

Abstract
Neutrophils are evolutionarily conserved innate immune cells playing pivotal roles in host defense. Zebrafish models have contributed substantially to our understanding of neutrophil functions but similarities to human neutrophil maturation have not been systematically characterized, which limits their applicability to studying human disease. Here we show, by generating and analysing transgenic zebrafish strains representing distinct neutrophil differentiation stages, a high-resolution transcriptional profile of neutrophil maturation. We link gene expression at each stage to characteristic transcription factors, including C/ebp-β, which is important for late neutrophil maturation. Cross-species comparison of zebrafish, mouse, and human samples confirms high molecular similarity of immature stages and discriminates zebrafish-specific from pan-species gene signatures. Applying the pan-species neutrophil maturation signature to RNA-sequencing data from human neuroblastoma patients reveals association between metastatic tumor cell infiltration in the bone marrow and an overall increase in mature neutrophils. Our detailed neutrophil maturation atlas thus provides a valuable resource for studying neutrophil function at different stages across species in health and disease.
Similar content being viewed by others


Introduction
Neutrophils are the most abundant immune cell population in humans and the first responders to injury and infection1,2. In mammals, neutrophils mature in the bone marrow (BM), during the final steps of a cascade where hematopoietic stem cells differentiate through a granulocyte-monocyte progenitor towards neutrophils3,4. Characteristic granules form throughout development from the promyelocyte to segmented neutrophil stage5,6,7. Transcription factors of the C/EBP family are key for the expression of granule enzymes8,9. Surface marker expression, nuclear morphology, and granule content have long been used to define neutrophil maturation stages, but recent studies using single-cell RNA sequencing (scRNA-seq) and proteome analysis (CyTOF) have questioned this classification scheme instead suggesting a sequence of continuous transcriptional stages10,11,12,13,14,15.
Functional diversity of neutrophils at different maturation grades is still understudied16,17. A potential role for different maturation states becomes apparent in cancer, where immature neutrophils often accumulate in blood and tumors, and can have altered effector functions such as reduced phagocytosis, ROS production, NETosis, granularity, chemokine receptor expression and increased suppressive functions6,16,18. Neutrophils are now known to be involved in almost every stage of cancer such as tumor initiation by ROS production, growth, angiogenesis, and the conditioning of the pre-metastatic niche6,17. Different tumor-associated neutrophil (TAN) populations polarized by TGF-β or IFN-β towards pro- or anti-tumor roles have been observed. Interestingly, their opposing roles in cancer progression have been linked to different maturation stages and densities18,19,20.
Zebrafish are a versatile model to study neutrophil functions in infection and tumorigenesis thanks to the availability of many fluorescent transgenic lines and the prospect of intravital imaging21,22,23. Hematopoiesis in zebrafish occurs in the kidney marrow, where all major immune cell types known from humans are present and are generally considered evolutionarily conserved24,25. However, to date the transcriptional states of zebrafish neutrophils during maturation and associated functions in vivo have not been mapped to their human equivalents, thus limiting cross-species comparisons. Comparative transcriptomic studies have been hampered by under-representation of neutrophils in many human datasets due to technical difficulties and by a lack of suitable zebrafish lines allowing effective sorting of neutrophils comprising multiple maturation stages26.
Here, we establish Tg(lysC:CFPNTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 double transgenic zebrafish, which allow us to distinguish immature from mature neutrophils in vivo, visualize their interactions with bacteria and tumor cells, and isolate maturing neutrophil populations for morphological and transcriptional analysis. We use scRNA-seq to catalog transcriptional changes during maturation and to identify critical transcription factors, including C/ebp-β. Finally, cross-species comparison enables the definition of a conserved gene signature, which we apply to analyze bulk human tumor transcriptome data and to correlate neutrophil maturation stage with BM metastasis.
Results
The Mmp9 transgene identifies mature neutrophils in zebrafish
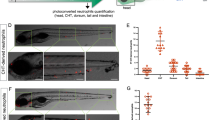
In order to distinguish immature from mature neutrophils in vivo, we generated transgenic zebrafish expressing membrane-directed Citrine under the control of regulatory elements for mmp9 (Tg(BACmmp9:Citrine-CAAX)vi003), a tertiary granule protein in mammalian mature neutrophils and expressed in zebrafish mature heterophils27,28. Tg(BACmmp9:Citrine-CAAX)vi003 zebrafish embryos/larvae report mmp9 transcription from 2 days post fertilization (dpf) in the epithelia of the tail fin and the distal gut (Supplementary Fig. 1a). Additionally, a dotted pattern became apparent along the head, yolk, and in the caudal hematopoietic tissue (CHT) suggesting expression in a leukocyte population. We confirmed that Citrine fluorescence specifically reports mmp9 expression by detecting mmp9 RNA in FACS-purified Citrine+ but not in Citrine- cells (Supplementary Fig. 1b).
To examine the identity of Mmp9+ leukocytes, mmp9:Citrine fish were crossed with fish double-transgenic for myeloid markers Tg(lysC:CFP-NTR)vi002 (labeling neutrophils), and Tg(mpeg:mCherry)gl23 (macrophages)29. Live imaging of triple-transgenic larvae at 3 and 5 dpf confirmed the existence of both Mmp9- and Mmp9+ subpopulations of lysC:CFP+ neutrophils with a more pronounced co-localization in the head than in the caudal hematopoietic tissue region (Fig. 1a). Mmp9+LysC+ cells had low mpeg:mCherry expression, while bona-fide macrophages were highly (2.5-fold higher signal intensity) positive for mpeg:mCherry and few of the latter showed detectable mmp9 expression (Fig. 1b and Supplementary Fig. 1c). LysC+Mpeg+Mmp9+ triple-positive neutrophils were enriched in the head region at 3 dpf (mean = 36%; range = 24.3−52.3%) as well as in the head and tail at 5 dpf (mean = 37%; range = 29−46.8%; and 32%, range = 20.5−40%, respectively), whereas the CHT population decreased from 3 to 5 dpf (from mean = 20%, range = 11.1−30.3%, to 13%, range = 10−17.6%) (Supplementary Fig. 1c). Pre-dominant localization of triple-positive neutrophils in the head points towards an already more progressed maturation stage of RBI-derived head neutrophils compared to HSC-derived CHT neutrophils. Interestingly, a recent publication also described transcriptional differences between these two neutrophil populations30.

a Confocal images of triple transgenic larvae Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003/ Tg(mpeg:mCherry)gl23 reveal myeloid cells with different expression levels of the three analyzed markers. Stitched whole-mount images of a 5 dpf (days post fertilization) larva. n = 3. b High magnification showing presence of three different cell types: #1 neutrophil: lysC:CFP+/ mpeg:mCherry+/ mmp9:Citrine+; #2 neutrophil: lysC:CFP+/ mpeg:mCherry-/ mmp9:Citrine-; #3 macrophage: lysC:CFP-/ mpeg:mCherry+/ mmp9:Citrine-. c Flow cytometric analysis (BD LSRFortessa) of myeloid cells from mpeg:mCherry-/ lysC:CFP+ control larvae, lysC:CFP-/ mpeg:mCherry+ controls, or triple transgenic larvae lysC:CFP+/ mmp9:Citrine+/ mpeg:mCherry+(dot plots left to right) at 3 dpf. Histogram and box plot showing expression of mmp9:Citrine in myeloid cells (combined gates of Q1, Q2, Q3; Minimum = 12.4%, Maximum = 17.1%, Median = 15.7%; Box 25th−75th percentile). Violin plots indicating the distribution and expression levels of mmp9:Citrine among myeloid populations (mmp9:Citrine+ gate; n = 5 with pools of 15 triple transgenic larvae each). mfi = mean fluorescence intensity (d) mmp9:Citrine+ neutrophils (red histogram) have a higher side scatter (SSC) and therefore granularity compared to mmp9:Citrine- neutrophils (blue histogram). Flow cytometric analysis of cells isolated from Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 at 2 dpf. Cells were gated on the lysC:CFP+/ mmp9:Citrine+ or lysC:CFP+/ mmp9:Citrine- populations and analyzed for SSC (n = 3). e Cells were isolated from kidneys, spleen or blood of adult Tg(lysC:dsRed)nz50Tg/ Tg(BACmmp9:Citrine-CAAX)vi003 and analyzed by flow cytometry for frequency of Mmp9+ cells. f Neutrophils were isolated from whole kidney marrow (WKM) of adult Tg(lysC:dsRed)nz50Tg/ Tg(BACmmp9:Citrine-CAAX)vi003 zebrafish and sorted into lysC:dsRed+/ mmp9:Citrine+ or lysC:dsRed+/ mmp9:Citrine- populations, cytospins were prepared and Pappenheim stained. Different neutrophil maturation stages were scored blinded, showing that the lysC:dsRed+/ mmp9:Citrine+ population consists of highly differentiated neutrophil stages. INT = intermediate; HI = high. Graph presents average percentages of n = 4 kidneys (721 Mmp9+ cells and 819 Mmp9- cells were analyzed in total). g Representative ultrastructural images show an unsegmented nucleus and round and elongated granules in lysC:CFP+/ mmp9:Citrine- cells (n = 5 from one TEM transmission electron microscopy experiment) contrasting with a segmented nucleus and predominantly elongated granules in lysC:CFP+/ mmp9:CitrineHIcells (n = 2); *, round granules; #, elongated granules.
Complementary analysis by flow cytometry revealed that 15.4% (mean; range: 12.4−17.1%) of myeloid cells were expressing Mmp9; of those 67% (mean; range 61.8−69.5%; Q2) belonged to the LysC+Mpeg+ neutrophil population (Fig. 1c, Supplementary Fig. 2a). Fewer Citrine-positive cells were part of the LysC+Mpeg- neutrophil (mean= 29.06%; range: 25.8−34.8% Q1) and Mpeg+LysC- macrophage (mean = 3.9%; range 2.8−5.8%; Q3) populations. Mmp9 expression was restricted to a subpopulation of cells stained with Sudan Black, a lipophilic dye labeling granules present in maturing neutrophils (Supplementary Fig. 1d)31. Furthermore, Mmp9+ cells showed an increased side scatter (SSC) compared to Mmp9- cells indicating a higher granularity and intracellular complexity, which suggests a mature phenotype (Fig. 1d).
Time-lapse imaging of cellular behavior after wounding showed that both, LysC+Mmp9+ and LysC+Mmp9- neutrophils had a similar round morphology and moved to and from the wound quickly with amoeboid motility as described for neutrophils32 (Supplementary Movie 1). In contrast, Mpeg+ cells showed the protrusions typical of macrophages and stayed close to the wound edge.
