
Abstract
While it has been well characterized that chemosensory receptors in guts of mammals have great influence on food preference, much remains elusive in insects. Insect chemosensory proteins (CSPs) are soluble proteins that could deliver chemicals to olfactory and gustatory receptors. Recent studies have identified a number of CSPs expressed in midgut in Lepidoptera insects, which started to reveal their roles in chemical recognition and stimulating appetite in midgut. In this study, we examined expression patterns in midgut of 21 Spodoptera litura CSPs (SlitCSPs) characterized from a previously reported transcriptome, and three CSPs were identified to be expressed highly in midgut. The orthologous relationships between midgut expressed CSPs in S. litura and those in Bombyx mori and Plutella xylostella also suggest a conserved pattern of CSP expression in midgut. We further demonstrated that the expression of midgut-CSPs may change in response to different host plants, and SlitCSPs could bind typical chemicals from host plant in vitro. Overall, our results suggested midgut expressed SlitCSPs may have functional roles, likely contributing to specialization and adaption to different ecosystems. Better knowledge of this critical component of the chemsensation signaling pathways in midguts may improve our understanding of food preference processes in a new perspective.
Similar content being viewed by others


Introduction
The tobacco cutworm, Spodoptera litura (F.) (Lepidoptera: Noctuidae), is a generalist herbivore and one of the most important pests in many countries. The economic importance of S. litura is owing to its high increase rate and wide host spectrum, encompassing a large assortment of agricultural crops, including vegetables, green manures, and horticultural plants, as well as miscellaneous wild plants and weeds1. Chemical cues, emitted from host plants of phytophagous insects, may elicit a wide range of behavioral and physiological responses of insects, including feeding, oviposition and courtship2. Among those behaviors, feeding is a complex behavior that could be regulated by several internal mechanisms3. Recently, the discovery of chemosensory related proteins in the gut has led to intensive researches on their roles in gut chemical discriminations, stimulating appetite and conditioning food preferences4. A myriad of genes in midgut could help maintain balance by prompting foraging and feeding, or by encouraging the cessation of feeding behaviors of insects. Studies suggested that the exposure to plant defense chemicals could have negative effects on insect gut microbial community composition, thus affecting the subsequent feeding behaviors of insects5. In addition to this homeostatic regulation, a post-feeding reward system in midgut could also positively reinforce feeding activities6. Slight adjustments to midguts system can tilt the balance to affect the nutrient adversity6. Extensive studies have been carried out with the upper (oral) regions of chemosensory process to illuminate the mechanisms of host and food preference7, 8. However, much remains to be discovered about the lower (gastric, intestinal) regions of the alimentary canal, which were also proved to be critical for the stimulation or inhibition of feeding behaviors9.
Normally, the reception of chemical messages in insect starts when chemosensory-related proteins bind the chemicals and transport them through the aqueous hemolymph10. Chemosensory protein (CSPs) and odorant binding protein (OBPs) are often speculated to play roles in initial stage of chemosensory perception by insects11, 12. While many OBPs have restricted expression pattern in main chemosensory tissues such as antennae, CSPs could express in a variety of tissues and may be involved in divergent functions13, 14. One intriguing possibility is that, in midgut, CSPs also could perceive signal chemicals from food resources, thus to mediate feeding behaviors. The analysis of gene expression in response to different treatments have potential to understand the biological function of such genes in adapting surrounding environments15 and the binding of an external ligand to the CSPs could result in action potentials and contribute to subsequent behaviors16. Eventually, studying of CSPs expression variations in response to different plant resources and binding affinities may elucidate the role of CSPs in midgut.
In our study, 21 CSPs were identified from the transcriptome data we previously conducted. On the quest to challenge our hypothesis, we examined whether the expression levels of selected CSPs in midguts could response to different host plants. And the binding activities between CSPs and typical host plant chemicals were investigated by competitive binding essay in vitro. This study could potentially uncover the chemosensory protein variations in midguts in response to host plants, which could surely provide foundation for facilitating the understanding of host recognition and feeding preferences.
