
Abstract
Time-course transcriptome expression data were constructed for four parts of the silk gland (anterior, middle, and posterior parts of the middle silk gland, along with the posterior silk gland) in the domestic silkworm, Bombyx mori, from days 0 to 7 of the last-instar larvae. For sample preparation, silk glands were extracted from one female and one male larva every 24 hours accurately after the fourth ecdysis. The reliability of these transcriptome data was confirmed by comparing the transcripts per million (TPM) values of the silk gene and quantitative reverse transcription PCR results. Hierarchical cluster analysis results supported the reliability of transcriptome data. These data are likely to contribute to the progress in molecular biology and genetic research using B. mori, such as elucidating the mechanism underlying the massive production of silk proteins, conducting entomological research using a meta-analysis as a model for lepidopteran insect species, and exploring medical research using B. mori as a model for disease species by utilising transcriptome data.
Similar content being viewed by others
Background & Summary
The domestic silkworm, Bombyx mori, is a lepidopteran insect renowned for silk production. Additionally, B. mori serves as a bioreactor, enabling the development of a baculovirus vector system with silkworm1 and a transgenic silkworm system2,3, leading to the production of recombinant proteins. Understanding the mechanisms regulating silk gene expression through its high-quality genome sequences is essential for improving the production of recombinant proteins. Therefore, B. mori draft genome sequence data were first published in 20044,5 and have since been continuously updated with additional data6,7. Consequently, several related datasets, such as full-length cDNA data8, also have been published. These data are now available in public databases such as DDBJ/NCBI/ENA and in silkworm databases such as KAIKOBase, SilkBase, and SilkDB9,10,11. The development of these genome data and genetic tools has elevated the status of B. mori as a model lepidopteran species, contributing to advances in entomology (not limited to B. mori) and medical science12,13. To further expand the silkworm genome and related data, we constructed a chromosome-level reference genome and transcriptome datasets7,14. Using a new gene annotation workflow, we performed functional annotations on the silkworm gene15. Furthermore, we generated transcriptome expression data for multiple tissues in day-3 last-instar (fifth-inster) larvae (p50T (Daizo) strain, used for the genome project), when the silk gene was abundantly expressed14. The development of genome editing techniques for B. mori has enabled us to perform precise gene or genome modifications16. The w1 pnd is an experimental strain with non-diapause and white-eye traits, and is often employed for transgenic and genome editing experiments because the former trait allows us to save the treatment of the embryonic diapause disruption and the latter permits the easy observation of the fluorescent marker gene.
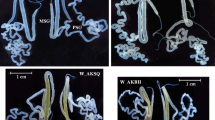
The silk gland (SG) is a key tissue for silk production and cocoon formation. The days 0 to 7 of the last-instar larvae are important stages for them. Actually, SG became larger and got more weight until day4 or day5 in this stages (Fig. 1a)17. SG comprises three parts: the anterior silk gland (ASG), middle silk gland (MSG), and posterior silk gland (PSG); the MSG is further divided into the anterior part (MSG_A), middle part (MSG_M), and posterior part (MSG_P)18,19 (Fig. 1b). Each SG component plays an important role in silk production and possesses a unique silk gene expression profile. During the life cycle of B. mori, the last-instar larvae produce silk by expressing silk genes, including Sericin1, Sericin2, Sericin3, Fibroin-H, Fibroin-L, and fibrohexamerin (fhx). While Sericin1, Sericin2, and Sericin3 are primarily expressed in MSG18, Fibroin-H, Fibroin-L, and fhx genes are expressed in PSG20,21,22. Considering these features, the time-course reference transcriptome expression data of MSG_A, MSG_M, MSG_P, and PSG from the last-instar larvae of the w1 pnd strain hold potentially high value and thus can serve as important reference datasets. Therefore, we conducted to construct these data. Actually, we prepared total RNA samples extracted from multiple SG territories from days 0 to 7 of the last-instar larvae (Fig. 1b). After performing RNA sequencing (RNA-seq) on these samples, we calculated the expression values of the reference transcripts for each sample using pre-prepared RNA-seq data (trimmed and quality-controlled) and previously reported reference transcript data14. The expression data for each sample were merged into one matrix as the time-course expression data for SG (Fig. 2). These expression data are invaluable for various aspects of B. mori research, including the molecular mechanism of silk gene expression control, lepidopteran or insect basic research (e.g., comparative genomics), basic medical research (e.g., disease models using B. mori), and producing and breeding more valuable B. mori strains with enhanced silk production capabilities.