In adult zebrafish LysC+Mmp9+neutrophils were detectable in the whole kidney marrow (WKM), the primary hematopoietic organ of teleost fish, as well as in the spleen and blood (Fig. 1e). We investigated morphology and maturation grade of FACS-sorted LysC+Mmp9+ and LysC+Mmp9- populations from WKM on stained cytospins and by electron microscopy (Fig. 1f, g, Supplementary Fig. 1e−i). Only 3% (mean, range = 0−9%) of Mmp9- cells contained segmented nuclei compared to 23% (mean, range = 3−47%) of the Mmp9+ fraction, indicating an advanced maturation grade of the latter (Fig. 1f). LysC+Mmp9HI neutrophils were also of smaller cell size than Mmp9- cells another parameter for a more differentiated stage (Supplementary Fig. 1g). The ultrastructure of LysC+Mmp9HI cells revealed more developed cigar-shaped granules, while granules in Mmp9- cells were generally rounder (Fig. 1g, Supplementary Fig. 1i). Collectively, our data show that Mmp9 is a suitable marker for mature neutrophils in zebrafish consistent with previous data for human cells33.
Mmp9+ neutrophils are highly phagocytic and rapidly recruited to wounds
Neutrophils are rapidly recruited to sites of injury through chemotactic cues34. When zebrafish larvae were wounded at the ventral fin, Mmp9HI neutrophil tracks came closer to the wound (mean = 8.3 µm HI, 45.8 µm NO, 43 µm INT; n = 6) and showed a lower linearity of movement (mean = 0.23 HI, 0.42 NO, 0.39 INT; n = 6) than Mmp9INT or NO cells resulting from a meandering movement in the wound area, while their speed was similar (mean = 0.055 µm/s HI, 0.045 µm/s NO, 0.053 µm/s INT) (Fig. 2a−d). The phagocytic capacity of neutrophils increases with their maturation grade35. We performed in vivo phagocytosis assays by injecting live mCherry-labeled E. coli into 2 dpf Tg(lysC:CFPNTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 larvae (Fig. 2e−g; Supplementary Fig. 2b). Both neutrophil subpopulations, LysC+Mmp9- and LysC+Mmp9+, were able to ingest bacteria (Fig. 2e) and to produce reactive oxygen species (Supplementary Fig. 2c). Quantification of E. coli uptake as seen by overlap of mCherry (E. coli) and CFP (neutrophil) fluorescence by flow cytometry showed a significantly higher percentage of Mmp9+ neutrophils with bacterial cargo compared to Mmp9- neutrophils (mean = 23.8% and 10.7%, respectively; n = 5; paired t test, P = 0.007) (Fig. 2f; Supplementary Fig. 2b). Furthermore, Mmp9+ neutrophils were more efficiently recruited to sites of bacterial infection (67.3% Mmp9+ vs. 32.7% Mmp9- of all LysC+ neutrophils at infection site; n = 33; paired t test; P = 0.002) (Fig. 2g) and the overall frequency of Mmp9+ cells was increased during infection (mean = 56.4% Mmp9+ in infected and 39.2% Mmp9+ in PBS treated, n = 5; paired t test; P = 0.009) (Supplementary Fig. 2d, e).

a−d Recruitment of neutrophils to an injury of the fin at 2 dpf (days post fertilization) lysC:CFP+ mmp9:CitrineHI, lysC:CFP+ mmp9:CitrineINT and lysC:CFP+ mmp9:Citrine- neutrophil tracks (n = 21; 13; 26 cells, df = 57, respectively; n = 6 larvae; One-way ANOVA with Dunnett’s test) were analyzed over a 2 h period: b for distance to a wound ROI (P = 0.0002, F = 9.955), (c) linearity of forward progression (mean straight line speed/ track mean speed) (P = 0.0086, F = 5.172) and (d) speed (P = 0.1315, F = 2.103). ns = not significant. INT = intermediate expression; HI = high expression. e−g In vivo phagocytosis assays were performed by injecting mCherry-labeled E. coli into the caudal vein or otic vesicle of Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 zebrafish larvae at 2 dpf. e E. coli were observed inside both, lysC:CFP+ mmp9:Citrine+ and lysC:CFP+ mmp9:Citrine- neutrophils 6 hpi (hours post infection). f Phagocytosis experiments analyzed by flow cytometry (n = 5, each approx. 20 larvae; two-tailed paired t test, P = 0.007; df = 4); g Neutrophil recruitment to E.coli-mCherry injected into the otic vesicle of 3 dpf larvae was analyzed by confocal microscopy (n = 33, two-tailed paired t test, P = 0.002, df = 32). h−k Neutrophil- pre-neoplastic cell interactions were observed by confocal microscopy in Et(kita:GAL4)hzm1/ Tg(UAS:EGFP-HRAS_G12V)io006/ Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 zebrafish larvae starting at 78 hpf. h Still Images of z-stack maximum projections from a time-lapse movie showing Mmp9+ neutrophils forming dynamic contacts with GFP+ kita tumor cells. White arrows point at a GFP+ cell tether. Z-stacks were acquired every 88 s. i Snapshots and cell footprints were taken from the same time-lapse movie (t = 5 h). Superimposition of lysC:CFP and mmp9:Citrine from all time frames generating neutrophil footprints (bottom). White arrow points out how the movements of an Mmp9+ neutrophil copied the outline of the RAS-GFP+ cluster seen in the snapshot (top). j Close-up clippings showing an Mmp9+ neutrophil spreading over the pre-neoplastic cell cluster marked by a white arrow in (i). k Quantification of neutrophil-pre-neoplastic interactions were performed from maximum projections of different time-lapse movies. The frequency of interacting neutrophils of each subpopulation (no, intermediate or high mmp9:Citrine levels) getting into close, intense interactions with GFP+ kita/RAS skin pre-neoplastic cells. (n = 8, one-way ANOVA, P = 0.0022, F = 8.561, df = 19).
Next, we examined neutrophils in a model of pre-neoplastic melanoma (Et(kita:GAL4)hzm1x Tg(HRAS_G12V:UAS:CFP)vi004)36. As during infection, we found an increase in Mmp9+ cells (mean 46.0% Mmp9+ in the presence of HRASG12V versus 37.4% Mmp9+ in controls; n = 5; paired t test; P = 0.005) (Supplementary Fig. 2f). Live imaging in transparent zebrafish larvae showed that some Mmp9+ cells stayed in contact with transformed cells by thin tethers (Fig. 2h) and others interacted over a long period (Supplementary Movie 2), consistent with previous data for LysC+ neutrophils37. We found that some neutrophils got into close contact, spread out, and crawled over HRASG12V+ cells, seemingly scanning their surface (Fig. 2j). Cell footprints of those neutrophil movements trace the outline of HRASG12V+ clusters (Fig. 2i). Notably, this spreading behavior around HRASG12V+ cells was significantly enriched in Mmp9HI cells (mean 56.5%; n = 8; one-way ANOVA, P = 0.0022) compared to Mmp9INT (17.3%) or Mmp9- (17.9%) cells (Fig. 2k, Supplementary Movie 3).
In summary, we observed that mmp9 transgene expression correlates with a high maturation grade in neutrophils with increased effector functions, such as recruitment and phagocytosis during bacterial infections and augmented interactions with pre-neoplastic cells.
Single-cell transcriptomics defines zebrafish neutrophil maturation stages
To scrutinize neutrophil maturation in zebrafish at the molecular level and to enable cross-species comparison to mammals, we performed droplet-based scRNA-seq on WKM and on LysC+ neutrophil populations expressing different Mmp9 levels (NO, INT, HI) sorted from the WKM of two adult Tg(lysC:CFPNTR)vi004/ Tg(BACmmp9:Citrine-CAAX)vi003 zebrafish (Fig. 3a, Supplementary Fig. 3a). The generated dataset comprised a total of 18,150 cells passing quality control (Fig. 3b, Supplementary Fig. 3b−g, Supplementary Data 9).

a Schema showing the workflow for cell isolation, multiplexing and scRNA-seq, as well as the bioinformatics analyses applied. NO = no expression; INT = intermediate expression; HI = high expression. WKM = whole kidney marrow. b Uniform Manifold Approximation and Projection (UMAP) of single-cell RNA-seq data (n = 18,150 cells) showing cells from FACS-sorted neutrophil populations (lysC+/ mmp9 = NO, INT, HI) and unsorted whole-kidney marrow cells. The cluster labeled with an asterisk (*) represents a group of cells dominated by cell cycle effects (cp. Supplementary Fig. 3k–m). c, d Reference-based labeling of cell types by projecting cells to two zebrafish hematopoietic reference atlases in the same UMAP as in panel b24,38. As expected, a strong overlap with genetically labeled Tg(mpx:GFP) neutrophils is observed (highlighted in color). e Harmony82 plot of the inferred neutrophil maturation trajectory in zebrafish (n = 15,876 cells). A continuous trajectory indicating a maturation continuum is observed. f Line plots of the distribution of sorted (NO, INT, HI) and unsorted neutrophil populations along the inferred trajectory. g Smoothed expression of lyz and mmp9 in neutrophils along the inferred trajectory.
Sorted neutrophil populations and neutrophils from WKM overlapped in our dataset, indicating that our sorting strategy captured all neutrophils present in WKM (Fig. 3b). To investigate the maturation dynamics, we focused on cells consistently annotated as neutrophils (n = 15,876 cells) based on a bioinformatic mapping to two independent reference datasets24,38 (Fig. 3c, d) and used the Slingshot algorithm to infer the structure of the underlying trajectory (Fig. 3e, f)39. Consistent results were also obtained with other trajectory inference algorithms (Supplementary Fig. 4) and the validity and directionality of this trajectory was supported by (i) the sequential order of the sorted subpopulations along the trajectory (in order: Mmp9NO, Mmp9INT, Mmp9HI; Fig. 3f), (ii) decreasing levels of lysozyme C (lyz) and increasing levels of mmp9 (Fig. 3g), and (iii) a decrease in the number of cycling cells (inferred bioinformatically using the CellCycleScoring function from the Seurat package; Supplementary Fig. 3h). A cluster separated from the main neutrophil population was mainly formed by cells in the G2/M phase (labeled by *, Fig. 3b, Supplementary Fig. 3e, k, l, m).
We used the tradeSeq40 algorithm to define the genes associated with neutrophil maturation (Padj < 0.05, top 1500 in descending order based on Wald-statistic; Supplementary Data 1). Based on expression patterns of these genes, we partitioned the cells into four maturation phases along the trajectory (P1 = early, P2 = early/intermediate, P3 = intermediate/late, P4 = late). Similarly, we partitioned the genes into three modules (M1 = early, M2 = intermediate, M3 = late) (Fig. 4a). While M1 and M2 were almost exclusively expressed in phase P1 and P2, respectively, M3 genes were expressed in both P3 and P4 (Fig. 4b). Functional enrichment analysis using hypeR41 indicated that earlier modules M1 and M2 related to cell cycle and proliferation while module M3 comprised genes associated with functions of mature neutrophils like migration, immune activation, and inflammation (Fig. 4c, Supplementary Data 3).

