Experimental location and setup
The investigation was set up in an agricultural field situated in southern China (latitude 26.204 and longitude 119.078). The study site lies at 30 m above sea level. Figure S1 shows the average monthly rainfall days and precipitation amounts (Fig. S1a) and the average monthly high and low temperatures during the experimental year (Fig. S1b) (the experiment was conducted from April to July). The initial properties of the soils before land preparation are shown in Table S2.
The experiment was conducted on field plots with dimensions of 1.5 × 1.5 m per plot. One meter space was maintained between the plots across the experimental area. The plots were demarcated on a pre-harrowed and ploughed field to ensure the soils were lose enough for biochar incorporation. The biochar and its modified forms were incorporated and mixed with the soils within each plot up to 20 cm depth at a rate of 40 t ha−1 when the soil was still dry to ensure uniform mixing. Within three days of biochar amendment, the field was subjected to light rainfall and an early maturing maize (Zea mays L.) variety was planted three days after the rains. The treatments evaluated for this study included a control treatment (C), single super phosphate (SSP) fertilization (F), pristine biochar + P fertilization (BF), MgO-optimized biochar + P fertilization (MgOBF), and sepiolite-optimized biochar + P fertilization (SBF). The treatments were randomized with three replications per treatment across the experimental area. P fertilization was carried out with single super phosphate (SSP) at 75 kg P ha−1 using the side placement method 2 weeks after maize sowing. Aside from the control treatment, the plants were also fertilized uniformly with urea (for N) and potassium chloride (KCl) (for K) at 150 and 75 kg of N and K, respectively. Planting was done with two seeds per hole which were later thinned to 1, at a spacing of 25 and 75 cm for intra-row and inter-row (53,333 plant stands per hectare), respectively. The plants were grown until physiological maturity under natural rainfall for 3 months. All cultural practices were carried out to maintain healthy maize stands throughout the entire growth period.
2.3 Sampling of soil and plant and chemical analyses
To measure P availability in soil and its concentration in the plant across different growing stages, sampling was carried out at three key growth stages characterized by different nutrient uptake patterns: the vegetative stage (12-leaf stage), the reproductive (blister stage), and physiological maturity stage at 59, 75 and 95 days after sowing, respectively. Soil sampling was randomly done from several points around the plant root zone at 20 cm depth and bulked per plot across each treatment. Subsamples per plot were obtained and separated into two parts. One part was labeled and stored for measuring soil chemical properties, while the other subsample was transferred into an ice pack and subsequently subsequently stored at − 80 °C for microbial assay (at the blister and physiological maturity stages). Plant sampling was done by the destructive sampling of a plant stand located in the central section of the ridge during each sampling period. The whole plant was cut into small parts and dried for 12 h at 60 °C in labeled envelopes. The grains obtained after harvest were also oven-dried under similar conditions. They were subsequently weighed, ground, sieved (0.15 mm), and stored for total P analysis.
2.3.1 Soil and plant analyses
A soil:water ratio of 1:2.5 was mixed, stirred, and allowed to stand for 30 min before soil pH measurement. Sodium bicarbonate solution (pH 8.5) was used for the extraction of soil available P (Do Carmo Horta and Torrent, 2007), and was followed by the development of blue color using molybdate-ascorbic acid reagent before colorimetry at 880 nm using a microplate reader (BioTeck Epoch, Thermo Fisher Scientific). Soil and plant total P were determined as done for the biochar but with 1 g of soil (Enders and Lehmann 2012).
The FastDNA Spin extraction kit was used for DNA extraction (MP Biomedicals, CA, USA) as outlined by the manufacturers. The PCR amplification of the bacterial 16S rRNA hypervariable regions (V3-V4) was done with the primer pairs, 338F and 806R, and the PCR conditions were documented in an earlier study (Ibrahim et al. 2020b). The polymerized and purified paired-end equimolar amplicons (2 × 250 bp) were sequenced on an Illumina MiSeq platform (Allwegene Technologies, BJ, China). The raw sequence reads can be assessed using PRJNA781060 on the NCBI database (sequence read archive).
2.4 Statistical analyses and bioinformatics
We employed the analysis of variance (one-way) to explore the variations among soil and plant properties and visualized the output using OriginPro 2021. Comparisons among means were executed using the Tukey’s test (p < 0.05). The raw data from the X-ray diffraction analysis were processed using the JADE software (v6.5) and the patterns were generated and visualized using OriginPro 2021.
The raw sequence data obtained were processed using the QIIME2 pipeline to trim and eliminate low-quality sequences (quality threshold value < 20 and length < 250 bp). Uparse (v 7.0) was used to remove chimeric sequences, and the operational taxonomic units (OTUs) were clustered from the derived clean tags at 97% similarity. The OTUs classification was done via the Silva database (SILVA Release 138). The alpha diversity was denoted by the number of observed species in the soil samples collected during the blister and physiological maturity stages of maize. The beta diversity assessment was based on the unweighted unifrac distance (BrayCurtis) for each pair of OTUs. The number of key bacterial functional gene copies regulating P cycling during the blister and physiological growth stages of maize were predicted using PICRUSt2 against the KEGG (Kyoto Encyclopedia of Genes and Genomes) database. The algorithm of PICRUSt2 can precisely predict functional genes using a huge database of reference genomes (Douglas et al. 2020). The multivariate relationships between soil P cycling genes and the P in the soil and plant in the different treatments were explored using the Redundancy analysis (RDA) on XLSTAT software.
3 Results
3.1 Characteristics of biochar and its modified forms
Compared to the unmodified biochar, the pH of the MgO and sepiolite-modified biochars slightly increased, but with no significant (p = 0.07) difference between the modified forms (Table S1). The CEC and BET surface area of the Mg-O and sepiolite-biochars significantly increased compared to its pristine form. Also, elemental composition (P, nitrogen (N), and potassium (K)) was significantly highest in MgOB (Table S1). To confirm the incorporation of Mg-based nanoparticles on the surfaces and matrix of the optimized biochars, X-ray diffraction revealed the presence of Mg-associated compounds on the optimized biochars compared to the unmodified biochar (Fig. 1a and b). Also, SEM images revealed the presence of deposited nano-particles of sepiolite and Mg incorporated onto the surface and matrix of the optimized biochars (Fig. 1c–e).