Materials and Methods
Sample preparation and RNA isolation
S. litura (F.) larvae were reared on an artificial diet consisting of soybean, yeast extract, wheat bran, and maintained at 27 °C and 70% RH with a 14: 10 h L: D photoperiod17. Adults were transferred to Chinese cabbage [Brassica campestris L. ssp. Chinensis (L.)] and raised in a greenhouse at 25 °C and 60–70% RH and the honey was added as a dietary supplement.
The total RNA was extracted using the E.Z.N.A.TM total RNA isolation system kit (Omega, USA) according to the manufacturer’s instructions. The concentration of isolated RNA was examined by Nanophotometer. All tissues were stored at −80 °C until to be used experimentally. One μg of the isolated RNA was transcribed to first-strand cDNA by M-MLV reverse transcriptase (TaKaRa, China) and oligo(dT)18 as primer at 42 °C for 60 min. The reaction was terminated by heating at 95 °C for 5 min, and the products were stored at −20 °C.
Identification of CSPs from transcriptome and genome datasets
CSP genes in S. litura were identified from the de novo transcriptome assembly previously reported by our group which was based on mixed RNA samples from multiple developmental stages, including larva18. The original study was centered on Cry toxin receptor, and to our knowledge, this transcriptome dataset has not been used for any CSP related study. Likely coding regions in the transcriptome assembly were annotated using TransDecoder v2.0.1 and translated protein sequences were checked for the presence of the characteristic domain of CSPs (IPR005055) using InterProScan v5. Protein sequences of CSPs in S. litura (identified in the previous step) were used as queries to perform TBLASTN search to identify putative CSP coding regions. For such region, homology-based gene prediction was performed using GeneWise v2.2.0 with the most similar query sequence as reference. All predicted genes were further examined for the presence of the characteristic CSP domain in their translated protein sequences. CSPs in Bombyx mori and Plutella xylostella were identified from their genome assemblies (downloaded from the LepBase: http://ensembl.lepbase.org/) using the same approach. The identified CSPs in S. litura and P. xylostella were named after orthologous genes in B. mori.
CSPs expressions in midguts
For each sample, the midguts from five S. litura (fourth-instar larva of S. litura) were dissected and immediately transferred into eppendorf tubes immersed in liquid nitrogen, and all treatments were conducted in three replicates. The expression levels of all identified SlitCSPs in the midguts of S. litura were examined by PCR. The primers of all 21 identified SlitCSPs were designed and synthesized as recommended, and the actin gene of S. litura was used to normalize the target gene expression (Table 1). The concentration of the primes used in the reaction is 10 nmol. Amplification was performed by denaturing at 94 °C for 5 min, followed by 27 cycles of 94 °C for 30 s, 60 °C for 30 s and 72 °C for 45 s, with a final extension at 72 °C for 10 min. PCR products were analyzed on 1.2% agarose gels.
To measure the midgut expression levels of B. mori and P. xylostella, the following larval midgut transcriptome data were downloaded from the NCBI Sequence Read Archive database: B. mori-SRR1805030, SRR1806712, SRR1806713, SRR1806715, and SRR1806736, and P. xylostella-SRR835315, SRR835316, SRR835317. Reads were trimmed for low-quality positions using Trimmomatic v0.35, aligned to respective genome assemblies using STAR v2.4.2a with gene annotations as guidance, and uniquely mapped reads were counted using HTSeq v0.6.1p1. For all genes, the reads per kilobase per million mapped reads (RPKM) values were calculated as a measure of expression level, and the percentile ranks within their respective transcriptomes were determined accordingly.