a Heatmap showing differentially expressed genes (tradeSeq;40Padj < 0.001, top 1500 based on Wald-statistic) along the maturation trajectory. Cells have been allocated to four distinct phases (P1-P4) and into three distinct gene modules (M1-M3) by hierarchical clustering. The columns of the heatmaps correspond to 100 ordered discrete bins covering the maturation trajectory. The bottom line-annotation shows the distribution of sorted subpopulations along the maturation trajectory (see Fig. 3f) and the bar shows the pseudotime score (see Fig. 3e). b Heatmap summarizing average gene expression per module and phase (from a). c Top-5 enriched gene sets per module from the indicated source pathway databases (hypeR41 over-representation analysis). d Heatmaps of differentially expressed genes (excerpts from a) associated with selected neutrophil-related functional pathways.
We next examined genes associated with certain neutrophil functions in detail (Fig. 4d)6,8,12,14. First, we found proliferation factor myca expressed during early maturation (P1), while advancing maturation (P3, P4) was associated with anti-apoptotic genes such as mcl1a, mcl1b and mxi1, a negative regulator of myc42. Second, we detected genes related to oxidative stress response (selenoh43, abcc13, prdx1) early in P1, which could aid to protect progenitor cells from oxidative damage. Third, we found a down-regulation of the marrow retention factor cxcr4b in P4, and conversely an upregulation of cxcr1, lyn, and the migration-related transcription factor atf3 in P4, in line with a putative switch towards a migratory phenotype44.
Finally, we also found genes encoding granule-related proteins expressed in specific phases (Fig. 4d), these granules have conventionally been used for neutrophil staging in mammals5. At the beginning of the trajectory lyz from P1 on (compare Fig. 3g), primary granule-related genes cd63, srgn, and mpx from P2 on, secondary granule-related NADPH oxidase subunits cybb and cyba from P3 on, and the gelatinase mmp9 and mmp13a.1 (a putative orthologue of human MMP1) in P4. The expression of these genes is consistent with a staging of human neutrophils from myeloblasts (P1) through pro-myelocytes (P2) and myelocytes/metamyelocytes (P3) to banded/segmented neutrophils (P4). The last phase (P4) was also associated with the upregulation of the pro-inflammatory cytokine il1b (Fig. 4d), confirming data from mouse neutrophils12,14.
Taken together, our data demonstrate that neutrophils in the kidney marrow of zebrafish mature in a continuous process advancing from a proliferative stage to a post-mitotic, anti-apoptotic, and migratory phase. Detection of genes associated with certain granule types suggests a similar sequence of granule production as in mammals.
C/ebp-β governs expression of late granule genes during maturation
Next, we sought to identify the key regulators of the neutrophil maturation process in zebrafish. Many known neutrophil TFs displayed dynamic expression patterns along the trajectory (Fig. 5a)8. We reasoned that the expression of key regulatory TFs was closely correlated with the expression of their target genes, albeit possibly with a temporal delay. We therefore compared TF expression with the expression of genes in each module using dynamic time warping (DTW) analysis, which highlighted ybx1, also known as key splicing factor in hematopoietic development45, hmgb2b and dnajc1 as top regulators of early maturation genes in module M1 (Fig. 5b, c; Supplementary Fig. 5; Supplementary Data 4, 5). In contrast, cebpb and atf3 emerged as the top-ranked transcription factors for late maturation (M3). C/EBP-β governs demand-driven granulopoiesis during infection in zebrafish and in mammals46,47 and has also been identified as a late-expressed transcription factor in neutrophils in mice12,14. However, a role during steady-state neutrophil maturation has not been described to date.

a Heatmap showing regulation of selected transcription factors (TFs) during neutrophil maturation. The bottom line-annotation shows the distribution of sorted subpopulations along the maturation trajectory (see Fig. 3f) and the bar shows the pseudotime score (see Fig. 3e). b Ridge plot showing top candidate regulators and their specificity to target genes in each module. Here, “module specificity” measures the similarity of the expression of a TF to all target genes in the neutrophil maturation modules compared to other TFs. It was calculated as 1 minus the dynamic time warping (DTW) distance and scaled relative to all other TFs. Thus, a high score on the x-axis indicates a target gene with a closely matching and specific expression pattern for the TF. The y-axis shows the distribution density for all genes in each module (M1, M2, M3; different colors). c Expression of top TFs ybx1, compared to its putative target genes in module M1 (top), and cebpb, compared to genes in M3 (bottom). d qPCR validation of cebpb expression in sorted lysC:CFP+/ mmp9:Citrine+ and lysC:CFP+/ mmp9:Citrine- neutrophils. (n = 3, each sample sorted from a pool of 160 larva; repeated measures ANOVA).
We hypothesized that C/ebp-β is involved in the regulation of neutrophil maturation in zebrafish. After confirming that cebpb expression in sorted LysC+Mmp9+ neutrophils was significantly higher than in LysC+Mmp9- neutrophils and unsorted cells (repeated measures ANOVA, P < 0.05, Fig. 5d), we targeted cebpb translation using a published AUG-binding morpholino (Fig. 6a). No systemic adverse effects were observed, knockdown did not influence steady-state neutrophil numbers, in line with previous data (Fig. 6b)46. We examined whether C/ebp-β regulates the augmented phagocytic function of Mmp9+ neutrophils but found no effect on uptake of mCherry-labeled E.coli (Fig. 6c). Cebpb knock down affected expression of the published C/ebp-β target genes14, mmp9 and fcer1gl, but not of spi1b and mcl1a in FACS-sorted LysC+Mmp9+ neutrophils (Fig. 6d). Strikingly, we found that cebpb morpholino treatment reduced the frequency of LysC+Mmp9+ neutrophils at 2 and 3dpf (Fig. 6e), indicating an instructive role of C/ebp-β in the differentiation towards mature Mmp9-expressing neutrophils. Conversely, cebpb RNA overexpression led to higher LysC+Mmp9+ neutrophil frequencies and mmp9 expression levels, further supporting this role (Fig. 6f). To confirm the function of C/ebp-β in adult neutrophils we generated cebpbMUT vi006 fish by CRISPR-Cas9 introducing a 76 bp deletion plus a 2 bp insertion leading to a frameshift and premature stop codon (Supplementary Fig. 6a, b). In agreement with the results in larvae, we observed a strongly diminished LysC+Mmp9+ population in the WKM of homozygous cebpbMUT vi006 fish (in mmp9:CitHOM mean = 1.4%; in mmp9:CitHET mean = 5.1%) compared to cebpbWT (in mmp9:CitHOM mean = 32%; in mmp9:CitHET mean = 25.1%) (Fig. 6h), but no change in the frequency of LysC+ neutrophils (Fig. 6i). Together, these data indicate that C/ebp-β – in addition to its role in demand-driven hematopoiesis – regulates aspects of steady-state neutrophil maturation such as generation of mature Mmp9+ neutrophils and expression of granule proteins but not their phagocytic function.

a–e Analyses of standard (Std)-morpholino (MO) or cebpb-morpholino treated Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 larvae at 3 dpf (days post-fertilization). a Schematic drawing of subsequent experiments. b ImageJ analysis of LysC+ leukocyte numbers in the tail region of untreated, control (ctrl) standard morpholino or cebpb morpholino-injected larvae (n = 28; 23; 28 larvae, respectively, one-way ANOVA with Tukey´s test, P = 0.1343, F = 2.041, representative image shown to the left). Leica SP8 confocal images with a HC PL APO CS 10x/0.40 DRY; Zoom 0.85x. c In vivo phagocytosis assay with E.coli-mCherry injected into the caudal vein after cebpb morpholino treatment. Representative flow cytometry plots (left). d qPCR of selected target genes of C/ebp-β (selected from Xie et al.14.) in FACS-sorted cells (n = 4; sorted from 65-102 larvae each; one-way ANOVA with Tukey´s test; all P < 0.0001, df = 18) after morpholino treatment at 3 dpf. ns = not significant. e Representative flow cytometry plots show reduction in LysC+Mmp9+ neutrophils after cebpb morpholino treatment (left). Plots summarizing four independent experiments each with pools of approx. 20 larvae per group analyzed by flow cytometry at 2 and 3 dpf after morpholino treatment (right). Two-tailed paired t test. f Frequencies of LysC+Mmp9+ neutrophils and mean mmp9:Citrine expression are increased after cebpb full-length RNA overexpression. Analyzed by flow cytometry 3 dpf after mRNA injection (n = 7; pools of 20 larvae each), two-tailed paired t test. g−i Analyses of neutrophils from adult Cebpb mutant fish. WKM = whole kidney marrow. g Schematic drawing. WKM of adult Cebpb wildtype (cebpbWT) or Cebpb mutant (cebpbMUT vi006) Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 fish was evaluated by flow cytometry. h Representative dot plots (left). Violin plot summarizing percentages of LysC+Mmp9+ neutrophils of all LysC+ neutrophils in mmp9:CitHOM (n = 5) mmp9:CitHET (n = 6) cebpbWT and mmp9:CitHOM (n = 4) mmp9:CitHET (n = 6) cebpbMUT vi006 kidneys. (One-way ANOVA with Tukey´s test P = 0.0006, F = 9.656, df = 17). i Representative dot plots (left). Violin plot summarizing percentages of LysC+ neutrophils of all live cells in cebpbWT (n = 11) and cebpbMUT vi006 (n = 10) kidneys (two-tailed unpaired t test; P = 0.3797).
Cross-species comparison allows staging of zebrafish neutrophil maturation phases and identifying conserved gene signatures
Next, we wanted to gauge the conservation of neutrophil maturation trajectories between zebrafish and mammalian species, an important piece of information for investigating and modeling granulopoiesis across species. To address this point, we collected transcriptome datasets capturing progenitor, early, and late neutrophil maturation stages in human and mice. We included both in vivo and in vitro data in this selection11,13,48,49,50,51. We then followed two complementary approaches to assess similarities at the cellular and molecular level.