Characterization of biochar and optimized biochar. a X-ray diffraction (XRD) of raw biochar, b XRD of optimized biochar, and scanning electron microscopy (SEM) images of c biochar, d MgO-optimized biochar and e sepiolite-optimized biochar. The SEM images show the formation of MgO and sepiolite nanoparticles on the surfaces of the optimized biochar. In b, MgOB indicates MgO-optimized biochar, and SB indicates sepiolite-optimized biochar. The red spheres and arrows in a and b indicate the occurrence of Mg-nanocomposites on the surface and pores of the modified biochars, respectively
3.2 Effect of biochar/optimized biochar on soil–plant P dynamics and uptake
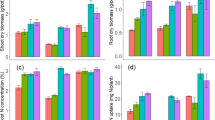
During the 12LS and blister stage of maize growth, the modified biochar treatments (MgOBF and SBF) significantly increased soil available P compared to the BF, F, and C treatments. At the 12-LS, available P in the MgOBF treatment increased by 49.1%, 35.5%, and 26.5% compared to the C, F, and BF treatments, respectively (Fig. 2a). Also, available P in SBF increased by 35.1%, 22.9%, and 14.7% in C, F, and BF, respectively. A similar pattern in available P content in the SBF and MgOBF treatments was obtained during the blister stage relative to the C, F, and BF treatments. However, during the PM stage, the SBF treatment maintained the significantly highest soil available P content (15.5% higher than the BF), while insignificant variation was recorded among the other treatments. Across the three growth stages of maize, AP was 15.6% and 13.2% higher in the MgOBF and SBF treatments, respectively, than in the BF. A notable increase in soil total P in treatments incorporated with biochar compared to the non-biochar amended (C and F treatments) across the three growth stages of maize evaluated was observed (Fig. 2b). Despite the high soil available P in the SBF treatment at the 12LS (Fig. 2a), there was no corresponding increase in P accumulation in the plant (Fig. 2c). The high soil available P in the MgOBF treatment at the 12LS and blister stage corresponds with the high plant P concentration (Fig. 2c) during these stages. Plant total P did not significantly vary across the treatments at the blister stage (Fig. 2c). The reduction in the soil available P in the MgOBF treatment during the PM stage (Fig. 2a) unlike during the earlier growth stages was associated with a significant reduction in plant total P concentration (Fig. 2c). This reduction was also associated with an increase in soil total P compared to the other treatments (Fig. 2b). Among the biochar-amended treatments (BF, MgOBF, and SBF), soil available P was highest in SBF at the PM stage (Fig. 2a). However, there was insignificant variation in plant total P concentration among the C, F, BF, and SBF treatments at this stage (PM), while the lowest value was obtained in MgOBF (Fig. 2c). Notably, the SBF had the highest grain total P, which was followed by F and MgOBF, while the C and F treatments had similar and significantly least grain P concentrations (Fig. 2d).

Soil and plant phosphorus (P) concentration at three growth stages of maize. Soil a available P concentration and b total P concentration, c plant biomass P concentration, d grain total P concentration, e plant biomass yield, and f plant P uptake. The error bars represent the standard error of the mean, and differences among the means at the same growth stage are indicated by the alphabets on violin plots and bars with similar colors using Tukey’s test (n = 3, p < 0.05). C: control; F: fertilizer (single super phosphate); BF: biochar + F; MgOBF: MgO-optimized biochar + F; SBF: sepiolite-optimized biochar + F. 12LS: 12-leaf stage, BS: blister stage, PM: physiological maturity
At the 12LS, plant biomass was least in the control treatment. Among the biochar-amended soils, the plant biomass was lower in the SBF and F treatments than in the BF and MgOBF (Fig. 2e). No significant variation was recorded between the BF and MgOBF treatments. The plant biomass was higher in BF than in the F and SBF treatments at the blister stage (Fig. 2e). The MgOBF maintained the highest biomass and grain yield across the treatments. Notably, P uptake was higher in the BF than in the SBF treatment at the 12LS and BS, while the highest P uptake among the treatments at these stages was recorded in MgOBF (Fig. 2f). However, the increase in P availability in SBF at the PM stage (Fig. 2a) was associated with a high P uptake in the biomass and grain compared to other treatments (Fig. 2f).
3.3 Alpha and beta diversity of 16S rRNA genes and dynamics of P functional genes
The alpha diversity of 16S rRNA genes, indicated by the number of observed species revealed insignificant variation in bacterial OTUs distribution across the treatments during the blister stage (Fig. 3a). However, relative to the other treatments, the diversity of the observed bacterial OTUs significantly increased in MgOBF during the PM stage of maize (Fig. 3b). Notably, the F treatment significantly reduced the number of observed species of bacteria in rhizosphere soil at the PM stage (Fig. 3b). Beta diversity analysis, represented by the unweighted unifrac distance heatmap, showed a more distinct soil bacterial OTU structure during the PM stage (Fig. 3d) relative to the blister stage (Fig. 3c). We observed a similar bacterial structure in the C and F treatments during the PM and blister stages. While the heatmap revealed that the MgOBF treatment possesses a distinct bacterial structure, a closer distance matrix of bacterial structure existed in the SBF and BF treatments. Additionally, Venn diagrams were constructed to explore the number of unique OTUs shared by the treatments at the different maize growth stages (Fig. 3e and f). The OTUs abundance increased in the pattern: C > F > BF > SBF > MgOBF at both growth stages. While the OTUs increased towards physiological maturity, the number of shared OTUs decreased. Besides, unlike their abundance, the analysis of similarity (ANOSIM) of bacterial OTUs across the blister and PM stages showed no significant difference in the identities of bacterial 16S rRNA across these growth stages in the evaluated treatments (Tables S3 and S4).

Diversity analysis of 16S rRNA genes. a The alpha diversity indicated by the number of observed bacteria species in rhizosphere soil at the blister stage and b physiological maturity stage of maize; c the beta diversity analysis showing the unweighted unifrac distance heat map among treatments at the blister stage and d physiological maturity stage of maize. e Venn diagrams showing the number of unique and shared OTUs in soil among treatments at the blister stage and f at the physiological maturity stage of maize. Each box plot in a and b represents the mean of each treatment at the indicated growth stage (n = 3). The error bars represent the standard error of the mean, and differences among the means are indicated by the lowercase letters using Tukey’s test (p < 0.05). C: control; F: fertilizer (single super phosphate); BF: biochar + F; MgOBF: MgO-optimized biochar + F; SBF: sepiolite-optimized biochar + F
The top four (4) most abundant P cycling genes copies selected from the pool of P cycling genes predicted across the blister and PM growth stages of maize exhibited different growth stage-related responses in their expression patterns across the treatments (Fig. 4). Just like the number of OTUs (Fig. 4e and f), the number of P cycling genes reduced at the PM stage compared to the blister stage. During the blister stage, the P starvation genes (phoR, phoU, and phoP) did not show significant response to fertilizer (F) or optimized biochar/biochar amended soils, while phoB was most expressed in the F treatment (Fig. 4a). However, the P starvation genes phoU and phoP were significantly reduced equally in the SBF and MgOBF relative to C, BF, and F treatments, while phoB was most expressed in C (Fig. 4b). We observed no significant variation in the number of gene copies (pstB, pstC, pstA, and phnD) regulating P uptake and transport at the blister stage (Fig. 4c). However, pstA, pstB, and pstC were significantly reduced similarly in the MgOBF and SBF relative to C, F, and BF treatments (lower, but statistically similar in BF) (Fig. 4d). Similar to the blister stage, no significant treatment effect was observed on the expression of phnD (Fig. 4d). The gene copies regulating P solubilization and mineralization (phoD, phnJ, phnM, and phnP) were significantly reduced (p < 0.043) in the control treatment at the blister stage (Fig. 4e) compared to those in other treatments. However, there was a significant stimulation of gene copies regulating P solubilization (phoD, phnJ, phnM, and phnP) and mineralization during the PM stage similarly in MgOBF and in SBF (Fig. 4f). Particularly, the phoD gene which regulates alkaline phosphatase and plays a key role in P solubilization was significantly stimulated in the SBF treatment at the PM growth stage of maize.