Expression pattern of three SlitCSPs in other tissues
The expression patterns of three selected candidate CSPs were further investigated by quantitative real-time PCR (qRT-PCR). RNA samples were isolated from different developmental stages (including first to sixth-instar larvae, pre-pupae, pupae and adult), and different tissues (including cuticle, midguts, fatbody, antennae, heads (without antennae), wings, legs, abdomens, testis and ovary). qRT-PCR was performed using iCycler iQ Real-Time PCR Detection System (Bio-Rad) with SYBR green dye (Taraka, China) binding to double-strand DNA at the end of each elongation cycle. Amplification process was carried out by using the same primers as previously mentioned (Table 1). For all of the tested samples, the concentration of total RNA used in reverse transcription to get first-strand cDNA is 1 μg. All amplifications were performed with three biological replicates. Relative gene expression data were analyzed using the 2−ΔΔCT method as described by Livak19.
Bioassay
As fourth-instar larva of S. litura have active feeding behaviors and just enter the first stage of gluttony20,21,22, we used fourth-instar larvae to examine the expression levels of candidate CSPs in midgut. Therefore, fourth-instar larvae from the same piece of egg fraction were collected. The starvation group was set as negative control, which fed on nothing, while the group reared on artificial diet was set as positive control. The other two groups were reared on cabbage (Brassica camperstris ssp.pekinens) and tobacco (Nicotiana tabacum L.), respectively. Every group has five larvae, and repeated for three times. After 24 h feeding, the midguts of the tested S. litura were dissected, and the remains in the midguts were removed. By qRT-PCR, the expression levels of SlitCSPs in midguts after treatments were examined as described previously.
Expression of recombinan CSPs
The sequences encoding three mature SlitCSPs with EcoR I (GAATTC) and Xho I (CTCGAG) were connected to pET32a (Invitrogen, US) by T4 DNA ligase (Takara, China) at 14 °C, and then transformed to BL21 (DE3) competent cells (Takara, China). The selected positive bacterial colony was then inoculated in liquid LB overnight at 37 °C, then transferred 50 μL overnight bacterial liquid to 50 mL fresh LB (Ampicilin 100 μg/mL) until its OD600 reached 0.4–0.6. Isopropyl-D-thiogalactoside (IPTG) (0.6 mmol/L) was added and then incubated at various times at 28 °C. After breaking by sonic oscillator, 30 μg of expression product of protein was examined by SDS-PAGE and Western blot. The recombinant protein was purified by affinity chromatography using HisTrap columns prepacked with Ni Sepharose (GE Healthcare) according to the specifications. After overnight dialysis in Tris-HCL (pH = 7.4), the protein was subjected to the Bovine Enterokinase overnight to remove the His-tag. The purified protein was collected and examined by 12% SDS–PAGE. Bradford method was used to determine protein concentration23. Purified recombinant SlitCSPs protein was used to immunize rabbit as described previously. The sera of the immunized rabbit was collected as SlitCSP sera24. The serum titer was showed to have an enzyme linked immunosorbent assay (ELISA) end point of 1:12, 000 using the method of indirect ELISA25. Western-blotting analysis was modified according to the methods previously described26. Samples were electrophoresed on 12% SDS polyacrylamide mini-gels and transferred to PVDF membranes using Tris–glycine transfer buffer on a mini-Trans-Blot electrophoretic transfer tank (Bio-RAD, USA). Blots were blocked in TBS (100 mM Tris–HCl, pH 7.5, 0.9% NaCl) containing 5% nonfat powdered milk and 0.1% Tween-20 for 1 h. The immunoreactivity was tested with the anti-SlitCSP serum (diluted 1: 5000), and incubated with the filter overnight at 4 °C. Blots were washed with TBST three times. An IgG anti-rabbit antibody conjugated with HRP was used as a secondary antibody (Tiangen, China) and finally visualized by ECL (enhanced chemiluminescence).