Size changes of silk glands in last-instar larvae and preparation of RNA-seq data of the silk grands (a) Silk glands in last-instar larvae were photographed. One silk gland pair was used for this photograph and measuring the weight shown in our previous report17,30, and the other pair used for RNA extraction. (b) Schematic workflow of sample preparation for extraction of total RNA. Fifth-instar larvae, one female and one male, from day 0 to day 7, were used as one biological replicate with three replicates prepared. SGs were dissected from both larvae and separated into MSG_A, MSG_M, MSG_P and PSG. Female and male dissections were combined to form one sample. Total RNA was extracted from each prepared sample (upper panel). Abbreviations of sample names in this work are shown in the lower panel.

Schematic workflow of transcriptome analysis. Total RNA from the samples shown in Fig. 1 was sequenced. The raw RNA-seq data were deposited to SRA. Quality control and trimming of the raw RNA-seq data were performed using fastp. Calculations for the reference transcripts of each sample were performed using processed RNA-seq and reference transcript data by kallisto. The expression data from each sample were merged into a matrix data (“Time-course transcriptome data of the silk gland in the last-instar larvae”)26. These expression data were deposited in the GEA and figshare27.
Methods
Sample preparation and total RNA extraction
The w1 pnd strain of B. mori silkworms was raised on an artificial diet (Nihon Nosan Kogyo, Yokohama, Japan) at 25 °C under 12:12 h light/dark conditions. This strain shows standard developmental characteristics as an experimental strain, including SG. The series of the silkworm samples were prepared from a single experimental population. Immediately after the fourth molting, the larvae were individually placed in plastic petri dishes containing the artificial diet. Every time a larva moulted during the last-instar period, one male and one female insect were prepared and maintained for 24 h (days 0–7). Subsequently, SGs were dissected from the prepared male and female larvae in a physiological saline solution. The extracted SGs were further dissected into MSG_A, MSG_M, MSG_P, and PSG. Each SG part was then homogenised in a TRIzol (Invitrogen, Carlsbad, CA, USA) solution and stored at -80 °C. Afterwards, the supernatant treated with chloroform was mixed in equal volumes (300 µL). RNA was extracted with the removal of genomic DNA by the RNeasy Plus Mini Kit (Qiagen, Hilden, Germany). Three-biological replicates were prepared for each sample category (e.g., MSG_A_d0_1 to MSG_A_d0_3) using the different larvae (Fig. 1).
Library preparation and RNA-seq
For RNA-seq, cDNA libraries were constructed from the total RNA samples using the TruSeq Stranded mRNA Library Prep Kit, following the procedures outlined in its Reference Guide. Sequencing was performed using an Illumina NovaSeq 6000 (San Diego, California, USA.). Both library construction and sequencing were performed by Macrogen Japan Corp (Kyoto, Japan).
Data analysis for transcriptome data
The workflow for analysing the transcriptome data is shown in Fig. 2. All raw RNA-seq sequence data (FASTQ files) have been deposited in the Sequence Read Archive (SRA) (for details, see Data Records) (Fig. 2). Pre-processing of the raw sequence data involved removing adapter sequences, trimming, and quality control using fastp (version 0.22.0) with default settings23. Subsequently, using processed RNA-seq data and reference transcript sequence data of B. mori (Transcriptome Shotgun Assembly (TSA) accession ID ICPK01000001-ICPK01051926)14, TPM values of the reference transcripts in each sample were calculated using kallisto (version 0.44.0) with a bootstrap value of 100 and others as defaults24. A TPM column in abundance.tsv of each sample, which is a kallisto output data file (available in figshare)25, was extracted, which contains time-course transcript data of each sample. Another column data including transcript IDs of the reference transcript were extracted from abundance.tsv of one sample. Finally, the time-course data for all samples and the transcript ID data were merged into a single matrix dataset as “Time-course transcriptome data of the silk gland in B. mori last-instar larvae”26. The matrix data was deposited in the Genomic Expression Archive (GEA) of the DNA Data Bank of Japan (DDBJ)27 and uploaded in figshare26 (Fig. 2). All the commands used in these analyses are available28.