The abundance of soil phosphorus (P) metabolizing gene copies as affected by different soil amendments. Soil abundance of a, b P starvation genes, c, d P uptake and transport genes, and e, f P solubilization and mineralization genes at the blister stage and physiological maturity stages, respectively. The error bars represent the standard error of the mean, and differences among the means of each gene represented by bars with similar colors are indicated by the alphabets on bars using Tukey’s test (n = 3, p < 0.05). C: control; F: fertilizer (single super phosphate); BF: biochar + F; MgOBF: MgO-optimized biochar + F; SBF: sepiolite-optimized biochar + F. P starvation response genes (phoR, phoU, phoB, phoP); P uptake and transport genes (pstA, pstB, pstC, phnD), P solubilizing and mineralizing genes (phoD, phnJ, phnM, phnP)
3.4 Relationship between P functional genes and soil properties
To establish the relationships among the treatments, soil P-cycling gene copies, and soil–plant P content, a multivariate RDA was conducted from datasets obtained at the blister and PM stages of maize (Fig. 5). During the blister stage, we observed a positive relationship between the key P solubilization gene (phoD) and soil available P, total P, and plant P, particularly in MgOBF and SBF (Fig. 5a). The C and F treatments had a positive relationship with the P uptake and transport genes (pstA, pstB, and pstC) and the P starvation response gene (phoP and phoU). At the physiological maturity stage, positive associations between soil total P, and P solubilization and mineralization gene copies (phoD, phnJ, phnM, and phnP) in the MgOBF were recorded. The soil available P and grain total P showed a positive association in the SBF treatment (Fig. 5b). The control, F, and BF treatments showed positive associations with the P uptake and transport gene copies and the P starvation gene (Fig. 5b). Plant total P did not show any association with the P cycling genes at physiological maturity.

Redundancy analysis showing the relationship between treatments, phosphorus (P) cycling genes and the P content of soil and plant at a blister stage b physiological maturity stage of maize. TP: total P; C: control; F: fertilizer (single super phosphate); BF: biochar + F; MgOBF: MgO-optimized biochar + F; SBF: sepiolite-optimized biochar + F; P starvation response genes (phoR, phoU, phoB, phoP); P uptake and transport genes (pstA, pstB, pstC, phnD), P solubilizing and mineralizing genes (phoD, phnJ, phnM, phnP)
4 Discussion
4.1 Biochar’s modification effect on soil–plant P dynamics
The effect of biochar on soil pH plays a vital role in its impact on soil P availability (Chan et al. 2007). Soil pH is adjudged as one of the most crucial regulators of soil P availability (Zwetsloot et al. 2016). Raw biochar did not significantly influence soil P availability or plant P concentration compared to inorganic fertilization, despite inducing a considerable increase in soil pH (ranging from 6.16 to 6.42 in BF, compared to 5.19 to 5.38 in the F treatment). The deprotonation of the phenolic and lactonic functional groups of the raw biochar with increasing soil pH may have reduced the P retention capacity of the biochar-applied soil and increased its loss relative to uptake, resulting in limited P availability for plant use. When biochar is applied to acidic soils, its functional groups undergo deprotonation, involving the loss of a proton (H⁺) from the functional groups (Xiong et al. 2021), and thus, they become negatively charged and exhibit poor adsorption of negatively charged phosphates (Lalley et al. 2016). Also, the insignificant difference in P availability under BF compared to F could partly be associated with the entrapment of the added inorganic P fertilizer in the pores and surface of biochar, and its co-precipitation with ash in the biochar (Yao et al. 2013). Such entrapment of inorganic P by biochar resulting in the reduction in soil P availability has been reported (Chintala et al. 2014; Xu et al. 2016). However, some reports have documented the positive effects of biochar on soil P availability (Bornø et al. 2018a; Glaser and Lehr 2019; Kahura et al. 2018; Liu et al. 2019; Sarfraz et al. 2020). The diverse effects of biochar amendments on soil P availability primarily depend on biochar properties and quantity applied, soil types, and management regimes (DeLuca et al. 2015; Zhu et al. 2018).
Modifying biochar to optimize its surface and sorption characteristics is essential to enhance its potential benefits for environmental management (Tan et al. 2016). Information from the XRD peaks of the modified biochar (Fig. 1a and b) showed that MgO and sepiolite-based modifications were effective in incorporating Mg-bound compounds on the biochar’s surfaces. These compounds could react with soil phosphates to form magnesium phosphates (such as Mg2PO4OH) which can be subsequently desorbed for plant use (Chen et al. 2018). Also, the presence of the nanoparticles of Mg and sepiolite on the biochar’s surface and matrix (Fig. 1d and e) compared to the pristine biochar (Fig. 1c) would serve as sorption sites for negatively charged phosphate ions and their subsequent release for plant uptake (Wu et al. 2019). Thus, there is an associated increase in available P in the functionalized biochars-treated soils, particularly, in MgOBF at the 12LS and the blister stage. However, the increase in available P in the SBF treatment at the PM stage translated to higher total plant and grain P compared to MgOBF, indicating its potential for extended P-release.
As soil pH increases after the amendment with optimized biochars (having high pH values) (Fig. S2), the deprotonation of biochar functional groups intensifies and there is increasing electrostatic repulsion between the phosphate ions and the surface of biochar, which limits phosphate adsorption and increases its availability (Lalley et al. 2016; Shi et al. 2023). However, introducing Mg2+ on the surface and matrix of the optimized biochars ensured the retention of the repelled phosphates as Mg-bound phosphates. The extent of the release of the P bound to Mg tended to increase during the PM stage of maize where the soil pH significantly increased, particularly in the SBF treatment. Also, the increase in available P during the PM stage in the SBF treatment could be due to the high pH recorded compared to other treatments (Fig. S2). The higher pH in SBF would induce more deprotonation of biochar functional group and increase the rate of P repulsion, thus increasing soil available P as observed in the SBF treatment. Thus, the rise in the pH of soils amended with modified biochar further promoted P availability. Notably, the higher P content of the MgO-optimized biochar (MgOB) compared to the raw and sepiolite-biochars may have also played a vital role in its P supply capacity. This is because the constituent P in biochar can serve as an additional P source for plant and microbial use when released into the soil (Sun et al. 2018; Zhang et al. 2016). Besides, the significantly higher CEC and specific surface area of MgOB than the raw or sepiolite-biochar provided a higher reactive surface area for retaining P on the biochar’s exchange complex. The adsorption of phosphates to Mg-functionalized biochar is a heterogeneous surface adsorption process and is highly influenced by the biochar’s surface area (Cui et al. 2016). The improved surface area and CEC of biochar after modification (as observed in MgOB) are vital for increased retention of P ions either by complexation, electrostatic or capillary forces, or a combination of these forces (Dey et al. 2023), and its subsequent release for plant use. Higher CEC of the modified biochar provides it with a higher capacity to retain more Mg2+, whose presence accelerates phosphate retention. In agreement with our observations, Mg-based modifications of biochar have been shown to increase its surface area, thus promoting its sorption properties (Ibrahim et al. 2023b; Tao et al. 2019; Yin et al. 2018; Yang et al. 2019; Zheng et al. 2020). Compared to soil mineral-bound P, P sorbed on biochar surfaces is more readily released in available forms (Gao et al. 2019). Hence, the retained P on the modified biochar surface will be slowly released over time to meet plants’ needs. However, we noted that the extent of P retention and release varied across maize growth stages and modification types.