Fluorescence competitive Binding Assays
The fluorescence spectra were recorded on an F-4500 FL Fluorescence Spectrophotometer (HITACHI) in a 1 cm light path quartz cuvette at 23 °C. The slit width used for excitation and emission was 5 nm. The compounds used to investigate the binding abilities of SlitCSPs were purchased from Sigma-Aldrich with the highest purity and stored as specified instruction by the manufacturer. The selected chemicals were listed in Table 2, which are the typical volatiles of Chinese cabbage (Brassica camperstris ssp.pekinens) and tobacco (Nicotiana tabacum L.) based on previous investigations. The fluorescent probe N-phenyl-1-naphthylamine (1 − NPN) and all ligands used in competition experiments were dissolved in HPLC purity grade methanol. To measure the affinities of 1 − NPN to three purified SlitCSPs protein, the fluorescence of 2 μM 1 − NPN in 50 mM Tris-HCl was excited at 337 nm and emission spectra were recorded between 350 nm and 480 nm. And then, 2 μM of protein was added and titrated with aliquots of 1 mM 1 − NPN to final concentrations of 2 to 20 μM. The affinities of the chemicals were measured by competitive binding assays in presence of three candidate SlitCSPs protein at 2 μM and 1 − NPN at 4 μM by adding ligands from 0 to 20 μM. All values reported were obtained from three independent measurements. The corresponding to the maximum fluorescence emission was plotted against the ligand concentrations for the determination of the binding constants. The curves were linearized by Scatchard plots. The dissociation constants of the competitors were calculated using the corresponding IC50 values according to the equation: KD = [IC50]/(1 + [1 − NPN]/K1−NPN), where [1 − NPN] is the free concentration of 1 − NPN and K1−NPN is the dissociation constant of the complex protein/1 − NPN27.
Statistical analysis
All the results from experimental replicates ere expressed as the mean (±S.E.M.) and analyzed by one-way analysis of variance (ANOVA) and t-test using SPSS 17.0 for Windows software (SPSS Inc, Chicago).
Result
Identification Midgut Expressed Chemosensory Proteins in S. litura
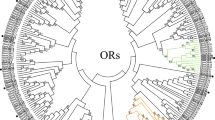
By analyzing the de novo transcriptome assembly of S. litura we reported previously18, we identified 21 non-redundant CSP coding transcripts (hereafter referred to as SlitCSP genes) (Table 1S, Fig. 1) including 14 SlitCSPs characterized in another transcriptome study28 and seven newly discovered ones. We then performed RT-PCR to determine the midgut expression levels of these CSPs. As shown in Fig. 2, 14 of the 21 SlitCSPs had detectable expression in midgut. Among them, SlitCSP11, SlitCSP3 and SlitCSP8 could express highly in midgut. Therefore, these CSPs (SlitCSP11, SlitCSP3 and SlitCSP8) were selected for further studies.

Phylogenetic analysis of chemosensory protein in three Lepidoptera insects. The phylogenetic tree was constructed in MEGA 6.0 using neighbour-joining method. Bootstrap values >50% (1000 replicates) are indicated at the nodes. ● The results showed the RPKM values of BmorCSP4, BmorCSP17 from two or more transcriptome data could reach to top 25%. ○ The RPKM values of BmorCSP3, BmorCSP9, BmorCSP14 from at least one transcriptome dataset were above average value. ■ The RPKM values of PxylCSP4, PxylCSP5, PxylCSP7, PxylCSP8, PxylCSP13, PxylCSP22, PxylCSP23, PxylCSP24, PxylCSP26, PxylCSP27, PxylCSP32, PxylCSP41, PxylCSP42, PxylCSP43 from two or more transcriptome dataset could reach to top 25%, which indicated their high expression were reliable. □ The RPKM values of PxylCSP10 and PxylCSP29 from at least one transcriptome dataset were above average value.

Expression levels of identified SlitCSPs in midguts by RT-PCR. Detection of identified SlitCSPs in midguts by RT-PCR. A: actin gene of S. litura. M: DNA maker. 1, SlitCSP11, SlitCSP3 3: SlitCSP8 4: SlitCSP5 5: SlitCSP12.1; SlitCSP12.2; 7, SlitCSP6; 8, SlitCSP18; 9: SlitCSP19; 10, SlitCSP1; 11, SlitCSP14; 12: SlitCSP15; 13: SlitCSP9; 14: SlitCSP17; 15, SlitCSP13; 16: SlitCSP18; 17: SlitCSP2; 18: SlitCSP26; 19: SlitCSP10; 20: SlitCSP25; 21: SlitCSP4. The red box indicated three candidate CSPs were selected to subsequent study.