First, we integrated all datasets with the four phases of neutrophil maturation that we defined in zebrafish (aggregating transcriptome profiles of single cells per phase) and used unsupervised hierarchical clustering to group similar stages in an unbiased manner (Fig. 7a, Supplementary Data 10; zebrafish phases P1-P4 highlighted with dotted lines). We found that phase P1 clustered together with early stages of human in vitro differentiation (Neu3h-12h51) and unipotent neutrophil precursors and early neutrophils in mice (ly6c_neg_gmp13, preNeu11, proNeu-1/250, proneu113) and human (NCP1-448, eNeP49). Phase P2 corresponds to the transition stage that includes preNeu-150 and proneu213 in mouse and promyelocytes (PM)48, myelocytes (MY)48 and N1-neutrophils excluding the early progenitor portion (“N1 w/o eNeP”)49 in human. Finally, both P3 and P4 phases clustered with cells reaching the end of in vitro differentiation (Neu96h)51 and the differentiated and mmp9 positive mouse neutrophils described by Muench et al. 2020 (immNeu)50, as well as banded cells (BC)48. Other cells captured from peripheral blood (blood_neu11, mature_neu11, Neuts49) as well as segmented neutrophils (SN)48 and PMN48 also clustered with the zebrafish neutrophils at stages P3-P4 although at higher distance, which is consistent with ongoing maturation after exiting the bone marrow, a process missing in the zebrafish WKM neutrophil data. Altogether, this analysis indicated that the four zebrafish phases mirrored major stages of neutrophil maturation in mouse and human.

a Heatmap of hierarchically clustered neutrophil maturation stages across studies11,13,48,49,50,51 based on zebrafish neutrophil maturation signatures (M1ortho, M2ortho, M3ortho). The four aggregated zebrafish phases are highlighted by a dotted box. Column annotations on the top show dataset annotation and row annotations on the left show module membership of each gene. Panels display zebrafish kidney marrow (b), mouse (c) and human (d) bone marrow (BM) scRNA-seq data ordered by their own maturation trajectory10,12,14,52,53,54. Ordered from top to bottom, column annotations display dataset annotations, inferred trajectories per dataset with pseudotime, cell density (histograms) and quality-flagged bins (<= 3 cells; potentially less reliable), heatmaps for modules M1ortho-M3ortho, and author-provided annotations. Left side annotation bars show module membership of each gene orthologue. e, f Average cross-correlation lag between mouse and human datasets (from panels c, d) and the corresponding zebrafish gene.
Second, we asked whether the neutrophil maturation followed similar dynamics in all species at the transcriptomic level by comparing the maturation trajectory from our zebrafish model (Fig. 7b) to those from mouse10,12,14 (Fig. 7c) and human52,53,54 (Fig. 7d) based on published bone marrow scRNA-seq data (Supplementary Data 10). For each trajectory, we aligned the expression patterns of genes along the maturation trajectory to the corresponding homolog along the zebrafish trajectory using cross-correlation (Fig. 7e, f). We found that gene expression in early development (M1ortho) was highly congruent with their mouse and human orthologues in all datasets. At later stages (M2ortho and M3ortho modules), expression profiles diverged, but a subset of genes in each module was found conserved across all datasets (Fig. 7e, f). For instance, this analysis confirmed the strong agreement of identified candidate regulators of M1 (ybx1) and M3 (cebpb) across the three species (Supplementary Fig. 7a). mmp9 expression lagged slightly behind in zebrafish compared to some human (Tabula Sapiens and Xie 2021) and mouse (Grieshaber−Bouyer 2020) datasets that contain peripheral neutrophils that were not analyzed in our WKM isolates. Conversely, other genes showed species-specific differences; for instance, tgfbi was expressed earlier in human but later in mouse trajectories compared to zebrafish. Other genes showed a lag (e.g., txnipa) or advance (e.g., amd1) compared to zebrafish. To assess the similarity of maturation trajectories at species-level, we calculated the mean cross-correlation coefficient per dataset for each module (Fig. 7e, f, Supplementary Fig. 7b) and used hierarchical clustering to group datasets with similar maturation dynamics (Supplementary Fig. 7c−e). We found that the range of cross-correlation lags for M1ortho and M2ortho was smaller than for M3ortho, for which differences between all datasets increased. Taken together, these results indicate a strong conservation (across all species) of early neutrophil maturation, while differences arise at late maturation stages.
As an external validation, we sought to compare our orthologous gene modules (M1ortho-M3ortho) to frequently used gene signatures for human immature neutrophils (HAY_BONE_MARROW_IMMATURE_NEUTROPHIL; M39200)55. Surprisingly, the majority of putative “immature neutrophil” genes were attributable to late modules (M2ortho, M3ortho) of zebrafish neutrophil maturation (Supplementary Fig. 8a), but also to late time points of human in vitro neutrophil differentiation (Supplementary Fig. 8b), highlighting the necessity for alternative gene signatures of different maturation grades. Our cross-species comparison allowed us to define a pan-species gene signature of neutrophil maturation (absolute cross-correlation lag <= 50; M1pan = 304, M2pan = 176, M3pan = 122; Supplementary Data 1, 6; see Cross-species integration and comparison for details).
Finally, we went on to show the utility of the pan-species neutrophil maturation signature by using it to infer the maturation stage of neutrophils in heterogeneous tissues from bulk RNA sequencing data. To this end, we analyzed metastatic neuroblastoma samples56. Neuroblastoma is a childhood cancer derived from the sympathetic nervous system that frequently disseminates into the BM. Comparison of BM with (n = 17 datasets) and without tumor cell infiltration (n = 21 controls) thus presents an in vivo test case to examine our gene modules. Single-sample Gene Set Enrichment Analysis (ssGSEA)57 indicated a differential enrichment of early modules (M1pan and M2pan) in control samples opposed to an enrichment of the mature module M3pan in tumor-infiltrated samples (Fig. 8a, Supplementary Fig. 9a). For validation we scored the percentage of segmented neutrophils on BM cytospins from patients with localized (n = 9) and infiltrated (n = 12) neuroblastoma by imaging mass cytometry (IMC; Iridium-intercalator = nuclear; CD15 = granulocytic; Fig. 8b, c, Supplementary Fig. 9b). This confirmed a significant increase of segmented neutrophils in the metastatic group compared to control (Unpaired t test; P = 0.025).

a Application of the pan-species neutrophil maturation signatures (M1pan, M2pan, M3pan) derived from maturing zebrafish neutrophils (see Methods for details on signature definition) on bulk RNA sequencing data from 38 bone marrow (NB-BM) samples of patients with metastatic (n = 17, infiltrated) and localized (n = 21, control) neuroblastoma. The heatmap displays the ssGSEA57 score of the maturation signatures on RNA-seq samples from 38 neuroblastoma patients with (n = 17) and without (n = 21) tumor cell bone marrow infiltration. b, c Assessment of neutrophil maturation based on nuclear morphology in BM cytospin samples from patients with metastatic (n = 12, infiltrated) and localized (n = 9, control) neuroblastoma. Samples were stained with DNA-intercalator Iridium and anti-CD15-Bi209, analyzed with IMC and counted blinded. c Analysis of the frequency of segmented neutrophils in control NB-BM (mean = 3.48%; range = 0.8−10.1%) versus infiltrated (mean = 13.48%; range 2−41.3%, two-tailed unpaired t test, P = 0.025).
In summary, we established a comprehensive cross-species transcriptome comparison of neutrophil maturation, suggesting a high degree of conservation between zebrafish and mammalian models. The pan-species gene signature derived from this comparison will present a valuable and robust alternative to existing gene signatures of neutrophil maturation and is applicable for examination of human samples.
Discussion
In this study, we generated lysC:CFP/mmp9:Citrine transgenic zebrafish, which enabled us to identify and study neutrophils of different maturation grades non-invasively in a living organism. The importance of neutrophils is underlined by their long evolutionary conservation: Granular cells are present already in invertebrates such as lancelets (Branchiostoma), and myeloperoxidase is even produced by invertebrates58. However, to use animal models for human disease, it is crucial to first delineate similarities and differences between species. Our transgenic zebrafish made it possible to separate neutrophils into populations that differed in maturation-dependent neutrophil functions (e.g., phagocytic capability, recruitment towards bacterial infections, and the ability to interact with oncogene-expressing cells). As antibody markers for maturation states are not available in zebrafish, our Mmp9-reporter was thus instrumental in dissecting these different cell populations. Notably, among myeloid cells, Mmp9 expression was almost completely restricted to neutrophils (LysC+ Mpeg+ and LysC+ Mpeg-) in our model and only found in a small population of steady-state macrophages (LysC- Mpeg+) at low levels. In interaction with malignant cells, macrophages might start expressing Mmp9 as observed59,60.
Powered by the new model, our subsequent scRNA-seq analyses revealed a continuous neutrophil maturation trajectory in zebrafish akin to what has been observed in mice12. We found that maturing zebrafish neutrophils went through multiple stages equivalent to those in mammals (Fig. 9), including also cells mimicking early, unipotent progenitors described recently in mice13,50 and human48,49. All three species opted for similar gene expression programs at these early stages, while transcriptomic differences became more pronounced with increasing maturity. For instance, the expression of transcription factors ybx1 and cebpb were particularly well synchronized across species, while the expression of tgfbi diverged. As in mice we found late maturation under the control of the transcription factor cebpb and associated with inflammatory pathways and increasing levels of il1b and csf3r suggesting a pro-inflammatory state of neutrophils awaiting release from the marrow12. Additionally, we also discovered that cebpb expression was required to drive tertiary granule genes such as mmp9 and fcer1gl61 and for the generation of Mmp9+ neutrophils, suggesting a role beyond emergency granulopoiesis46,47 that may be underappreciated in other organisms. Another family member, cebpe is known to regulate early neutrophil maturation in mice9 and its putative zebrafish orthologue cebp162 has been reported to regulate granule genes lyz and srgn in zebrafish63. Indeed, lyz and srgn were also expressed in early neutrophil maturation in our study (Fig. 4) and cebp1 itself was weakly expressed throughout maturation. Given that there were no strong changes in cebp1 expression during maturation, our algorithms prioritized other well-known hematopoietic regulators such as myb64, myca, and ybx145 as driving the early phase of development in our analyses.

Association of gene expression with modules and phases as determined by scRNA-seq. Granule stage and morphology were assumed by comparing gene expression (as in Fig. 4d, 7a, Supplementary Data 2) with previously published data for neutrophils. TF = transcription factor; module = M; P = phase; KM = kidney marrow; GMP = granulocyte-monocyte progenitor; MB = myeloblast; PM = promyelocyte; MC = myelocyte; MM = metamyelocyte; BC = band cell; SC = segmented cell.
Our cross-species comparison identified a subset of genes with highly synchronous expression dynamics in zebrafish, mice, and humans (M1pan-M3pan). We found this conserved gene signature helpful in interpreting neutrophil states also in human bulk RNA sequencing data, as illustrated here using data from BM metastases of pediatric neuroblastoma patients56 (Supplementary Fig. 17). Intriguingly, we detected the mature neutrophil signature (M3pan) in patients with disseminated tumor cells. We speculate that this enrichment in mature neutrophils could either result from prolonged retention of mature TANs in the BM or increased recruitment of peripheral neutrophils in response to metastasizing tumor cells. The presence of TANs in BM metastases has been associated with a pro-inflammatory and concurrently immuno-suppressive environment in the BM metastatic niche56,65. In the future, our signature could be applied to relate neutrophil maturation grade to disease progression or outcome. However, care must be taken when performing such analysis, as our signatures were designed to identify neutrophil maturation stages and not to distinguish between neutrophils and other cell types. Other cells, in particular closely related myeloid phagocytes, utilize similar genes in their maturation and may influence analyses of bulk tissues. (Please refer to our Supplementary Figs. 10−15 for additional assessments of the specificity and sensitivity of the pan-species neutrophil maturation signatures.) Therefore, we advise validation of putative differences in neutrophil maturation states using orthogonal assays (e.g., using imaging as illustrated in our study).
In conclusion, the strong homology of neutrophil maturation supports the translatability of zebrafish models to study neutrophil biology. The combination of live imaging and transcriptomic approaches in zebrafish will further enable the dissection of the role of immature and mature neutrophils in the tumor microenvironment and the consequences of their interactions with tumor cells.
Methods
Experimental model and subject details
Zebrafish model and zebrafish transgenic lines
Zebrafish (Danio rerio) were maintained at standard conditions66 according to the guidelines of the local authorities (Vienna Magistrat MA58) under licenses GZ:565304/2014/6 and GZ:534619/2014/4 in a research fish facility (Tecniplast, Italy). Experiments on larval zebrafish were carried out at developmental stages, which do not require ethical approval. Single-cell RNAseq or flow cytometry was performed on kidney marrow cells from adult zebrafish post mortem.
Larvae were kept in egg medium with 20 mg/l phenylthiourea (PTU) (Merck) from 22 hpf to avoid pigmentation. Tricaine was used as an anesthetic.
The following transgenic lines were used: Tg(lysC:CFP-NTR)vi002,Tg(lysC:dsRed)nz50Tg, Tg(mpeg1:mCherry)gl23, Et(kita:GAL4)hzm1, Tg(UAS:EGFP-HRAS_G12V)io006, Tg(HRAS_G12V:UAS:CFP)vi004, Tg(BACmmp9:Citrine-CAAX)vi003. Transgenic lines were on a AB* (Tg(lysC:CFP-NTR)vi002,Tg(lysC:dsRed)nz50Tg, Tg(mpeg1:mCherry)gl23, Et(kita:GAL4)hzm1, Tg(UAS:EGFP-HRAS_G12V)io006, Tg(HRAS_G12V:UAS:CFP)vi004) or AB*x SAT mixed background (Tg(BACmmp9:Citrine-CAAX)vi003). Tg(BACmmp9:Citrine-CAAX)vi003 fish (abbreviated mmp9:Citrine) were generated by BAC transgenesis according to published protocols67,68. In short: Identity of annotated BAC CH211-269M15 (105.8 kb) containing mmp9 gene and regulatory regions (BACPAC Resources) was confirmed by sequencing, recombineered with iTol2 sites and with membrane-targeted fluorescent Citrine-CAAX DNA inserted at the start codon of mmp9. BAC DNA (67 ng/µl) was micro-injected into fertilized zebrafish eggs together with Tol2 transposase mRNA as previously described69. pDEST-lysC:CFPNTR (#54) was generated by gateway recombination of p5-lysC (#31), pENTR1A-CFPNTR (#51) and pDEST-Tol2pA2 (kind gift of Chi-Bin Chien). Tg(HRAS_G12V:UAS:CFP) plasmid was constructed and kindly provided by the Mione lab. The transgenesis constructs were injected into fertilized zebrafish eggs at 25 ng/µl together with 25 ng/µl Tol2 RNA. Injected zebrafish were grown up to adulthood and screened for germline transmission.
Generation of Cebpb CRISPR-Cas9 mutant zebrafish
Cebpbmut vi006 was generated by microinjection of Cas9 RNP complexes into one-cell stage eggs from Tg(lysC:CFP-NTR)vi002x Tg(BACmmp9:Citrine-CAAX)vi003 crosses. RNPs were assembled by first incubating Alt-R tracrRNA ATTO550 (IDT) (3 µM) with each of ALT-R crRNA dr.cebpb_AB and crRNA_ dr.cebpb AA (1.5 µM each) in Duplex buffer (IDT) for 5 min at 95 °C followed by mixing equal amounts of assembled gRNA and of Cas9 (0.5 µg/ml) at 37 °C for 10 min. F0 larvae were checked for mutation efficiency by PCR using genotyping primers cebpb_fw (CTGGGCAGGCAACCTATCAC) and cebpb_rev (CATTTTACCGCCCGCTTGAG) and T7 mismatch assays. Adult F0 and F1 fish were typed by PCR and sequencing using the aforementioned primers (Supplementary Figs. 6b, 18).
Human research participants
In this study bone marrow aspirates from children and adolescents (age at diagnosis 0−18 years) with neuroblastoma or ganglioneuroma were analysed. The cohort used for imaging (n = 21) contained samples from 10 female and 11 male patients. Additionally, we collected neuroblastoma bulk RNA-seq data for 38 samples (17 with and 21 without bone marrow infiltration) from accession GSE172184 [Gene Expression Omnibus]”. Sex was not included in the parameters for the data analysis results presented.
Patients were recruited previously: Patients with high-risk metastatic (stage M) neuroblastoma were enrolled in the SIOPEN/HR-NBL-1 trial (NCT01704716) according to the trial’s inclusion and exclusion criteria. Samples were available as left over material from diagnostic procedures and selected for this study based on their availability in the CCRI Biobank. Ethics oversight by the Ethics committee of the Medical University of Vienna, Austria.
Method details
Flow cytometry and FACS
Single-cell suspensions from adult zebrafish (3 months) spleens or kidneys were prepared by mashing or pipetting, respectively, and pipetting through a cell strainer. To isolate cells from larvae, they were immersed in 10 mM DTT (Merck) in E3 to remove mucus, then digested with Liberase Blendzyme TM at 1.1 μ/ml and Dnase I at 40 μg/ml (Merck) in HBSS under constant shaking at 37 °C for 40 min. 7-AAD (Invitrogen) was used as a live stain. Samples were run on an LSRFortessa cytometer (Becton-Dickinson) with BD FACSDiva Software v9.0 and analyzed in FlowJo_v10.8.1 or sorted using a FACSAria Fusion.
Wounding, injection of bacteria and phagocytosis assay
Zebrafish larvae at 2 dpf were wounded in the ventral fin area using a scalpel blade or glass needle and were imaged from 30 mpi (minutes post injury) to 150 mpi on a Leica SP8 confocal microscope. Tracking of LysC+ Mmp9-, INT or HI neutrophils was performed using the ImageJ Fiji TrackMate plugin;70 The trackmate feature linearity of forward progression = mean straight line speed/ mean speed, where the mean straight line speed is net distance/total track time.
E. coli labeled with mCherry (pZS*12-mCherry-KANr: PRlambda-mCherry in pZSstar, SC101* ORI)71 were injected at OD = 2 into the caudal vein area or otic vesicle as previously described72. To study the phagocytic capacity of neutrophils, cells from Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 larvae were analyzed after 6 hpi (hours post infection) by flow cytometry as described above or imaged using confocal microscopy. For ROS detection E.coli-mCherry OD = 3 were injected together with CellROX Deep red (Thermo Fisher) at 5 µM into the otic vesicle of 3 dpf larvae.
Morpholino and mRNA micro-injection
We used a previously published C/ebp-β translation blocking morpholino (5′-GATCTTAACACCCGCCGGATTGCG-3′) and a negative control morpholino (5′-CCTCTTACCTCAGTTACAATTTATA-3′) from Gene Tools LCC46,73. Efficacious doses (1 pmole and 0.5 pmole, respectively) were determined empirically. Full-length cebpb mRNA (50 pg), prepared by mMessage mMachine T7 Ultra transcription (ThermoFisher) or morpholinos were injected into one to two-cell stage embryos from Tg(lysC:CFP-NTR)vi002x Tg(BACmmp9:Citrine-CAAX)vi003 crosses.
Cytospin, Electron microscopy and Sudan Black staining
Kidney marrow cells of adult Tg(lysC:dsRed)nz50Tg/ Tg(BACmmp9:Citrine-CAAX)vi003 fish (aged three months) were FACS sorted into Mmp9+LysC+ or Mmp9-LysC+ fractions and spun in a cytocentrifuge (Fisher Scientific) onto slides according to the manufacturer´s instructions and stained using Pappenheim solution (Merck).
For TEM Mmp9HILysC+ or Mmp9-LysC+ cells were FACS sorted from adult kidneys of Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 fish and embedded in liquid low melting agarose (3% in PBS), fixed with 2.5% glutaraldehyde in 0.1 M cacodylate buffer containing 5 mM CaCl2, post-fixation in 1% aqueous OsO4 and embedded in Epon812 resin. Semi-thin sections were stained with toluidine blue and images were recorded using a 100x/N.A. 1.4 lens. Ultrathin sections were contrasted with uranyl acetate and lead citrate and imaged using a FEI Tecnai G2 20 TEM. Measurements were performed using Fiji software.
For analysis of granulated neutrophils whole larvae Tg(lysC:CFP-NTR)vi002/ Tg(BACmmp9:Citrine-CAAX)vi003 were PTU treated, and stained at 2 dpf with Sudan Black B (Merck) according to a published protocol74 and imaged on a confocal microscope.
IMC analysis of neuroblastoma BM cytospin preparations
Archived cytospin preparations of human BM aspirates from patients with localized and metastatic neuroblastoma (leftover samples from standard-of-care diagnostic procedures) have been obtained after institutional review board approval and informed consent by patients or their guardians from the CCRI Biobank (EK1853/2016). Inclusion criteria: male and female patients aged 0 to 18 years with clinically, histologically, and biologically confirmed high-risk neuroblastoma with bone marrow metastasis or ganglioneuroblastom, ganglioneuroma or localized neuroblastoma with no detectable BM metastasis. Ethical approval for the use of bone marrow aspirates for imaging and clinical data (histological diagnosis) was obtained from the local institutional review board of the Medical University of Vienna (EK1216/2018, EK1224/2020). Patients were not compensated for study participation. Patients’ sex was recorded, but not used to disaggregate the data, due to the limited number of patients and lack of statistical power. Gender was not assessed since NB is a tumor of early childhood.
Samples were thawed for 15 min at RT, fixed in 4% PFA (Carl Roth) at 4 °C for 30 min, washed twice in TBS and then blocked with 2% BSA (Carl Roth) and 0.1% Tween-20 (Merck) in TBS at RT for 1 h. Samples were incubated overnight at 4 °C with diluted CD15 antibody (BioLegend; catalog number: 301902; clone: HI98) at 5 µg/ml in 0.1% Tween-20 in TBS. On the following day, samples were stained with the DNA intercalator Iridium (Fluidigm) and then dried with pressured air before IMC measurement with the Hyperion imaging system (Fluidigm) and CyTOF Software v7.
Morphological assessment was subsequently performed in the image analysis software QuPath75.
Nuclear morphology of CD15+ cells was assessed, and cells were classified as segmented cells, band cells or metamyelocytes and myelocytes (3 ROI/patient; n = 248−330 cells/patient). Cells were counted using the QuPath counting tool.
Quantitative real-time PCR
RNA from FACS sorted cells was prepared using RNeasy Micro kit (Qiagen) and transcribed with High Capacity cDNA transcription kit. qPCR was performed using Maxima SYBR green mix (Applied Biosystems) and gene-specific primers (Supplementary Table 1 in Supplementary Information) on a 7500 Fast real-time PCR machine (Applied Biosystems).
Imaging
For imaging larvae were pre-treated in E3/PTU, anaesthetized with 0.02% tricaine and embedded in 1.2% low-melting agarose (Merck) on glass bottom dishes (D35-14-1.5-NJ, Cellvis, USA) as described previously76. Images were acquired with a Leica TCS SP8 WLL microscope (HCX PL APO CS 10x/0.40 DRY objective or HC PL APO CS2 40x/1.10 WATER objective). Maximum projections were performed in the Leica LASX 3.7.0.20979 software. Images were rendered using Leica LAS software, Photoshop CS6 (Adobe) and Fiji ImageJ, e.g., for preparing cell tracks. Leukocyte units were analyzed according to77.
Single-cell RNA sequencing
Kidney marrow from 2 adult, six-month old, male fish was isolated, labeled using lipid-tagged indices as published (MULTI-seq)78. In short: each kidney marrow was split into four portions, labeled with lipid anchor plus individual barcode solution (2 µM) for 5 min on ice and then incubated with lipid co-anchor (2 µM) for 5 min on ice. Each cell portion was individually FACS-sorted to obtain one population (Mmp9 NO, INT, HI or WKM). All cells were gated on live gate, for WKM debris was excluded in a FSC/SSC gate, and Mmp9 NO, INT, HI were gated on LysC:CFP positivity and different levels of Mmp9:Citrine-CAAX expression (Supplementary Fig. 3a). For multiplexing, the two WKM populations were sorted into one well (A2) and processed together (20,000 cells each; total: 40,000). Similarly, the six LysC+ populations were sorted into one well (A1) and processed together (total: 41,000 cells). Here, cell numbers ranged from 1700 to 10,000 cells/population.
Single cell suspensions were immediately subjected to scRNA-seq using the Chromium Single Cell Controller and Single Cell 3’ Library & Gel Bead Kit v3.1 (10x Genomics, Pleasanton, CA), according to the manufacturer’s protocols (10x Genomics). Sequencing was performed at the Biomedical Sequencing Facility of the CeMM Research Center for Molecular Medicine of the Austrian Academy of Sciences (Vienna, Austria) using the Illumina NovaSeq platform and the 50 bp paired-end configuration with adapted read-lengths for both forward and reverse reads.
Bioinformatics analyses
Read processing, quality control, and normalization
We used the CellRanger v3.1.0 software (10x Genomics) for cell demultiplexing and alignment to GRCz11-3.1.0 zebrafish reference transcriptome that had been expanded to include the sequences of reporter genes (Citrine and CFPNTR, sequences from snapgene.com). The R statistics software v4.0.3 was used to carry out the entire analysis workflow. Processed data was loaded into R for sample demultiplexing (package deMULTIplex78 v1.0.2; cell classification step was stopped when negative cell number dropped below 100). Only cells classified as singlets or negatives were retained, while doublets were excluded. Next, we loaded the counts into Seurat v4.0.279 and performed quality control by only including features detected in at least 20 cells (n = 12,619 out of 25,109 features retained), and cells with a minimum of 500 features, mitochondrial reads proportion less than 10%, and doublet score, calculated using function doubletCells (package scran80 v1.18.3;default parameters), below 3 (n = 19,373 out of 28,534 cells retained). Remaining negative cells (that is, cells without an assigned sample label; n = 4723) from each sequencing run were reclassified separately using Linear Discriminate Analysis (function lda from the package MASS v7.3-53; default parameters), only relabeling negative cells with a posterior probability >= 0.95 (n = 3500 cells), yielding a final dataset of 18,150 cells. Cell cycle phase inference was done based on expression of G2M and S phase markers saved in Seurat package and using CellCycleScoring function from Seurat package. Cells with high level of expression of either G2M or S markers, as they are anti-correlated, are likely to be cycling cells. Next, we normalized raw read counts using SCTransform81 v0.3.2 and integrated using Harmony v1.082 (default parameters) the batch effect of different sequencing runs and fish. We performed low-dimensional projection using UMAP based on the top 30 Harmony components. Reference-based cell annotation using Seurat79 v4.0.2 was carried out by mapping our data to two atlases of hematopoiesis in zebrafish24,38 (preprocessing as described above) according to the workflow recommended by the developers (https://satijalab.org/seurat/articles/multimodal_reference_mapping.html; accessed on 18-Nov-2022). Cells identified as neutrophils in both atlases (n = 15,876) were selected for downstream analysis. Finally, after subsetting neutrophils, we excluded hemoglobin-related genes (based on the following regular expression vector “^hb[ba]|si:ch211.5k11.8”) and features detected in less than 20 neutrophils to minimize the effect of other cell types on the downstream analysis of neutrophils.
Inference of neutrophil maturation trajectories, associated genes, and top regulators
Continuous maturation trajectories were inferred using Slingshot39 v1.8.0 (default parameters) based on the top two Harmony components of the dataset82. We compared Slingshot output to three other methods: Component 1, TSCAN (default parameters), and Monocle3 (use_partition = FALSE, verbose = FALSE, close_loop = FALSE)83,84. The first, cluster-free approach uses Harmony 1 coordinate values (akin to principal components), while the other two approaches use pre-defined clusters as inputs (Supplementary Fig. 4e). We then discretized the trajectory into 100 bins by averaging overlapping cells at each bin. We used the functions fitGAM (package tradeSeq40 v1.4.0, parameters: knots = 6, cellWeights = rep(1, #genes)) and associationTest (package tradeSeq40 v1.4.0) to carry out differential expression analysis along the inferred trajectory using pseudotime values from Slingshot. In more detail, first, expression patterns of each gene were modeled as non-linear functions of the pseudotime using generalized additive model (GAM). Second, genes were tested (two-tailed) for the equality of all smoother coefficients within the inferred lineage. We excluded mitochondrial and ribosomal genes, as well as genes with less than 5 cells with 3 or more reads. Genes that passed the cutoff (BH (Benjamini Hochberg)-adjusted P < 0.05) were put in descending order based on Wald-statistic and the top 1500 genes were selected as maturation-associated genes for further analysis (Supplementary Data 1). To define gene modules and maturation phases, we used the binned expression of these maturation-associated genes, calculated the DTW distance using function dist (package proxy v0.4-25; method = “dtw”), and used hierarchical clustering (parameters: method = “ward.D2”). The resulting dendrograms were cut into three gene modules and four maturation phases, which was the optimal number based on the lowest Kelley-Gardner-Sutcliffe penalty using function kgs (package maptree v1.4-7; default parameters) (Supplementary Fig. 16). To interpret and characterize the genes in each maturation module, we used the function hyper (package hyper41 v2.0.1; default parameters) with five datasets (CP:KEGG, GO:BP, CP:WIKIPATHWAYS,CP:REACTOME, HALLMARK) retrieved from Molecular Signatures Database (MSigDB) using msigdbr function (package msigdbr v7.5.1; species = “Danio rerio”), to carry out over-representation analysis (upper one-tailed hypergeometric test: FDR (false discovery rate)-adjusted p < 0.1; Supplementary Data 3), selected the top five genesets with lowest FDR, and plotted their ratio of overlap with the gene modules. To identify the transcription factors (“top regulators”) that best explain the variability in the expression patterns of each gene module, we collected a list of zebrafish transcription factors based on both AnimalTFDB3 database (date of retrieval: 21 June 2021)85 and manual selection from literature6,8,12,14, and calculated the DTW distance between each transcription factor and all target genes85. DTW allows to account for the expected lag between transcription factor expression and target gene activation. Next, we transformed the DTW distance dil for gene i and transcription factorlinto a “scaled DTW similarity” sil, as follows: sil = 1 – (dil / max(di)). Finally, to rank putative regulators per module, we used the function dunn_test (package rstatix v0.6.0; default parameters: p.adjust.method = “holm”) to run a post-hoc Kruskal-Wallis test (two-tailed) of the distribution of similarity values between transcription factors and putative target genes across the three modules (Supplementary Data 5).
Cross-species integration and comparison
For maturation staging of the zebrafish phases, we downloaded relevant datasets13,48,49,50 (see Data Availability section) and selected neutrophil-related samples. Samples were then mean-aggregated per maturation stage followed by quantile normalization across all datasets. Specifically, genes were first ranked per column and sorted ascendingly. Next, we replaced each gene with its mean rank and restored the original ranking of genes. This was followed by scaling quantile-normalized genes across the maturation stages of each dataset. We then used hierarchical clustering (parameters: method = “ward.D2”) to group maturation stages across all datasets based on the Pearson distance. Finally, each stage was assigned a continuous score reflecting its maturation order (earliest = 0 and latest = 1) in the corresponding study and the leaves of the resulting hierarchical clustering tree were then reordered under the constraints of the tree using function reorder (package: stats, parameters: agglo.FUN = mean).
For gene-level alignment, scRNA-seq10,12,14,52,53,54 (see Data Availability section) data were preprocessed following the same approach described under Read processing, quality control, and normalization section, with the following exceptions: We did not carry out further quality control filtration on data from Suo 202252 (minimum of 501 features and 2001 UMI counts per cell) and Tabula Sapiens53 (minimum of 200 features and 2500 UMI counts per cell). In the case of neutrotime12 model, we used the pre-defined embedding provided by the authors via ImmGen single cell explorer (https://singlecell.broadinstitute.org/single_cell/study/SCP1019/ly6-neutrophils-from-bone-marrow-blood-spleen?scpbr=immunological-genome-project#study-download).
We then selected only healthy bone-marrow neutrophils (using provided cell annotations) (Supplementary Data 8). Subsequently, we carried out trajectory inference using Slingshot based on the top two PCA components and discretization into 100 bins, as explained above. Next, we performed homology mapping across zebrafish, mouse, and human using getLDS function (package biomaRt v2.46.3; default parameters). Common genes from human and mouse were renamed to the corresponding homologous zebrafish gene using a modified version of RenameGenesSeurat function (package Seurat.utils v1.4.7; https://doi.org/10.5281/zenodo.7228243). Cases of 1-to-many homology were resolved by mapping the genes to the most expressed homolog based on the attribute “detection_rate” from the sctransform model. We kept only the set of genes that are common across all datasets, scaled each of the datasets separately, and ordered the genes based on their expression pattern across all datasets using the seriate function (seriation 1.3.1; method = “PCA”) and the corresponding module. Finally, we used cross-correlation to align the gene expression pattern along maturation trajectory in mouse and human datasets onto zebrafish using the ccf function (stats v4.0.3; default parameters) and recorded the mean lag at which highest cross-correlation was achieved among mouse and human datasets per gene (Supplementary Data 7).
The pan-species neutrophil maturation signatures (M1pan, M2pan, M3pan) were defined based on two inclusion criteria: First, consistent expression pattern between human and zebrafish ( | maximum cross-correlation lag | <= 50). Second, we excluded unique known markers of other cell types (Additional file 5 from xCell86). We then obtained normalized count data from neuroblastoma bulk RNA-seq56 and used this signature to carry out single-sample gene set enrichment analysis (ssGSEA) using function ssgsea (package corto57 v1.1.11;default parameters). To examine the specificity of neutrophil maturation modules, we calculated module scores for M1pan and M3pan across different cell types and cell states. First, we used two zebrafish scRNA-seq datasets24,38 and computed module score using AddModuleScore function (package Seurat v4.0.2). Second, we used two human scRNA-seq datasets52,53 that we aggregated into pseudobulk samples of cell types across tissues (>= least 50 neutrophils cells) and one bulk RNA-seq dataset51. The human bulk/pseudobulk datasets were then normalized using vst function (package DESeq2 v 1.30.0)87 and a module score was computed using gsva function (package GSVA v 1.38.2; method = “ssgsea”). Sensitivity analysis was carried out by mixing cells from the three neutrophil maturation stages (promyelocyte, myelocyte, neutrophil) at defined ratios (0%, …, 100%) with stromal cells from different tissues52 to create pseudo-bulk samples with known neutrophil contributions. The mixing was done via random sampling of cells to a sum of 1000 cells and the process was repeated 20 times. Module scores were computed using gsva function (package GSVA v 1.38.2; method = “ssgsea”), and the median value presented in Supplementary Fig. 15.
Quantification and statistical analysis
GraphPad Prism Version 8.3.0 was used for statistical analysis. Statistical details of experiments can be found in the results, methods and figure legends.
Resource availability
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Martin Distel (martin.distel@ccri.at). Raw data underlying the figures will be shared upon reasonable request.
Material availability
Transgenic zebrafish lines generated in this study are deposited in the European Zebrafish Resource Center (EZRC) (https://www.ezrc.kit.edu/), Tg(lyz:CFP-NTR)vi002 #37873 and TgBAC(mmp9:Citrine-CAAX)vi003 #37874.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The single-cell RNA sequencing data generated in this study was deposited in the Gene Expression Omnibus (GEO) database under accession code GSE252788. The two samples generated are accessible under GSM8007849 (MF317_A1_GEX_zebrafish_multiseq) and GSM8007850 (MF317_A2_GEX_zebrafish_multiseq). We made use of the following publicly available datasets in our study: Single-cell RNA-seq: E-GEOD-100911, E-MTAB-5530, GSE137539, GSE165276, GSE149938, GSE142754, fetal-immune (https://developmental.cellatlas.io/fetal-immune), Tabula sapiens (https://cellxgene.cziscience.com/collections/e5f58829-1a66-40b5-a624-9046778e74f5), Murine neutropenia (https://www.synapse.org/#!Synapse:syn16816566) ; Bulk RNA-seq: GSE79044, GSE109467, GSE172184 neuroblastoma data provided by the authors (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE172184), GSE153263, GSE151682, GSE175880 ; Others: neutrotime model (https://singlecell.broadinstitute.org/single_cell/study/SCP1019/ly6-neutrophils-from-bone-marrow-blood-spleen?scpbr=immunological-genome-project), AnimalTFDB3.0-zebrafish (http://bioinfo.life.hust.edu.cn/static/AnimalTFDB3/download/Danio_rerio_TF), xCell signatures Additional file 5 (https://static-content.springer.com/esm/art%3A10.1186%2Fs13059-017-1349-1/MediaObjects/13059_2017_1349_MOESM5_ESM.xlsx), additional metadata of GSE109467 was kindly provided by the authors. Source data are provided with this paper.
Code availability
Computer code used for the data analysis in this paper can be accessed on our GitHub page https://github.com/cancerbits/Kirchberger_Shoeb2024_neut and via Zenodo.
References
Mantovani, A., Cassatella, M. A., Costantini, C. & Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 11, 519–531 (2011).
Nathan, C. Neutrophils and immunity: challenges and opportunities. Nat. Rev. Immunol. 6, 173–182 (2006).
Taschner, S. et al. Down-regulation of RXRalpha expression is essential for neutrophil development from granulocyte/monocyte progenitors. Blood 109, 971–979 (2007).
Cowland, J. B. & Borregaard, N. Granulopoiesis and granules of human neutrophils. Immunol. Rev. 273, 11–28 (2016).
Borregaard, N., Sorensen, O. E. & Theilgaard-Monch, K. Neutrophil granules: a library of innate immunity proteins. Trends Immunol. 28, 340–345 (2007).
Coffelt, S. B., Wellenstein, M. D. & de Visser, K. E. Neutrophils in cancer: neutral no more. Nat. Rev. Cancer 16, 431–446 (2016).
Faurschou, M. & Borregaard, N. Neutrophil granules and secretory vesicles in inflammation. Microbes Infect. 5, 1317–1327 (2003).
Ai, Z. & Udalova, I. A. Transcriptional regulation of neutrophil differentiation and function during inflammation. J. Leukoc. Biol. 107, 419–430 (2020).
Gombart, A. F. et al. Regulation of neutrophil and eosinophil secondary granule gene expression by transcription factors C/EBP epsilon and PU.1. Blood 101, 3265–3273 (2003).
Ballesteros, I. et al. Co-option of neutrophil fates by tissue environments. Cell 183, 1282–1297.e1218 (2020).
Evrard, M. et al. Developmental analysis of bone marrow neutrophils reveals populations specialized in expansion, trafficking, and effector functions. Immunity 48, 364–379.e368 (2018).
Grieshaber-Bouyer, R. et al. The neutrotime transcriptional signature defines a single continuum of neutrophils across biological compartments. Nat. Commun. 12, 2856 (2021).
Kwok, I. et al. Combinatorial single-cell analyses of granulocyte-monocyte progenitor heterogeneity reveals an early uni-potent neutrophil progenitor. Immunity 53, 303–318.e305 (2020).
Xie, X. et al. Single-cell transcriptome profiling reveals neutrophil heterogeneity in homeostasis and infection. Nat. Immunol. 21, 1119–1133 (2020).
Zhu, Y. P. et al. Identification of an early unipotent neutrophil progenitor with pro-tumoral activity in mouse and human bone marrow. Cell Rep. 24, 2329–2341.e2328 (2018).
Mackey, J. B. G., Coffelt, S. B. & Carlin, L. M. Neutrophil maturity in cancer. Front. Immunol. 10, 1912 (2019).
Quail, D. F. et al. Neutrophil phenotypes and functions in cancer: a consensus statement. J. Exp. Med. 219, e20220011 (2022).
Sagiv, J. Y. et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 10, 562–573 (2015).
Fridlender, Z. G. et al. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 16, 183–194 (2009).
Jablonska, J., Leschner, S., Westphal, K., Lienenklaus, S. & Weiss, S. Neutrophils responsive to endogenous IFN-beta regulate tumor angiogenesis and growth in a mouse tumor model. J. Clin. Investig. 120, 1151–1164 (2010).
Feng, Y. & Martin, P. Imaging innate immune responses at tumour initiation: new insights from fish and flies. Nat. Rev. Cancer 15, 556–562 (2015).
Henry, K. M., Loynes, C. A., Whyte, M. K. & Renshaw, S. A. Zebrafish as a model for the study of neutrophil biology. J. Leukoc. Biol. 94, 633–642 (2013).
Lieschke, G. J., Oates, A. C., Crowhurst, M. O., Ward, A. C. & Layton, J. E. Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood 98, 3087–3096 (2001).
Tang, Q. et al. Dissecting hematopoietic and renal cell heterogeneity in adult zebrafish at single-cell resolution using RNA sequencing. J. Exp. Med. 214, 2875–2887 (2017).
Langenau, D. M. et al. Myc-induced T cell leukemia in transgenic zebrafish. Science 299, 887–890 (2003).
Wigerblad, G. et al. Single-cell analysis reveals the range of transcriptional states of circulating human neutrophils. J. Immunol. 209, 772–782 (2022).
Dewald, B., Bretz, U. & Baggiolini, M. Release of gelatinase from a novel secretory compartment of human neutrophils. J. Clin. Investig. 70, 518–525 (1982).
Yoong, S. et al. Characterization of the zebrafish matrix metalloproteinase 9 gene and its developmental expression pattern. Gene Expr. Patterns 7, 39–46 (2007).
Ellett, F., Pase, L., Hayman, J. W., Andrianopoulos, A. & Lieschke, G. J. mpeg1 promoter transgenes direct macrophage-lineage expression in zebrafish. Blood 117, e49–e56 (2011).
Garcia-Lopez, J. P. et al. Ontogenetically distinct neutrophils differ in function and transcriptional profile in zebrafish. Nat. Commun. 14, 4942 (2023).
Le Guyader, D. et al. Origins and unconventional behavior of neutrophils in developing zebrafish. Blood 111, 132–141 (2008).
Barros-Becker, F., Lam, P. Y., Fisher, R. & Huttenlocher, A. Live imaging reveals distinct modes of neutrophil and macrophage migration within interstitial tissues. J. Cell Sci. 130, 3801–3808 (2017).
Borregaard, N., Kjeldsen, L., Lollike, K. & Sengelov, H. Granules and secretory vesicles of the human neutrophil. Clin. Exp. Immunol. 101, 6–9 (1995).
de Oliveira, S., Rosowski, E. E. & Huttenlocher, A. Neutrophil migration in infection and wound repair: going forward in reverse. Nat. Rev. Immunol. 16, 378–391 (2016).
Glasser, L. & Fiederlein, R. L. Functional differentiation of normal human neutrophils. Blood 69, 937–944 (1987).
Santoriello, C. et al. Kita driven expression of oncogenic HRAS leads to early onset and highly penetrant melanoma in zebrafish. PLoS One 5, e15170 (2010).
Feng, Y., Santoriello, C., Mione, M., Hurlstone, A. & Martin, P. Live imaging of innate immune cell sensing of transformed cells in zebrafish larvae: parallels between tumor initiation and wound inflammation. PLoS Biol. 8, e1000562 (2010).
Athanasiadis, E. I. et al. Single-cell RNA-sequencing uncovers transcriptional states and fate decisions in haematopoiesis. Nat. Commun. 8, 2045 (2017).
Street, K. et al. Slingshot: cell lineage and pseudotime inference for single-cell transcriptomics. BMC Genom. 19, 477 (2018).
Van den Berge, K. et al. Trajectory-based differential expression analysis for single-cell sequencing data. Nat. Commun. 11, 1201 (2020).
Federico, A. & Monti, S. hypeR: an R package for geneset enrichment workflows. Bioinformatics 36, 1307–1308 (2020).
Lee, T. C. & Ziff, E. B. Mxi1 is a repressor of the c-Myc promoter and reverses activation by USF. J. Biol. Chem. 274, 595–606 (1999).
Cox, A. G. et al. Selenoprotein H is an essential regulator of redox homeostasis that cooperates with p53 in development and tumorigenesis. Proc. Natl. Acad. Sci. USA 113, E5562–E5571 (2016).
Boespflug, N. D. et al. ATF3 is a novel regulator of mouse neutrophil migration. Blood 123, 2084–2093 (2014).
Lummertz da Rocha, E. et al. CellComm infers cellular crosstalk that drives haematopoietic stem and progenitor cell development. Nat. Cell Biol. 24, 579–589 (2022).
Hall, C. J. et al. Infection-responsive expansion of the hematopoietic stem and progenitor cell compartment in zebrafish is dependent upon inducible nitric oxide. Cell Stem Cell 10, 198–209 (2012).
Hirai, H. et al. C/EBPbeta is required for ‘emergency’ granulopoiesis. Nat. Immunol. 7, 732–739 (2006).
Calzetti, F. et al. CD66b(-)CD64(dim)CD115(-) cells in the human bone marrow represent neutrophil-committed progenitors. Nat. Immunol. 23, 679–691 (2022).
Dinh, H. Q. et al. Coexpression of CD71 and CD117 identifies an early unipotent neutrophil progenitor population in human bone marrow. Immunity 53, 319–334.e316 (2020).
Muench, D. E. et al. Mouse models of neutropenia reveal progenitor-stage-specific defects. Nature 582, 109–114 (2020).
Ramirez, R. N. et al. Dynamic gene regulatory networks of human myeloid differentiation. Cell Syst. 4, 416–429.e413 (2017).
Suo, C. et al. Mapping the developing human immune system across organs. Science 376, eabo0510 (2022).
Tabula Sapiens, C. et al. The Tabula Sapiens: a multiple-organ, single-cell transcriptomic atlas of humans. Science 376, eabl4896 (2022).
Xie, X. et al. Single-cell transcriptomic landscape of human blood cells. Natl. Sci. Rev. 8, nwaa180 (2021).
Hay, S. B., Ferchen, K., Chetal, K., Grimes, H. L. & Salomonis, N. The Human cell Atlas bone marrow single-cell interactive web portal. Exp. Hematol. 68, 51–61 (2018).
Lazic, D. et al. Landscape of bone marrow metastasis in human neuroblastoma unraveled by transcriptomics and deep multiplex imaging. Cancers 13, 4311 (2021).
Mercatelli, D., Lopez-Garcia, G. & Giorgi, F. M. corto: a lightweight R package for gene network inference and master regulator analysis. Bioinformatics 36, 3916–3917 (2020).
Fingerhut, L., Dolz, G. & de Buhr, N. What Is the evolutionary fingerprint in neutrophil granulocytes? Int. J. Mol. Sci. 21, 4523. (2020).
Hiratsuka, S. et al. MMP9 induction by vascular endothelial growth factor receptor-1 is involved in lung-specific metastasis. Cancer Cell 2, 289–300 (2002).
Nozawa, H., Chiu, C. & Hanahan, D. Infiltrating neutrophils mediate the initial angiogenic switch in a mouse model of multistage carcinogenesis. Proc. Natl. Acad. Sci. USA 103, 12493–12498 (2006).
Lominadze, G. et al. Proteomic analysis of human neutrophil granules. Mol. Cell Proteom. 4, 1503–1521 (2005).
Lyons, S. E. et al. Molecular cloning, genetic mapping, and expression analysis of four zebrafish c/ebp genes. Gene 281, 43–51 (2001).
Jin, H. et al. c-Myb acts in parallel and cooperatively with Cebp1 to regulate neutrophil maturation in zebrafish. Blood 128, 415–426 (2016).
Oelgeschlager, M., Nuchprayoon, I., Luscher, B. & Friedman, A. D. C/EBP, c-Myb, and PU.1 cooperate to regulate the neutrophil elastase promoter. Mol. Cell Biol. 16, 4717–4725 (1996).
Fetahu, I. S. et al. Single-cell transcriptomics and epigenomics unravel the role of monocytes in neuroblastoma bone marrow metastasis. Nat. Commun. 14, 3620 (2023).
Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B. & Schilling, T. F. Stages of embryonic development of the zebrafish. Dev. Dyn. 203, 253–310 (1995).
Bussmann, J., Schulte-Merker, S. & Rapid, B. A. C. selection for tol2-mediated transgenesis in zebrafish. Development 138, 4327–4332 (2011).
Suster, M. L., Abe, G., Schouw, A., Kawakami, K. & Transposon-mediated, B. A. C. transgenesis in zebrafish. Nat. Protoc. 6, 1998–2021 (2011).
Mayr, V., Sturtzel, C., Stadler, M., Grissenberger, S. & Distel, M. Fast dynamic in vivo monitoring of Erk activity at single cell resolution in DREKA Zebrafish. Front. Cell Dev. Biol. 6, 111 (2018).
Ershov, D. et al. TrackMate 7: integrating state-of-the-art segmentation algorithms into tracking pipelines. Nat. Methods 19, 829–832 (2022).
Guet, C. C. et al. Minimally invasive determination of mRNA concentration in single living bacteria. Nucleic Acids Res. 36, e73 (2008).
Levraud, J. P., Colucci-Guyon, E., Redd, M. J., Lutfalla, G. & Herbomel, P. In vivo analysis of zebrafish innate immunity. Methods Mol. Biol. 415, 337–363 (2008).
Eisinger, A. L. et al. The adenomatous polyposis coli tumor suppressor gene regulates expression of cyclooxygenase-2 by a mechanism that involves retinoic acid. J. Biol. Chem. 281, 20474–20482 (2006).
Buchan, K. D. et al. A transgenic zebrafish line for in vivo visualisation of neutrophil myeloperoxidase. PLoS One 14, e0215592 (2019).
Bankhead, P. et al. QuPath: open source software for digital pathology image analysis. Sci. Rep. 7, 16878 (2017).
Distel, M. & Koster, R. W. In vivo time-lapse imaging of zebrafish embryonic development. CSH Protoc. 2007, pdb-prot4816 (2007).
Ellett, F. & Lieschke, G. J. Computational quantification of fluorescent leukocyte numbers in zebrafish embryos. Methods Enzymol. 506, 425–435 (2012).
McGinnis, C. S. et al. MULTI-seq: sample multiplexing for single-cell RNA sequencing using lipid-tagged indices. Nat. Methods 16, 619–626 (2019).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e3529 (2021).
Lun, A. T., McCarthy, D. J. & Marioni, J. C. A step-by-step workflow for low-level analysis of single-cell RNA-seq data with Bioconductor. F1000Res 5, 2122 (2016).
Hafemeister, C. & Satija, R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 20, 296 (2019).
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 16, 1289–1296 (2019).
Cao, J. et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 566, 496–502 (2019).
Ji, Z. & Ji, H. TSCAN: Pseudo-time reconstruction and evaluation in single-cell RNA-seq analysis. Nucleic Acids Res. 44, e117 (2016).
Hu, H. et al. AnimalTFDB 3.0: a comprehensive resource for annotation and prediction of animal transcription factors. Nucleic Acids Res. 47, D33–D38 (2018).
Aran, D., Hu, Z. & Butte, A. J. xCell: digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 18, 220 (2017).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Acknowledgements
We would like to thank Dieter Printz for FACS sorting; the Biomedical Sequencing Facility of the CeMM Research Center for Molecular Medicine of the Austrian Academy of Sciences for next-generation sequencing; Cristina Santoriello and Marina Mione (University of Trento) for the kind gift of the HRAS_G12V:UAS:CFP plasmid. We would like to acknowledge the CCRI Biobank and Marie Bernkopf. Peter Repiscak for providing support on pre-processing of NB-BM sequencing data. We would like to thank the members of the Innovative Cancer Models group and Dietmar Herndler-Brandstetter for their insightful comments on the manuscript; Adam Varady for providing illustrations; Ulrike Pötschger for advice on statistical tests. This study was supported by the St. Anna Kinderkrebsforschung (to S.T.M., R.L., F.H. and M.D.), the Austrian Research Promotion Agency (FFG) (project 7940628, Danio4Can to M.D.), a German Academic Exchange Service postdoctoral fellowship and an EMBO fellowship (to M.D.), the Austrian Science Fund (FWF) through grants TAI454 (to F.H. and M.D.), TAI732 (to F.H.), I4162 (ERA-NET/Transcan-2 LIQUIDHOPE; to S.T.M.), P35841 (MAPMET; to S.T.M.), P34152 (to T.L.), P30642 (to C.S.) and the Alex’s Lemonade Stand Foundation for Childhood Cancer 20-17258 (to F.H. and M.D.), and the Swiss Government Excellence Scholarship (to D.L.), and the EC H2020 grant no. 826494 (PRIMAGE; to R.L.), and by the European Commission within the FP7 Framework program (Fungitect-Grant No 602125 to T.L.).
Author information
Authors and Affiliations
Contributions
SK designed and performed experiments, analyzed data and wrote the manuscript. MS performed bioinformatics analysis including collection and preprocessing of public data, and wrote the manuscript. DL performed IMC on neuroblastoma BM samples. KF supported BAC screening and AWW cebpbmut vi006 genotyping. LS performed the 10x Genomics workflow. FR designed the graphical abstract. FR and EB developed new imaging methods. RL has provided patient samples and clinical data. BB provided access to IMC instruments. FN, TL and DT provided reagents. CS performed and analyzed electron microscopy imaging. MF supervised scRNA-seq analysis. STM and BB conceptualized and supervised the analysis of human BM samples. FH and MD equally contributed to the manuscript. They designed and analyzed experiments, supervised all work, and wrote the manuscript. All authors contributed to writing the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kirchberger, S., Shoeb, M.R., Lazic, D. et al. Comparative transcriptomics coupled to developmental grading via transgenic zebrafish reporter strains identifies conserved features in neutrophil maturation. Nat Commun 15, 1792 (2024). https://doi.org/10.1038/s41467-024-45802-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-45802-1
- Springer Nature Limited
This article is cited by
-
Understanding vertebrate immunity through comparative immunology
Nature Reviews Immunology (2024)