Additionally, P retention was higher over time in MgOBF-amended soil as verified by the higher total soil P compared to the other treatments. In addition to its high P content, the formation of Mg-bound phosphates on the surface and matrix of the MgO-doped biochar is a key mechanism for its higher P retention and increase in soil total P content. The high concentration of Mg in MgO-modified biochar is essential for accelerated inorganic P adsorption on the modified biochar (Li et al. 2017). This observation is consistent with previous enhanced retention of inorganic P by MgO-optimized biochar compared to unmodified biochar in sorption experiments (Li et al. 2017; Wu et al. 2019). The formation of Mg3(PO4)2, Mg2PO4OH, and MgHPO4 crystalline phases with the Mg (Chen et al. 2018) by MgO and sepiolite incorporated into biochar was the primary pathway of P retention, and regulating subsequent P release and the increase in soil total P. It is noted that the P supply potential of the MgOBF treatment was greater at the 12LS and blister stage of maize, while SBF treatment provided more available P towards grain filling during maize physiological maturity. This was also associated with higher plant biomass P and P uptake in the PM stage in the SBF than in the MgOBF treatment (Fig. 2c). During the initial stages of Mg-phosphate interaction, the formation of Mg-phosphate complexes on the biochar’s surface is via weak chemical bonds (Yao et al. 2011), and thus P could be released more readily during the 12LS in MgOBF than the later stage (as shown in Fig. 2a). This did not apply to the Mg in sepiolite as P release was delayed in the SBF treatment to the later stage of maize growth, thus significantly increasing biomass and grain yield as well as grain P content at the PM stage in this treatment. The delayed P availability in SBF could in part be due to the high adsorption properties of sepiolite (Frost et al. 1998), associated with the dual binding of phosphate on the mineral lattice, and with Mg ions in the modified surface of the biochar. Thus, compared to pristine biochar, the higher capacity of MgO-functionalized biochar for P retention and its slow release will be useful for meeting plant needs over time.
4.2 Optimized biochars enhance microbial P solubilization and mineralization
The shifts in functional genes regulating P supply are vital for understanding the microbial mediation of P cycling and provide deeper insights into how biochar’s modification regulates P retention and supply, and the mechanisms involved at different plant growth stages. The increase in bacterial 16S rRNA genes in the optimized biochar treatments across maize growth stages, especially at the PM stage in the MgOBF treatment is linked to the higher nutrient composition of the MgO-modified biochar, its increased surface area and CEC (Table S1), and thus, its capacity to retain more nutrients and promoting substrates availability for more microbial colonization. The high nutrient requirement of maize during its reproductive stage could shape soil microbial distribution in its rhizosphere (Li et al. 2014). Hence, the high nutrient concentrations in the modified biochars, particularly the MgO-optimized biochar were also key in promoting the 16S rRNA gene abundance in the MgOBF treatments. This assumption supports the distinct bacterial diversity in the modified biochar soils (Fig. 3a–d), particularly at the PM stage where the MgOBF possessed a higher bacterial diversity (Fig. 3b) and a unique bacterial OTU clustering compared to the other treatments across both growth stages (Fig. 3c and d).
Microorganisms are the key drivers of soil P cycling due to their ability to mineralize organic P and immobilize inorganic P (Kehler et al. 2021; Willey et al. 2014). The cycling of P in the soil is primarily regulated by three groups of P cycling genes: those regulating P starvation response, uptake and transport, and mineralization and solubilization (Dai et al. 2020). We found that, unlike at the blister stage where P starvation genes did not significantly vary among treatments, the reduction in their expression (particularly, phoU and phoP) in the MgOBF as well as in the SBF treatments during the PM stage suggests that the increase in the release of retained P (as earlier observed) in these treatments provided available P for microbial and plant use. Hence, there was a reduction in the expression of the P starvation response genes in the modified biochar-amended soils compared to the raw biochar, inorganic fertilizer, or control treatments where P availability was lower. When there is a limitation in available P supply, the increase in the expression of P-starvation genes is directly linked with the enhancement of P availability, which could concurrently stimulate P uptake and transport genes, as well as indirectly connected with its mineralization and solubilization (Eder et al. 1996; Helfenstein et al. 2018). Thus, P uptake in the C, F, and BF treatments where less P was available during the PM stage was mainly mediated and promoted by the stimulation of P starvation genes to promote P availability.
The reduced expression of gene copies regulating the uptake and transport of P (pst A, B, and C) in the SBF and MgOBF treatments, particularly during the PM stage, suggests that microbial P uptake was reduced in favor of plant uptake under the optimized biochar-treated soils compared to in the BF, F, and C treatments (Fig. 4d). Thus, the stimulation of P uptake and transport gene copies in the C, F, and BF treatments alike, limited P availability in these treatments due to higher microbial immobilization. Microbial functional genes regulating the uptake and transport of P are primarily involved in efficient P utilization and immobilization into microbial biomass (Richardson and Simpson 2011). When their expression is high, it could result in a reduction in P availability. Therefore, stimulation of these genes in C, F, and BF, unlike in the MgOBF and SBF treatments was associated with less P availability, resulting in the stimulation of the P starvation response genes in these treatments as shown in Fig. 4b. This observation was further supported by the positive associations between the P uptake and transport gene copies in the control, BF, and F treatments during the maize PM growth stage (Fig. 5b). While P availability has been shown to positively correlate with the affinity of transporter proteins, such as those contained in genes regulating the uptake and transport of P (Dai et al. 2020), the persistence of this relationship could ultimately result in the reduction in P availability due to increased P immobilization as recorded in our study.
The significant stimulation of gene copies regulating P solubilization and mineralization (phoD, phnJ, phnM, and phnP) during maize physiological maturity in the MgOBF and SBF treatments shows that these genes were essential in promoting the availability of P that is held in the soil, as well as the constituent biochar P that is held on its modified surfaces. Evidence has shown that microbial genes regulating P solubilization and mineralization can dissolve inorganic P or express enzymes that could promote the mineralization of organic P to increase P availability (Elias et al. 2001). The expression of PhoD, which was significantly highest in the SBF treatment during the PM stage where P availability was highest indicates its dominant role in the solubilization and mineralization of P in this treatment, and supports the conclusion that its expression positively correlates with soil-available P components (Wan et al. 2021). Such important roles of phosphate-solubilizing microorganisms in ensuring P availability through slow release when coupled with biochar have also been observed (Liu et al. 2023). The increase in soil pH in the soils amended with the optimized biochar, particularly during the PM growth stage (Fig. S2) was important in inducing P availability through increased microbial metabolism under more favorable soil pH conditions. Similar findings have shown that the pH of the soil is a primary factor that induces the increased expression of gene copies regulating organic P-mineralization (Wan et al. 2021). Thus, the key strategy for P availability and utilization by plants under the Mg-doped biochars is their ability to stimulate the expression of microbial functional genes regulating P solubilization and mineralization.
Our RDA revealed two distinct patterns in P availability under the MgOBF and SBF treatments. In support of higher P availability recorded up to the blister stage under MgOBF, the positive relationship between the dominant P solubilization and mineralization gene (phoD), soil AP, and total P in the MgOBF treatment indicated that P solubilization and mineralization was the main driver of P availability during this growth stage. However, at the PM stage, the positive correlation, especially between the MgOBF treatment, total P, and P solubilization and mineralization gene copies (phoD, phnJ, phnM, and phnP) (Fig. 5b) indicated continuous mineralization of organic P sources, while simultaneously promoting its solubilization. However, our RDA revealed that the higher P availability in the SBF treatment at the PM stage was partially microbially driven, as it could have resulted more from the desorption of the inorganic P held on the sepiolite-biochar composite and in the soil. Sepiolite is effective in the adsorption and slow release of ions (Frost et al. 1998; Mohammadi et al. 2020; Yin et al. 2017) and could ensure P availability over a longer period. Thus, the P supply strategies of different biochar optimization could vary, and would require adequate knowledge of the P requirement times of plants to ensure effective P utilization. However, a key mechanism for increased P availability under MgO-optimization of biochar is its potential to optimize the retention and release of P while simultaneously promoting microbial solubilization and mineralization of P. Sepiolite-optimization increased soil P availability mostly via increasing soil pH resulting in the potential deprotonation of biochar functional groups and the release of P, particularly, at the PM growth stage of maize.