To compare the midgut expressed CSPs in S. litura with that in other closely related insects, we also identified 23 CSP genes in (BmorCSPs) and 43 CSP genes in P. xylostella (PxylCSPs) from previous reported genome assemblies compared with previous reports29, 30, these include one new BmorCSP and 11 new PxylCSPs. Phylogenetic analysis of all 87 CSPs from the three insect revealed 17 well-supported clades (bootstrap support ≥70%) displaying clear orthologous relationships between CSPs from different species (Fig. 1). Almost all these clades consist of a single effective gene (or group of very recent duplicates in the case of P. xylostella) from each species. Interestingly, the B. mori and P. xylostella orthologs of SlitCSP3, SlitCSP8, and SlitCSP11 are all highly expressed in midgut (Fig. 1).
Expression pattern of three SlitCSPs
Three SlitCSPs with highest midgut expression levels (SlitCSP11, SlitCSP3 and SlitCSP8) were selected for in-depth analysis of their expression patterns. For S. litura, early instar larvae were fed on the lower part of leaf layers. For 2nd and 3rd instar larvae of S. litura, they started to proliferation. And from 4th to 6th instar larvae, the moths start to enter one period of gluttony, that is, the insect could eat many kinds of plants without special selectivity31. As its relatively stable expression level of 2nd instar larvae, RT-qPCR was used to investigate the expression levels of these three SlitCSPs in various developmental stages and tissues by using the 2nd instar larvae sample as the calibrator32. Although SlitCSPs were found to be expressed at multiple stages, their expression levels varied greatly from each other. For SlitCSP11, the highest expression was observed at the 1st instar larva, reaching to 13.56-fold higher compared with the 2nd instar larvae. The highest expression level of SlitCSP3 was observed in pre-pupae, which was 195.87-fold higher than that of the 2nd instar larvae. Other high expression level of SlitCSP3 was observed in 1st instar larvae and pupae, reached to 26.95-fold and 19.61-fold higher compared with 2nd instar larvae. Likewise, high expression of SlitCSP8 was also observed in 1st instar larvae, which was 30.28-fold higher than 2nd instar larvae. However, the highest expression level of SlitCSP8 was observed in pupae, which is 170.40-fold higher than 2nd instar larvae (Fig. 3a).

Quantitative real-time RT-PCR analysis the expression pattern of three CSPs in S. litura. (a–c) Different developmental stages: 1st: 1st instar larva; 2nd: 2nd instar larva; 3rd: 3rd instar larva; 4th: instar larva; 5th: 5th instar larva; 6th: 6th instar larva; pp: pre-pupae, p: pupae, ad: adult. (d–f) Different adult tissues: M: midguts; B: body wall; F: fat body; A: antenna; H: head (without antenna); AB: abdomen; L: leg; W: wing; T: testis; O: ovary. (a,d,b,e,c and f) represent the SlitCSP11, SlitCSP3 and SlitCSP8 respectively.
The expression levels of three candidate SlitCSPs in various tissues were also examined, including cuticle, midguts, fatbody, antennae, heads (without antennae), wings, legs, abdomens, testis and ovary (Fig. 3b). Besides high expression in the chemosensory organs, these three SlitCSPs were also expressed in non-chemosensory organs, while the highest expression was observed in cuticle, fatbody and midguts. The SlitCSP3 was expressed in both chemosensory and non-chemosensory organs, namely, fatbody and antennae. However, unlike SlitCSP11 and SlitCSP3, the SlitCSP8 was expressed in all tested tissues except ovary and cuticle.
CSPs expression level analysis after different treatments
Three SlitCSPs were examined for expression variations in the midgut of fourth instar larvae upon feeding with different diets. After treatment, the survival rate for the treatment group and control group are 100%. The results showed that the expression of SlitCSP11 in midgut was up-regulated by 2.15-fold, 6.63-fold and 2.16-fold after the moths were fed with artificial diet, cabbage and tobacco, respectively (Fig. 4). When compared with negative control, the expression of SlitCSP3 in midgut was up-regulated by 3.68-fold, 2.71-fold and 2.09-fold after the treatment of artificial diet, cabbage and tobacco, respectively. The expression of SlitCSP8 in midgut was up-regulated by 9.26-fold and 6.05-fold after fed with cabbage and tobacco respectively. As control, the expression levels of CSPs in fat body increased compared with the starvation group, however, significant differences could not be observed among different treatments (Fig. 4).

Relative expression of SlitCSPs after different treatments in midgut and fatbody. (a) midgut; (b) fatbody. CK was negative control, which fed on nothing. 1, SlitCSP11; 2, SlitCSP3; 3, SlitCSP8. All the data represent the mean values ± S.E.M. of replicates. Different letters indicated significant differences of expression levels of candidate CSPs between the treated by treatments and CK, as determined using a t-test (p < 0.05).
Fluorescence Binding Assays
The result showed that three SlitCSPs recombinant protein were successfully induced and expressed (Supplementary Fig. 1a–c). These SlitCSPs were resolved as a single band with molecular weight of around 33 kDa by western blot (Supplementary Fig. 1d), and then the proteins were purified successfully (Supplementary Fig. 1e). After subjected to the Bovine Enterokinase, the result of SDS-PAGE showed the molecular weight of the recombinant protein SlitCSP11, SlitCSP3 and SlitCSP8 was 12.1, 13.9, and 14.9 KDa, respectively, after removed the His-tag (18 kD) successfully (Supplementary Fig. 1f). Three proteins could be used for further investigation.
The protein was expressed with good yield (20.5 μg/μl, 12.3 μg/μl, 22.7 μg/μl). By titrating the SlitCSPs with increasing concentration of 1 − NPN, a saturation (Fig. 5a) and linear Scatchard plot were observed (Fig. 5a), indicating a single binding site and no allosteric effect. Three SlitCSPs could bind to the probe with dissociation constants of 2.860 μM, 3.337 μM and 4.756 μM (Fig. 5b). By using 1 − NPN as the fluorescent reporter, the affinities of SlitCSPs to a series of compounds were measured in competitive binding assays (Fig. 6). The IC50 values (the concentration of the ligand that yielded 50% of the initial fluorescence value) and calculated binding constants were reported in Table 2. From the results, the SlitCSP8 showed better binding activities with the typical odors from both cabbage and tobacco when compared with SlitCSP11 and SlitCSP3. Many chemicals could complete the 1 − NPN from the binding site of SlitCSP8 at low concentrations, namely, cis-3-Hexen-1-ol, benzyl alcohol and isonicotinamide. Most of the selected ligands could bind to the SlitCSP8, except for Hexanal, 2-Ethylfuran, Benzonitrile and Decanonitrile, which showed poor affinities with all the tested protein. For SlitCSP11, it showed high affinities with the five compounds of tobacco and three compounds of cabbage. SlitCSP3 had good binding activities with four ligands of tobacco and four ligands of cabbage.

Ligand-binding assays of the three SlitCSPs. (a,b,c) Binding curve for different concentration of 1 − NPN to SlitCSP11, SlitCSP3 and SlitCSP8. (d,e,f) Scatchard plot of these three CSPs. The binding curve of 1 − NPN and relative Scatchard plot analysis (insert). To measure the affinity of 1 − NPN to three SlitCSPs, the fluorescence of 2 μM 1 − NPN in 50 mM Tris-HCl was excited at 337 nm and emission spectra were recorded between 350 nm and 480 nm. Then, 2 μM of protein was added and titrated with aliquots of 1 mM 1 − NPN to final concentrations of 2 to 20 μM. The experiment was replicated for at least three times, and the data were analyzed using Prism software and indicated the presence of a single binding site. The solution was excited at 337 nm.

Competitive binding activities of the selected ligands with three candidate SlitCSPs. (a) SlitCSP11; (b) SlitCSP3; (c) SlitCSP8. 1: Competitive binding activities of the candidate protein with typical odor chemicals from tobacco. 1: β-Ionone; 2: Hexadecanoic acid; 3: Menthol; 4: Isonicotinamide; 5: Benzyl alcohol; 6: Benzaldehyde; 7: Pyridine; 8: Furan-2-carboxaldehyde; 2: Competitive binding activities of the candidate protein with typical odor chemicals from cabbage. A: cis-3-Hexen-1-ol; B: 2-Ethylfuran; C: Phenol; D: Phenethyl isothiocyanate; E: Styrene; F: Benzonitrile; G: Decanenitrile; H: Hexanal; I: 5-(Hydroxymethyl)furfural; J: trans-2-Hexenoic acid; K: Butyl isothiocyanate; L: Isothiocyanicacid; M: 6,10-Dimethyl-5,9-undecadien-2-one.
Discussions
Midgut, as a dynamic tissue, was suggested to play a vital role in metabolism, digestion and detoxification33, 34. In Lepidoptera, previous studies have focused on the role of proteases, lipases and carbohydrases in digestion, carboxylesterases, glutathione-S-transferases and cytochrome P450s in midgut35,36,37. Other than those, our study focused on physiological roles of chemosensory-related protein in midgut. In the present study, we identified 21 CSPs from the transcriptome assembly of S. litura, 23 CSPs from the genome assembly of B. mori and 43 CSPs from the genome assembly of P. xylostella (Fig. 1). Midgut expression levels of CSPs in S. litura were examined by RT-PCR. Our results showed that not all the identified CSPs could be detected to be expressed in midgut (Fig. 2). The same phenomenon was observed in Bactrocera dorsalis, among the identified four CSPs in B. dorsalis, only two of them could be detected in abdomen38.
Three candidate CSPs (SlitCSP11, SlitCSP3, SlitCSP8), which showed highest expression levels in midguts, were selected to further investigation. These three CSPs expressed highly in early pupae (Fig. 3), which implied their function in chemoreception in this period39, 40. Consistent with our results here, a previous RNA sequencing (RNAseq) study in S. litura revealed that the expression levels of many SlitCSPs could be enhanced in whole insect bodies, including thorax, wings, labial palps, tarsi, proboscis, pheromone glands and ejaculatory ducts38, 41, 42. One interpretation of those data is that SlitCSPs are functional in nonhead tissues where they are used in noncanonical chemosensory roles. In fact, these proteins have been shown to be involved in development43, 44 and immune protection44. Other similarly fancy experiments in moths have suggested a role of CSP as wetting agent to reduce the surface tension of aqueous sugar solutions and thereby reduce the pressure involved in sucking nectar45.
As previous reports suggested that the repertoires of chemosensory related proteins, especially CSPs, could be under the selection pressure that is influenced by the ecological status of different insect species46. Comparative gene expression studies enable the identification of biological functions involved in the adaptation of organisms to their surrounding environments15. By proteomics approach, Celorio-Mancera et al. demonstrated the expression of CSP1 changed in response to the caterpillar diet in the mandibular glands. In the meanwhile, the CSP2 abundances also changed in both labial and mandibular glands after changed to different diet resources16. Some CSPs might also act in a sort of immune protection in gut against insecticides, as their gene expressions have been reported to be upregulated in the gut of some insect species by such insecticides47, 48. Fourteen CSP genes in the silkworm moth were significantly up-regulated in various non-chemosensory tissues in response to avermectins, which suggested the roles of such protein in xenobiotic degradation and insect defense in the whole body48. In this study, the expression levels of three SlitCSPs in midgut were regulated when switched to different feeding resources (Fig. 4), which suggested those CSPs could be functional in midgut. This may be the case when these proteins act as sequestering agents for noxious compounds or as nutrient solubilisers, but it is also likely that they might play the role of carriers for specific hormones49. The function of CSPs in regulating nutrient signals is also supported by the fact that the identification of abundant CSPs in the proboscis of Lepidoptera, which was suggested to be involved in helping solubilizing important hydrophobic nutrients49. In this study, the changes in expression levels of CSPs in midguts in response to different diets provided us with initial evidence, which could support our hypothesis that CSPs may play roles in midguts and may eventually control insect behaviors by influencing nutrient utilization or inhibiting appetite. The detailed mechanism of CSPs in midgut needs further studies.
All these functions, although unrelated to chemical communication, may still be linked to the binding capacities of CSPs for all sorts of hydrophobic chemicals. Generally consistent with bioassay, in the fluorescence binding assays in this study, SlitCSP6 also showed good binding activities with typical volatile signals from cabbage. The SlitCSP8 could bind well with typical chemicals from tobacco, and when the S. litura was fed by tobacco, the abundance of SlitCSP8 in midgut was up-regulated. However, when fed with Cabbage, the expression level of SlitCSP8 did not show significant change. This may be due to the reason that the SlitCSP8 could not bind well with typical odors from cabbage in competitive binding assays (Table 2). Different host plants may cause changes in expression levels of CSPs, due to their recognition abilities and binding preferences to different chemical signals from different food resources. The internal hydrophobic binding cavity composed of α-helices, which could bind the relevant chemicals, enabled CSPs execute such function50.
Overall, 21 CSPs from S. litura were identified in this study. Moreover, the expression variations of CSPs in midguts responding to different diet treatments and the binding affinities between CSPs and typical odors from host plants were investigated. The results of this study could preclude or reinforce that the changes of CSP expression levels in midgut may act in concert to modulate host specialization to adapt different ecosystem.
References
Tuan, S.-J. et al. Effects of green manure cover crops on Spodoptera litura (Lepidoptera: Noctuidae) populations. Journal of economic entomology 107, 897–905 (2014).
Gerofotis, C. D., Ioannou, C. S., Nakas, C. T., Papadopoulos, N. T. The odor of a plant metabolite affects life history traits in dietary restricted adult olive flies. Scientific Reports. 6 (2016).
Sakurai, T. et al. Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 92, 573–585 (1998).
Xu, W., Zhang, H.-J. & Anderson, A. A sugar gustatory receptor identified from the foregut of cotton bollworm Helicoverpa armigera. Journal of chemical ecology 38, 1513–1520 (2012).
Xu, L. et al. Pine Defensive Monoterpene α-Pinene Influences the Feeding Behavior of Dendroctonus valens and Its Gut Bacterial Community Structure. International Journal of Molecular Sciences 17, 1734 (2016).
Slade, J., Staveley, B. Manipulation of components that control feeding behavior in Drosophila melanogaster increases sensitivity to amino acid starvation. Genetics and molecular research: GMR. 15 (2016).
Zhang, Y. V., Raghuwanshi, R. P., Shen, W. L. & Montell, C. Food experience-induced taste desensitization modulated by the Drosophila TRPL channel. Nature neuroscience. 16, 1468–1476 (2013).
Drewnowski, A., Mennella, J. A., Johnson, S. L. & Bellisle, F. Sweetness and food preference. The Journal of nutrition 142, 1142S–1148S (2012).
Sclafani, A. & Ackroff, K. Role of gut nutrient sensing in stimulating appetite and conditioning food preferences. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 302, R1119–R1133 (2012).
Leal, W. S. et al. Kinetics and molecular properties of pheromone binding and release. Proceedings of the National Academy of Sciences of the United States of America 102, 5386–5391 (2005).
Yi, X. et al. Involvement of a specific chemosensory protein from Bactrocera dorsalis in perceiving host plant volatiles. Journal of chemical ecology 40, 267–275 (2014).
Viano, M. et al. Deciphering the odorant binding protein-olfactory receptor interactions. Chemical Senses. 40, 242 (2015).
Angeli, S. et al. Purification, structural characterization, cloning and immunocytochemical localization of chemoreception proteins from Schistocerca gregaria. European Journal of Biochemistry 262, 745–754 (1999).
Zhang, T. et al. