
Abstract
Alcohol abuse is a global health problem causing a substantial fraction of chronic liver diseases. Abundant TGF-β—a potent pro-fibrogenic cytokine—leads to disease progression. Our aim was to elucidate the crosstalk of TGF-β and alcohol on hepatocytes. Primary murine hepatocytes were challenged with ethanol and TGF-β and cell fate was determined. Fluidigm RNA analyses revealed transcriptional effects that regulate survival and apoptosis. Mechanistic insights were derived from enzyme/pathway inhibition experiments and modulation of oxidative stress levels. To substantiate findings, animal model specimens and human liver tissue cultures were investigated. Results: On its own, ethanol had no effect on hepatocyte apoptosis, whereas TGF-β increased cell death. Combined treatment led to massive hepatocyte apoptosis, which could also be recapitulated in human HCC liver tissue treated ex vivo. Alcohol boosted the TGF-β pro-apoptotic gene signature. The underlying mechanism of pathway crosstalk involves SMAD and non-SMAD/AKT signaling. Blunting CYP2E1 and ADH activities did not prevent this effect, implying that it was not a consequence of alcohol metabolism. In line with this, the ethanol metabolite acetaldehyde did not mimic the effect and glutathione supplementation did not prevent the super-induction of cell death. In contrast, blocking GSK-3β activity, a downstream mediator of AKT signaling, rescued the strong apoptotic response triggered by ethanol and TGF-β. This study provides novel information on the crosstalk between ethanol and TGF-β. We give evidence that ethanol directly leads to a boost of TGF-β’s pro-apoptotic function in hepatocytes, which may have implications for patients with chronic alcoholic liver disease.
Similar content being viewed by others

Introduction
Alcohol abuse is a major burden of Western countries. Excessive alcohol consumption may lead to fatty liver, fibrosis, and hepatitis. In many patients, continuous drinking causes cirrhosis, which significantly increases the risk for development of liver decompensation, hepatocellular carcinoma (HCC), and mortality1,2. Ethanol provokes an array of detrimental effects, systemically and on specific organs and cell types. Key features of alcoholic liver diseases (ALD) are upregulated oxidative stress levels, disturbed hepatocyte metabolism, increased influx of bacterial products to the liver (endotoxins, e.g., LPS), hepatic inflammation, and compromised regeneration as well as high mortality2.
Hepatocytes constitute the major cell type for the biotransformation of ethanol to acetaldehyde. For this, three major enzymatic systems are available3. In normal liver, alcohol dehydrogenase (ADH) is most relevant, using NADP+ as electron acceptor to oxidize ethanol to acetaldehyde. Additionally, catalase is capable of oxidizing ethanol. Finally, the microsomal ethanol oxidation systems (MEOS) accounts for rapid ethanol oxidation, with CYP2E1 being the key enzyme. Importantly, CYP2E1 is directly induced by ethanol. Ethanol metabolism increases reactive oxygen species (ROS) production, especially of H2O2 and superoxide anion (O2−), therewith weakening the cell’s defense system and causing direct cell damage, e.g., by lipid peroxidation4,5,6.
In the context of hepatocyte functionality, a key effect of ethanol is promotion of steatosis by increased fatty acid biosynthesis (via fatty acid synthase) and disturbed fatty acid oxidation (β-oxidation). Finally, it is well accepted that ethanol contributes to hepatic insulin resistance7.
Transforming growth factor beta1 (TGF-β) is a key pro-fibrotic cytokine in a broad range of chronic liver diseases and is abundant in ALD8,9,10,11,12,13. TGF-β dimers bind to receptor complexes (TGF-β receptors II and I) at the cell surface to initiate intracellular signaling cascades. The canonical pathway starts with C-terminal phosphorylation of SMAD2 and SMAD3 by TβRI, which enables interaction with SMAD4, forming a complex that translocates to the nucleus to regulate gene expression. Furthermore, several other pathways are triggered by TGF-β exposure in liver and other tissues, such as TAK1, PI3K/AKT, and small GTPase-dependent signaling networks14,15. TGF-β exerts a cell type specific response. It is a potent immune suppressor and mediates activation of quiescent hepatic stellate cells, therewith enhancing fibrogenesis. In epithelial cells, TGF-β may induce epithelial to mesenchymal transition (EMT), growth arrest, and promote apoptosis. In line with the latter, pro-apoptotic factors are induced by TGF-β in hepatocytes16, such as NOX4 leading to generation of ROS17,18 and other pro-apoptotic factors, such as BIM and BMF19. During liver regeneration, TGF-β accounts for ceasing of hepatocyte proliferation16,20,21.
Owing to the abundance of TGF-β in chronic liver diseases and the impact of ethanol on cellular integrity, we asked whether alcohol may modulate the hepatocyte’s response to this cytokine. In a previous investigation, we already showed that TGF-β signaling is induced in mouse livers after chronic ethanol insult as well as in human alcoholics. TGF-β treatment of mouse hepatocytes induced lipid-, oxidative stress metabolism-, and fibrogenesis-gene expression signatures. Interestingly, TGF-β downregulated expression of the alcohol metabolizing enzyme ADH1 via ALK5/SMAD2/3 signaling, thereby increasing alcohol-induced lipid accumulation and CYP2E1-dependent toxicity22. We now analyzed the molecular mechanism, by which TGF-β signaling and alcohol challenge integrate in cell fate decisions of hepatocytes. We found that TGF-β’s pro-apoptotic function is massively enhanced by ethanol. Noteworthy, this finding is not explained by increased SMAD signaling alone but rather by an alcohol-triggered communication with other signaling cascades leading to a distinct apoptosis-related gene expression signature. Curiously, this effect is not mediated by increased oxidative stress or ethanol’s oxidation to acetaldehyde, but rather involves an interference with AKT signaling and GSK-3β, which have previously been linked to apoptosis or survival, respectively23. In summary, we provide evidence that ethanol is a crucial modulator of TGF-β effects, which may have clinical implications in patients with pre-existing liver fibrosis even when consuming only moderate amounts of alcohol.
Results
Ethanol sensitizes culture-stressed hepatocytes for TGF-β-mediated apoptosis
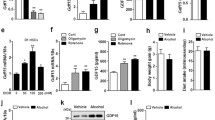
As ethanol abundance affects cellular integrity and functionality, we aimed at investigating its consequences on TGF-β-mediated cellular phenotypes. Initially, we used a biosensor chip (BIONAS 2500, Bionas GmbH, Rostock) to follow real-time changes in cellular morphology, adhesion, cell–cell interactions, and membrane functionality of mouse hepatocytes via impedance measurement in response to ethanol and TGF-β (Fig. 1a)24,25. Although TGF-β and ethanol alone did only marginally reduce cell impedance, combined application of both effectively reduced cell impedance. Estimated values of the combined treatment started to deviate from single substance treatment at 30 h (22 h following exposure) and resulted in a loss of impedance at 66 h (58 h). Changes in impedance are a good indicator for the initiation of apoptosis25,26, but can also reflect other changes in cellular interaction and adhesion properties, including EMT24. Because TGF-β is known as initiator of epithelial cell apoptosis, we next measured caspase-3 activity (Fig. 1b). Interestingly, only TGF-β led to activation of caspase-3 in individual treatments after 48 h (Fig. 1b). This was further increased significantly after combined application of ethanol and TGF-β. This pro-apoptotic signaling was further confirmed using immunoblot (Supplementary Fig. 1a, c). The effect of ethanol was dose-dependent (Supplementary Fig. 1b). Investigating the apoptotic route responsible we found by immunoblotting that mitochondrial cytochrome C release was strongest with co-treatment (72 h, Fig. 1d), which further indicates that ethanol sensitizes hepatocytes for TGF-β-mediated apoptosis. Next, Annexin-V staining with Hoechst 33342 counterstain of nuclei was performed. In agreement with the former results (Fig. 1b), strongest positivity for Annexin-V was detected upon co-treatment (Fig. 1e; Supplementary Fig. 1c). BCL2 had been demonstrated to counteract TGF-β-mediated apoptosis27. Hence, we overexpressed BCL2 in mouse hepatocytes utilizing an adenoviral construct. BCL2 over-expression completely blunted TGF-β-triggered cell death and prevented ethanol-dependent super-induction (Fig. 1f; Supplementary Fig. 1d).

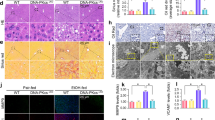
a Real-time response profile of cellular impedance upon ethanol/TGF-β treatment of mouse hepatocytes. Exposure to TGF-β (5 ng/ml) and/or ethanol (100 mM) started after 8 h equilibration of hepatocytes in the biosensor chip system and was continued over 58 h. Beginning of treatment is indicated by the dashed vertical line. RM = running medium (medium without substances). At beginning of treatment, RM is replaced by medium containing ethanol or TGF-β or both at the indicated concentrations. b Caspase-3 activity was measured in mouse hepatocytes stimulated with either TGF-β (5 ng/ml), ethanol (150 mM) or both for 48 h. The average activity of three independent experiments ± SEM is shown. Values were normalized to the total protein content of each sample. RFU = relative fluorescence units. c Detection of cleaved caspase-3 by immunoblot analysis using hepatocyte lysates after 72 h treatment as described for b. Detection of β-actin was performed as loading control. d Cellular lysates of mouse hepatocytes treated as in c were separated into cytoplasmic and mitochondrial fractions, and presence of cytochrome C was detected by immunoblot analyses. Ab antibody. e Mouse hepatocytes treated with 5 ng/ml TGF-β1, 100 mM ethanol or a combination of both were stained with Annexin-V-Cy3 and Hoechst 33342 (nuclei). Resulting fluorescent signals were analyzed using the ImageJ Software (particle analyzer). Relative amounts (in %) of apoptotic cells are shown (N = 9 pictures analyzed per group). P-values relate to control group. ¥ indicates P < 0.001 between TGF-β and TGF-β + EtOH group. Normal distribution was tested by Kolmogorov–Smirnov test, followed by one-way ANOVA with Bonferroni’s multiple comparisons test. f Hepatocytes were transfected with adenoviral vectors either expressing LacZ (Ad-ctrl) or Bcl2 (Ad-Bcl2). 2 h after infection, viruses were removed, and after 2 h starvation, cells were stimulated with TGF-β and/or ethanol as indicated for 48 h. Caspase-3 activity was measured as in b and c. *** indicates a significant difference between LacZ + TGF-β and LacZ + TGF-β + EtOH, as measured by two-way ANOVA test, P < 0.001. g Effect of TGF-β, ethanol or both on caspase activation in extracts of human HCC tissues (N = 6). Combined stimulation with TGF-β and ethanol resulted in significantly increased caspase activation compared to all other treatment groups. Analyzed by one-way ANOVA, coupled with uncorrected Fisher’s least significant difference (LSD) test (in comparison to all other treatment groups; *P < 0.05)
We next investigated the effect of TGF-β and ethanol in fresh liver tissue explants from human HCC (N = 6). No significant differences in caspase-3/-7 activity were found between HCC tissues treated with ethanol or TGF-β and untreated control. This is probably due to a more robustly transdifferentiated phenotype in HCC cells, where the anti-proliferative function of TGF-β is replaced by tumor promoting actions of the same pathway. However, as in cultured cells, co-treatment with TGF-β and ethanol lead to resensitization with significantly increased caspase-3/-7 activity (Fig. 1g). These results demonstrate that the apoptosis sensitizing role of ethanol on TGF-β signaling is relevant also in the tissue in situ. The importance of TGF-β signaling is underlined by elevated phospho-Smad levels already in untreated HCC tissues (Supplementary Fig. 1e, f). The possible therapeutic impact of this finding on therapy of HCC patients needs to be determined in another study.
Ethanol- and TGF-β-mediated apoptosis gene expression signature in mouse hepatocytes
To more comprehensively delineate pathway components participating in the TGF-β apoptotic response of mouse hepatocytes upon sensitization with ethanol, we assessed the transcription profile of a selected panel of pro- and anti-apoptotic genes 12, 24, and 48 h after treatment, using the Fluidigm real-time PCR platform. Strongest changes occurred at early time points (Fig. 2a, b). Compared to single treatments, a substantial fraction of target genes (including Bim, Bik, and Bmf) was significantly increased by co-treatment. Noteworthy, although several anti-apoptotic genes were also regulated, pro-apoptotic factors were stronger induced at all time points. At 48 h, ethanol alone did not affect apoptotic genes any more, except for increased Cyp2e1 expression (Fig. 2c). In contrast, TGF-β still kept pro-apoptotic factors at higher expression levels, e.g. Nox4, Bim, and Bmf, which was more pronounced in the presence of ethanol. These changes over time and the boosting effect of ethanol on TGF-β target gene expression is illustrated in Fig. 2d for Bim, Bik, Bmf, and Gadd45a. At 12 h, super-induction from co-treatment is evident for all targets, whereas at 24 h, this effect only increased further for Bim. After 48 h, only Gadd45a was still super-induced, but did not reach significance any more (Fig. 2c). With regard to known ethanol target genes, Cyp2e1 was increased at all time points and anti-apoptotic Bcl2 only at 12 h (Fig. 2e). Upon co-treatment, TGF-β did not further increase expression of these ethanol target genes, indicating that the synergistic effect only occurs for TGF-β-regulated genes, but not vice versa. To summarize, ethanol is a potent sensitizer for the TGF-β-regulated apoptosis-related gene expression program in hepatocytes, leading to super-induction, especially at early time points (12 and 24 h).

a–c Mouse hepatocytes were treated with 5 ng/ml TGF-β, 150 mM ethanol, and both for 12 h (a), 24 h (b), and 48 h (c). Expression profiling was conducted with Fluidigm qPCR technology. Changes in expression are shown (fold) relative to untreated. The lists are sorted by TGF-β group descending in fold expression. Analyses were conducted with one-way ANOVA with multiple comparison and Tukey correction. ns. not significant; # anti-apoptotic genes. d Time course analyses (assembled from data of a–c) of strongly altered pro-apoptotic genes (Bim, Bik, Bmf, and Gadd45a). For all apoptosis-promoting targets, co-treatment of TGF-β and ethanol triggered strongest induction. e Time-resolved pattern of ethanol metabolizing enzyme Cyp2e1 and the anti-apoptotic gene Bcl2 (data derived from a–c)
Ethanol sensitizes hepatocytes for TGF-β signaling by upregulating TβRII expression
Next, we tested whether ethanol directly modulates the TGF-β signaling pathway in mouse hepatocytes. In time course experiments, we found mRNA and protein levels of TβRII significantly increased upon ethanol treatment (direct TGF-β-induced upregulation was very mild), as measured by conventional RT-PCR (Fig. 3a), real-time PCR (Supplementary Fig. 2a), and immunoblotting (Fig. 3b). In mice, ethanol intoxication similarly significantly induced TβRII expression in liver tissue, 12 and 24 h post gavage (Fig. 3c). These data indicate that acute alcohol intoxication induces expression of rate limiting components of the TGF-β signaling pathway, thereby enhancing responsiveness of the cells. This was further supported by significantly increased phosphoprotein levels of SMADs −1 and −3 in cultured mouse hepatocytes upon TGF-β treatment when the cells were pretreated with ethanol (Fig. 3d, Supplementary Fig. 2b). In line with this finding, TGF-β downstream signaling via SMAD3/4 was enhanced by ethanol co-treatment, as shown in reporter assays upon adenoviral infection of cultured mouse hepatocytes with a SMAD binding element (CAGA) luciferase reporter construct (Fig. 4a).

a–c Mouse hepatocytes were incubated with ethanol (150 mM) for 12, 24, 48, and 72 h, as indicated. Subsequent mRNA analyses (conventional PCR, a) and immunoblot analyses (b) revealed significant induction of TBRII (most pronounced at the earliest time point; P < 0.05 for all time points, two-sided unpaired t-test). c Livers from mice having received an alcohol binge were isolated 12 and 48 h post gavage. mRNA expression of TβRII was significantly elevated at both time points (two-sided unpaired t-test). d Immunoblot analysis of TGF-β-treated (time as indicated, 5 ng/ml) or -untreated (ctrl) mouse hepatocytes, pretreated or not with ethanol (48 h, 150 mM), showing phospho-SMAD1 and phospho-SMAD3 levels and GAPDH as control

a TGF-β/SMAD3 signaling reporter assay using Ad(CAGA)9-MLP-Luc infection of mouse hepatocytes (for 2 h during attachment phase) and TGF-β/ethanol (5 ng/ml; 150 mM) treatment, as indicated. Obtained values (relative light units, RLU) were normalized to protein content. The experiment was performed in triplicates. b Immunoblot analysis of CTGF expression and c real-time RT-PCR analyses of Smad6 and Smad7 expression in mouse hepatocytes treated with TGF-β and ethanol as indicated and described in Fig. 1b
Finally, co-treatment significantly increased canonical TGF-β target gene expression, as measured at the protein level for CTGF (Fig. 4b, Supplementary Fig. 2c) and at the mRNA level for Smad6 and Smad7 (Fig. 4c). Thus, ethanol potently enhances the TGF-β signaling cascade in mouse hepatocytes.
Ethanol affects survival-signaling networks in hepatocytes and redirects TGF-β signaling towards apoptosis
Culture of mouse hepatocytes in 2D as monolayer represents a cellular stress situation with upregulated survival signaling and downregulated cytostatic responsiveness28,29,30. Moreover, thorough expression profiling indicated that this culture condition phenocopies cellular stress in mouse models and human patients with liver diseases29. Owing to our finding that in combination, ethanol challenge and TGF-β signaling are central in twisting the balance of the homeostasis of the hepatocyte phenotype towards a higher sensitivity for cell damage, we speculated that the survival pathway via AKT might be involved. We and others have previously shown that AKT activity is crucial for resistance to TGF-β-mediated apoptosis31,32. Co-treatment demonstrated significant abrogation of AKT phosphorylation at Ser473 (P < 0.01, N = 6)—a finding not observed by single treatments (Fig. 5a), with unchanged total AKT levels (Supplementary Fig. 3a). Using the PI3K/AKT small molecule inhibitor LY294002 in two different concentrations, we could phenocopy the ethanol effect on sensitizing mouse hepatocytes for apoptotic TGF-β signaling (Supplementary Fig. 3b). AKT is a negative regulator of GSK-3β and thus blocking GSK-3β function using a chemical inhibitor (SB216763) significantly reduced caspase-3 activity upon co-treatment (Fig. 5b, P = 0.005). Noteworthy, SB216763 neither affected basal nor direct TGF-β or ethanol induced apoptosis (also Supplementary Fig. 3c). Thus, AKT/GSK-3β signaling is pivotal in mediating the synergistic TGF-β and ethanol triggered hepatocyte death. In line with this, PTEN, a negative regulator of AKT activation was directly induced by ethanol treatment (Supplementary Fig. 3d).

a A representative immunoblot of TGF-β (5 ng/ml)- and ethanol (150 mM)-treated murine hepatocytes (48 h) shows alterations in phospho-AKT levels upon treatment, normalized (to GAPDH). Densitometric analyses of N = 6 experiments indicates a significant (P < 0.01) reduction in AKT phosphorylation (Ser473) in the TGF-β/EtOH compared to the untreated group (two-tailed t-test). b The GSK-3β inhibitor SB216763 (10 µM) was used to study the influence of GSK-3β on TGF-β- and ethanol-triggered apoptosis. A caspase-3 assay revealed that only upon co-treatment, blockage of GSK-3β reduced programmed cell death in murine hepatocytes (treatment for 48 h, P > 0.001, two-tailed t-test). Co, DMSO control; SB216763, 10 µM
Ethanol, but not acetaldehyde, triggers the apoptotic effect
Owing to the rapid metabolism of ethanol to acetaldehyde and the known toxicity of this metabolite, we aimed at exploring whether the apoptotic phenotype is a direct effect of ethanol or is due to its conversion to acetaldehyde. Treating hepatocytes directly with 0.2 and 2 mM acetaldehyde instead of ethanol, did not increase TGF-β-mediated induction of caspase-3 activity (Fig. 6a). This indicates that the super-induction of hepatocyte death is directly mediated by ethanol itself. To further support this hypothesis, we investigated whether generation of oxidative stress might have a role in this phenomenon.

a The ethanol metabolite acetaldehyde does not affect TGF-β-mediated apoptosis in murine hepatocytes. Co-treatment of TGF-β (5 ng/ml) with ethanol (150 mM) led to enhanced hepatocyte apoptosis (P < 0.01). Its metabolite acetaldehyde was tested with 0.2 and 2 mM in combination with TGF-β for 48 h. Caspase-3 activity was not altered compared to only TGF-β treatment (ANOVA with multiple comparison and Tukey correction). b–e The role of oxidative stress in ethanol’s enhancement of apoptosis. b Caspase-3 assay was performed to analyze the effect of oxidative stress on TGF-β-mediated apoptosis. Increased activity of GOX (glucose oxidase) leads to production of reactive oxygen species in the cell medium, with catalase (CAT) supplementation balancing this effect. Reduced catalase levels led to increased oxidative stress, and upon TGF-β (5 ng/ml) addition, a GOX dose-dependent effect on caspase-3 activity was measured. ¥¥P < 0.01 indicates comparison of GOX/CAT 1:200,000 + TGF-β vs. no-GOX/no-CAT + TGF-β. TGF-β + GOX is highly significant compared with all other treatment groups (one-way ANOVA with multiple comparison and Tukey correction). c Supplementation of the antioxidant N-acetylcystein (NAC, 2 and 4 mM) significantly reduced TGF-β-mediated apoptosis, as measured by caspase-3 activity (48 h treatment). * indicates comparison with untreated group, ¥ indicates comparison with TGF-β only treatment. 4 mM NAC supplementation entirely blunted TGF-β-mediated apoptosis in murine hepatocytes (one-way ANOVA with multiple comparison and Tukey correction). d Blocking ADH (with 4-MP, 0.75 mM) and CYP2E1 (with DAS, 0.5 mM) enzymatic activities did not affect ethanol (150 mM)-mediated elevation of TGF-β-triggered hepatocyte apoptosis, as measured by caspase-3 activity (after 48 h). No significant difference was found in the TGF-β + ethanol group with inhibitor treatments. e NAC treatment (4 mM) in the context of TGF-β and ethanol-triggered apoptosis (48 h) revealed that the ethanol boost in programmed cell death is not blocked by reducing oxidative stress. ¥ indicates a highly significant difference between TGF-β treatment and TGF-β + ethanol treatment. NAC blunted TGF-β-mediated apoptosis, as also shown in c (one-way ANOVA with multiple comparison and Tukey correction). f Schematic illustration of ethanol’s mode of action on TGF-β-mediated apoptosis. Ethanol co-treatment leads to downregulation of the pro-survival AKT pathway, which enables GSK-3β functionality. This event then leads to enhancement of apoptosis. In parallel, ethanol affects TGF-β pathway components, which may also enhance apoptotic signaling. TGF-β/SMAD signaling regulates anti- and pro-apoptotic genes, although the latter are stronger induced and push the balance towards programmed cell death. Oxidative stress is not involved in ethanol’s modulation of TGF-β-triggered apoptosis
Oxidative stress and ethanol metabolism
It is well accepted that oxidative stress is an essential component of TGF-β-mediated apoptosis. Furthermore, Nox4, a TGF-β target gene, has previously been documented as trigger for hepatocyte apoptosis17,18. Hence, we applied an enzymatic system, which generates a stable level of reactive oxygen species, the so called GOX/CAT system33. With that, we could demonstrate that indeed increased oxidative stress is able to amplify TGF-β-induced caspase-3 activity (Fig. 6b). Looking for gene expression effects, we found that GOX/CAT treatment induced TβRII and Cyp2E1 expression and blunted TGF-β-induced expression of the survival gene Bcl2 (Supplementary Fig. 4). Furthermore, interfering with oxidative stress formation by application of N-acetylcysteine (NAC) yielded in reduced caspase-3 activity upon TGF-β exposure (Fig. 6c). As ethanol metabolism increases ROS levels, we blocked the major ethanol metabolizing enzymes, namely ADH and CYP2E1 with the inhibitors 4-MP22 and diallyl sulfide (DAS), respectively. Interestingly, neither blocking ADH or CYP2E1, nor their combined inhibition reduced ethanol-mediated hyperactivation of caspase-3 upon TGF-β exposure (Fig. 6d). Finally, we assessed the consequences of NAC on apoptosis super-induction. As shown before (Fig. 6c), TGF-β-mediated apoptosis was abolished in the presence of NAC, but NAC did not affect apoptosis induction by co-treatment (Fig. 6e). In conclusion, ethanol triggers TGF-β-mediated apoptosis neither by formation of its metabolite acetaldehyde nor via the production of oxidative stress, suggesting that the observed switch of TGF-β signaling from survival to death is a direct effect of alcohol, whereby mechanistic details still largely remain undetermined.
Discussion
We describe a detrimental crosstalk between TGF-β signaling and ethanol exposure. Such scenario is relevant for alcohol patients with existing fibrosis, being frequently associated with increased TGF-β levels. Previously, we showed that cell damage-mediated lactate dehydrogenase release increases by such co-treatment due to elevated oxidative stress6,34. Here, we found that mouse hepatocytes underwent apoptosis upon co-treatment, whereas ethanol alone had no effect. Super-induction of apoptosis was mediated by mitochondrial cytochrome C release, supporting our previous results22, and confirmed by the blunting effect of BCL235 on TGF-β and co-stimulation triggered apoptosis. We, however, do not take into account crosstalk with other resident liver cells or hepatocyte organization in lobular structures. Monolayer hepatocytes present with cellular stress and rapid dedifferentiation, reflecting a damage and plasticity fate, as present in human and mouse liver diseases, including liver cancer36. We further observed that stressed hepatocytes display activated survival signaling pathways, including ERK and AKT28,30. This is a prerequisite for switching TGF-β signaling from cytostatic to tumorigenic in the liver. The pro-apoptotic-signaling branch is abrogated and the alternate tumorigenic pathway is activated, comprising production of secreted growth factors for tumorigenic stroma formation, e.g., CTGF, PDGF, and VEGF37,38,39,40 and enhanced growth factor receptor expression supporting cell survival. In addition, gene expression changes occur for epithelial to mesenchymal transition, including upregulated snail and vimentin, and decreased ZO-1 and E-cadherin41,42,43, a TGF-β response previously summarized as late TGF-β signature and that significantly correlated with bad prognosis in HCC patients44.
To translate our in vitro findings to human disease, we analyzed liver tissue explants from HCC patients for the apoptotic response to TGF-β and ethanol. Noteworthy, in HCC tissue co-treatment induced the strongest response, indicating that this mechanism is conserved in primary tissue as well. Although being aware that TGF-β effects in HCC in principle are pro-tumorigenic, the effect of ethanol on cell death was obvious and may hint for a mechanism switching back the TGF-β pathway to cytostasis. Moreover, a therapeutic option for HCCs which cannot be treated by resection or radiofrequency ablation is the injection of pure ethanol. According to current knowledge, the mechanism of ethanol action is via triggering necrosis, partially by damaging small vessels with subsequent ischemia45,46. In our approach, we applied lower doses of ethanol, and found the in vitro described synergism with TGF-β is valid in HCC tissue as well, inducing cancer cell apoptosis. In high dose treatments, this may have implications at tumor margins, where only low concentrations of ethanol are reached.
To gain more insight on transcriptional regulation by ethanol, we screened for gene expression changes of the TGF-β pathway components. TβRII was upregulated by ethanol in vivo and in vitro. These observations point to a direct stimulating function of ethanol on the TGF-β pathway. Indeed, it has been shown that acetaldehyde, the major metabolite of ethanol, can trigger TβRII expression in hepatic stellate cells and can also cause release and activation of latent TGF-β147,48,49. A positive regulation of the TGF-β signaling cascade by ethanol is also concluded from our analyses of SMAD2/3 phosphorylation, SMAD binding element reporter activation, and CTGF expression. CTGF is known to foster fibrotic processes and is a direct target of TGF-β50,51. Although we found highest induction of CTGF upon co-treatment with TGF-β and ethanol, we do not have evidence that other pro-fibrotic events in hepatocytes were regulated similarly (e.g., enhanced expressions of Snail, Col1a1, or Vimentin; data not shown). Elevated expression was also determined for Smad6 and Smad7. These inhibitory SMADs provide negative feedback on the TGF-β/BMP-pathways52. Noteworthy, in addition to enhanced Smad2/3 phosphorylation, we observed elevated activation of Smad1 by TGF-β upon ethanol pretreatment. Although Smad1 is the classical mediator of the canonical BMP signaling pathway, it can also be activated by TGF-β, especially at very early time points53. Besides TGF-β signaling components, we addressed whether the ethanol induced pro-apoptotic phenotype is accompanied by changes in expression of pro- and anti-apoptotic genes. Indeed, especially after 12 and 24 h, a super-induction of apoptosis-related genes was found in the co-treatment cohort. Although at 48 h, this effect had mainly faded out, the apoptotic phenotype was still manifest. As this analysis handled gene expression data, a conclusion on protein expression or activation status could not be drawn. Yet, the alcohol effect is evident at early time points, therewith paving the way for abrogated cell survival and increased TGF-β-mediated apoptotic rates of the hepatocyte population. Execution of apoptosis is balanced by pro- and anti-apoptotic factors35. Thus, it is not surprising that also anti-apoptotic genes are partially upregulated by TGF-β and alcohol in our experimental setting, but in summary, the equilibrium got dysbalanced in favor of apoptosis execution.
As non-SMAD pathways have fundamental roles in mediating TGF-β effects, especially on cell survival, we analyzed AKT and ERK signaling pathway activities. ERK activity was not affected by ethanol treatment (not shown), but a significant reduction in AKT Ser473 phosphorylation was identified in co-treated hepatocytes. A major role of AKT interference on directing TGF-β signaling towards cell death has been acknowledged31, making it a likely candidate for mediating the super-induction of caspase-3. An interesting regulatory mechanism was demonstrated with AKT reducing NOX4 activity, thus counteracting programmed cell death17. Reduced AKT activity upon co-treatment might deplete such preventive mechanism. Downstream signaling events of AKT comprise inactivation of GSK-3β by Ser9 phosphorylation. GSK-3β has previously been linked to promoting apoptosis23 by controlling expression of a panel of apoptosis regulating factors. The targeted inhibition of GSK-3β activity by SB216763 yielded in loss of co-treatment triggered apoptosis super-induction. On the contrary, TGF-β-mediated apoptosis was not affected, providing strong support for the AKT/GSK-3β axis having a dominant role especially in ethanol’s enhancement effect on TGF-β-mediated cell death.
The question whether ethanol directly accounts for the rise in TGF-β-triggered death or whether this was rather executed by its metabolism was also addressed. As acetaldehyde was described as major cell-damaging effector upon ethanol oxidation, we replaced ethanol by acetaldehyde and tested the apoptotic response. Although, as discussed above, acetaldehyde is capable of triggering TβRII induction as well as TGF-β1 activation48,49, in our experimental setting, acetaldehyde did not cause increased hepatocyte death. Furthermore, next to excluding acetaldehyde as mediator, we could rule out a role of ethanol metabolism (and thus oxidative stress). Oxidative stress by its own increased TGF-β driven apoptosis and in a model with over-expression of CYP2E1, TGF-β exerted enhanced cell damage54. Further, stabilization of cellular stress defense capacity by addition of NAC protected against TGF-β driven programmed cell death. However, this treatment regimen could not overcome the co-stimulatory effect on apoptosis. The two major ethanol oxidizing systems, MEOS and ADH1, produce ROS. Thus, blocking the enzymatic systems by inhibitors should prevent alcohol-dependent production of reactive oxygen and acetaldehyde. Addition of these inhibitors did not prevent the co-treatment effect on hepatocyte survival, which further demonstrates that ethanol and not its metabolism is directly mediating the enhanced TGF-β phenotype.
In conclusion, we unravel a crosstalk of TGF-β signaling and ethanol in hepatocytes with detrimental consequences for survival (a schematic illustration is provided in Fig. 6f). We further show that this crosstalk integrates at the AKT/GSK-3β axis to interfere with cell dedifferentiation associated survival, thus sensitizing hepatocytes for TGF-β-induced cell death. We and others have previously shown that in liver, fine tuning of TGF-β downstream signaling via specific adaptor proteins and activity of survival signaling pathways triggers cellular fate and subsequent consequences for the organ. Mishra and coworkers have shown that the adapter protein β2 spectrin is required for SMAD3-mediated cytostatic TGF-β signaling in hepatocytes, and that its downregulation, e.g., by induced expression of the E3 ligase PRAJA and subsequent β2 spectrin ubiquitinylation and proteasomal degradation facilitates cancer stem cell development and HCC formation55,56,57. Similarly, in an investigation of enhanced liver damage upon combined HCV infection and alcohol consumption, loss of cytostatic TGF-β signaling was the critical step in cancer stem cell origination and HCC development. In detail, HCV infection induced TLR4 expression in hepatocytes, thus sensitizing hepatocytes for alcohol-induced LPS. Downstream signaling induced expression of the stemness factor Nanog, which in turn induces the oncogenes Igfbp3 and Yap1. Both integrate at the TGF-β signaling pathway, YAP1 via stabilizing SMAD7 and IGFBP3 via activating AKT/mTOR signaling, together efficiently interfering with the cytostatic TGF-β response58,59. These findings underline that controlled cytostatic TGF-β signaling is required for hepatocyte physiology. On the other hand, recent work from Seki and colleagues indicates that counteracting survival signaling that dampens cytostasis is also necessary for cellular homeostasis in liver. In detail, they show that cell stress induced by high fat diet, similarly as in our setting with alcohol, results in enhanced TGF-β-mediated cell death. Furthermore, they found that high fat diet downregulates TAK1-mediated survival signaling, thus directing TGF-β signaling towards lipogenic gene expression and abrogation of the hepatocyte antioxidant response60. The group then fully depleted hepatocytes for TAK1 survival signaling in vivo and these mice rapidly developed aggressive liver cancer, which was completely rescued by additionally depleting hepatocytes of TβRII. In TAK1 knockout mice, TGF-β induced massive hepatocyte apoptosis, which in consequence resulted in strong inflammation and compensatory proliferation, together causing hepatocarcinogenesis61.
Thus, the presented data in the context of the current state of the art further indicate that the TGF-β signaling pathway has strong and critical impact on hepatocyte fate decisions that are directly related to liver diseases, including ASH, NASH, and HCC. Thereby, TGF-β downstream signaling is integrated into and controlled from a network of survival signaling pathways. Future studies need to shed light on the relevance of these findings in patients suffering from alcoholic liver disease and the possible consequences of continued drinking in a pre-damaged liver scenario with TGF-β abundance.
Material and methods
Reagents
Antibodies against pAKT (#4051), pGSK-3β (#9323), cytochrome C (#4280), and cleaved caspase-3 (#9664) were purchased from Cell Signaling (Leiden, Netherlands), pSMAD1/3 (#1880-1) was obtained from Epitomics (Abcam, Germany), GAPDH from SantaCruz (sc-47724, Heidelberg, Germany), and β-actin (#A5441) from Sigma (Taufkirchen, Germany). Secondary antibodies carrying HRP were obtained from SantaCruz. Diallyl sulfide (DAS), 4-Methylpyrazole hydrochloride (4-MP), N-Acetyl-l-cysteine (NAC), and SB216763 were from Sigma. Ethanol (absolute) and acetaldehyde were purchased from Merck (Darmstadt, Germany). TGF-β1 (“TGF-β”) was from Peprotech (Hamburg, Germany) and used at a concentration of 5 ng/ml. LY294002 was from Sigma-Aldrich and used as indicated.
Cell isolation/culture/treatment procedures
Primary hepatocytes (from C57BL/6 mice) were isolated by collagenase perfusion technique as described elsewhere28. For cell culture experiments, cells were seeded as monolayer on collagen coated plates. Culture plates were pre-coated with a solution of 250 μg/ml rat tail collagen I, followed by UV light-mediated crosslinking28, yielding a thin layer of monomeric collagen. Williams E served as medium, supplemented during attachment phase (4 h) with Pen/Strep, l-glutamine (2 mM), dexamethasone (100 nM), and FCS (10%). After attachment, cells were treated with TGF-β or ethanol (50–150 mM as indicated) in Williams E medium supplemented with l-glutamine (2 mM) and Pen/Strep. Wells were covered with parafilm to reduce ethanol evaporation62.
Adenovirus infection
The adenovirus encoding Bcl2 was used at an MOI of 10 to prevent TGF-β-mediated cell death. As control, AdLacZ was utilized at the same MOI. Cells were infected for 2 h during the attachment phase. Subsequently, viruses were removed, and cells were washed and used for stimulation experiments.
Caspase-3 assay
A fluorimetric-based enzymatic caspase-3 assay was used as described previously31. In brief, cells were lysed and equal protein amounts were used for the assay. As caspase-3 substrate, AC-DEVD-AFC (Biomol, Hamburg, Germany) was used and after incubation for 1 h with cell lysate, fluorescence was measured (excitation 400 nm; emission 505 nm) with a Tecan infinite M200 (Thermo Scientific, Braunschweig, Germany) plate reader (96 well format). Y-axis in arbitrary units (relative fluorescence units; RFU, normalized to protein concentration).
Annexin-V staining
As another technique to analyze hepatocyte apoptosis, Annexin-V staining was performed. After over night serum-starvation, mouse hepatocytes were treated with 5 ng/ml TGF-β1, 100 mM ethanol, or a combination of both for 24 h. Residual culture medium was washed off with PBS (2 mM CaCl2). Cells were stained for 5 min with 0.25 µg/ml Annexin-V-Cy3, 5 µg/ml Hoechst 33342, and 150 nM Sytox Green in PBS. Unbound stain was removed by washing with PBS (2 mM CaCl2) and fluorescent signals were detected immediately and analyzed using the ImageJ Software (particle analyzer).
Cytochrome C release
To quantify cytoplasmic and mitochondrial cytochrome C, we used the Qproteome Mitochondria Isolation Kit from Qiagen (Hilden, Germany). Therewith, we were able to generate cytosolic and mitochondrial protein lysates. Subsequently, cytochrome C content was assessed by immunoblotting using a cytochrome C-specific antibody.
SMAD luciferase reporter assay
The Ad(CAGA)9-MLP-Luc was used as a SMAD3 luciferase reporter (adenoviral infection for 2 h during attachment phase) to estimate SMAD transcriptional activity63. Luciferase substrate was purchased from Promega (Mannheim, Germany). Luminometric measurements were performed using Tecan infinite M200. Obtained values (relative light units, RLU) were normalized to protein content. Experiments were performed in triplicates. Shown are the means of three experiments.
PCR (conventional and Fluidigm qRT-PCR)
Total RNA from hepatocytes was isolated using RNAeasy Mini Kit (Qiagen). Overall, 600 ng total RNA was reverse transcribed to cDNA with TaqMan Reverse Transcription Reagents (Applera GmbH, Darmstadt, Germany). Conventional PCR was performed as previously described64. For quantitative Real-time PCR, the Fluidigm Biomark high throughput qRT-PCR platform (Fluidigm Corporation, San Francisco, USA) was used with pre-designed taqman probes from Applied Biosystems, according to the manufacturer’s instructions65. Data were analyzed using the ΔΔCt method and expression values were normalized to the expression levels of Gapdh and β-actin. For each condition, 4–6 independent hepatocyte isolation sets were used (N = 6 for 48 h time point; N = 4 for 12 and 24 h time points). For each time point, four treatment groups were used (control, TGF-β, ethanol, TGF-β + ethanol). Expression changes upon treatment were calculated relative to the corresponding untreated control.
Impedance measurement
Impedance was measured using a microfluidic flow biosensor chip system (BIONAS 2500, Bionas GmbH, Rostock, Germany) containing two interdigitated electrodes, which provide a measure of the cell layer permeability and changes in cell shape based on extracellular ion mobility (flux), using electrical properties of the cell layer as described earlier24,25. Real-time response profiling of cellular impedance upon ethanol/TGF-β treatment of primary mouse hepatocytes was performed as follows. Cells were seeded as monolayer at high density on the chip surface and were equilibrated in sensor chip culture medium with continuous medium exchange in 4 and 4 min (for equilibration) and 4 and 30 min (for long-term cultivation) flow/stop cycles. Exposure to ethanol/TGF-β started after 8 h of equilibration of hepatocytes in the biosensor chip system and was continued over 58 h. Beginning of treatment is indicated by the dashed vertical line. At beginning of treatment, RM (running medium without substances) is replaced by medium containing ethanol and TGF-β at indicated concentrations; control experiments are performed with RM; treatment is continued until the end of experiment after 66 h.
Culture of liver explants and caspase-3/7 assay
We investigated freshly isolated, unfixed HCC tissues from HCC patients (N = 6) who underwent partial hepatectomy. This study was approved by the ethics committee of Hannover Medical School. Briefly, HCC explants were precisely divided into 125 mm3 cubes under sterile conditions and incubated with modified Eagle’s medium supplemented with TGF-β, ethanol, or both for 8 h. Following incubation, liver tissue was pulverized in liquid nitrogen and lysed with lysis buffer as described previously66,67. Tissue extracts were diluted in buffer containing 50 mM Tris-HCl (pH 7.4), 10 mM KCl, and 5% glycerol to reach a final protein concentration of 1 mg/ml. Activity of caspase-3/-7 was measured in duplicates by a luminescent substrate assay (Caspase-Glo; Promega, Mannheim, Germany). For this purpose, 10 µl of tissue extracts were incubated with 10 µl of caspase substrate DEVD-luciferin and luciferase reagent for 2 h at room temperature. Luminescence of the samples was measured in relative light units (RLU) by a luminometer (LB 960, Berthold, Bad Wildbad, Germany).
GOX/CAT system
This system makes use of glucose oxidase (GOX) in the culture supernatant, producing reactive oxygen species (H2O2). To achieve a steady level, catalase (CAT) is added aiming at degrading reactive oxygen species. Enzymatic rates of the enzymes are described in ref.33. We used GOX with a 1:300,000 and CAT with a 1:50,000 or 1:200,000 dilution, as indicated.
Acute alcohol intoxication—in vivo experiment
All procedures were approved by the local Institutional Animal Care and Use Committee (IACUC) and the Regierungspräsidium Stuttgart (application number V250/07 EM). Experiments were carried out in a facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) at the University of Hohenheim. Female C57BL/6J mice (8 weeks old) received 6 g ethanol per kg body-weight (once) per oral gavage as previously described68. After 12 and 48 h, animals were sacrificed, the livers preserved and further processed for RNA isolation. Experiments were not blinded.
Immunoblotting
Cell lysates were prepared using RIPA buffer (50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Nonidet P-40 (NP40), 0.5% sodium deoxycholate, 0.1% SDS, Proteases Inhibitor Cocktail, and Phosphatase Inhibitor Cocktail II (Roche, Mannheim, Germany))69. Protein concentration was determined using the DC protein assay (Bio-Rad). Up to 30 μg protein per sample were separated by SDS-PAGE (12%). Subsequently, proteins were transferred to a nitrocellulose membrane and blocked in 5% non-fat milk in TBST. Then, membranes were incubated with indicated antibodies (first antibodies overnight at 4 °C and second antibodies at room temperature for 1 h) and developed with ECL solution (Pierce) using Chemi-Smart imager (Vilber Lourmat, Eberhardzell, Germany). Aida Image Analyzer v. 4.25 was used to perform densitometric analyses.
Data availability
The data sets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Statistics
Analyses were performed using GraphPad Prism Software. Two-sided unpaired t-test was applied as indicated. Normal distribution of values was tested with the Kolmogorow–Smirnov test. Subsequently, one-way ANOVA tests were used with multiple comparison and Tukey’s correction or with uncorrected Fisher’s least significant difference (LSD) test. Shown are data of three experiments (or as indicated) with SEM. Significance: *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001.
References
Stickel, F. Alcoholic cirrhosis and hepatocellular carcinoma. Adv. Exp. Med. Biol. 815, 113–130 (2015).
Beier, J. I. & McClain, C. J. Mechanisms and cell signaling in alcoholic liver disease. Biol. Chem. 391, 1249–1264 (2010).
Louvet, A. & Mathurin, P. Alcoholic liver disease: mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 12, 231–242 (2015).
Bailey, S. M. & Cunningham, C. C. Acute and chronic ethanol increases reactive oxygen species generation and decreases viability in fresh, isolated rat hepatocytes. Hepatology 28, 1318–1326 (1998).
Seth, D., Haber, P. S., Syn, W. K., Diehl, A. M. & Day, C. P. Pathogenesis of alcohol-induced liver disease: classical concepts and recent advances. J. Gastroenterol. Hepatol. 26, 1089–1105 (2011).
Nussler, A. K. et al. Protective role of HO-1 for alcohol-dependent liver damage. Dig. Dis. 28, 792–798 (2010).
Sugimoto, K. & Takei, Y. Pathogenesis of alcoholic liver disease. Hepatol. Res. 47, 70–79 (2017).
Flisiak, R. & Prokopowicz, D. Transforming growth factor-beta1 as a surrogate marker of hepatic dysfunction in chronic liver diseases. Clin. Chem. Lab. Med. 38, 1129–1131 (2000).
Daniluk, J., Szuster-Ciesielska, A., Drabko, J. & Kandefer-Szerszen, M. Serum cytokine levels in alcohol-related liver cirrhosis. Alcohol 23, 29–34 (2001).
Meyer, C., Meindl-Beinker, N. M. & Dooley, S. TGF-beta signaling in alcohol induced hepatic injury. Front. Biosci. 15, 740–749 (2010).
Dooley, S. & ten Dijke, P. TGF-beta in progression of liver disease. Cell. Tissue. Res. 347, 245–256 (2012).
Zhang, S., Sun, W. Y., Wu, J. J. & Wei, W. TGF-beta signaling pathway as a pharmacological target in liver diseases. Pharmacol. Res. 85, 15–22 (2014).
Chen, W. X. et al. Quantitative analysis of transforming growth factor beta 1 mRNA in patients with alcoholic liver disease. World J. Gastroenterol. 8, 379–381 (2002).
Meyer, C., Liu, Y. & Dooley, S. Caveolin and TGF-beta entanglements. J. Cell Physiol. 228, 2097–2102 (2013).
Mu, Y., Gudey, S. K. & Landstrom, M. Non-Smad signaling pathways. Cell. Tissue. Res. 347, 11–20 (2012).
Arendt, E., Ueberham, U., Bittner, R., Gebhardt, R. & Ueberham, E. Enhanced matrix degradation after withdrawal of TGF-beta1 triggers hepatocytes from apoptosis to proliferation and regeneration. Cell Prolif. 38, 287–299 (2005).
Caja, L. et al. Overactivation of the MEK/ERK pathway in liver tumor cells confers resistance to TGF-{beta}-induced cell death through impairing upregulation of the NADPH oxidase NOX4. Cancer Res. 69, 7595–7602 (2009).
Carmona-Cuenca, I. et al. Upregulation of the NADPH oxidase NOX4 by TGF-beta in hepatocytes is required for its pro-apoptotic activity. J. Hepatol. 49, 965–976 (2008).
Ramjaun, A. R., Tomlinson, S., Eddaoudi, A. & Downward, J. Upregulation of two BH3-only proteins, Bmf and Bim, during TGF beta-induced apoptosis. Oncogene 26, 970–981 (2007).
Russell, W. E., Coffey, R. J. Jr, Ouellette, A. J. & Moses, H. L. Type beta transforming growth factor reversibly inhibits the early proliferative response to partial hepatectomy in the rat. Proc. Natl Acad. Sci. USA 85, 5126–5130 (1988).
Nakamura, T. et al. Suppression of transforming growth factor-beta results in upregulation of transcription of regeneration factors after chronic liver injury. J. Hepatol. 41, 974–982 (2004).
Ciuclan, L. et al. TGF-beta enhances alcohol dependent hepatocyte damage via downregulation of alcohol dehydrogenase I. J. Hepatol. 52, 407–416 (2010).
Rahmani, M. et al. Dual inhibition of Bcl-2 and Bcl-xL strikingly enhances PI3K inhibition-induced apoptosis in human myeloid leukemia cells through a GSK3- and Bim-dependent mechanism. Cancer Res. 73, 1340–1351 (2013).
Schatzschneider, U. et al. Cellular uptake, cytotoxicity, and metabolic profiling of human cancer cells treated with ruthenium(II) polypyridyl complexes [Ru(bpy)2(N–N)]Cl2 with N–N=bpy, phen, dpq, dppz, and dppn. ChemMedChem 3, 1104–1109 (2008).
Alborzinia, H. et al. Real-time monitoring of cisplatin-induced cell death. PLoS ONE 6, e19714 (2011).
Holenya, P. et al. Detailed analysis of pro-apoptotic signaling and metabolic adaptation triggered by a N-heterocyclic carbene-gold(I) complex. Metallomics 6, 1591–1601 (2014).
Lafon, C. et al. Transforming growth factor beta 1-induced apoptosis in human ovarian carcinoma cells: protection by the antioxidant N-acetylcysteine and bcl-2. Cell Growth Differ. 7, 1095–1104 (1996).
Godoy, P. et al. Extracellular matrix modulates sensitivity of hepatocytes to fibroblastoid dedifferentiation and transforming growth factor beta-induced apoptosis. Hepatology 49, 2031–2043 (2009).
Godoy, P. et al. Gene network activity in cultivated primary hepatocytes is highly similar to diseased mammalian liver tissue. Arch. Toxicol. 90, 2513–2529 (2016).
Zellmer, S. et al. Transcription factors ETF, E2F, and SP-1 are involved in cytokine-independent proliferation of murine hepatocytes. Hepatology 52, 2127–2136 (2010).
Meyer, C., Liu, Y., Kaul, A., Peipe, I. & Dooley, S. Caveolin-1 abrogates TGF-beta mediated hepatocyte apoptosis. Cell Death Dis. 4, e466 (2013).
Valdes, F. et al. Transforming growth factor-beta activates both pro-apoptotic and survival signals in fetal rat hepatocytes. Exp. Cell Res. 292, 209–218 (2004).
Mueller, S., Millonig, G. & Waite, G. N. The GOX/CAT system: a novel enzymatic method to independently control hydrogen peroxide and hypoxia in cell culture. Adv. Med. Sci. 54, 121–135 (2009).
Ehnert, S. et al. The right choice of antihypertensives protects primary human hepatocytes from ethanol- and recombinant human TGF-beta1-induced cellular damage. Hepatic Med. 5, 31–41 (2013).
Cosentino, K. & Garcia-Saez, A. J. Mitochondrial alterations in apoptosis. Chem. Phys. Lipids 181, 62–75 (2014).
Godoy, P. et al. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch. Toxicol. 87, 1315–1530 (2013).
Mazzocca, A., Fransvea, E., Lavezzari, G., Antonaci, S. & Giannelli, G. Inhibition of transforming growth factor beta receptor I kinase blocks hepatocellular carcinoma growth through neo-angiogenesis regulation. Hepatology 50, 1140–1151 (2009).
Mazzocca, A. et al. Down-regulation of connective tissue growth factor by inhibition of transforming growth factor beta blocks the tumor-stroma cross-talk and tumor progression in hepatocellular carcinoma. Hepatology 51, 523–534 (2010).
Aigner, K. et al. The transcription factor ZEB1 (deltaEF1) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity. Oncogene 26, 6979–6988 (2007).
van Zijl, F. et al. Hepatic tumor-stroma crosstalk guides epithelial to mesenchymal transition at the tumor edge. Oncogene 28, 4022–4033 (2009).
Giannelli, G., Bergamini, C., Fransvea, E., Sgarra, C. & Antonaci, S. Laminin-5 with transforming growth factor-beta1 induces epithelial to mesenchymal transition in hepatocellular carcinoma. Gastroenterology 129, 1375–1383 (2005).
Fransvea, E., Angelotti, U., Antonaci, S. & Giannelli, G. Blocking transforming growth factor-beta up-regulates E-cadherin and reduces migration and invasion of hepatocellular carcinoma cells. Hepatology 47, 1557–1566 (2008).
Lahsnig, C. et al. ILEI requires oncogenic Ras for the epithelial to mesenchymal transition of hepatocytes and liver carcinoma progression. Oncogene 28, 638–650 (2009).
Coulouarn, C., Factor, V. M. & Thorgeirsson, S. S. Transforming growth factor-beta gene expression signature in mouse hepatocytes predicts clinical outcome in human cancer. Hepatology 47, 2059–2067 (2008).
Llovet, J. M., Sala, M. & Bruix, J. Nonsurgical treatment of hepatocellular carcinoma. Liver Transplant. 6, S11–S15 (2000).
Dusheiko, G. M., Hobbs, K. E., Dick, R. & Burroughs, A. K. Treatment of small hepatocellular carcinomas. Lancet 340, 285–288 (1992).
Ceni, E., Mello, T. & Galli, A. Pathogenesis of alcoholic liver disease: role of oxidative metabolism. World J. Gastroenterol. 20, 17756–17772 (2014).
Svegliati-Baroni, G. et al. Early response ofalpha2(I) collagen to acetaldehyde in human hepatic stellate cells is TGF-beta independent. Hepatology 42, 343–352 (2005).
Chen, A. Acetaldehyde stimulates the activation of latent transforming growth factor-beta1 and induces expression of the type II receptor of the cytokine in rat cultured hepatic stellate cells. Biochem. J. 368, 683–693 (2002).
Liu, Y. et al. Transforming growth factor-beta (TGF-beta)-mediated connective tissue growth factor (CTGF) expression in hepatic stellate cells requires Stat3 signaling activation. J. Biol. Chem. 288, 30708–30719 (2013).
Brigstock, D. R. Connective tissue growth factor (CCN2, CTGF) and organ fibrosis: lessons from transgenic animals. J. Cell Commun. Signal. 4, 1–4 (2010).
Yan, X., Liu, Z. & Chen, Y. Regulation of TGF-beta signaling by Smad7. Acta. Biochim. Biophys. Sin. (Shanghai). 41, 263–272 (2009).
Chen, S. et al. Transforming growth factor-beta1 (TGF-beta1) activates hepcidin mRNA Expression in Hepatocytes. J. Biol. Chem. 291, 13160–13174 (2016).
Zhuge, J. & Cederbaum, A. I. Increased toxicity by transforming growth factor-beta 1 in liver cells overexpressing CYP2E1. Free Radic. Biol. Med. 41, 1100–1112 (2006).
Baek, H. J. et al. Transforming growth factor-beta adaptor, beta2-spectrin, modulates cyclin dependent kinase 4 to reduce development of hepatocellular cancer. Hepatology 53, 1676–1684 (2011).
Thenappan, A. et al. Role of transforming growth factor beta signaling and expansion of progenitor cells in regenerating liver. Hepatology 51, 1373–1382 (2010).
Kitisin, K. et al. Disruption of transforming growth factor-beta signaling through beta-spectrin ELF leads to hepatocellular cancer through cyclin D1 activation. Oncogene 26, 7103–7110 (2007).
Machida, K. et al. Toll-like receptor 4 mediates synergism between alcohol and HCV in hepatic oncogenesis involving stem cell marker Nanog. Proc. Natl Acad. Sci. USA 106, 1548–1553 (2009).
Chen, C. L. et al. Reciprocal regulation by TLR4 and TGF-beta in tumor-initiating stem-like cells. J. Clin. Invest. 123, 2832–2849 (2013).
Yang, L. et al. Transforming growth factor beta signaling in hepatocytes participates in steatohepatitis through regulation of cell death and lipid metabolism in mice. Hepatology 59, 483–495 (2014).
Yang, L. et al. Transforming growth factor-beta signaling in hepatocytes promotes hepatic fibrosis and carcinogenesis in mice with hepatocyte-specific deletion of TAK1. Gastroenterology 144, 1042–1054 e4 (2013).
Eysseric, H. et al. There is not simple method to maintain a constant ethanol concentration in long-term cell culture: keys to a solution applied to the survey of astrocytic ethanol absorption. Alcohol 14, 111–115 (1997).
Meyer, C. et al. Distinct role of endocytosis for Smad and non-Smad TGF-beta signaling regulation in hepatocytes. J. Hepatol. 55, 369–378 (2011).
Li, Q. et al. Bone morphogenetic protein-9 induces epithelial to mesenchymal transition in hepatocellular carcinoma cells. Cancer Sci. 104, 398–408 (2013).
Spurgeon, S. L., Jones, R. C. & Ramakrishnan, R. High throughput gene expression measurement with real time PCR in a microfluidic dynamic array. PLoS ONE 3, e1662 (2008).
Wahl, K. et al. Increased apoptosis induction in hepatocellular carcinoma by a novel tumor-targeted TRAIL fusion protein combined with bortezomib. Hepatology 57, 625–636 (2013).
Volkmann, X. et al. Increased hepatotoxicity of tumor necrosis factor-related apoptosis-inducing ligand in diseased human liver. Hepatology 46, 1498–1508 (2007).
Wagnerberger, S. et al. Sex-specific differences in the development of acute alcohol-induced liver steatosis in mice. Alcohol Alcohol. 48, 648–656 (2013).
Meyer, C. et al. Distinct dedifferentiation processes affect caveolin-1 expression in hepatocytes. Cell Commun. Signal. 11, 6 (2013).
Acknowledgements
We highly appreciate technical assistance of Mei Han, Weronika Piorońska, Katarina Abramovic, Alexandra Müller, and Sabine Wagnerberger. This work was supported by the DFG (ME4532/1-1), the BMBF to SD (Virtual Liver Network, VLN, Grant No. 0315764 and 0315755), and EU funding FP7 to SD (IT-Liver consortium, Marie Curie Training Network).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing financial interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gaitantzi, H., Meyer, C., Rakoczy, P. et al. Ethanol sensitizes hepatocytes for TGF-β-triggered apoptosis. Cell Death Dis 9, 51 (2018). https://doi.org/10.1038/s41419-017-0071-y
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-017-0071-y
- Springer Nature Limited
This article is cited by
-
Maternal Ethanol Exposure-Induced Cardiac Fibrosis is Associated with Changes in TGF-β and SIRT1/FOXO3a Signaling in Male Rat Offspring: A Three-Month Follow-up Study
Cardiovascular Toxicology (2022)
-
Berberine inhibits epithelial-mesenchymal transition and promotes apoptosis of tumour-associated fibroblast-induced colonic epithelial cells through regulation of TGF-β signalling
Journal of Cell Communication and Signaling (2020)