4.3 Applicability of Mg-doped biochars across soil-crop systems and future research directions
Currently, most applications of Mg-doped biochars have focused on the removal of contaminants in soil and wastewater (e.g., Zhang et al. 2012; Li et al. 2017; Zhou et al. 2022). Soil application of these optimized biochars for crop production will largely depend on the soil type and its properties, as well as the crop species cultivated. Although the effect of biochar on soil pH largely depends on the source of its feedstock (Zhang et al. 2021), our observed increase in soil pH towards the physiological maturity of maize after the addition of the Mg-doped biochars indicates their suitability for acidic soils. Also, MgO is hygroscopic (Hornak 2021), as such, its doping on biochar which possesses water retention capacity in its pores will further increase biochar-mediated soil water retention. Thus, these biochars will be more suitable for soils with poor water retention such as those with high sand proportion, as well as soils in drought-prone regions. However, it is important to note that the specific effects of Mg-doped biochars on different soil types can vary depending on their dosage, application method, soil texture/structure, and plant species involved. Generally, Mg-doped biochars could be beneficial for crop production due to their capacity to optimize nutrient retention/release while increasing soil CEC. In addition, the Mg provided by these optimized biochars could also mitigate the widespread deficiency of Mg, especially, in acidic soils (Ishfaq et al. 2022). Mg also plays a fundamental role in the metabolism (a constituent of chlorophyll vital for photosynthesis and enzyme activation), growth, and development of plants (Tian et al. 2021). However, the possibility of introducing significant amounts of Mg into the soil by Mg-doped biochar makes it inapplicable for growing crops that are hyperaccumulators of Mg. For example, Spinach (Spinacia oleracea) and Soybean (Glycine max) are hyperaccumulators of Mg which can interfere with the uptake and utilization of other essential nutrients, such as calcium and potassium, leading to nutrient imbalances and potential physiological disorders (Marschner 2012).
Generally, our findings provide experimental and scientific proof that P limitation in cropland soils could be alleviated using Mg-doped biochars compared to raw biochars. Hence, soil application of these optimized biochars would be vital for promoting soil P use efficiency, increasing plant yield and biomass, and ultimately resulting in higher carbon capture by plants. Besides, these optimized biochars also show great promise in stimulating soil microbial diversity and structure and could be vital for promoting ecosystem sustainability. However, there is a need to further evaluate the effect of the high Mg concentration introduced into the soil by these modified biochars and to establish a threshold for Mg modification to avoid its toxicity effects on plants' metabolic processes and soil biological and chemical processes across soil types and climatic conditions. Despite these concerns, the use of Mg-doped biochar in soil presents a potential strategy to alleviate P limitation, particularly in acidic soils. Therefore, future research should focus on optimizing the amount of Mg used for doping biochars to achieve high performance while protecting the environment from excessive Mg accumulation. At the same time, field-scale evaluations of other appropriate optimized biochars aimed at relieving P limitation in highly acidic and alkaline soils should be explored. Also, the residual effect of the Mg-modified biochars and their P supply capacity during biochar aging needs to be evaluated to provide their long-term effects on P cycling and supply.
5 Conclusion
Amending maize-grown field soils with MgO and sepiolite biochar-based composites promoted soil phosphorus (P) availability through the retention and slow release of inorganic P to meet plant needs across the growth stages of maize compared to pristine biochar or inorganic P fertilization. Even though amending the soil with the modified biochars increased microbial 16S rRNA abundance, particularly during the physiological maturity stage of maize, they suppressed the expression of gene copies regulating P starvation response and P uptake and transport. However, their capacity to stimulate the genes regulating P solubilization and mineralization was a key strategy that ensured P availability during the reproductive stage of maize. At the physiological maturity stage of maize, sepiolite-optimization promoted the release of inorganic P held on biochar and in the soil relative to microbial-mediated P release as observed in MgO-optimized biochar treatment. Thus, the modified biochars increased P availability and plant uptake via the combined processes of slow-release of surface-retained Mg-bound phosphates while also inducing microbial P solubilization and mineralization. Our study provides a mechanistic understanding of soil P supply strategies under MgO and sepiolite-optimized biochars and provides a scientific basis for their potential field-scale utilization for crop production.
Data availability
The datasets used or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Agegnehu G, Nelson PN, Bird MI (2016) Crop yield, plant nutrient uptake and soil physicochemical properties under organic soil amendments and nitrogen fertilization on Nitisols. Soil till Res 160:1–13. https://doi.org/10.1016/j.still.2016.02.003
Andersson H, Bergström L, Djodjic F, Ulén B, Kirchmann H (2013) Topsoil and subsoil properties influence phosphorus leaching from four agricultural soils. J Environ Qual 42:455–463. https://doi.org/10.2134/jeq2012.0224
Arif M, Ilyas M, Riaz M, Ali K, Shah K, Haq IU, Fahad S (2017) Biochar improves phosphorus use efficiency of organic-inorganic fertilizers, maize-wheat productivity and soil quality in a low fertility alkaline soil. Field Crops Res 214:25–37. https://doi.org/10.1016/j.fcr.2017.08.018
Bornø ML, Eduah JO, Müller-Stöver DS, Liu F (2018a) Effect of different biochars on phosphorus (P) dynamics in the rhizosphere of Zea mays L. (maize). Plant Soil 431:257–272. https://doi.org/10.1007/s11104-018-3762-y
Bornø ML, Müller-Stöver DS, Liu F (2018b) Contrasting effects of biochar on phosphorus dynamics and bioavailability in different soil types. Sci Total Environ 627:963–974. https://doi.org/10.1016/j.scitotenv.2018.01.283
Chan KY, Van Zwieten L, Meszaros I, Downie A, Joseph S (2007) Agronomic values of greenwaste biochar as a soil amendment. Soil Res 45:629–634. https://doi.org/10.1071/sr07109
