

Abstract
Tissue injury triggers activation of mesenchymal lineage cells into wound-repairing myofibroblasts, whose unrestrained activity leads to fibrosis. Although this process is largely controlled at the transcriptional level, whether the main transcription factors involved have all been identified has remained elusive. Here, we report multi-omics analyses unraveling Basonuclin 2 (BNC2) as a myofibroblast identity transcription factor. Using liver fibrosis as a model for in-depth investigations, we first show that BNC2 expression is induced in both mouse and human fibrotic livers from different etiologies and decreases upon human liver fibrosis regression. Importantly, we found that BNC2 transcriptional induction is a specific feature of myofibroblastic activation in fibrotic tissues. Mechanistically, BNC2 expression and activities allow to integrate pro-fibrotic stimuli, including TGFβ and Hippo/YAP1 signaling, towards induction of matrisome genes such as those encoding type I collagen. As a consequence, Bnc2 deficiency blunts collagen deposition in livers of mice fed a fibrogenic diet. Additionally, our work establishes BNC2 as potentially druggable since we identified the thalidomide derivative CC-885 as a BNC2 inhibitor. Altogether, we propose that BNC2 is a transcription factor involved in canonical pathways driving myofibroblastic activation in fibrosis.
Similar content being viewed by others
Introduction
Fibrosis is characterized by the excessive accumulation of structurally abnormal extracellular matrix (ECM) that can lead to disruption of tissue architecture and function, and ultimately organ failure1. As such, fibrosis has a major clinical impact on patients’ morbidity and mortality. This pathological condition is a consequence of an excessive wound healing response caused by repeated or chronic injuries that can affect a wide range of organs, including the kidneys, the lungs, the heart, and the liver1,2. Cirrhosis is the terminal stage of progressive liver fibrosis, which has various etiologies. While the incidence of fibrosis caused by viral hepatitis is declining, fibrosis triggered by a spectrum of liver diseases ranging from alcoholic-related liver diseases (ARLD) to non-alcoholic steatohepatitis (NASH) is on the rise. In particular, NASH is rapidly becoming one of the leading causes of liver failure worldwide with respect to the ever-increasing prevalence of obesity3,4. While fibrotic-related diseases have been estimated to account for as much as 45% of all-cause mortality rates in developed countries5, few therapeutic strategies are currently available and the medical need for effective anti-fibrotic therapies is high2,5. As of today, only bariatric surgery has been able to trigger significant liver fibrosis regression in a cohort of patients with obesity-associated NASH6. In this context, a better knowledge of the molecular mechanisms involved in fibrosis development is urgently needed.
Despite being initially triggered in an organ- and condition-specific manner, fibrosis converges toward conserved core cellular and molecular pathways1,7. A central and common event in fibrogenesis is the unrestrained activation of local tissue fibroblasts or mesenchymal-like cells into highly contractile and ECM-producing myofibroflasts (MFs)1,8. Myofibroblastic activation is triggered by intercellular crosstalks within inflamed injured organs where transforming growth factor (TGF)-β is considered the most potent pro-fibrotic cytokine9. Importantly, mechanosensing of the microenvironment stiffness through the Hippo/Yes-Associated Protein 1 (YAP1) pathway has recently emerged as a central trigger of both initial and self-sustained myofibroblastic activation10,11. Indeed, expression of ECM encoding genes and ECM-associated genes such as modifying enzymes, together defined as the matrisome12, allows MFs to increase both ECM amount and stiffness13. In particular, MFs massively secrete fibrillar collagens such as type I and III collagens (COL-I and COL-III) and promote their stabilization through cross-linking by lysyl-oxidases (LOX)9,13.
Myofibroblastic activation relies on a profound transcriptomic reprogramming allowing cells often considered as “quiescent” in healthy conditions to adopt the MF phenotype. This has triggered a growing interest in defining the transcriptional regulators involved in this process. In that regard, despite reported tissue specificities, MFs harbor conserved functionalities across organs such as ECM production pointing to a potentially conserved transcriptional regulatory program. This is exemplified by the described role for TGFβ-induced SMAD transcription factors and for the YAP1 co-regulator in MFs from diverse tissues8. In this study, we have implemented an unbiased functional genomics approach to assess candidate transcription factors (TFs) driving the MF phenotype. Our study pointed to Basonuclin 2 (BNC2), an understudied TF that belongs to the Zinc-finger family14, as specifically expressed in MFs with a predicted role in ECM synthesis. In line, we have found that myofibroblastic activation requires BNC2 up-regulation and that BNC2 expression is a characteristic of MFs from mouse and human fibrotic organs. We have also defined how BNC2 integrates pro-fibrotic signals to directly drive the transcriptional induction of matrisome genes in MFs, and finally we have shown that, as a consequence, BNC2 plays a key role in ECM deposition during liver fibrogenesis. Altogether, our study has uncovered BNC2 as a core TF driving myofibroblastic activation in fibrosis.
Results
Functional genomics analyses identify BNC2 as a MF identity TF
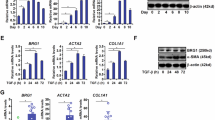
Leveraging epigenomic data has recently been established as a robust mean to identify genes defining the identity and specific functionalities of a given cell type15,16. In particular, identity genes are characterized by broad histone H3 lysine 4 trimethylated (H3K4me3) domains around their transcriptional start site (TSS), a feature functionally linked to enhanced transcriptional activity and consistency15,17. Hence, in order to identify key genes for MF identity and function, we mined H3K4me3 chromatin immunoprecipitation experiments coupled to DNA sequencing (ChIP-seq) data from human primary MFs, obtained by spontaneous in vitro activation of hepatic stellate cells (HSCs)18, the main source of MFs in the liver19,20. MFs obtained upon in vitro activation lack induction of environmentally induced inflammatory genes, but the transcriptional program responsible for the cell-intrinsic and canonical MF functions is appropriately induced8,21,22,23. Chromatin regions enriched for H3K4me3 signal in MFs were identified and split into sharp and broad H3K4me3 domains (Fig. 1A and Supplementary Fig. 1A). As expected, promoters characterized by broad H3K4me3 domains were more active as indicated by wider H3K27ac ChIP-seq signals around their TSS (Supplementary Fig. 1B) and higher mRNA expression levels of associated genes (Supplementary Fig. 1C) when compared to genes with sharp H3K4me3 domains. Also, promoters with broad H3K4me3 were characterized by shorter distances to nearest super-enhancers (Supplementary Fig. 1D), another defining feature of identity genes24. Among genes labeled with broad H3K4me3 domains, we have previously defined that identity genes are best captured by broad H3K4me3 domains which are not ubiquitously conserved across cell types25. Indeed, conserved broad H3K4me3 domains rather point to genes, such as tumor suppressor genes, involved in essential cell functions25,26. Hence, in order to more specifically discriminate potential identity genes, well-annotated genes linked to broad MF H3K4me3 domains were retrieved (n = 1278), and length of H3K4me3 labeling at these genes was monitored in 76 other human cell types and tissues26. Hierarchical clustering was next used to sort both genes and samples according to the relative length of gene H3K4me3 domains in the different analyzed cell types. This allowed to identify 3 main gene clusters denoted as clusters 1, 2, and 3, comprising 130, 318, and 830 genes, respectively (Fig. 1B and Supplementary Data file 1). Cluster 1 stood out as comprising genes whose H3K4me3 domain length was remarkably and preferentially high in a cluster of samples comprising MFs together with MF-like cells such as in vitro grown mesenchymal cells and muscle stem cells27 (Fig. 1B and Supplementary Data File 2). Consistently, mining transcriptomic data from 561 primary human cells (Supplementary Data File 3) indicated that MFs were characterized by high expression of cluster 1 genes. Indeed, when cells were independently ranked according to the average expression of genes from clusters 1, 2, and 3, a biased ranking of MFs towards highly expressing cells was evidenced for cluster 1 (Fig. 1C). Moreover, pathway and gene ontology enrichment analyses revealed that, while genes from clusters 2 and 3 were specifically enriched for essential cell functions or signaling pathways, genes from cluster 1 were more significantly enriched for terms related to acquisition of a mesenchymal phenotype including ECM synthesis and organization, i.e., MF canonical functions (Fig. 1D and Supplementary Fig. 1E). In line, cluster 1 comprised the greatest proportion of matrisome genes (Fig. 1E). Altogether, these data pointed to cluster 1 genes as particularly relevant with regards to the MF molecular identity.

A Breadth distribution of H3K4me3 domains called from ChIP-seq data from human primary MF-HSCs. The cumulative frequency distribution of H3K4me3 domain lengths is shown. The inflexion point of the curve was used to separate sharp from broad H3K4me3 domains. B Heatmap showing the relative length of H3K4me3 domains in 76 human cell types or tissues for 1278 genes linked to a broad H3K4me3 domain in primary MFs identified in (A). Hierarchical clustering was used to cluster both genes (rows) and samples (columns). Clusters were visualized on the heatmap using dotted lines. Three main gene clusters were identified and labeled clusters 1–3 (Supplementary Data File 1). At the top of the heatmap, the identity of cell types or tissues within the 5 main sample clusters are summarized (details are provided in Supplementary Data File 2). C 561 primary human cells were ranked based on increasing median expression levels of genes from cluster 1, 2, or 3. Next, the position of 64 MF cell types was indicated by purple horizontal bars within the ranking obtained using genes from cluster 1, cluster 2 or cluster 3. D Top 20 terms retrieved by Metascape88 when searching for pathways enriched within genes from clusters 1–3 (Hallmark Gene Sets, Reactome Gene Sets, KEGG Pathway, WikiPathways, Canonical Pathways, and PANTHER Pathways were used). Hierarchical clustering based on statistical significance is shown. White color indicates a lack of significance. Terms colored in red are preferentially enriched in cluster 1. E Proportion of matrisome genes in clusters 1–3. The percentage of matrisome genes in each cluster was retrieved by comparing genes from clusters 1–3 with a list of 1028 genes encoding ECM and ECM-associated proteins12. Statistical significance was assessed using two-sided Fisher exact tests with Benjamini–Hochberg correction for multiple testing.
In order to define potential key transcriptional regulators operating in MFs, we screened the list of MF identity genes from cluster 1 for the presence of TFs. 25 TFs were identified, among which several had already been described to play a role in the control of myofibroblastic activation or had been linked to fibrosis, including Forkhead Box (FOX), T-Box (TBX), Small Mothers Against Decapentaplegic (SMAD), TWIST and SNAIL family members (Fig. 2A). Importantly, our strategy also pointed out TFs more seldomly or never previously reported to be important in MFs, including the poorly studied zinc-finger TFs called Zinc Finger Homeobox 4 (ZFHX4) and Basonuclin 2 (BNC2). Further supporting these predictions, we found BNC2 expression levels to be high in MFs in general regardless of their tissue of origin (Fig. 2B). In line, BNC2 expressing cells also showed higher expression of the MF signature genes Actin Alpha 2, Smooth Muscle (ACTA2 also known as α-SMA) and Collagen Type I Alpha 1 Chain (COL1A1) (Fig. 2B). Moreover, Gene-Module Association Determination annotation (G-MAD), which predicts gene functions by analyzing co-expressed genes across large-scale transcriptomic data from human samples28, pointed out genes linked to ECM-related functions when interrogated using BNC2 (Fig. 2C). These predicted functional relationships hold true for many different human organs when they were individually considered (Supplementary Fig. 2A). While ZFHX4 was also largely expressed across MFs (Supplementary Fig. 2B), G-MAD scores linking ZFHX4 to ECM were not as strong across different human organs (Supplementary Fig. 2A). Moreover, similar analyses using TBX4, a previously described lung MF-specific TF (Supplementary Fig. 2A, C, D)29, further highlighted the strong and widespread association of BNC2 with ECM-related terms (Supplementary Fig. 2A). Finally, in addition to being characterized by a broad H3K4me3 domain and H3K27ac super-enhancer, we found that the BNC2 gene was also densely bound by the acetylated histone epigenetic reader BRD4 for its expression in MFs (Supplementary Fig. 3A). Consequently, induction of BRD4 degradation with MZ1 led to reduced BNC2 levels (Supplementary Fig. 3B, C). This is in line with BNC2 defining an identity TF since expression of identity TF genes is characterized by its sensitivity to BRD4 inhibition25,30.

A Network-based presentation of the 25 TFs identified by our approach. The connection between TFs was provided by String108. TFs are colored according to the number of PubMed citations linking a given TF to MFs or fibrosis as detailed in the Methods section. TFs of the HOXB family were grouped together. TFs in the green background have no reported link to MFs or fibrosis. B Analysis performed as in (C) except cells were ranked according to increasing BNC2 expression levels. COL1A1 and ACTA2 expression levels are displayed using the same color scale. Ranking of 64 MF cell types is indicated at the bottom using purple horizontal bars. C Prediction of BNC2-associated functions by the Gene-module association Determination (G-MAD) tool. The top five terms (highest G-MAD scores) are shown.
Altogether, these data prompted us to perform an in-depth characterization of the role exerted by BNC2 in myofibroblastic activation and fibrosis.
Induction of BNC2 expression specifically characterizes myofibroblastic activation in fibrotic tissues
To characterize BNC2 expression with regards to myofibroblastic activation, we performed in-depth analyses of BNC2 expression upon both murine and human liver fibrogenesis. Increased Col1a1 expression was used throughout these experiments as a molecular marker of fibrosis. We found that Bnc2 expression was increased in mouse fibrotic livers obtained through repeated injection of carbon tetrachloride (CCl4) for 8 weeks, a classical model of hepatotoxicity-induced liver fibrosis (Fig. 3A and Supplementary Fig. 4A). Importantly, this induction specifically stemmed from myofibroblastic hepatic stellate cells (MF-HSCs). Indeed, sorting the main liver cell types (hepatocytes (Hep), Kupffer cells (KC), HSC, cholangiocytes (Chol), and liver sinusoidal endothelial cells (LSEC)) from CCl4-injected mice showed that Bnc2 gene expression was specifically induced in MF-HSCs (Fig. 3B). In line, single-cell RNA-seq analyses of the livers of CCl4 treated mice31 indicated that Bnc2 transcripts were specifically found in MF-HSCs expressing the highest levels of Col1a1 (Supplementary Fig. 5A–D). Of note, BNC2 induction upon in vivo myofibroblastic activation extended to lung or heart injury (Supplementary Fig. 5E) in line with BNC2 being a marker of MFs across organs. In the liver, Bnc2 expression was also induced when mice were subjected to diet-induced fibrosis, i.e., high fat, sucrose and cholesterol diet (HFSC)32 and choline-deficient l-amino-acid-defined HFSC (CDAA-HFSC) diet33 (Fig. 3C, D and Supplementary Fig. 4B, C). Single-nuclei RNA-seq analysis of liver cells from mice fed a HFSC diet34 further pointed to Bnc2 being expressed in MFs (Supplementary Fig. 6). Hence, Bnc2 expression was consistently induced in different mouse liver fibrosis models.

A RT-qPCR data showing increased Bnc2 (n = 12 mice) and Col1a1 (n = 11 mice) expression in the fibrotic livers of mice injected with CCl4. Control mice were injected with olive oil (n = 4 mice). **P = 0.011. B RT-qPCR analysis of Bnc2 and Col1a1 expression in FACS-sorted cells obtained from the livers of mice injected with CCl4 (n = 6 mice) or olive oil (n = 7 mice). Hep. hepatocytes, KC Kupffer cells, HSC hepatic stellate cells, Chol. cholangiocytes, LSEC liver sinusoidal endothelial cells. *P = 0.0111. C RT-qPCR data showing increased Bnc2 and Col1a1 expression in the livers of mice fed the HFSC diet for 24 weeks (n = 11 mice) or a standard rodent chow diet (control, n = 10 mice). ***P < 0.0001. D RT-qPCR data showing increased Bnc2 and Col1a1 mRNA expression in the livers of mice fed the CDAA-HFSC diet for 10–12 weeks (n = 8 mice) or a standard rodent chow diet (control, n = 7 mice for Bnc2 and 6 for Col1a1). *P = 0.0289 and ***P = 0.0007. Graphs in panels A–D show means ± SD. Statistical significance was assessed using two-tailed Mann–Whitney U test (A and C, D) or two-way ANOVA with Sidak multiple comparison post hoc test (B, CCl4 vs control). Source data are provided as a Source Data file.
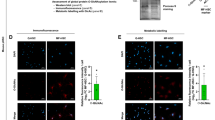
To monitor BNC2 expression in human livers, we first used samples from a cohort of patients with alcohol-related cirrhosis (Supplementary Data File 4)35. RT-qPCR assays showed that BNC2 expression is significantly induced in human cirrhotic livers when compared to histologically normal areas of livers from patients who underwent tumor resections (Fig. 4A). In line, BNC2 protein expression levels were specifically detected in livers with alcohol-related cirrhosis together with that of type I collagen (COL-I) and ACTA2 (Fig. 4B; see Supplementary Fig. 7 for anti-BNC2 specificity controls). Importantly, RNAscope mRNA in situ hybridization performed on four human cirrhotic livers indicated that BNC2 and COL1A1 mRNA expression co-localize to the fibrous tissue, while their expression is low within the hepatocyte areas and in non-fibrotic control livers (Fig. 4C and Supplementary Fig. 8A).

A RT-qPCR data showing increased BNC2 and COL1A1 expression in the livers of patients presenting alcohol-related cirrhosis (n = 42 biologically independent samples) versus control biopsies from the nontumoral areas of liver resections (n = 20 biologically independent samples). Box plots are composed of a box from the 25th to the 75th percentile with the median as a line and min to max as whiskers. Statistical significance was assessed using two-tailed Mann–Whitney U test. ***P < 0.0001. B Western blot assays showing BNC2, ACTA2 and type I Collagen (COL-I) protein expression in the livers of patients with alcohol-related cirrhosis (= 8 biologically independent samples) and control samples from the nontumoral areas of liver resections (= 6 biologically independent samples). Sub-cellular fractionation was performed for the detection of BNC2 (chromatin fraction) and ACTA2 (cytosolic fraction). H3 and GAPDH were used as loading controls. MW molecular weight markers. C Localization of BNC2 (white staining) and COL1A1 (red staining) mRNA in human alcohol-related cirrhotic livers. In situ RNA hybridation (RNAscope) was performed on paraffin-embedded liver sections obtained from patients presenting alcohol-related cirrhosis (n = 4 biologically independent samples) or control samples from the nontumoral areas of liver resections (n = 2 biologically independent samples). Dotted lines delimitate the hepatocyte and fibrous areas. C shows a representative image obtained from the liver of a patient with alcohol-related cirrhosis (Patient #A). Data obtained with livers from additional donors are shown in Supplementary Fig. 8A. D RNAscope analyses of BNC2 and COL1A1 mRNA expression in liver biopsies from patients with obesity-associated NASH showing bariatric surgery-induced loss of liver fibrosis (Kleiner Score F3 before surgery to F0/F1 1–5 years after surgery; n = 4 biologically independent sets of samples). D shows representative images obtained from the liver of one patient (Patient #G). Data obtained with livers from additional donors (n = 3) are shown in Supplementary Fig. 8C. The number of dots/cell at baseline and after surgery are shown in the right panel. Statistical significance was assessed two-tailed ratio paired t test. *P = 0.0416 or 0.0426 for BNC2 and COL1A1, respectively. Source data are provided as a Source Data file.
Interrogating our previous transcriptomic analyses36 indicated that BNC2 is also significantly upregulated in human NASH-associated fibrosis (Supplementary Fig. 8B and Supplementary Data file 5). Since liver fibrosis resolution involves loss of MF-HSCs through different mechanisms, including MF-HSC apoptosis, senescence, or deactivation37, we hypothesized that fibrosis resolution would be linked to a loss of BNC2 expression. To test this hypothesis, we leveraged liver biopsies obtained from a long-term follow-up study of a subset of patients with NASH-associated fibrosis who underwent bariatric surgery resulting in liver fibrosis regression (Kleiner score F3 at the time of surgery to Kleiner score F0/1 when monitored 1–5 years after surgery)6. Interestingly, liver fibrosis regression obtained through bariatric surgery was accompanied by reduced BNC2 expression levels as demonstrated by RNAscope assays in four patients (Fig. 4D and Supplementary Fig. 8C).
Altogether, these data indicated that induction of BNC2 expression is an intrinsic feature of myofibroblastic activation which characterizes mouse and human liver fibrosis.
BNC2 integrates pro-fibrotic stimuli in MF-HSCs
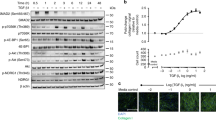
In order to define the functional role exerted by BNC2 in MFs, a combination of cistromic and epigenomic analyses were performed in the human MF-HSC cell line LX2. ChIP-seq was used to define the genome-wide binding sites of BNC2 using an antibody qualified through rapid immunoprecipitation mass spectrometry of endogenous protein (RIME) experiments38 as being able to immunoprecipitate cross-linked BNC2 (Supplementary Data File 6). We identified 1479 BNC2 binding sites, which occurred in active transcriptional regulatory regions as judged through measurement of chromatin accessibility using Column Purified chromatin (CoP)-seq39 and histone acetylation using H3K27ac ChIP-seq (Fig. 5A and Supplementary Data File 7). Enrichment for H3K27ac at identified BNC2 binding sites also specifically extended to primary human MF-HSCs (Supplementary Fig. 9A). De novo motif discovery retrieved the previously proposed BNC2 consensus binding motif40 as the top enriched motif (Supplementary Fig. 9B). To further mine the BNC2 cistrome, we next used the CistromeDB ChIP-seq database toolkit41 to monitor overlaps with known cistromes. Interestingly, transcriptional regulators involved in the fibrogenic TGFβ (SMAD) and Hippo/YAP1 (TEAD/YAP1) signaling pathways were identified in these analyses (Fig. 5B). Accordingly, DNA motifs used by the SMAD or TEAD TF families are enriched within the BNC2 cistrome (Supplementary Fig. 9B, C). This is in line with previous observations indicating that the TGFβ and Hippo/YAP1 signaling pathways converge in the regulation of myofibroblastic activation8,42 and shows this occurs at BNC2-bound regulatory regions. Indeed, recruitment of SMAD3 and YAP1 (ChIP-seq data from LX2 or IMR90 cells)43,44 is an overall feature of active regulatory elements bound by BNC2 in MFs (Fig. 5C). In line, RIME assays used to define BNC2 chromatin-bound partners in LX2 cells retrieved several cofactors involved in signal transduction from stiff ECM including YAP1 (Fig. 5D, Supplementary Fig. 10, and Supplementary Data Files 6 and 8). A physical interaction between BNC2 and the co-regulator YAP1 was confirmed by co-immunoprecipitation experiments (Fig. 5E and Supplementary Fig. 9D). Identification of genes bound by BNC2 using the Genomic Regions Enrichment of Annotations Tool (GREAT)45 pointed to ECM-related genes such as COL1A1, COL5A1, or Lysyl Oxidase Like 1 (LOXL1) (Fig. 5F and Supplementary Fig. 9E). BNC2 binding to these genes was verified using additional ChIP-qPCR assays (Supplementary Fig. 9F). Interestingly, concomitant recruitment of BNC2, SMAD3, and YAP1 was also observed at the BNC2 gene itself (Fig. 5G). This was consistent with identity TFs being involved in auto-regulatory loops46 and suggested that induction of BNC2 expression upon myofibroblastic activation could be driven by the TGFβ and Hippo/YAP1 signaling pathways. Indeed, induction of Bnc2 expression was recapitulated upon spontaneous in vitro activation of purified primary quiescent murine HSCs (Q-HSCs) into MF-HSCs (Fig. 6A and Supplementary Fig. 11A, B), which involves mechanotransduction through the Hippo/YAP1 pathway due to the high stiffness substrate provided by plastic culture plates8,47. Furthermore, treatment with the YAP1 inhibitor verteporfin48 and transfection of a small interfering RNA (siRNA) targeting Yap1 (siYap1) led to downregulation of Bnc2 expression in in vitro activated primary murine MF-HSCs (Fig. 6B, C). Moreover, growing primary murine Q-HSCs in non-embedded 3D-spheroids, which reduces YAP1 activation47, was also accompanied by lower Bnc2 expression levels when compared to conventional 2D cultures (Fig. 6D). In addition, we found that Bnc2 expression in MF-HSCs was further induced by treatment with TGFβ (Fig. 6E) and inversely reduced by treatment with SB431542 or SD208, which are TGFβ receptor kinase inhibitors (Fig. 6F).

A Input and BNC2 ChIP-seq signals together with CoP-seq and H3K27ac ChIP-seq signals from LX2 cells were visualized at the BNC2 cistrome (1479 binding sites). Heatmaps at the bottom show the signals in 5 kb regions centered on the BNC2 peaks. Average signals are plotted on top of the heatmaps. B The BNC2 cistrome was used to search for proteins with similar genomic binding profiles in CistromeDB and the top 20 hits were ranked according to the GIGGLE similarity score. Proteins involved in TGFβ signaling (SMADs) and Hippo/YAP1 (TEAD and YAP1) were highlighted in red and green, respectively. C Analyses performed as in A to monitor SMAD3 and YAP1 ChIP-seq signals from MFs at the BNC2 cistrome. D Transcriptional regulators specifically identified in BNC2 RIME but not IgG control RIME were clustered according to the percent protein coverage in individual biological replicates (n = 3 biologically independent experiments; anti-BNC2 antibody 55220-1-AP, Proteintech). Percent coverage obtained in an additional BNC2 RIME experiment using the anti-BNC2 antibody HPA018525 (Sigma-Aldrich) is shown on the right. E Nuclear extracts from LX2 cells expressing recombinant BNC2 and YAP1 were subjected to immunoprecipitation with an antibody against BNC2 (55220-1-AP, Proteintech). Immunoprecipitated material was analyzed by western blot using antibodies directed against BNC2 or YAP1. The presented data are representative of two biologically independent experiments. MW, molecular weight markers. F, G The Integrated Genome Browser (IGB) was used to visualize ChIP-seq profiles for BNC2 (black track, LX2 cells), SMAD3 (red track, LX2 cells), YAP1 (green track, IMR90 cells), and H3K27ac (blue track, MF-HSCs) at the COL1A1 (F) and BNC2 (G) genes.

A RT-qPCR data showing changes in gene expression upon murine primary Q-HSCs spontaneous in vitro activation into MF-HSCs (n = 6 biologically independent experiments). Log2 fold changes (FC) between MF-HSCs (6 days of culture) and Q-HSCs (1 day of culture) are shown. B RT-qPCR data showing changes in gene expression upon YAP1 inhibition in MF-HSCs. Isolated mouse primary HSCs were treated with 1 µM verteporfin or vehicle for 6 days before being harvested [n = 3 (Yap1), 4 (Bnc2 and Ctgf), or 5 (Col1a1 and Acta2) biologically independent experiments]. Log2 FC between verteporfin and vehicle-treated cells are shown. C RT-qPCR data showing changes in gene expression upon Yap1 silencing in MF-HSCs (n = 3 biologically independent experiments). Log2 FC between siYAP1 and siCTRL-transfected cells are shown. D RT-qPCR data showing differences in gene expression in mouse primary HSCs grown for 9 days in 3D (spheroids) or in 2D [n = 3 (Bnc2) or 4 (Col1a1, Acta2, Yap1, Ctgf) biologically independent experiments). Log2 FC between cells grown in 3D and 2D are shown. B–D Expression of Ctgf, an established YAP1 target, was assessed. E RT-qPCR data showing changes in gene expression induced by treatment of MF-HSCs with TGFβ (1 ng/mL) for 24 h (n = 4 biologically independent experiments). Log2 FC between cells treated with TGFβ and vehicle are shown. F RT-qPCR data showing changes in gene expression induced by treatment of EMS404 MF-HSCs with the indicated TGFβ signaling inhibitors for 24 h (n = 3 biologically independent experiments). Log2 FC between cells treated with TGFβ signaling inhibitors and vehicle are shown. In all panels, bar graphs show means ± SD. Statistical significance was assessed using two-sided one-sample t test with Benjamini–Hochberg correction for multiple testing to determine if the mean log2 FC was statistically different from 0. *P < 0.05, **P < 0.01, ***P < 0.001. Source data are provided as a Source Data file.
Hence, fibrogenic signaling pathways converge at BNC2 binding sites including those found at the BNC2 gene itself, defining BNC2 as an integrator of fibrogenic stimuli in MF-HSCs.
BNC2 transcriptionally drives the expression of matrisome genes in MF-HSCs
We next more broadly characterized potential BNC2 target genes. In addition to the previously mentioned assignation of BNC2-bound regions to proximal genes using GREAT, we also performed potential target gene assignation taking into account promoter-enhancer interactions predicted by the FDR-corrected OLS with Cross-validation and Shrinkage (FOCS) tool49. These complementary procedures retrieved, as top hits, genes enriched for similar biological processes as revealed by gene ontology term enrichment analyses (Supplementary Fig. 12A). These biological processes were further grouped based on gene-set similarity, which indicated that predicted BNC2 target genes were enriched for functions directly relevant to MF activities such as ”response to wounding” and “ECM” (Fig. 7A, Supplementary Fig. 12B, and Supplementary Data File 9). We next combined these data with transcriptomic analyses of LX2 cells transfected with a siRNA targeting BNC2 (siBNC2; silencing efficiency is shown in Fig. 8A). Gene Set Enrichment Analysis (GSEA)50 indicated that most biological processes predicted to be targeted by BNC2 using its cistrome were indeed enriched for deregulated (i.e., up- or downregulated) genes in BNC2-deficient LX2 cells (Fig. 7A). To further mine BNC2 predicted targets, we monitored enrichment for molecular function gene sets among genes defined as both bound and regulated by BNC2 in these analyses. This notably highlighted an enrichment for genes related to ECM, cell adhesion and signaling such as the TGFβ/SMAD signaling pathway (Fig. 7B, Supplementary Fig. 12C, and Supplementary Data File 9). In line, GSEA indicated that the matrisome gene set was significantly biased towards downregulated genes upon BNC2 silencing (Fig. 7C). Calling differentially regulated genes upon BNC2 silencing and performing gene enrichment analyses also retrieved matrisome genes as specifically enriched among downregulated genes (Supplementary Fig. 13). Similar conclusions were reached when GSEA analyses were performed using a restricted set of genes encoding ECM components effectively detected in human fibrotic livers51 (Fig. 7D). GSEA analyses performed on transcriptomic data obtained upon Bnc2 silencing in the mouse MF-HSC cell line EMS404 (Supplementary Fig. 14) also pointed to downregulation of ECM-related genes (Fig. 7E, F). Both ECM constituents such as collagen encoding genes and ECM regulators including ECM-remodeling enzymes were downregulated in both LX2 and EMS404 cells (Supplementary Fig. S15). The requirement of BNC2 for expression of matrisome-related genes was verified using RT-qPCR in primary human MF-HSCs. Indeed, similar to LX2 cells, BNC2 silencing in primary human MF-HCSs led to decreased expression of a selected set of matrisome genes encoding ECM constituents (COL1A1, additional COL genes and Laminin Subunit Alpha 5 (LAMA5)), the ECM receptor Integrin Subunit Alpha 1 (ITGA1) and the ECM cross-linking enzyme LOXL1 (Fig. 8A). The importance of BNC2 was conserved across species since its silencing led to significant downregulation of most of these ECM-related genes both in EMS404 cells and in mouse primary MF-HSCs (Fig. 8B). In line with reduced expression of Col1a1 and Col1a2, BNC2 silencing decreased protein levels of COL-I in both LX2 and EMS404 cells (Fig. 8C). Of note, downregulation of matrisome gene expression upon BNC2 silencing was also found in LL-29 MF from human idiopathic pulmonary fibrosis (Supplementary Fig. 16) further substantiating the conserved function of BNC2 in MFs. Since Zinc-finger TFs were recently shown to be targeted to cereblon (CRBN)-mediated protein degradation by thalidomide and its derivatives52, we used thalidomide and the pomalidomide, CC-885 or Iberdomide derivatives as additional potential means to control BNC2 expression in MFs. We found that BNC2 levels were specifically downregulated by CC-885 in LX2 cells (Fig. 8D and Supplementary Fig. 17A). This was in line with in silico predictions based on docking analyses performed using a structural model of the BNC2 zinc finger-CRBN interaction, which indicated that only CC-885 was able to adopt a bridging position between BNC2 and CRBN which could promote this interaction (Supplementary Fig. 17B). Thalidomide and its derivatives, including CC-885, were ineffective at decreasing BNC2 in mouse EMS404 cells (Supplementary Fig. 17A), in line with mouse cells being resistant to these compounds at least in part because of differences in the CRBN sequence53. Interestingly, CC-885 treatment of LX2 cells also selectively led to a decrease in matrisome gene expression (Fig. 8E and Supplementary Fig. 17C).

A Biological Process GO term enrichment analysis was performed using genes linked to the BNC2 cistrome. The main recovered terms are indicated under “BNC2 cistrome predicted targets”. Next, microarray-based analysis was used to define how BNC2 predicted target genes were deregulated upon BNC2 silencing. Dot plot depicts the results of GSEA performed for each term using transcriptomic changes induced by BNC2 silencing in LX2 cells, which were transfected with the BNC2-targeting siRNA (siBNC2) or a non-targeting control siRNA (siCTRL) (n = 4 biologically independent experiments). Dot areas are proportional to the normalized encrichment score (NES), while colors indicate the false discovery rate (FDR). A random list of 500 human genes was used as a negative control. B Genes both bound and regulated by BNC2, i.e., genes from the GSEA core enrichments from panel A, were further mined to search for enriched molecular pathways using ToppGene. Enriched terms were clustered according to the similarity of gene lists (Supplementary Fig. 12C), and main clusters pointing to related common molecular functions were further grouped. C, D Enrichment plots from GSEA performed using transcriptomic changes induced by siBNC2 in LX2 cells as the ranking measure and the Matrisome (ECM and ECM-associated genes, n = 1028; C) or human proteomics-based identified fibrotic liver ECM genes (n = 71; D) as the gene sets. NES and FDR are the normalized enrichment score and the false discovery rate provided by GSEA, respectively. E, F Enrichment plots from GSEA similar to those shown in (C, D) obtained using mining of transcriptomic data from EMS404 cells transfected using siBNC2 or siCTRL (control).

A, B RT-qPCR data showing changes in gene expression upon BNC2 silencing in human (A) and mouse (B) MF-HSC cell lines and primary cells. Cells were transfected with BNC2-targeting siRNA (siBNC2) or a non-targeting control siRNA (siCTRL) for 48 h. Data for LX2 are from biologically independent experiments [n = 4 (COL1A1, COL3A1, COL5A1, LOXL1) or 5 (BNC2, COL1A2, COL5A2, LAMA5, ITGA1)] while data for human primary MF-HSCs were obtained using cells from different donors (n = 3). Data for EMS404 are from biologically independent experiments (n = 3 or 2 for Col5a1) while data for mouse primary MF-HSCs were obtained using cells from independent isolations [n = 3 (Col5a1, Col5a2, Lama5, Itga1, Loxl1) or 4 (Bnc2, Col1a1, Col1a2, Col3a1)]. Log2 FC between siBNC2 and siCTRL-transfected cells are shown. C Simple western immunoassays used to monitor changes in BNC2 and type I collagen (COL-I) levels upon BNC2 silencing in LX2 (human) and EMS404 (mouse) cell lines. The lower panel shows the results of protein quantifications obtained from biologically independent experiments [(n = 3 (COL-I in EMS404), 4 (BNC2), or 5 (COL-I in LX2)]. BNC2 and COL-I levels were normalized to that of HSP90 or LMNA expression. Log2 FC between siBNC2 and siCTRL-transfected cells are shown. MW, molecular weight markers. D Simple Western immunoassays used to monitor changes in BNC2 levels in LX2 cells treated with 1 μM pomalidomide (Pom.), thalidomide (Thal.), CC-885, Iberdomide (Iber.) or vehicle for 24 h. Lower panel shows the results of protein quantification obtained from three biologically independent experiments. Data were normalized to LMNA protein expression, and Log2 FC between drug and vehicle-treated cells are shown. MW, molecular weight markers. E RT-qPCR data showing changes in gene expression upon treatment of LX2 cells as described for (D) (n = 3 biologically independent experiments or 2 for iberdomide). Log2 FC between drug and vehicle-treated cells are shown. Pom., pomalidomide; Thal.; thalidomide; Iber., iberdomide. In all panels, bar graphs show means ± SD. Statistical significance was assessed using two-sided one-sample t test with Benjamini–Hochberg correction for multiple testing to determine if the mean log2 FC was statistically different from 0. *P < 0.05, **P < 0.01, ***P < 0.001. Source data are provided as a Source Data file.
Collectively, these data define a role for BNC2 in the control of matrisome gene expression in MFs by integrating pro-fibrotic signals through both its expression and its transcriptional regulatory activities.
BNC2 controls the development of liver fibrosis
In light of our results, we next assessed whether the role for BNC2 in MF-HSCs in vitro translates to a role in fibrosis development in vivo. Ayu21-18 mice, which carry an insertion of a gene trap vector between exons 2a and 3 of the Bnc2 gene leading to its disruption and inactivation54,55 were studied. Since Bnc2 knock-out animals prematurely die within 24 h after birth55, heterozygous (Bnc2+/−) mice were used to define whether reduced Bnc2 expression is sufficient to modulate liver fibrogenesis. The CDAA-HFSC diet was used since it triggers induction of Bnc2 expression (Fig. 3D) concomitant with moderate NASH-associated liver fibrosis. Bnc2+/− mice and their wild-type (WT; Bnc2+/+) littermates were fed the CDAA-HFSC diet for 7.5 weeks (Fig. 9A). Hepatic steatosis (Fig. 9B and Supplementary Fig. 18A) and plasma transaminase activities (Fig. 9C and Supplementary Fig. 18B) were not significantly different between Bnc2+/+ and Bnc2+/− mice. However, a significant decrease in the level of collagen deposition was measured in Bnc2+/− mice, as determined by Sirius red staining of liver sections (Fig. 9D). Bnc2+/− mice were also characterized by reduced liver collagen deposition in an independent experiment where mice were fed the CDAA-HFSC diet for an extended period of 12 weeks (Supplementary Fig. S19A, B). Moreover, liver transcriptomic analyses revealed a significant downregulation of genes relevant to fibrogenesis in Bnc2+/− mice including inflammation and ECM-related functions, as evidenced using GSEA (Fig. 9E and Supplementary Fig. 18C, D). Consistently, GSEA using the matrisome and the human fibrotic liver ECM gene sets indicated a significant bias towards genes downregulated in livers of Bnc2+/− mice, i.e stronger expression in the livers of WT mice (Fig. 9F, G). This was further confirmed by RT-qPCR analyses which indicated that reduced Bnc2 expression in the livers of Bnc2+/− mice was accompanied by decreased expression levels of several of our previously selected set of BNC2-dependent MF matrisome genes (Fig. 9H and Supplementary Fig. 19C).

A Schematic summary of the experimental protocol. Heterozygous mice (+/−, n = 13 mice) and WT mice (+/+, n = 12 mice) were fed for 7.5 weeks with the CDAA-HFSC diet. Control WT mice were fed a chow diet (n = 6 mice). Blood and livers were collected the day of sacrifice for biochemistry, histological, and gene expression analyses shown in panels B–H. A mouse was depicted using “Vector diagram of the laboratory mouse (black and white)“, https://commons.wikimedia.org/wiki/File:Vector_diagram_of_laboratory_mouse_(black_and_white).svg, available under the Creative Commons Attribution-Share Alike 4.0 International license (https://creativecommons.org/licenses/by-sa/4.0/deed.en). B Liver triglycerides (TG) content. C Aspartate aminotransferase (AST) plasma levels. D Quantification of collagen deposition was performed using Sirius red staining of two liver sections per mouse. Ten fields were randomly chosen within each section. The average percentage of fibrotic area is shown. The right panel shows representative images obtained for WT (+/+; n = 11 mice) or heterozygous (+/−; n = 12 mice) mice. E GSEA was used to define biological processes enriched for genes deregulated in the livers of Bnc2 heterozygous mice analyzed in (B–D). Transcriptomic analyses were performed using livers from ten heterozygous (+/−) and ten WT (+/+) mice. Transcriptomic changes induced by Bnc2 deficiency were used as the ranking measure and biological processes (MiSigDB, v7.2) as the gene sets. Normalized enrichment scores (NES) for terms with FDR < 10−4 are shown. F, G Enrichment plots from GSEA performed using transcriptomic changes induced by Bnc2 deficiency in mice as the ranking measure and the Matrisome (ECM and ECM-related genes, n = 1028; F) or human fibrotic liver ECM genes (n = 71; G) as the gene sets. NES and FDR are the normalized enrichment score and the false discovery rate provided by GSEA, respectively. H RT-qPCR data showing the expression of matrisome genes in WT (+/+; n = 11 mice) and heterozygous (+/−; n = 13 mice) mice. B–D, H Graphs show means ± SD. Statistical significance was assessed using one-way ANOVA with Tukey multiple comparison post hoc test (B–D) or one-tailed Mann–Whitney U test with Benjamini–Hochberg correction for multiple testing (H). *P < 0.05, **P < 0.01, ***P < 0.001. Source data are provided as a Source Data file.
Altogether, these data establish that BNC2-mediated control of matrisome genes is also observed in vivo and, consequently, that this TF is required for liver fibrogenesis in mice.
Discussion
By combining multi-omics analyses in experimental models of myofibroblastic activation and organ fibrosis, our study has allowed to uncover BNC2 as a TF critical for MF identity and fibrogenesis. Our strategy, leveraging epigenomic and transcriptomic features of identity TF genes, allowed us to capture poorly studied TFs such as BNC2. This is in contrast with previous studies relying on the enrichment of TF-recognition motifs and regulatory network reconstruction (e.g., refs. 56,57). Indeed, many TFs of the Zinc-finger family including BNC2 have been understudied and are consequently not yet represented in databases of known TF-binding motifs58,59. As an illustration, motif-based identification of TFs cooperating with the cofactor YAP1 in MFs did not identify BNC260. In that regard, and in line with ref. 14, our study highlights that multi-omics approaches can be leveraged to provide an in-depth characterization of a given TF’s functions by defining how this TF bridges signaling pathways and transcriptional regulatory activities. Indeed, we report that BNC2 acts as an integrator of pro-fibrotic stimuli in MFs where the TGFβ and Hippo/YAP1 pathways converge both towards the BNC2 gene itself and BNC2-bound regulatory regions at downstream targets (Fig. 10).

Schematic depicting how BNC2 controls the establishment of the MF transcriptome and development of fibrosis.
Despite originating from various cell types, MFs across different organs acquire a similar phenotype with canonical functions which include ECM synthesis9. Therefore, myofibroblastic activation defines a common theme in fibrogenesis across organs1. Our study indicates that this is linked to MFs being characterized by commonalities in their epigenomic and transcriptomic programs. Importantly, we posit that BNC2 is required at the molecular level to establish MF canonical functionalities and more specifically for high expression of ECM constituents and modifying enzymes. In mice, deletion of Bnc2 is lethal due to the development of a cleft palate and abnormalities of craniofacial bones and the tongue55. In humans, BNC2 mutations have been linked to congenital lower urinary-tract obstruction, adolescent idiopathic scoliosis, and ovarian cancer40,61,62. Therefore, we suspect that some of the molecular functions described in our study may also be relevant to these additional BNC2-related phenotypes. Indeed, the developmental functions of BNC2 are linked to its expression in cells of the mesenchymal lineage in the developing craniofacial bones55. Moreover, reminiscent to several other MF identity TFs (Fig. 2A; e.g., TWIST, SNAI), BNC2 pro-tumorigenic activities40,63 may be linked to its ability to promote mesenchymal-like transcriptional programs triggering epithelial–mesenchymal transitions (EMT)64. However, beyond this unifying program, single-cell transcriptomic analyses are now also pointing to MF adopting a myriad of heterogenous phenotypes. In particular, MFs are emerging as instrumental signaling cells in injured organs where adaptation to the local microenvironment may define labile heterogeneous populations of MFs across different stages of the fibrogenic process65,66. Our data suggest that BNC2 might also be involved in additional MF activities such as those pertaining to cell signaling and dialog (Figs. 7A, B and 9E). In this context, how BNC2 activities and that of MF subtype-specific TFs are orchestrated will be important to define in order to better understand the molecular mechanisms underpinning MF heterogeneity and plasticity. Indeed, cell identity and fine tuning of the cellular transcriptional program are defined through combinatorial activities of multiple TFs. In that regard, we have obtained evidences that additional TFs identified in our study including the other poorly characterized Zinc-finger TF ZFHX4 also represent bona fide regulators of the MF transcriptional program. Indeed, their silencing in MF-HSCs leads to reduced type I collagen encoding gene mRNA and protein levels (Supplementary Fig. 20).
Fibrosis, which can affect virtually all organs, is a major health issue in desperate need for widely applicable therapeutic treatment options2. The ability to reverse fibrosis to avoid life-threatening complications is particularly important but challenging in patients with advanced fibrosis. In this context, targeting myofibroblasts is considered a promising strategy. Considering the high costs of drug development, the interest of targeting myofibroblasts also lies in their conserved role as drivers of fibrosis development across organs1,5,7. Hence, identification of TFs such as BNC2 which drive the common MF identity program and whose expression decreases upon fibrosis regression in humans is of primary interest. While developing drugs targeting TFs outside the nuclear receptor family still represents a tremendous challenge67, our observation that BNC2 expression levels and MF activities are sensitive to the thalidomide derivative CC-885 suggests this TF could be a potential actionable target in fibrosis.
Methods
Resources and reagents
All resources and reagents used in this study are listed in Supplementary Data File 10.
Animal studies
Mice were housed in standard cages in a temperature-controlled room (22-24 °C) with a 12-h dark–light cycle. Wild-type (WT) C57BL/6J mice were purchased from Charles River and were allowed to acclimate for at least 1 week prior to any experiment. The Ayu21-18 mouse line (B6;CBA-Bnc2Gt(pU21)18Imeg/Orl) was purchased from Infrafrontier (EM-05043). In this model, insertion of the gene trap vector pU21 in the intron separating exons 2a and 3 leads to disruption and inactivation of the Bnc2 gene55. For genotyping, DNA was extracted from the tail with REDExtract-N-Amp Tissue PCR Kit (Sigma-Aldrich) and PCR was performed as described in ref. 55 using the primers listed in Supplementary Data File 10. Primers pairs allowing to detect the WT allele or the mutated allele (directed against the pU21 vector) were mixed in the same PCR (35 repetitions of the following amplification cycle were used: 95° for 15 s, 55° for 15 s, and 72° for 30 s). Amplicons were visualized after migration in ethidium bromide-stained agarose gels as shown in Supplementary Fig. 21. Experiments were performed using 13–19 weeks old heterozygous male mice (Bnc2+/−) and their wild-type littermates (Bnc2+/+) as control.
To induce liver fibrosis, WT mice (females, 10–14 weeks old) were intraperitoneally injected with CCl4 (Sigma-Aldrich) three times a week for 8 weeks. Increasing doses of CCl4 were used according to our previously established procedure68 as follows: 0.1 mL/kg (week 1), 0.2 mL/kg (week 2), 0.25 mL/kg (week 3), 0.3 mL/kg (week 4) and 0.4 mL/kg (weeks 5–8). The HFSC diet and its associated NASH-like liver phenotype have been described previously32. The CDAA-HFSC diet consisted of ad libitum feeding with a choline-deficient, l-amino acid-defined (CDAA) diet supplemented with 35% sucrose, 21% fat, and 2% cholesterol. Control mice were fed a chow diet containing 12% sucrose and 7% fat (Ssniff, custom diets). Mice body weight was measured weekly throughout the course of the experiments.
All animal studies were performed in compliance with EU specifications regarding the use of laboratory animals and have been approved by the Nord-Pas de Calais Ethical Committee (APAFIS#15539-2018053011323354).
Human liver samples
Fibrotic liver samples were obtained from NASH patients from the ABOS prospective cohort (“Atlas Biologique de l’Obésité Sévère”; ClinGov NCT01129297)69 and from patients of the TargetOH cohort (“Comparison of Inflammatory Profiles and Regenerative Potential in Alcoholic Liver Disease”; ClinGov NCT03773887; and DC-2008-642)70,71. Patients with obesity enrolled in the ABOS cohort were deemed eligible for weight loss surgery69. For transcriptomic analyses of biopsies from the ABOS cohorts36, 53 NASH patients with a Kleiner score F3-F4 were matched for clinical parameters with 53 control patients with obesity and F0 Kleiner score (age, sex, BMI, alcohol, diabetes, statin, fenofibrate; Supplementary Data File 5) as in ref. 72. Follow-up liver biopsies from patients who underwent bariatric surgery were obtained 1 or 5 years after surgery6. Liver samples with alcohol-related cirrhosis were obtained from 42 patients with decompensated alcoholic-related cirrhosis who underwent liver transplantation at Huriez Hospital’s Liver Unit (Lille, France). Fragments of healthy liver samples were obtained from 20 patients who underwent liver resection for hepatic tumors (control patients; Supplementary Data File 4). The liver samples were immediately fixed for histology or frozen for RNA and protein extraction. Study is authorized by the Lille ethical committee (Lille University Hospital), and informed consent was obtained from all subjects.
Primary HSC isolation, cell culture, transfection, and treatments
Primary mouse HSCs were purified from C57BL/6J mice (male, 15–18 weeks old) according to the protocol described in73. Briefly, livers were digested in situ with 14 mg pronase (Sigma-Aldrich) and 3.7 U collagenase D (Roche) followed by in vitro digestion with 0.5 mg/mL pronase, 0.088 U/mL collagenase D and 0.02 mg/mL DNase I (Roche). HSCs were then separated by a Nycodenz gradient and sorted on a FACS Aria II SORP (BD Biosciences) based on retinoid autofluorescence using the 355 nm laser for excitation and 450/50 nm band-pass filter for detection. Cytometry data were analyzed using FlowJo v10.5.3 (FlowJo, LLC). The purity of HSCs was assessed by measuring the percentage of ultraviolet (UV; retinol autofluorescence) and Desmin-positive cells (Supplementary Fig. 11A).
Primary HSCs and HSC cell lines [human LX2 (Merck, scc064) and murine EMS404 (Kerafast, #EMS404)] were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 5% fetal bovine serum (FCS, Deutscher). Human primary HSC (Supplementary Data file 10) were grown in DMEM supplemented with 15% FCS. LL-29 cell line was grown into F-12 media supplemented with 15% FCS (Deutscher). AML12 and IHH cells were cultured as described in74,75. All cells were grown in the presence of 100 U/mL penicillin-streptomycin (Gibco) at 37 °C in a humidified atmosphere with 5% CO2. As the EMS404 cell line76 had not been thoroughly used in the literature, we verified the proper expression of MF markers and their ability to respond to TGFβ (Supplementary Fig. 14). For TGFβ stimulation, cells were serum-starved for 24 h in DMEM supplemented with 0.5% FCS and 0.2% BSA before incubation with TGFβ (R&D systems) at 1 ng/mL for 24 h in serum-free media. Primary mouse MF-HSCs were subjected to this protocol 4 days after isolation. Additional treatments with 1 μM verterporfin (Sigma-Aldricht), 0.5 μM MZ1 (Tocris) or 1 μM pomalidomide, thalidomide, CC-885 or Iberdomide (MedChemExpress) were directly added to the growth medium. The TGFβ signaling inhibitors SD208 (Tocris) and SB431542 (Stemgent) were used at 1 and 10 µM, respectively. Spheroids were obtained by growing mouse primary HSCs in U-bottom cell repellent plates (Greiner) for 9 days as described in ref. 47. Non-embedded spheroids were used to ensure inactivation of YAP147,77. Reagents used for cell culture treatments are listed in Supplementary Data File 10.
Transfection of siRNA (used at 20 nM) was performed using lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) for EMS404 cells, JetPrime (Polyplus transfection) for LX2 cells and HiPerfect (Qiagen) for primary HSCs, according to the manufacturer’s instructions. Transfected cells were cultured for 48 h prior to terminal assays. For Yap1 silencing in mouse primary HSCs, cells were transfected twice (at day 3 and day 6 after isolation) with a siRNA targeting Yap1 (siYAP1) or non-targeting control siRNA (siCTRL), and cells were harvested 48 h after the last transfection. Control and target siRNAs directed against human or mouse genes were purchased from Thermo Fisher Scientific (Supplementary Data File 10). Plasmid transfection in LX2 cells was performed using JetPEI (Polyplus transfection) according to the manufacturer’s instructions. Transfected cells were cultured for 48 h prior to terminal assays. Plasmids used in these experiments are listed in Supplementary Data File 10.
Isolation of primary mouse liver cell types
Simultaneous purifications of murine primary cell populations from the same livers were performed using a modified version of the method described in ref. 74. Briefly, livers were digested in situ with 100 U/ml of collagenase type IV (Sigma). Hepatocytes (Hep.) were enriched through a 50×g centrifugation prior to size-based FACS sorting, whereas the non-parenchymal fraction was further processed for the isolation of hepatic stellate cells (HSC), Kuppffer cells (KC), cholangiocytes (Chol) and endothelial cells (LSEC). The purity of each cell type was assessed by RT-qPCR for cell-specific markers (Supplementary Fig. 11B).
RNA extraction and RT-qPCR
Tissues were homogenized using a T10 Ultra-Turrax (Ika). The superior right lobe was systematically used for RNA extraction from mouse livers. Total RNA was extracted using Trizol (Eurobio) for cell lines and liver tissues or using the Nucleospin® RNA Plus XS kit (Macherey-Nagel) for primary cells. RNA was reverse-transcribed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystem). RT-qPCR was performed in technical triplicates using the SYBR green Brilliant II fast kit (Agilent Technologies) on an Mx3005p apparatus (Agilent Technologies) or a QuantStudio 3 (Applied Biosystems). The specificity of the amplification was checked by recording the dissociation curves, and the efficiency was verified to be above 95% for each primer pair. mRNA levels were normalized to the expression of housekeeping genes as indicated in Supplementary Data File 10, and the fold induction was calculated using the cycle threshold (ΔΔCT) method. The sequences of primers used are listed in Supplementary Data File 10.
Gene expression microarray
Purified RNA was further digested with rDNase (Macherey-Nagel). RNA integrity and quantity were evaluated using the Agilent 2100 Bioanalyser (Agilent Technologies). RNA was then processed for transcriptomic analysis using Affymetrix GeneChip arrays (HTA 2.0 or MoGene 2.0) according to the manufacturer’s instructions. All liquid handling steps were performed by a GeneChip Fluidics Station 450 and GeneChips were scanned with a GeneChip Scanner 3000-7 G using Command Console v4.1.2 (Affymetrix). Quality controls were performed using the Affymetrix expression console.
Public transcriptomic and functional genomic data recovery
Public data used in this study were downloaded from Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/)78, ENCODE79, Roadmap epigenome (http://www.roadmapepigenomics.org/)80, or CistromeDB (http://cistrome.org/db/#/)41 and are listed in Supplementary Data File 11. RLE normalized expression data (CAGE-seq) from 561 primary cells listed in Supplementary Data File 3 were downloaded from the FANTOM5 website (https://fantom.gsc.riken.jp/5/sstar)81.
Transcriptomic data analyses
RNA-seq raw data were processed using a local instance of Galaxy82. Data were analyzed using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc) and reads were trimmed when necessary. Mapping of reads to the human genome (hg38) was achieved using Tophat 2.0.983 using the Gencode annotation (GRCh38.84). Read counting was then performed using the Htseq-count tool v1.0.084. Normalization taking into account gene lengths and differential expression analyses were performed using EdgeR v0.0.385.
Affymetrix raw data were processed through the GIANT v0.0.2 tools suite86 on a local instance of Galaxy82. Normalization was performed using the apt-probeset-summarize tool of Affymetrix Power Tools (https://www.affymetrix.com/support/developer/powertools/changelog/index.html) using “gc correction+scale intensity+rma”. Normalization level was set to “probeset” and probesets assigned to the same gene (NetAffx Annotation Release 36) were averaged for human data while normalization level was set to “Core genes” for mouse data in order to use the “Collapse to gene symbols” function of the GSEA tool. For differential expression analysis, the LIMMA tool from GIANT v0.0.2 was used87. When necessary, human gene symbols were attributed to each murine gene using the Orthologue Conversion software (https://biodbnet-abcc.ncifcrf.gov/db/dbOrtho.php).
Pathway and gene-set enrichment analyses
Enrichments for biological or molecular pathways were defined using Metascape (http://metascape.org)88 or the ToppGene Suite89.
Gene-set enrichment analyses (GSEA) were performed using the GSEA software (v3.0) developed at the Broad Institute50 essentially as described previously25. We used 1000 gene-set permutations and the following settings: “weighted” as the enrichment statistic and “difference of classes” or “signal to noise” as the metric for ranking genes. Ranking was performed by the GSEA software using the gene average expression when multiple probesets were present in the microarray. The following gene sets from MSigDB (https://www.gsea-msigdb.org/gsea/msigdb) were used: Biological process v7.2, Reactome v.7.2, and KEGG v7.2. The matrisome gene set comprising 1028 genes encoding for ECM and ECM-associated proteins (or selective subsets including collagen or ECM-regulator encoding genes) was retrieved from http://matrisomeproject.mit.edu/12. The human fibrotic liver ECM gene set comprising genes encoding ECM proteins that have been detected by proteomics analyses in fibrotic livers (71 genes) were retrieved from http://matrisomedb.pepchem.org/51. In addition to enrichment plots, figures also provide NES and FDR, which are the normalized enrichment score and the false discovery rate provided by GSEA, respectively. The GSEA core enrichment comprising genes that account for the gene set’s enrichment signal was retrieved in some of the performed analyses. Random gene lists used in GSEA were obtained using the Random Gene Set Generator tool (https://molbiotools.com/randomgenesetgenerator.php). Similar terms were grouped using an in-house procedure in R90 or using the GeneSetCluster package91.
Gene-module association determination (G-MAD)
Prediction of BNC2 functions was obtained using G-MAD analyses (https://www.systems-genetics.org/gmad)28, which were performed using default parameters and “Homo sapiens” datasets. Additional runs were performed to monitor the G-MAD score obtained for “GO:0044420 extracellular matrix component” in individual organs.
Single-cell and single-nuclei RNA-seq analyses
Single-cell RNA-seq data of HSCs from control and CCl4-treated mice have been described in31. Briefly, dissociated livers from CCl4 or vehicle-treated mice were enriched by flow cytometry for LSECs, KCs/MDMs (Kupffer cells/monocyte-derived macrophages), and HSCs prior to 10x Genomics Single-Cell 3’ v2 library preparation and sequencing. Sequencing data were aligned and quantified using the Cell Ranger Single Cell Software Suite (ver. 2.1), and cell types were separated using the Scanpy implementation of the Louvain algorithm. UMAPs were generated using Seurat v. 4.0.3.
Single-nuclei RNA-seq data of liver cells obtained from mice fed a HFSC NASH diet were retrieved from https://www.livercellatlas.org/download.php34. Only annotated cells were used, and cell populations with less than 50 individual cells were removed. Counts were then normalized using SCTransform from the Seurat package v. 4.0.392. UMAP were generated using Seurat.
Protein extraction
Total cell extracts were obtained by washing cells with ice-cold Phosphate-Buffered Saline (PBS) and scraping in Pierce IP lysis buffer (Thermo Fisher Scientific) containing a protease inhibitor cocktail (PIC, Roche). Cell lysates were then sonicated for 5 min (five cycles of 30 s ON/30 s OFF) using Bioruptor (Diagenode), and insoluble material was removed by 10 min centrifugation at 13,000×g.
The preparation of protein from the chromatin fraction was performed according to the protocol described in ref. 25. The cytosolic fraction issued from the same samples was also saved and analyzed.
Protein immunoblotting
Protein concentration was determined using the PierceTM BCA protein assay kit (Thermo scientific), and samples were used for immunoblotting. Regular western blotting or Simple Western immunoassays using the Wes system (Protein Simple), performed as in ref. 25, are described hereafter. Antibodies used and their dilutions are listed in Supplementary Data File 10. Uncropped and unprocessed images are provided in the Source Data file.
Western blotting
Ïn total, 40 µg of proteins were separated by 10% SDS-PAGE and immunodetected using the primary antibodies listed in Supplementary Data File 10. Detection was achieved using HRP-conjugated secondary antibodies (Sigma-Aldrich). Images were acquired using the iBrightTM CL1500 Imaging System (Thermo Fisher Scientific). Quantifications were performed using Image Studio Litev5.2 (LI-COR Biosciences, Lincoln, USA).
Simple western immunoassays
Protein extracts (0.4 µg/mL) were run on a Wes system (Protein Simple) according to the manufacturer’s instructions. Separation was performed using the 12–230 kDa capillary cartridges. The chemiluminescence-based electrophoretogram was autogenerated, and digitally rendered bands were produced from the chemiluminescent peaks using the Compass software (Protein Simple). Quantifications were obtained using the area under the peak of the protein of interest. Since antibodies against BNC2 had been poorly characterized in the literature, we performed experiments to demonstrate the specificity of #HPA059419 (Sigma) for western blotting (Supplementary Fig. 7).
Co-immunoprecipitation assays
Co-immunoprecipitation assays were conducted as in25. LX2 cells were transfected for 48 h with a plasmid coding for human BNC2 (custom construction, E-Zyvec) and a plasmid coding for constitutively active nuclear YAP1 (YAP1-S127A, Addgene). Alternatively, endogenous proteins were cross-linked by incubating LX2 cells with 1% formaldehyde for 10 min at room temperature. For nuclear extract preparation, cells were rinsed with ice-cold PBS and lysed with hypotonic buffer (20 mM Tris-HCl, pH 8.0, 10 mM NaCl, 3 mM MgCl2, 0.2% NP40) for 5 min at 4 °C. After 5 min centrifugation at 600×g, the pellet was resuspended in lysis buffer (25 mM Tris-HCl pH 8, 1 mM EDTA, 1.5 mM MgCl2) and incubated for 30 min at 4 °C. Following 10 min (30 s ON/30 s OFF) sonication with the Bioruptor, insoluble material was removed through 10 min centrifugation at 13000 g. Immunoprecipitation was performed using 2 µg of BNC2 antibody (55220-1-AP from Proteintech or HPA018525 from Sigma-Aldrich) or a control IgG and 500 µg of nuclear extract, which were incubated overnight at 4 °C in dilution buffer (25 mM Tris-HCl pH 8, 1 mM EDTA, 1.5 mM MgCl2). 10 µl of a 1:1 mixture of protein A and protein G magnetic beads (Dynabeads, Thermo Fisher Scientific) blocked overnight in PBS containing 5 mg/mL Bovine serum albumin (BSA) was then added to each sample. After incubating samples 4 h at 4 °C under agitation, beads were washed four times with cold washing buffer (25 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1 mM EDTA, 0.2% NP40). Elution was performed in Laemmli buffer 6× (175 mM Tris-HCl pH 6.8, 15% glycerol, 5% SDS, 300 mM DTT and 0.01% Bromophenol Blue). All buffers were supplemented with PIC (Roche). Input proteins and IPed materials were analyzed by western immunoblotting.
Chromatin immunoprecipation (ChIP) and ChIP-seq
Chromatin immunoprecipitation was performed as described previously25. LX2 cells were fixed for 10 min at room temperature with 1% formaldehyde (Thermo Fisher Scientific) followed by 5 min incubation with 125 mM glycine. After two washes with ice-cold PBS, cell pellets were incubated for 10 min in 0,25% Triton X-100, 10 mM EDTA, 10 mM HEPES and 0.5 mM EGTA followed by 10 min incubation in 0.2 M NaCl, 1 mM EDTA, 10 mM HEPES, 0.5 mM EGTA. Nuclei were then resuspended in Lysis buffer (50 mM Tris-HCl pH 8.0, 10 mM EDTA, 1% SDS) and sonicated for 30 min (three cycles 30 s ON/30 s OFF) using a Bioruptor (Diagenode). All buffers were supplemented with PIC (Roche). Chromatin was diluted tenfold in Dilution Buffer (20 mM Tris-HCl pH 8.0, 1% Triton X-100, 2 mM EDTA, 150 mM NaCl) and incubated overnight at 4 °C with 2 μg of H3K27ac antibody (Active Motif, #39685) or 3 μg of BNC2 antibody (Sigma, HPA018525 or Sigma, HPA059419). The next day, protein A/G sepharose beads (GE Healthcare) were added, and samples were incubated for 4 h at 4 °C under agitation in the presence of 70 μg/mL yeast tRNA (Sigma-Aldrich). Beads were washed three times with RIPA buffer (50 mM HEPES, 1 mM EDTA, 0.7% Na Deoxycholate, 1% NP40 and 500 mM LiCl) containing 10 μg/mL yeast tRNA and once with TE buffer (10 mM Tris-HCl pH 8.0, 1 mM EDTA). DNA was then eluted in 100 mM NaHCO3 containing 1% SDS and incubated overnight at 65 °C for reverse cross-linking. DNA purification was performed using the MinElute PCR purification kit (Qiagen). ChIP and input samples were subjected to high-throughput sequencing and were further analyzed as described hereafter. Selected binding sites were confirmed using qPCR and primers listed in Supplementary Data File 10.
CoP-seq
The CoP (Column Purified chromatin)-seq procedure was performed essentially as described in ref. 39. In brief, LX2 cells were cross-linked, lyzed and sonicated as described for ChIP except 2% formaldehyde were used. Soluble chromatin was loaded onto MinElute PCR purification kit (Qiagen) and eluates were treated with RNAse A, proteinase K and reverse cross-linked before final DNA purification with the MinElute PCR purification kit. Inputs were processed similarly except for the column-based purification of cross-linked material which was omitted.
ChIP-seq and CoP-seq data analysis
ChIP-seq and CoP-seq raw data were processed using a local instance of Galaxy82 essentially as described previously93. Data processing involved FastQC analysis (http://www.bioinformatics.babraham.ac.uk/projects/fastqc) and read mapping to hg38 using Bowtie2 version 1.0.094. Genome-wide signal tracks and enriched regions (peak calling) were obtained using model-based analysis of ChIP-seq version 2 (MACS2 v2.1.1.20160309)95. Input DNA was used as control, duplicate tags and those mapping to ENCODE blacklisted regions v296 were removed and parameters used for peak calling were as described in ref. 93. Signal tracks corresponding to enrichment over input were obtained using MACS2 bdgcmp. For H3K4me3 ChIP-seq processing, peak calling was performed using the broad option for histone marks (q < 0.001). Only H3K4me3 domains which could be assigned to a gene were considered in subsequent analyses, i.e., H3K4me3 domains with predicted gene TSS (GENCODE v24 database) overlapping or within 1 kb. ChIP-seq signals and called peaks were visualized at specific loci using the Integrated Genome Browser (IGB v9.0.1)97 or were analyzed at large scales using the deepTools v3.3.298. The promoters of housekeeping genes defined in Eisenberg and Levanon, 2013 were used as a control in some of these analyses. The coordinates of their TSS were retrieved using the UCSC Genome Browser99 and RefSeq as the gene annotation.
The BNC2 cistrome was compared with that of publicly available cistromes contained within the CistromeDB database using the CistromeDB toolkit41. « All peaks in each sample » was used for these analyses. De novo motif enrichment analyses were performed using the RSAT peak-motifs tool run with default parameters (http://rsat.sb-roscoff.fr/peak-motifs_form.cgi)100. BNC2 target genes were predicted by assigning peaks to genes using GREAT4.0.445 or FOCS49. With regards to the latest, BNC2 peaks within 2.5 kb of a Gencode v34 TSS101 were defined as promoter binding sites. The remaining peaks were monitored for predicted promoter interaction within the entire FOCS datasets (http://acgt.cs.tau.ac.il/focs/download.html; enhancers without assigned genes were removed). All retrieved genes were merged defining FOCS predicted BNC2 target genes.
Super-enhancers were defined as in our previous study102. First, model-based analysis of ChIP-seq version 2 (MACS2)95 was used to call peaks in the H3K27ac ChIP-seq data (FDR < 0.15) using input as control. Peaks matching to ENCODE blacklisted regions96 were discarded. Super-enhancers were then called using retained MACS2-derived peaks and Rank Ordering of Super-Enhancers (stitching set at 12.5 kb)24,103 leaving out regulatory regions mapping to promoters (TSS+/−2.5 kb) issued from GREAT45 and using Input DNA as control.
Bioinformatical identification of MF identity TFs
The cumulative frequency distribution of H3K4me3 domain lengths was used to define the inflexion point of the curve (3017 bp), which served to separate sharp from broad H3K4me3 domains (Fig. 1A)104. These broad H3K4me3 domains were assigned to genes using GREAT 2.045 (retrieving 1278 genes) in order to only use extremely high-confidence gene predictions as described in26. TSSs were used to monitor the distance to the center of the nearest super-enhancer, which were defined using primary MF-HSC H3K27ac ChIP-seq data18 following a procedure detailed hereabove. Note that we favored the use of broad H3K4me3 over that of super-enhancers to define identity genes to overcome issues highlighted in our previous study including uncertainty about target gene assignment to distal super-enhancers102.
Next, the length of H3K4me3 domains associated with the 1278 genes in 76 normal human cell types and tissues defined by26 was retrieved (Supplementary Data File 2). For each gene, the relative H3K4me3 domain length in a given cell type or tissue was obtained by dividing by the mean length of domains associated to this gene in all samples. A heatmap of relative H3K4me3 lengths was generated using the heatmap.2 function of the R package “gplots” (https://cran.r-project.org/web/packages/gplots/) and hierarchical clustering was performed using the hclust function of the R package “Stats” using ward.D2 agglomeration method105. This allowed to define three main clusters of genes listed in Supplementary Data File 1. TFs within these clusters were subsequently identified by comparison with the human TFs list provided by the Animal TFDB 2.0 database106.
Further analysis of TFs from cluster 1 was performed by mining the scientific literature to identify articles linking these TFs to MF or fibrosis using the easyPubMed package (https://www.data-pulse.com/dev_site/easypubmed/) in R. Articles with co-occurrence of a given TF gene name and “fibroblast”, “myofibroblast”, “fibrosis” or “extracellular matrix” in their title or abstract were retrieved. Results were visualized on a network generated using Cytoscape107. First, the TF network was generated using String 11.0 (https://string-db.org/; “full network”, “Medium confidence”)108. The network was then exported into Cytoscape where TFs without interactions were manually added back and where nodes were colored according to the number of previously retrieved PubMed articles.
Mouse liver histology and biochemical analyses
Histology
Liver slices were fixed with 4% paraformaldehyde for 48 h and embedded in paraffin using a STP 120 Spin Tissue Processor (Microm Microtech). Paraffin-embedded samples were cut at a thickness of 5-μm and sections were transferred on gelatin-coated slides. Fibrosis assessment was carried out by staining liver sections with a 0.1% solution of Sirius red in 1.3% saturated aqueous picric acid solution. To uncertain reliability of Sirius red staining quantifications, we followed guidelines provided in ref. 109. To avoid potential interlobular differences, we systematically used the same lobe (i.e., the right median lobe). Two large sections, including tissue margin and center but excluding the peripheral tissue and vessels, were used per mouse liver. The entire lobe sections were scanned using an Axioscan (Zeiss), and ten fields were randomly chosen (7.5-week experiment) or the entire section (12-week experiment) was used for quantification with Image J version 1.53c software (NIH, https://imagej.nih.gov/ij/). The analysis was systematically done on two different liver sections and all histological analyses were performed blinded.
Metabolic parameters
Before sacrifice, blood samples were collected from the retro-orbital sinus of mice. Plasma alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were measured using colorimetric assays (Thermo Fisher) on a Konelab 20 (Thermo Fisher).
Measurement of liver triglycerides
A weighted piece of liver was homogenized with T10 Ultra-Turrax (Ika) in PBS. Samples were transferred into glass tubes and mixed with a 2:1 chlorofrom:methanol mixture. After centrifugation, upper- and inter-phase were discarded. The lower organic phase was evaporated under nitrogen flow and reconstituted in 1% Triton X-100. Triglyceride content was measured with the LiquiColor Triglycerides Test (Interchim). Protein concentration was measured in parallel using the PierceTM BCA protein assay kit (Thermo scientific).
Rapid immunoprecipitation mass spectrometry of endogenous protein (RIME)
LX2 cells were processed using the protocol described in ref. 38. Briefly, 5 × 106 cells were grown for 2 days before being fixed in 1% formaldehyde (Sigma-Aldrich). Nuclear proteins were extracted in 10 mM Tris-HCl pH 8.0, 100 mM NaCl, 1 mM EDTA, 0,5 mM EGTA, 0,1% Na deoxycholate, 0.5% N-lauroylsarcosine and PIC (Roche). Immunoprecipitation was achieved using Dynabeads (Invitrogen) conjugated with 10 µg of BNC2 antibody (#55220-1-AP, Proteintech or #HPA018525, Sigma-Aldrich) or 10 μg of a non-immune control IgG (#2729, Cell Signaling). Beads were rinsed with RIPA buffer (50 mM HEPES, 1 mM EDTA, 0.7% Na Deoxycholate, 1% NP40 and 500 mM LiCl) and AMBIC solution (Sigma-Aldrich) before trypsin digestion.
Samples were analyzed by coupling a nanoflow liquid chromatography system (nanoElute, Bruker Daltonics) online to a trapped ion mobility spectrometry-quadrupole time of flight mass spectrometer (timsTOF Pro, Bruker Daltonics) equipped with a CaptiveSpray source operating in positive mode. Peptides were loaded on a trapping column (Acclaim PepMap 100, C18, 100 Å, 100 µm × 20 mm, Thermo Scientific) and separated on a reversed-phase C18 column (Aurora, 25 cm × 75 μm i.d., 1.6 μm, IonOpticks). Chromatographic separation was carried out using a gradient of 2-25% of solvent B (0.1% formic acid in acetonitrile) over 90 min, then 37% over 100 min and 95% over 110 min at a constant flow rate of 300 nl/min. The column temperature was controlled a 50 °C. Mass Spectrometry (MS) data were collected over a m/z range of 100–1700, and MS/MS range of 100–1700. LC-MS/MS data were acquired using the PASEF method with a total cycle time of 1.88 s, including 1 TIMS MS scan and 10 PASEF MS/MS scans. Ion mobility coefficient (1/K0) value was set from 0.7 to 1.25 Vs cm−2. Singly charged precursors were excluded by their position in the m/z-ion mobility plane and precursors that reached a “target value” of 17,000 a.u. were dynamically excluded for 0.2 min. The quadrupole isolation width was set between 2 and 3 m/z depending on precursor m/z. The collision energies varied between 20 and 52 eV depending on precursor mass and charge. TIMS, MS operation and PASEF were controlled and synchronized using the control instrument software OtofControl 6.0 (Bruker Daltonik).
Raw MS files were converted into mascot generic files (mgf) and subjected to a Mascot (2.6.2) (Mascot, Matrix Science) search using a target-decoy strategy in order to evaluate the false discovery rate (FDR) of the search. The human database was created with human proteins and present in SwisProt database (created 2019-09-12, containing 20490 target sequences plus the same number of reversed decoy sequences). Search parameters were as follows: trypsin was set as the cleavage enzyme and a maximum of one miscleavage was allowed. Cysteine carbamidomethylation was set as a fixed modification, whereas methionine oxidation, was set as a variable modification. The peptide mass tolerance (tolerance of mass measurement for precursor ion) was set to 15 ppm and the MS/MS mass tolerance (tolerance of mass measurement for fragment ion) set to 0.05 Da. Proline pipeline (http://www.profiproteomics.fr/proline/) was used to validate the identification results. This statistical validation was performed using the Target Decoy approach, which consists in creating a “decoy” sequence that does not exist in nature. The false identifications thus distribute evenly between the real database (target) and the decoy one, and so the number of decoy hits can be used to estimate the false discovery rate (FDR = decoy hits/decoy hits + target hits) and eliminate false positives. Peptide and protein identification validation parameters were set as follows: minimal length of seven amino acid, score ≥20, pretty rank ≤1, and FDR ≤ 1.
Only proteins detected in the BNC2 RIME, at least two out of three biological replicates, but in none of the non-immune control IgG RIME experiments were considered. Common non-specifically detected proteins in affinity purification mass spectrometry data from the Contaminant Repository for Affinity Purification (human CRAPome2.0) database110 were also discarded. BNC2 interactors were further filtered using the list of human transcriptional regulators from the AnimalTFDB3.0 database111. BNC2 interactors were clustered based on percent protein coverage in RIME assays using the heatmap.2 function of the R package “gplots” (https://cran.r-project.org/web/packages/gplots/) as described hereabove for H3K4me3 domain lengths. Biological triplicates were obtained using the anti-BNC2 antibody 55220-1-AP (Proteintech) and an additional experiment was performed using the anti-BNC2 antibody HPA018525 (Sigma-Aldrich). All non-filtered detected proteins for individual samples are provided in Supplementary Data Files 6 and 8.
RNA in situ hybridization
RNAscope assays on paraffin-embedded liver sections were performed using the RNAscope® Multiplex Fluorescent Reagent Kit v2 according to the manufacturer’s instructions (Bio-Techne). Briefly, the paraffin-embedded tissue sections were deparaffinized and pre-treated with hydrogen peroxide. Sections were incubated in RNAscope® 1× Target Retrieval Reagent for 15 min at 99 °C and treated with RNAscope® Protease Plus for 30 min at 40 °C in the HybEZ oven sequentially. Samples were then incubated with RNAscope® Probe-Hs-BNC2-C1 (#496801) and Probe-Hs_COL1A1-C2 (#401891-C2) for 2 h at 40 °C. After three steps of amplification, RNAscope® HRP-C1 was added on the sections that were incubated with Opal 650 (Akoya Biosciences) for 30 min. HRP-C2 was then added followed by incubation with Opal 570. When specified, RNA in situ hybridization was followed by immunofluorescence staining using an anti-ACTA2 antibody (ab76126, Abcam:1:50). Alcohol-related liver cirrhosis samples were analyzed using a CSU-W1 Spinning Disk (Gataca) with a 60x CFI PLAN APO LBDA objective (resolution 0.110 µM) and images were analyzed using ImageJ1.53c (https://imagej.nih.gov/ij/). For liver biopsies from NASH patients before and after bariatric surgery, the entire liver sections were scanned using the Axioscan (Zeiss) and images were analyzed using the ZEN 2 software (Zeiss). Quantification of red and white dots; illustrating the number of COL1A1 and BNC2 transcripts, respectively, was performed using ImageJ1.53c (https://imagej.nih.gov/ij/). The number of cells in each section was obtained by counting DAPI-positive nuclei.