Hierarchical clustering analysis
To assess the similarities in expression profiles among all samples, hierarchical clustering analysis was performed using R (version 4.2.3) in RStudio (version 2023.03.0+386) with expression matrix data as input. Spearman’s rank correlation coefficient and Ward’s method were used for measuring similarities between each sample and clustering, respectively. The R code for this analysis is uploaded in figshare28.
Data Records
The RNA-seq raw data in this study were deposited in SRA29. Accession IDs of BioSamples and SRA for each RNA-seq sample are described in the table file uploaded30. Additionally, expression data across the reference transcript in all samples can be accessed via GEA (Accession ID: E-GEAD-662)27 or figshare26.
Technical Validation
Sample preparation
As described in the “Methods” section, the series of larvae samples for RNA-seq were prepared from a single experimental population, and SG samples for RNA-seq were prepared under the strictly controlled timing. After the fourth moulting, each last-instar larva was individually housed and sampled for SG extraction at precise 24-hour intervals. Only one SG pair was used for RNA extraction, whereas the other pair was used for weight measurements to assess the degree of development17. As shown in Fig. 1a and our previous report17, SGs became larger and got more weight until day4 or day5 last-instar larvae. From these data, the silk synthesis must occur in these SG samples.
Transcriptome data validations using TPM values of silk genes compared to the silk genes expression results by qPCR
To confirm the reliability of the expression values, we compared the time-course TPM values of representative silk genes with the relative expression values obtained using quantitative reverse transcription PCR (RT-qPCR) from the total RNAs17. The TPM expression values of Sericin1, Sericin2, Sericin3, Fibroin-H, Fibroin-L, and fhx are shown in Fig. 3a–f, respectively. In Fig. 3a,the TPM time-course values (total TPM values of all isoforms) of Sericin1 (Transcript ID: KWMTBOMO06216, MSTRG.2477.1-MSTRG.2477.16) in MSG_A are shown in the upper left graph. Although TPM values exhibited variations in larvae expressing Sericin1, this gene began to be expressed in day 2 larvae, peaked in day 3 larvae, and decreased after day 4. Sericin1 expression in MSG_M increased and peaked on days 4 and 5 and decreased in day 6 larvae (lower left graph in Fig. 3a). These features were consistent with the qPCR results reported previously17, except for the timing of the decline, as Sericin1 expression decreased from day 5 larvae onward in the qPCR results. The expression profile of TPM values of Sericin1 showed an increase in MSG_P from days 0 to 5 and plateaued afterward (upper right graph in Fig. 3a), whereas Sericin1 qPCR results showed that the mRNA levels of Sericin1 in MSG_P increased from day 0 to day 2 larvae and plateaued afterward. Interestingly, the time points at which Sericin1 expression levels plateaued in the MSG_P group differed between the TPM and qPCR results17. These differences in the TPM and qPCR profiles may be because the qPCR values cannot contain all the isoforms described above with the primer pair used. Notably, Sericin1 TPM values in MSG_P and MSG_M from day 0 to day 7 larvae were notably higher than those in MSG_A, as observed in the qPCR results (lower right graph in Fig. 3a). Sericin1 TPM values in MSG_P from day 5 to day 7 were larger than those in MSG_M. However, this pattern was not observed in the qPCR results. The relative mRNA values in MSG_M were larger from day 0 to day 7 (but not significantly different)17. These discrepancies could be due to the methods used to measure gene expression values. As described above, TPM values include all isoforms, whereas qPCR values may not. Sericin1 mRNA levels in MSGs from day 0 to 7 larvae (w1 pnd strain) using qPCR showed that the amounts of Sericin1 mRNA increased from day 0 to day 6 and decreased on day 7, which was comparable to the MSG_P profile of the TPM results of this work2. The TPM values of Sericin2 (Transcript ID: KWMTBOMO06334, MSTRG.2627.1, and MSTRG.2627.2) in MSG_A of larvae from day 0 to day 4 were notably high (over 15000) and dropped to nearly zero from day 5 to day 7 (upper left graph in Fig. 3b). In MSG_M, Sericin2 TPM values decreased from approximately 11000 on day 0 to low levels in day 3 larvae and remained very low in day 7 larvae (lower left graph in Fig. 3b). Similarly, TPM values of Sericin2 in MSG_P were over 11000 in day 0 larvae, dropped below 1000 in day 1 larvae, and approached nearly zero in larvae of days 2 to 7 (upper right graph in Fig. 3b). Comparing the TPM values across samples tested, Sericin2 TPM values were the highest in day 0 larvae (MSG_A having the highest value and MSG_P the lowest), decreasing dramatically across larval stages (lower right graph in Fig. 3b). These trends of Sericin2 TPM values were consistent with the qPCR results17 and in a previous report18. Sericin3 (Transcript ID: KWMTBOMO06311 and MSTRG.2595.1-MSTRG.2595.9) TPM values in MSG_A were negligible in day 0 larvae and gradually increased in day 2 to day 4 larvae (ranging between 3000 and 65000). Subsequently, the TPM values dramatically increased in day 4 and day 5 larvae, plateauing in day 5 to day 7 larvae (upper left graph in Fig. 3c). In MSG_M, Sericin3 TPM values increased in day 1 to day 5 larvae, with a sharp increase in day 4 to day 5 larvae, followed by plateauing until day 7. The MSG_P, Sericin3 TPM values were very small or nearly zero at all stages tested (lower left and upper right graphs in Fig. 3c). Sericin3 TPM values in MSG_A and MSG_M from days 3 to 7 were notably higher than those in MSG_P (lower right graph in Fig. 3c), consistent with qPCR results17. In previous studies, Sericin3 expression profiles revealed that Sericin3 expression was only observed in MSG_A of days 5 to 7 larvae, with significantly higher expression observed in the day 6 and day 7 larvae compared with that in day 5 larvae19. These findings are consistent with the results of TPM data from our study. Specifically, Sericin3 TPM values in MSG_A of day 5 to day 7 larvae were notably higher than those in the other samples. In PSG, Fibroin-H (Transcript ID: KWMTBOMO08464 and MSTRG.14927.1-MSTRG.14927.23) TPM values increased from day 0 to day 2, remained relatively stable from day 2 to day 5, and slightly increased from day 5 to day 7 (Fig. 3d), whereas Fibroin-L (Transcript ID: KWMTBOMO15365 and MSTRG.5511.1) TPM values gradually increased from day 0 to day 6 and plateaued (Fig. 3e). The expression profiles of Fibroin-H and Fibroin-L TPM values from transcriptome analysis were similar to those determined by qPCR17. Fhx (Transcript ID: KWMTBOMO01001 and MSTRG.10154.1) TPM values increased from day 0 to day 5 and decreased and plateaued from day 5 to 6 and from day 6 to 7, respectively (Fig. 3f). Fhx is associated with Fibroin-H and Fibroin-H linked by disulfide bond20, suggesting that the Fhx expression profile was similar to the profiles of Fibroin-H and Fibroin-H. The expression profiles of these three genes determined based on the TPM values were similar. Taken together, these comparisons of the six silk gene expression data using TPM values in this study, qPCR results, and findings from other reports suggest that the TPM transcriptome expression data in this study were reliable.

Expression profiles of the major silk genes. Average TPM values representing the total TPM values of all isoforms for six silk genes per biological replicate were calculated (See main text). Sericin1 (a), Sericin2 (b) and Sericin3 (c) expression levels in MSG_A (left upper graph), MSG_M (left lower graph), MSG_P (right upper graph) across day 0 to day 7 last-instar larvae (three replicates) are presented. The right lower graph displays the average TPM values of the three SG samples. Average TPM values of Fibroin-H (d), Fibroin-L (e) and Fhx (f) in PSG are shown. Total TPM values of all isoforms for each silk gene per biological replicate were calculated. For example, total TPM value of Sericin1 of each biological replicate was calculated by using all TPM values of Sericin1 isoforms within that biological replicate. The vertical bar represents the standard deviation of each point.
Transcriptome data validations by hierarchical clustering analysis
Transcriptome data validation was performed through hierarchical clustering analysis using the TPM matrix data26 as input. The results are depicted in Fig. 4. The dendrogram results show that the three biological replicates of all samples, except one (MSG_P_d0 samples), form a single cluster or small closely neighbouring clusters. Specifically, MSG_P_d0_3 is located in the MSG_M_d0 cluster. We presumed that this was because the MSG_P_d0_3 sample contained a small number of MSG_M_d0 cells, and the contaminated MSG_M_d0 cells, which expressed a large number of genes, masked the transcriptome features of MSG_P_d0. If this is true, the samples are placed in different clusters. Typically, most samples from the same category, such as MSG_M, MSG_P, and PSG, clustered together or formed two distinct intermediate clusters (MSG_A). MSG_A_d0-4 and MSG_A_d5-7 samples were located in two separate intermediate clusters. The differences in the certain gene expressions, particularly evident in highly expressed genes such as Sericin2 and Sericin3 (Fig. 3), may reflect the two separate intermediate clusters. Several samples, including PSG_d0, MSG_P_d0-2, and MSG_M_d0-2, grouped together in clusters, possibly reflected similarities in the abundantly expressed genes with similar expression profiles across these samples rather than the same territories of SG. Indeed, Sericin1 and Sericin3 TPM values between MSG_P_d0-2 and MSG_M_d0-2 were not significantly different from those of the other samples (Fig. 3). Additionally, Fibroin-H, Fibroin-L and Fhx expression levels in PSG_d0 were markedly lower than those in the other PSG samples. In summary, the hierarchical clustering analysis demonstrated that, except for a few samples, there were no significant differences in transcriptome TPM values between biological replicates, suggesting that the reliability of the transcriptome data in this study.

Dendrogram of hierarchical clustering analysis. Hierarchical clustering analysis using time-course transcriptome data26 as input was performed. The resulting dendrogram was constructed based on the results of hierarchical clustering analysis. MSG_A, MSG_M, MSG_P and PSG samples in the dendrogram are coloured yellow, purple, green, and blue, respectively. A schematic representation of the SG (as shown in Fig. 1) is included in the upper right corner.
References
Maeda, S. et al. Production of human α-interferon in silkworm using a baculovirus vector. Nature 315, 592–594 (1985).
Tatematsu, K. et al. Construction of a binary transgenic gene expression system for recombinant protein production in the middle silk gland of the silkworm Bombyx mori. Transgenic Res. 19, 473–487 (2010).
Tamura, T. et al. Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat. Biotech. 18, 81–84 (2000).
Mita, K. et al. The genome sequence of silkworm, Bombyx mori. DNA Res. 11, 27–35 (2004).
Xia, Q. et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 306, 1937–1940 (2004).
International Silkworm Genome Consortium. The genome of a lepidopteran model insect, the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 38, 1036–1045 (2008).
Kawamoto, M. et al. High-quality genome assembly of the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 107, 53–62 (2019).
Suetsugu, Y. et al. Large scale full-length cDNA sequencing reveals a unique genomic landscape in a lepidopteran model insect, Bombyx mori. G3 (Bethesda) 3, 1481–1492 (2013).
Lu, F. et al. SilkDB 3.0: visualizing and exploring multiple levels of data for silkworm. Nucleic Acid Res. 48, D749–D755 (2020).
Kawamoto, M., Kiuchi, T. & Katsuma, S. SilkBase: an integrated transcriptomic and genomic database for Bombyx mori and related species. Database 2022, baac040 (2022).
Yang, C.-C., Yokoi, K., Yamamoto, K. & Jouraku, A. An update of KAIKObase, the silkworm genome database. Database (Oxford) 2021 (2021).
Tabunoki, H., Bono, H., Ito, K. & Yokoyama, T. Can the silkworm (Bombyx mori) be used as a human disease model? Drug Discov. Ther. 10, 3–8 (2016).
Hamamoto, H. et al. Lysocin E is a new antibiotic that targets menaquinone in the bacterial membrane. Nat Chem Biol 11, 127–133 (2015).
Yokoi, K., Tsubota, T., Jouraku, A., Sezutsu, H. & Bono, H. Reference Transcriptome Data in Silkworm Bombyx mori. Insects 12, 519 (2021).
Bono, H., Sakamoto, T., Kasukawa, T. & Tabunoki, H. Systematic Functional Annotation Workflow for Insects. Insects 13, 586 (2022).
Tsubota, T. & Sezutsu, H. Genome Editing of Silkworms. Methods in Molecular Biology 1630, 205–218 (2017).
Masuoka, Y. et al. Co-Expression Network and Time-Course Expression Analyses to Identify Silk Protein Regulatory Factors in Bombyx mori. Insects 13, 131 (2022).
Takasu, Y. et al. Identification and characterization of a novel sericin gene expressed in the anterior middle silk gland of the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 37, 1234–1240 (2007).
Takasu, Y., Hata, T., Uchino, K. & Zhang, Q. Identification of Ser2 proteins as major sericin components in the non-cocoon silk of Bombyx mori. Insect Biochem Mol Biol 40, 339–344 (2010).
Inoue, S. et al. Silk Fibroin of Bombyx mori Is Secreted, Assembling a High Molecular Mass Elementary Unit Consisting of H-chain, L-chain, and P25, with a 6:6:1 Molar Ratio *. J. Biol. Chem. 275, 40517–40528 (2000).
Gamo, T., Inokuchi, T. & Laufer, H. Polypeptides of fibroin and sericin secreted from the different sections of the silk gland in Bombyx mori. Insect Biochemistry 7, 285–295 (1977).
Maekawa, H. & Suzuki, Y. Repeated turn-off and turn-on of fibroin gene transcription during silk gland development of Bombyx mori. Dev. Biol. 78, 394–406 (1980).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884–i890 (2018).
Bray, N. L., Pimentel, H., Melsted, P. & Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat Biotechnol 34, 525–527 (2016).
Masuoka, Y. et al. Kallisto output files. Figshare https://doi.org/10.6084/m9.figshare.24978483 (2024).
Masuoka, Y. et al. Matrix data of the expression values of all reference transcripts in all samples. Figshare https://doi.org/10.6084/m9.figshare.24921435 (2024).
DNA Data Bank of Japan https://ddbj.nig.ac.jp/public/ddbj_database/gea/experiment/E-GEAD-000/E-GEAD-662/ (2024).
Masuoka, Y. et al. Script file for transcriptome and hierarchical clustering analyses. Figshare https://doi.org/10.6084/m9.figshare.24921459 (2024).
NCBI Sequence Read Archive https://identifiers.org/ncbi/insdc.sra:DRP011242 (2024).
Masuoka, Y. et al. List of metadata for all RNA-Seq data. Figshare https://doi.org/10.6084/m9.figshare.24921426 (2024).
Acknowledgements
This work was supported by the MAFF Commissioned project study on the “Research project for sericultural bio-industry” (Grant Number JP22680575) to Y.M., T.T., H.S. and K.Y., ROIS-DS-JOINT (024RP2023) to Y.M, A.J., H.O., H.C., H.B. and K.Y., by the Center of Innovation for Bio-Digital Transformation (BioDX), an open innovation platform for industry-academia co-creation (COI-NEXT) of JST (COI-NEXT, JPMJPF2010) to Y.M., A.J., T.T., H.S., H.B. and K.Y. and by and JSPS KAKENHI Grant Number 21H03831 to T.T., H.S. and K.Y. Parts of the Fig. 2 were drawn by using illustrations from TogoTV (© 2016 DBCLS TogoTV, CC-BY-4.0 https://creativecommons.org/licenses/by/4.0/deed.ja).
Author information
Authors and Affiliations
Contributions
Y.M., T.T., H.O., H.C., H.S., H.B. and K.Y. conceived the study. Y.M. prepared the RNA samples from silkworms for RNA-seq and photograph data of silk glands. Y.M., A.J., H.B., and K.Y. performed the bioinformatics data analysis and data registration. All authors curated and validated the data and metadata analysed in this study. K.Y wrote the original draft of the manuscript. All authors reviewed and edited the draft of the manuscript. H.O., H.S., H.B. and K.Y. contributed to funding acquisition. All the authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Masuoka, Y., Jouraku, A., Tsubota, T. et al. Time-course transcriptome data of silk glands in day 0–7 last-instar larvae of Bombyx mori (w1 pnd strain). Sci Data 11, 709 (2024). https://doi.org/10.1038/s41597-024-03560-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41597-024-03560-1
- Springer Nature Limited