Chen Q, Qin J, Cheng Z, Huang L, Sun P, Chen L, Shen G (2018) Synthesis of a stable magnesium-impregnated biochar and its reduction of phosphorus leaching from soil. Chemosphere 199:402–408. https://doi.org/10.1016/j.chemosphere.2018.02.058
Chintala R, Schumacher TE, McDonald LM, Clay DE, Malo DD, Papiernik SK, Clay SA, Julson JL (2014) Phosphorus sorption and availability from biochars and soil/biochar mixtures. Clean–soil Air, Water 42:626–634. https://doi.org/10.1002/clen.201300089
Chowdhury RB, Moore GA, Weatherley AJ, Arora M (2017) Key sustainability challenges for the global phosphorus resource, their implications for global food security, and options for mitigation. J Clean Prod 140:945–963. https://doi.org/10.1016/j.jclepro.2016.07.012
Cui X, Dai X, Khan KY, Li T, Yang X, He Z (2016) Removal of phosphate from aqueous solution using magnesium-alginate/chitosan modified biochar microspheres derived from Thalia dealbata. Biores Technol 218:1123–1132. https://doi.org/10.1016/j.biortech.2016.07.072
Dai Z, Liu G, Chen H, Chen C, Wang J, Ai S, Wei D, Li D, Ma B, Tang C (2020) Long-term nutrient inputs shift soil microbial functional profiles of phosphorus cycling in diverse agroecosystems. The ISME J 14:757–770. https://doi.org/10.1038/s41396-019-0567-9
Deinert L, Hossen S, Ikoyi I, Kwapinksi W, Noll M, Schmalenberger A (2024) Poultry litter biochar soil amendment affects microbial community structures, promotes phosphorus cycling and growth of barley (Hordeum vulgare). Europ J Soil Biol 120:103591. https://doi.org/10.1016/j.ejsobi.2023.103591
DeLuca TH, Gundale MJ, MacKenzie MD, Jones DL (2015) Biochar effects on soil nutrient transformations. Biochar Environ Manag Sci Technol Implement 2:421–454. https://doi.org/10.4324/9780203762264
Dey S, Purakayastha TJ, Sarkar B, Rinklebe J, Kumar S, Chakraborty R, Datta A, Lal K, Shivay YS (2023) Enhancing cation and anion exchange capacity of rice straw biochar by chemical modification for increased plant nutrient retention. Sci Total Environ 886:163681. https://doi.org/10.1016/j.scitotenv.2023.163681
Do Carmo Horta M, Torrent J (2007) Phosphorus desorption kinetics in relation to phosphorus forms and sorption properties of Portuguese acid soils. Soil Sci 172(8):631–638. https://doi.org/10.1097/ss.0b013e3180577270
Douglas GM, Maffei VJ, Zaneveld JR, Yurgel SN, Brown JR, Taylor CM, Huttenhower C, Langille MGI (2020) PICRUSt2 for prediction of metagenome functions. Nat Biotech 38:685–688. https://doi.org/10.1038/s41587-020-0548-6
Eder S, Shi L, Jensen K, Yamane K, Hulett FM (1996) A Bacillus subtilis secreted phosphodiesterase/alkaline phosphatase is the product of a Pho regulon gene, phoD. Microbiology 142:2041–2047. https://doi.org/10.1099/13500872-142-8-2041
Elias MD, Tanaka M, Sakai M, Toyama H, Matsushita K, Adachi O, Yamada M (2001) C-terminal periplasmic domain of Escherichia coli quinoprotein glucose dehydrogenase transfers electrons to ubiquinone. J Biol Chemist 276:48356–48361. https://doi.org/10.1074/jbc.M107355200
Enders A, Lehmann J (2012) Comparison of wet-digestion and dry-ashing methods for total elemental analysis of biochar. Commun Soil Sci Plant Anal 43:1042–1052. https://doi.org/10.1080/00103624.2012.656167
Frost RL, Cash GA, Kloprogge JT (1998) Rocky Mountain leather’, sepiolite and attapulgite—an infrared emission spectroscopic study. Vibrat Spectr 16:173–184. https://doi.org/10.1016/s0924-2031(98)00014-9
Gao S, DeLuca TH, Cleveland CC (2019) Biochar additions alter phosphorus and nitrogen availability in agricultural ecosystems: a meta-analysis. Sci Total Environ 654:463–472. https://doi.org/10.1016/j.scitotenv.2018.11.124
Glaser B, Lehr V-I (2019) Biochar effects on phosphorus availability in agricultural soils: a meta-analysis. Scien Rep 9:9338. https://doi.org/10.1038/s41598-019-45693-z
Helfenstein J, Tamburini F, von Sperber C, Massey MS, Pistocchi C, Chadwick OA, Vitousek PM, Kretzschmar R, Frossard E (2018) Combining spectroscopic and isotopic techniques gives a dynamic view of phosphorus cycling in soil. Nat Comm 9:3226. https://doi.org/10.1038/s41467-018-05731-2
Hornak J (2021) Synthesis, properties, and selected technical applications of magnesium oxide nanoparticles: a review. Int J Mol Sci 22(23):12752. https://doi.org/10.3390/ijms222312752
Hou E, Luo Y, Kuang Y, Chen C, Lu X, Jiang L, Luo X, Wen D (2020) Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems. Nat Comm 11:637. https://doi.org/10.1038/s41467-020-14492-w
Hou E, Wen D, Jiang L, Luo X, Kuang Y, Lu X, Chen C, Allen KT, He X, Huang X (2021) Latitudinal patterns of terrestrial phosphorus limitation over the globe. Ecol Lett 24:1420–1431. https://doi.org/10.1111/ele.13761
Hu W, Zhang Y, Xiangmin R, Fei J, Peng J, Luo G (2023) Coupling amendment of biochar and organic fertilizers increases maize yield and phosphorus uptake by regulating soil phosphatase activity and phosphorus-acquiring microbiota. Agric, Ecosys Environ 355:108582. https://doi.org/10.1016/j.agee.2023.108582
Ibrahim MM, Hu K, Tong C, Xing S, Zou S, Mao Y (2020a) De-ashed biochar enhances nitrogen retention in manured soil and changes soil microbial dynamics. Geoderma. https://doi.org/10.1016/j.geoderma.2020.114589
Ibrahim MM, Tong C, Hu K, Zhou B, Xing S, Mao Y (2020b) Biochar-fertilizer interaction modifies N-sorption, enzyme activities and microbial functional abundance regulating nitrogen retention in rhizosphere soil. Sci Total Environ 739:140065. https://doi.org/10.1016/j.scitotenv.2020.140065
Ibrahim MM, Guo L, Wu F, Liu D, Zhang H, Zou S, Xing S, Mao Y (2022) Field-applied biochar-based MgO and sepiolite composites possess CO2 capture potential and alter organic C mineralization and C-cycling bacterial structure in fertilized soils. Sci Total Environ 813:152495. https://doi.org/10.1016/j.scitotenv.2021.152495
Ibrahim MM, Liu D, Wu F, Chen Y, He Z, Zhang W, Xing S, Mao Y (2023a) Nitrogen retention potentials of magnesium oxide-and sepiolite-modified biochars and their impacts on bacterial distribution under nitrogen fertilization. Sci Total Environ 866:161358. https://doi.org/10.1016/j.scitotenv.2022.161358
Ibrahim MM, Wu F, Chen Y, Liu D, Zhang W, He Z, Hou E, Xing S, Mao Y (2023b) Impacts of MgO-and sepiolite-biochar composites on N-partitioning and dynamics of N-cycling bacteria in a soil-maize system: a field-based 15N-urea tracer study. Geoderma 429:116236. https://doi.org/10.1016/j.geoderma.2022.116236
Ishfaq M, Wang Y, Yan M, Wang Z, Wu L, Li C, Li X (2022) Physiological essence of magnesium in plants and its widespread deficiency in the farming system of China. Front Plant Sci 13:802274. https://doi.org/10.3389/fpls.2022.802274
Jin J, Fang Y, He S, Liu Y, Liu C, Li F, Khan S, Eltohamy KM, Liu B, Liang X (2023a) Improved phosphorus availability and reduced degree of phosphorus saturation by biochar-blended organic fertilizer addition to agricultural field soils. Chemosphere 317:137809. https://doi.org/10.1016/j.chemosphere.2023.137809
Jin J, Khan S, Eltohamy KM, He S, Liu C, Li F, Liang X (2023b) Biochar–coupled organic fertilizer reduced soil water-dispersible colloidal phosphorus contents in agricultural fields. Chemosphere 333:138963. https://doi.org/10.1016/j.chemosphere.2023.138963
Kahura MW, Min H-G, Kim M-S, Kim J-G (2018) Assessing phosphorus availability in a high pH, biochar amended soil under inorganic and organic fertilization. Ecol Resil Infrastr 5:11–18. https://doi.org/10.17820/ERI.2018.5.1.011
Kehler A, Haygarth P, Tamburini F, Blackwell M (2021) Cycling of reduced phosphorus compounds in soil and potential impacts of climate change. European J Soil Sci 72:2517–2537. https://doi.org/10.1111/ejss.13121
Khan MI, Afzal MJ, Bashir S, Naveed M, Anum S, Cheema SA, Wakeel A, Sanaullah M, Ali MH, Chen Z (2021) Improving nutrient uptake, growth, yield and protein content in chickpea by the co-addition of phosphorus fertilizers, organic manures, and Bacillus sp. MN-54. Agronomy 11:436. https://doi.org/10.3390/agronomy11030436
Lalley J, Han C, Li X, Dionysiou DD, Nadagouda MN (2016) Phosphate adsorption using modified iron oxide-based sorbents in lake water: kinetics, equilibrium, and column tests. Chem Eng J 284:1386–1396. https://doi.org/10.1016/j.cej.2015.08.114
Lehmann J, Rillig MC, Thies J, Masiello CA, Hockaday WC, Crowley D (2011) Biochar effects on soil biota–a review. Soil Biol Biochem 43:1812–1836. https://doi.org/10.1016/j.soilbio.2011.04.022
Li X, Rui J, Mao Y, Yannarell A, Mackie R (2014) Dynamics of the bacterial community structure in the rhizosphere of a maize cultivar. Soil Biol Biochem 68:392–401. https://doi.org/10.1016/j.soilbio.2013.10.017
Li R, Wang JJ, Zhou B, Zhang Z, Liu S, Lei S, Xiao R (2017) Simultaneous capture removal of phosphate, ammonium and organic substances by MgO impregnated biochar and its potential use in swine wastewater treatment. J Clean Prod 147:96–107. https://doi.org/10.1016/j.jclepro.2017.01.069
Liu J, Sui P, Cade-Menun BJ, Hu Y, Yang J, Huang S, Ma Y (2019) Molecular-level understanding of phosphorus transformation with long-term phosphorus addition and depletion in an alkaline soil. Geoderma 353:116–124. https://doi.org/10.1016/J.GEODERMA.2019.06.024
Liu Z, Wu Z, Tian F, Liu X, Li T, He Y, Li B, Zhang Z, Yu B (2023) Phosphate-solubilizing microorganisms regulate the release and transformation of phosphorus in biochar-based slow-release fertilizer. Sci Total Environ 869:161622. https://doi.org/10.1016/j.scitotenv.2023.161622
Lu J, Li Y, Cai Y, Jiang P, Yu B (2023) Co-incorporation of hydrotalcite and starch into biochar-based fertilizers for the synthesis of slow-release fertilizers with improved water retention. Biochar 5:44. https://doi.org/10.1007/s42773-023-00242-w
Luo S, Wang S, Tian L, Li S, Li X, Shen Y, Tian C (2017) Long-term biochar application influences soil microbial community and its potential roles in semiarid farmland. Appl Soil Ecol 117:10–15. https://doi.org/10.1016/j.apsoil.2017.04.024
Marschner P (2012) Marschner's mineral nutrition of higher plants (3rd ed.). Academic Press. https://doi.org/10.1016/C2009-0-63043-9
Mohammadi N, Shariatmadari H, Khademi H, Bazarganipour M (2020) Coating of sepiolite-chitosan nanocomposites onto urea increases nitrogen availability and its use efficiency in maize. Arch Agron Soil Sci 66:884–896. https://doi.org/10.1080/03650340.2019.1643842
Palansooriya KN, Wong JTF, Hashimoto Y, Huang L, Rinklebe J, Chang SX, Bolan N, Wang H, Ok YS (2019) Response of microbial communities to biochar-amended soils: a critical review. Biochar 1:3–22. https://doi.org/10.1007/s42773-019-00009-2
Penn CJ, Camberato JJ (2019) A critical review on soil chemical processes that control how soil pH affects phosphorus availability to plants. Agriculture 9:120. https://doi.org/10.3390/agriculture9060120
Qayyum MF, Haider G, Iqbal M, Hameed S, Ahmad N, Rehman MZu, Majeed A, Rizwan M, Ali S (2021) Effect of alkaline and chemically engineered biochar on soil properties and phosphorus bioavailability in maize. Chemosphere 266:128980. https://doi.org/10.1016/j.chemosphere.2020.128980
Rafique M, Ortas I, Rizwan M, Chaudhary HJ, Gurmani AR, Munis MFH (2020) Residual effects of biochar and phosphorus on growth and nutrient accumulation by maize (Zea mays L.) amended with microbes in texturally different soils. Chemosphere 238:124710. https://doi.org/10.1016/j.chemosphere.2019.124710
Richardson AE, Simpson RJ (2011) Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol 156:989–996. https://doi.org/10.1104/pp.111.175448
Sarfraz R, Yang W, Wang S, Zhou B, Xing S (2020) Short term effects of biochar with different particle sizes on phosphorous availability and microbial communities. Chemosphere 256:126862. https://doi.org/10.1016/j.chemosphere.2020.126862
Shi W, Ju Y, Bian R, Li L, Joseph S, Mitchell DRG, Munroe P, Taherymoosavi S, Pan G (2020) Biochar bound urea boosts plant growth and reduces nitrogen leaching. Sci Total Environ 701:134424. https://doi.org/10.1016/j.scitotenv.2019.134424
Shi Y, Yu Y, Chang E, Wang R, Hong Z, Cui J, Zhang F, Jiang J, Xu R (2023) Effect of biochar incorporation on phosphorus supplementation and availability in soil: a review. J Soil Sedim 23:672–686. https://doi.org/10.1007/s11368-022-03359-w
Song D, Tang J, Xi X, Zhang S, Liang G, Zhou W, Wang X (2018) Responses of soil nutrients and microbial activities to additions of maize straw biochar and chemical fertilization in a calcareous soil. Europ J Soil Biol 84:1–10. https://doi.org/10.1016/j.ejsobi.2017.11.003
Streubel JD, Collins HP, Tarara JM, Cochran RL (2012) Biochar produced from anaerobically digested fiber reduces phosphorus in dairy lagoons. J Environ Qual 41:1166–1174. https://doi.org/10.2134/jeq2011.0131
Sun K, Qiu M, Han L, Jin J, Wang Z, Pan Z, Xing B (2018) Speciation of phosphorus in plant-and manure-derived biochars and its dissolution under various aqueous conditions. Sci Total Environ 634:1300–1307. https://doi.org/10.1016/j.scitotenv.2018.04.099
Tan X-F, Liu Y-G, Gu Y-l, Xu Y, Zeng G-M, Hu X-J, Liu S-B, Wang X, Liu S-M, Li J (2016) Biochar-based nano-composites for the decontamination of wastewater: a review. Bioresource Technol 212:318–333. https://doi.org/10.1016/j.biortech.2016.04.093
Tao Q, Li B, Li Q, Han X, Jiang Y, Jupa R, Wang C, Li T (2019) Simultaneous remediation of sediments contaminated with sulfamethoxazole and cadmium using magnesium-modified biochar derived from Thalia dealbata. Sci Total Environ 659:1448–1456. https://doi.org/10.1016/j.scitotenv.2018.12.361
Tian J, Wang J, Dippold M, Gao Y, Blagodatskaya E, Kuzyakov Y (2016) Biochar affects soil organic matter cycling and microbial functions but does not alter microbial community structure in a paddy soil. Sci Total Environ 556:89–97. https://doi.org/10.1016/j.scitotenv.2016.03.010
Tian XY, He DD, Bai S et al (2021) Physiological and molecular advances in magnesium nutrition of plants. Plant Soil 468:1–17. https://doi.org/10.1007/s11104-021-05139-w
Vitousek PM, Naylor R, Crews T, David MB, Drinkwater LE, Holland E, Johnes PJ, Katzenberger J, Martinelli LA, Matson PA (2009) Nutrient imbalances in agricultural development. Science 324:1519–1520. https://doi.org/10.2307/20494176
Wan W, Hao X, Xing Y, Liu S, Zhang X, Li X, Chen W, Huang Q (2021) Spatial differences in soil microbial diversity caused by pH-driven organic phosphorus mineralization. Land Degrada Devel 32:766–776. https://doi.org/10.1002/ldr.3734
Willey JM, Sherwood LM, Woolverton CJ (2014) Prescott’s microbiology. McGraw-Hill, New York
Wu L, Wei C, Zhang S, Wang Y, Kuzyakov Y, Ding X (2019) MgO-modified biochar increases phosphate retention and rice yields in saline-alkaline soil. J Clean Prod 235:901–909. https://doi.org/10.1016/j.jclepro.2019.07.043
Xiong J, Zhou M, Qu C, Yu D, Chen C, Wang M, Tan W (2021) Quantitative analysis of Pb adsorption on sulfhydryl-modified biochar. Biochar 3:37–49. https://doi.org/10.1007/s42773-020-00077-9
Xu G, Zhang Y, Sun J, Shao H (2016) Negative interactive effects between biochar and phosphorus fertilization on phosphorus availability and plant yield in saline sodic soil. Sci Total Environ 568:910–915. https://doi.org/10.1016/j.scitotenv.2016.06.079
Yang X, Wan Y, Zheng Y, He F, Yu Z, Huang J, Wang H, Ok YS, Jiang Y, Gao B (2019) Surface functional groups of carbon-based adsorbents and their roles in the removal of heavy metals from aqueous solutions: a critical review. Chem Eng J 366:608–621. https://doi.org/10.1016/j.cej.2019.02.119
Yao Y, Gao B, Inyang M, Zimmerman AR, Cao X, Pullammanappallil P, Yang L. (2011) Removal of phosphate from aqueous solution by biochar derived from anaerobically digested sugar beet tailings. J Hazard Mater 190(1-3), 501–507. https://doi.org/10.1016/j.jhazmat.2011.03.083
Yao Y, Gao B, Chen J, Yang L (2013) Engineered biochar reclaiming phosphate from aqueous solutions: mechanisms and potential application as a slow-release fertilizer. Environ Sci Technol 47:8700–8708. https://doi.org/10.1016/j.chemosphere.2012.06.002
Yin X, Xu Y, Huang R, Huang Q, Xie Z, Cai Y, Liang X (2017) Remediation mechanisms for Cd-contaminated soil using natural sepiolite at the field scale. Environ Sci Process Impacts 19:1563–1570. https://doi.org/10.1039/C7EM00262A
Yin Q, Wang R, Zhao Z (2018) Application of Mg–Al-modified biochar for simultaneous removal of ammonium, nitrate, and phosphate from eutrophic water. J Clean Prod 176:230–240
Zhan F, Zeng W, Yuan X, Li B, Li T, Zu Y, Jiang M, Li Y (2019) Field experiment on the effects of sepiolite and biochar on the remediation of Cd-and Pb-polluted farmlands around a Pb–Zn mine in Yunnan Province, China. Environ Sci Poll Res 26:7743–7751. https://doi.org/10.1007/s11356-018-04079-w
Zhang M, Gao B, Yao Y, Xue Y, Inyang M (2012) Synthesis of porous MgO-biochar nano-composites for removal of phosphate and nitrate from aqueous solutions. Chem Eng J 210:26–32. https://doi.org/10.1016/j.cej.2012.08.052
Zhang H, Chen C, Gray EM, Boyd SE, Yang H, Zhang D (2016) Roles of biochar in improving phosphorus availability in soils: a phosphate adsorbent and a source of available phosphorus. Geoderma 276:1–6. https://doi.org/10.1016/j.geoderma.2016.04.020
Zhang Y, Wang J, Feng Y (2021) The effects of biochar addition on soil physicochemical properties: a review. CATENA 202:105284.https://doi.org/10.1016/j.catena.2021.105284
Zhao Z, Ibrahim MM, Wang X, Xing S, Heiling M, Hood-Nowotny R, Tong C, Mao Y (2019) Properties of biochar derived from spent mushroom substrates. BioResources 14:5254–5277. https://doi.org/10.15376/biores.14.3.5254-5277
Zheng Y, Wan Y, Chen J, Chen H, Gao B (2020) MgO modified biochar produced through ball milling: a dual-functional adsorbent for removal of different contaminants. Chemosphere 243:125344. https://doi.org/10.1016/j.chemosphere.2019.125344
Zhou F, Ye G, Gao Y, Wang H, Zhou S, Liu Y, Yan C (2022) Cadmium adsorption by thermal-activated sepiolite: application to in-situ remediation of artificially contaminated soil. J Hazard Mater 423:127104. https://doi.org/10.1016/j.jhazmat.2021.127104
Zhu J, Li M, Whelan M (2018) Phosphorus activators contribute to legacy phosphorus availability in agricultural soils: a review. Sci Total Environ 612:522–537. https://doi.org/10.1016/j.scitotenv.2017.08.095
Zwetsloot MJ, Lehmann J, Bauerle T, Vanek S, Hestrin R, Nigussie A (2016) Phosphorus availability from bone char in a P-fixing soil influenced by root-mycorrhizae-biochar interactions. Plant Soil 408:95–105. https://doi.org/10.1007/s11104-016-2905-2
Acknowledgements
We acknowledge the Key Research Laboratory of Soil Ecosystem Health and Regulation, College of Resources and Environment, Fujian Agriculture and Forestry University, Fuzhou, China for the support provided during the conduct of the experiment, and the Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou for providing the material support and working environment that helped in developing the core idea of this study and completing its write-up.
Funding
Financial support was received from the Guangdong Basic and Applied Basic Research Foundation (2022B1515020014), China Postdoctoral Science Foundation (2023M743544), Fujian Forestry Science and Technology Promotion Project (2020TG17), and the University-Industry Cooperation Project of Fujian Province (2021N5002).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study’s conception and design. Material preparation, data collection, and analysis, data curation, investigation, methodology, writing of the original draft, review, and editing, were performed by [Muhammed Mustapha Ibrahim]. Review and editing was done by [Huiying Lin], [Zhaofeng Chang], [Zhimin Li], [Asif Riaz]. Supervision, review and editing, and resources were provided by [Enqing Hou]. All authors read, commented on, and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Handling editor: Yilai Lou.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ibrahim, M.M., Lin, H., Chang, Z. et al. Magnesium-doped biochars increase soil phosphorus availability by regulating phosphorus retention, microbial solubilization and mineralization. Biochar 6, 68 (2024). https://doi.org/10.1007/s42773-024-00360-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42773-024-00360-z