Structural model of the BNC2-CRBN interaction and docking analyses
A structural model of the interaction between BNC2 zinc-finger 1 (BNC2ZF1) and CRBN was built using the crystallographic structure of DDB1-CRBN-pomalidomide bound to IKZF1 (Protein Data Bank ID 6h0f)52 and used for docking of thalidomide and its derivatives. First, we chose an experimentally resolved zinc finger-CRBN complex on the basis of its resolution and co-crystallized ligand, settling for DDB1-CRBN-pomalidomide bound to IKZF1 (Protein Data Bank ID 6h0f)52. Out of the six BNC2 zinc fingers, four were below a 25% identity threshold with the co-crystallized IKZF1 zinc finger. The two remaining BNC2 zinc fingers were the first (29% identity, 38.7% similarity) and fifth (25.8% identity and 38.7% similarity). We opted to keep the first (BNC2ZF1), as it was closer to the reference. BNC2ZF1 was modeled on the RMN structure of human zinc fingers and homeoboxes 1 (Protein Data Bank ID 2ghf; 20.83% identity and 50% similarity with BNC2ZF1). Interestingly, even with a higher identity percentage than the template used to build BNC2ZNF1, the model resulting from a homology with IKZF1 was marginally less interesting, with a slightly higher overall deviation from the Ramachandran optimal parameters. The BNCZF1-CRBN complex was next built by superimposing the BNC2 and IKZF1 zinc fingers and exchanging the co-crystallized IKZF1 for BNCZF1 and adjusting the side chains by a quick geometry optimization in two steps after adding hydrogens and partial charges. First, the side chains were adjusted by 1000 steps of steepest descent, then the whole zinc finger and the side chains of the DDB1-CRBN-pomalidomide complex were subjected to another 1000 steps of steepest descent. This protocol avoided a deformation of the skeletons of the proteins while minimizing the clashes between side chains. The resulting complex was used as the target for a docking study with GOLD (Genetic Optimisation for Ligand Docking)112, with the binding site defined as 10 Å sphere around the co-crystallized pomalidomide. A set of ligands was docked: pomalidomide as a control, the shorter thalidomide, iberdomide which was similar in size to CC-885, and CC-885. In all, 30 poses were generated for each compound. The docking was assessed on the basis of the number of clusters of closely related poses, the score of the clusters, and their spread around their average pose. Apart from CC-885, all the compounds were built in the same configuration as the co-crystallized pomalidomide (S enantiomer).
Statistical analyses
Statistical analyses were performed using the Prism software (GraphPad) and R105. The specific tests and corrections for multiple testing that were used are indicated in the figure legends. Unless specified in the figure legends, statistical significance was displayed as follow *P < 0.05, **P < 0.01, and ***P < 0.001.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Transcriptomic and cistromic data generated in this study have been deposited into Gene Expression Omnibus under SuperSeries accession number GSE185529. The mass spectrometry proteomics data generated in this study have been deposited to the ProteomeXchange Consortium via the PRIDE [1] partner repository with the dataset identifier PXD003624. Datasets analyzed in this study (Supplementary Data File 11) are available through the Gene Expression Omnibus (GSE6810818, GSE58680113, GSE3810343, GSE6185244, GSE6810818, GSE111059114, GSE63626115, GSE14508631, GSE19274234), ENCODE (https://www.encodeproject.org/experiments/ENCSR507UDH/)79, FANTOM5 (https://fantom.gsc.riken.jp/5/sstar)81, the matrisome database (http://matrisomeproject.mit.edu/ and http://matrisomedb.pepchem.org/)51, and manuscript supplementary data (https://www.nature.com/articles/ng.3385 and https://www.atsjournals.org/doi/10.1164/rccm.201712-2410OC)26,116. Source data are provided with this paper.
References
Rockey, D. C., Bell, P. D. & Hill, J. A. Fibrosis—a common pathway to organ injury and failure. N. Engl. J. Med. 372, 1138–1149 (2015).
Henderson, N. C., Rieder, F. & Wynn, T. A. Fibrosis: from mechanisms to medicines. Nature 587, 555–566 (2020).
Haas, J. T., Francque, S. & Staels, B. Pathophysiology and mechanisms of nonalcoholic fatty liver disease. Annu. Rev. Physiol. 78, 181–205 (2016).
Leslie, M. The liver’s weighty problem. Science 349, 18–20 (2015).
Distler, J. H. W. et al. Shared and distinct mechanisms of fibrosis. Nat. Rev. Rheumatol. 15, 705–730 (2019).
Lassailly, G. et al. Bariatric surgery provides long-term resolution of nonalcoholic steatohepatitis and regression of fibrosis. Gastroenterology 159, 1290–1301.e5 (2020).
Mehal, W. Z., Iredale, J. & Friedman, S. L. Scraping fibrosis: expressway to the core of fibrosis. Nat. Med. 17, 552–553 (2011).
Pakshir, P. et al. The myofibroblast at a glance. J. Cell Sci. 133, jcs227900 (2020).
Plikus, M. V. et al. Fibroblasts: origins, definitions, and functions in health and disease. Cell 184, 3852–3872 (2021).
Herrera, J., Henke, C. A. & Bitterman, P. B. Extracellular matrix as a driver of progressive fibrosis. J. Clin. Investig. 128, 45–53 (2018).
Santos, A. & Lagares, D. Matrix stiffness: the conductor of organ fibrosis. Curr. Rheumatol. Rep. 20, 2 (2018).
Naba, A. et al. The matrisome: in silico definition and in vivo characterization by proteomics of normal and tumor extracellular matrices. Mol. Cell Proteom. 11, M111.014647 (2012).
Ortiz, C. et al. Extracellular matrix remodeling in chronic liver disease. Curr. Tissue Microenviron. Rep. 1–12, https://doi.org/10.1007/s43152-021-00030-3 (2021).
Weidemüller, P., Kholmatov, M., Petsalaki, E. & Zaugg, J. B. Transcription factors: bridge between cell signaling and gene regulation. PROTEOMICS n/a, e2000034 (2021).
Beacon, T. H. et al. The dynamic broad epigenetic (H3K4me3, H3K27ac) domain as a mark of essential genes. Clin. Epigenetics. 13, 138 (2021).
Xia, B. et al. Machine learning uncovers cell identity regulator by histone code. Nat. Commun. 11, 2696 (2020).
Pekowska, A., Benoukraf, T., Ferrier, P. & Spicuglia, S. A unique H3K4me2 profile marks tissue-specific gene regulation. Genome Res. 20, 1493–1502 (2010).
Zhou, C. et al. Long noncoding RNAs expressed in human hepatic stellate cells form networks with extracellular matrix proteins. Genome Med. 8, 31 (2016).
Iwaisako, K. et al. Origin of myofibroblasts in the fibrotic liver in mice. Proc. Natl Acad. Sci. USA 111, E3297–E3305 (2014).
Mederacke, I. et al. Fate tracing reveals hepatic stellate cells as dominant contributors to liver fibrosis independent of its aetiology. Nat. Commun. 4, 2823 (2013).
Buchholz, M. et al. Transcriptome analysis of human hepatic and pancreatic stellate cells: organ-specific variations of a common transcriptional phenotype. J. Mol. Med. 83, 795–805 (2005).
De Minicis, S. et al. Gene expression profiles during hepatic stellate cell activation in culture and in vivo. Gastroenterology 132, 1937–1946 (2007).
Marcher, A.-B. et al. Transcriptional regulation of hepatic stellate cell activation in NASH. Sci. Rep. 9, 2324 (2019).
Hnisz, D. et al. Super-enhancers in the control of cell identity and disease. Cell 155, 934–947 (2013).
Dubois, V. et al. Endoplasmic reticulum stress actively suppresses hepatic molecular identity in damaged liver. Mol. Syst. Biol. 16, e9156 (2020).
Chen, K. et al. Broad H3K4me3 is associated with increased transcription elongation and enhancer activity at tumor-suppressor genes. Nat. Genet. 47, 1149–1157 (2015).
Denu, R. A. et al. Fibroblasts and mesenchymal stromal/stem cells are phenotypically indistinguishable. AHA 136, 85–97 (2016).
Li, H. et al. Identifying gene function and module connections by the integration of multispecies expression compendia. Genome Res. 29, 2034–2045 (2019).
Horie, M. et al. TBX4 is involved in the super-enhancer-driven transcriptional programs underlying features specific to lung fibroblasts. Am. J. Physiol. Lung Cell Mol. Physiol. 314, L177–L191 (2018).
Lee, J.-E. et al. Brd4 binds to active enhancers to control cell identity gene induction in adipogenesis and myogenesis. Nat. Commun. 8, 2217 (2017).
Terkelsen, M. K. et al. Transcriptional dynamics of hepatic sinusoid-associated cells after liver injury. Hepatology https://doi.org/10.1002/hep.31215 (2020).
Haas, J. T. et al. Transcriptional network analysis implicates altered hepatic immune function in NASH development and resolution. Nat. Metab. 1, 604–614 (2019).
L’homme, L. et al. Deletion of the nuclear receptor RORα in macrophages does not modify the development of obesity, insulin resistance and NASH. Sci. Rep. 10, 21095 (2020).
Guilliams, M. et al. Spatial proteogenomics reveals distinct and evolutionarily conserved hepatic macrophage niches. Cell 185, 379–396.e38 (2022).
Bou Saleh, M. et al. Loss of hepatocyte identity following aberrant YAP activation: a key mechanism in alcoholic hepatitis. J. Hepatol. S0168827821004165, https://doi.org/10.1016/j.jhep.2021.05.041 (2021).
Margerie, D. et al. Hepatic transcriptomic signatures of statin treatment are associated with impaired glucose homeostasis in severely obese patients. BMC Med. Genomics 12, 80 (2019).
Kisseleva, T. & Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 18, 151–166 (2021).
Mohammed, H. et al. Rapid immunoprecipitation mass spectrometry of endogenous proteins (RIME) for analysis of chromatin complexes. Nat. Protoc. 11, 316–326 (2016).
Zhang, Y. et al. HiCoP, a simple and robust method for detecting interactions of regulatory regions. Epigenetics Chromatin 13, 27 (2020).
Buckley, M. A. et al. Functional analysis and fine mapping of the 9p22.2 ovarian cancer susceptibility locus. Cancer Res. 79, 467–481 (2019).
Zheng, R. et al. Cistrome Data Browser: expanded datasets and new tools for gene regulatory analysis. Nucleic Acids Res. 47, D729–D735 (2019).
Piersma, B., Bank, R. A. & Boersema, M. Signaling in fibrosis: TGF-β, WNT, and YAP/TAZ Converge. Front. Med. 2, 59 (2015).
Ding, N. et al. A vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic response. Cell 153, 601–613 (2013).
Stein, C. et al. YAP1 exerts its transcriptional control via TEAD-mediated activation of enhancers. PLoS Genet. 11, e1005465 (2015).
McLean, C. Y. et al. GREAT improves functional interpretation of cis-regulatory regions. Nat. Biotechnol. 28, 495–501 (2010).
Arendt, D. et al. The origin and evolution of cell types. Nat. Rev. Genet. 17, 744–757 (2016).
Mannaerts, I. et al. The Hippo pathway effector YAP controls mouse hepatic stellate cell activation. J. Hepatol. 63, 679–688 (2015).
Wang, C. et al. Verteporfin inhibits YAP function through up-regulating 14-3-3σ sequestering YAP in the cytoplasm. Am. J. Cancer Res. 6, 27–37 (2016).
Hait, T. A., Amar, D., Shamir, R. & Elkon, R. FOCS: a novel method for analyzing enhancer and gene activity patterns infers an extensive enhancer–promoter map. Genome Biol. 19, 56 (2018).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Shao, X. et al. MatrisomeDB: the ECM-protein knowledge database. Nucleic Acids Res. 48, D1136–D1144 (2020).
Sievers, Q. L. et al. Defining the human C2H2 zinc finger degrome targeted by thalidomide analogs through CRBN. Science 362, eaat0572 (2018).
Krönke, J. et al. Lenalidomide induces ubiquitination and degradation of CK1α in del(5q) MDS. Nature 523, 183–188 (2015).
Taniwaki, T. et al. Characterization of an exchangeable gene trap using pU-17 carrying a stop codon-beta geo cassette. Dev. Growth Differ. 47, 163–172 (2005).
Vanhoutteghem, A. et al. Basonuclin 2 has a function in the multiplication of embryonic craniofacial mesenchymal cells and is orthologous to disco proteins. Proc. Natl Acad. Sci. USA 106, 14432–14437 (2009).
Kuppe, C. et al. Decoding myofibroblast origins in human kidney fibrosis. Nature 589, 281–286 (2021).
Neph, S. et al. Circuitry and dynamics of human transcription factor regulatory networks. Cell 150, 1274–1286 (2012).
Fornes, O. et al. JASPAR 2020: update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 48, D87–D92 (2020).
Kulakovskiy, I. V. et al. HOCOMOCO: towards a complete collection of transcription factor binding models for human and mouse via large-scale ChIP-Seq analysis. Nucleic Acids Res. 46, D252–D259 (2018).
Xiao, Y. et al. Hippo pathway deletion in adult resting cardiac fibroblasts initiates a cell state transition with spontaneous and self-sustaining fibrosis. Genes Dev. 33, 1491–1505 (2019).
Kolvenbach, C. M. et al. Rare variants in BNC2 are implicated in autosomal-dominant congenital lower urinary-tract obstruction. Am. J. Hum. Genet. 104, 994–1006 (2019).
Ogura, Y. et al. A functional SNP in BNC2 is associated with adolescent idiopathic scoliosis. Am. J. Hum. Genet. 97, 337–342 (2015).
da Silveira, W. A. et al. Transcription factor networks derived from breast cancer stem cells control the immune response in the basal subtype. Sci. Rep. 7, 2851 (2017).
Lu, W. & Kang, Y. Epithelial-mesenchymal plasticity in cancer progression and metastasis. Dev. Cell 49, 361–374 (2019).
Shaw, T. J. & Rognoni, E. Dissecting fibroblast heterogeneity in health and fibrotic disease. Curr. Rheumatol. Rep. 22, 33 (2020).
Tsukui, T. et al. Collagen-producing lung cell atlas identifies multiple subsets with distinct localization and relevance to fibrosis. Nat. Commun. 11, 1920 (2020).
Henley, M. J. & Koehler, A. N. Advances in targeting ‘undruggable’ transcription factors with small molecules. Nat. Rev. Drug Discov. https://doi.org/10.1038/s41573-021-00199-0 (2021).
Lefebvre, P. et al. Interspecies NASH disease activity whole-genome profiling identifies a fibrogenic role of PPARα-regulated dermatopontin. JCI Insight. 2, e92264 (2017).
Caiazzo, R. et al. Roux-en-Y gastric bypass versus adjustable gastric banding to reduce nonalcoholic fatty liver disease: a 5-year controlled longitudinal study. Ann. Surg. 260, 893–898 (2014).
Louvet, A. et al. Main drivers of outcome differ between short term and long term in severe alcoholic hepatitis: a prospective study. Hepatology 66, 1464–1473 (2017).
Louvet, A. et al. Combining data from liver disease scoring systems better predicts outcomes of patients with alcoholic hepatitis. Gastroenterology 149, 398–406.e8 (2015).
Vandel, J. et al. Hepatic molecular signatures highlight the sexual dimorphism of nonalcoholic steatohepatitis (NASH). Hepatology 73, 920–936 (2021).
Mederacke, I., Dapito, D. H., Affò, S., Uchinami, H. & Schwabe, R. F. High-yield and high-purity isolation of hepatic stellate cells from normal and fibrotic mouse livers. Nat. Protoc. 10, 305–315 (2015).
Ploton, M. et al. The nuclear bile acid receptor FXR is a PKA- and FOXA2-sensitive activator of fasting hepatic gluconeogenesis. J. Hepatol. 69, 1099–1109 (2018).
Samanez, C. H. et al. The human hepatocyte cell lines IHH and HepaRG: models to study glucose, lipid and lipoprotein metabolism. Arch. Physiol. Biochem. 118, 102–111 (2012).
Guo, J. et al. Functional linkage of cirrhosis-predictive single nucleotide polymorphisms of Toll-like receptor 4 to hepatic stellate cell responses. Hepatology 49, 960–968 (2009).
Pankova, D. et al. RASSF1A controls tissue stiffness and cancer stem-like cells in lung adenocarcinoma. EMBO J. 38, e100532 (2019).
Edgar, R., Domrachev, M. & Lash, A. E. Gene expression omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 30, 207–210 (2002).
Davis, C. A. et al. The Encyclopedia of DNA elements (ENCODE): data portal update. Nucleic Acids Res. 46, D794–D801 (2018).
Roadmap Epigenomics Consortium. et al. Integrative analysis of 111 reference human epigenomes. Nature 518, 317–330 (2015).
Lizio, M. et al. Update of the FANTOM web resource: expansion to provide additional transcriptome atlases. Nucleic Acids Res. 47, D752–D758 (2019).
Afgan, E. et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 46, W537–W544 (2018).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013).
Anders, S., Pyl, P. T. & Huber, W. HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Vandel, J. et al. GIANT: galaxy-based tool for interactive analysis of transcriptomic data. Sci. Rep. 10, 19835 (2020).
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47–e47 (2015).
Zhou, Y. et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 10, 1523 (2019).
Chen, J., Bardes, E. E., Aronow, B. J. & Jegga, A. G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 37, W305–W311 (2009).
Dubois-Chevalier, J. et al. A dynamic CTCF chromatin binding landscape promotes DNA hydroxymethylation and transcriptional induction of adipocyte differentiation. Nucleic Acids Res. 42, 10943–10959 (2014).
Ewing, E., Planell-Picola, N., Jagodic, M. & Gomez-Cabrero, D. GeneSetCluster: a tool for summarizing and integrating gene-set analysis results. BMC Bioinforma. 21, 443 (2020).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e29 (2021).
Dubois-Chevalier, J. et al. The logic of transcriptional regulator recruitment architecture at cis-regulatory modules controlling liver functions. Genome Res. 27, 985–996 (2017).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Zhang, Y. et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Amemiya, H. M., Kundaje, A. & Boyle, A. P. The ENCODE Blacklist: identification of problematic regions of the genome. Sci. Rep. 9, 9354 (2019).
Nicol, J. W., Helt, G. A., Blanchard, S. G., Raja, A. & Loraine, A. E. The Integrated Genome Browser: free software for distribution and exploration of genome-scale datasets. Bioinformatics 25, 2730–2731 (2009).
Ramírez, F. et al. deepTools2: a next generation web server for deep-sequencing data analysis. Nucleic Acids Res. 44, W160–W165 (2016).
Dreszer, T. R. et al. The UCSC Genome Browser database: extensions and updates 2011. Nucleic Acids Res. 40, D918–D923 (2012).
Thomas-Chollier, M. et al. A complete workflow for the analysis of full-size ChIP-seq (and similar) data sets using peak-motifs. Nat. Protoc. 7, 1551–1568 (2012).
Frankish, A. et al. GENCODE reference annotation for the human and mouse genomes. Nucleic Acids Res. 47, D766–D773 (2019).
Dubois-Chevalier, J., Dubois, V., Staels, B., Lefebvre, P. & Eeckhoute, J. Perspectives on the use of super-enhancers as a defining feature of cell/tissue-identity genes. Epigenomics 12, 715–723 (2020).
Whyte, W. A. et al. Master transcription factors and mediator establish super-enhancers at key cell identity genes. Cell 153, 307–319 (2013).
Belhocine, M. et al. Dynamic of broad H3K4me3 domains uncover an epigenetic switch between cell identity and cancer-related genes. Genome Res. gr.266924.120, https://doi.org/10.1101/gr.266924.120 (2021).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria (2022).
Zhang, H.-M. et al. AnimalTFDB 2.0: a resource for expression, prediction and functional study of animal transcription factors. Nucleic Acids Res 43, D76–D81 (2015).
Shannon, P. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Szklarczyk, D. et al. STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 47, D607–D613 (2019).
Clapper, J. R. et al. Diet-induced mouse model of fatty liver disease and nonalcoholic steatohepatitis reflecting clinical disease progression and methods of assessment. Am. J. Physiol. Gastrointest. Liver Physiol. 305, G483–G495 (2013).
Mellacheruvu, D. et al. The CRAPome: a contaminant repository for affinity purification mass spectrometry data. Nat. Methods 10, 730–736 (2013).
Hu, H. et al. AnimalTFDB 3.0: a comprehensive resource for annotation and prediction of animal transcription factors. Nucleic Acids Res. 47, D33–D38 (2019).
Jones, G., Willett, P., Glen, R. C., Leach, A. R. & Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 267, 727–748 (1997).
Ding, N. et al. BRD4 is a novel therapeutic target for liver fibrosis. Proc. Natl Acad. Sci. USA 112, 15713–15718 (2015).
Fu, X. et al. Specialized fibroblast differentiated states underlie scar formation in the infarcted mouse heart. J. Clin. Investig. 128, 2127–2143 (2018).
Higuchi, Y. et al. Gastrointestinal fibroblasts have specialized, diverse transcriptional phenotypes: a comprehensive gene expression analysis of human fibroblasts. PLoS ONE 10, e0129241 (2015).
Reyfman, P. A. et al. Single-cell transcriptomic analysis of human lung provides insights into the pathobiology of pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 199, 1517–1536 (2019).
Acknowledgements
The authors are indebted to Julien Devassine and Kim Letten of the EOPS2 animal facility (Plateformes Lilloises en Biologie et Santé (PLBS) - UMS 2014 - US 41, Univ. Lille), Sophie Salomé and Nathalie Jouy of the BioImaging Center Lille (BICeL) platform (Plateformes Lilloises en Biologie et Santé (PLBS) - UMS 2014 - US 41, Univ. Lille) for their assistance. The authors also would like to thank members of Inserm U1011 including Drs M. Johanns, L. L’Homme, N. Hennuyer, X. Maréchal, M. Rosa and M. Gimenez as well as F. Maggiotto (INFINITE, Lille) for technical help or helpful discussions. This work was supported by the Agence Nationale de la Recherche (ANR) grants “HSCreg” (ANR-21-CE14-0032), “MEdicAL” (ANR-21-CE17-0016), “DeCodeNASH” (ANR-20-CE14-0034), “PreciNASH” (ANR-16-RHUS-0006-PreciNASH), French Proteomic Infrastructure (ProFI; ANR-10-INBS-08–03) and “European Genomic Institute for Diabetes” E.G.I.D. (ANR-10-LABX-0046), a French State fund managed by ANR under the frame program Investissements d’Avenir I-SITE ULNE/ANR-16-IDEX-0004 ULNE. This project was also co-funded by The European Union under the European Regional Development Fund (ERDF) and by the Hauts de France Regional Council (contract 20000007 and contract 17003781), the MEL (contract_2020_ESR_02 and contract_2016_ESR_05) and the French State (contract no. 2019-R3-CTRL_IPL_Phase3 and contract no. 2017-R3-CTRL-Phase 1). Work in INFINITE was supported by a grant from the Fondation Recherche Médicale (FRM) (Equipe labellisée, EQU202003010299). BS is a recipient of an Advanced ERC Grant (694717).
Author information
Authors and Affiliations
Contributions
Conceptualization: M.B.-G., F.P.Z., F.P., L.D., B.S., P.L., and J.E.; methodology: M.B.-G., C.B., F.P.Z., J.D.-C., C.G., M.B.S., G.L., J.-M.S., A.F., M.P., J.V., L.F., L.C.N.-W., B.P., and J.T.H.; software: G.L., J.D.-C., J.V., and K.R.; validation: M.B.-G., C.B., J.D.-C., and C.G.; formal analysis: M.B.-G., J.D.-C., and J.E.; investigation: M.B.-G., C.B., F.P.Z., J.D.-C., C.G., M.B.S., J.-M.S., A.F., M.P., J.V., N.V., E.W., A.D., E.B., A.K.C., C.G., E.V., R.P., K.R., and J.T.H.; resources: L.G., P.M., V.R., R.C., S.C., F.P., L.D., V.G., and E.L.; data curation: M.B.-G., J.D.-C., and J.E.; writing—review & editing: M.B.-G. and J.E. with inputs from co-authors; visualization: G.L., M.B.-G., C.B., F.P.Z., A.F., K.R., J.C.-D., and J.E.; project administration: B.S., P.L., and J.E.; funding acquisition: B.S., P.L., and J.E.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Hisham Mohammed and the other, anonymous, reviewers for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bobowski-Gerard, M., Boulet, C., Zummo, F.P. et al. Functional genomics uncovers the transcription factor BNC2 as required for myofibroblastic activation in fibrosis. Nat Commun 13, 5324 (2022). https://doi.org/10.1038/s41467-022-33063-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-022-33063-9
- Springer Nature Limited
This article is cited by
-
Keratinocytes drive the epithelial hyperplasia key to sea lice resistance in coho salmon
BMC Biology (2024)
-
Deciphering the spatial landscape and plasticity of immunosuppressive fibroblasts in breast cancer
Nature Communications (2024)
-
O-GlcNAcylation controls pro-fibrotic transcriptional regulatory signaling in myofibroblasts
Cell Death & Disease (2024)
-
Transcription factors in fibroblast plasticity and CAF heterogeneity
Journal of Experimental & Clinical Cancer Research (2023)
-
An integrated gene-to-outcome multimodal database for metabolic dysfunction-associated steatotic liver disease
Nature Medicine (2023)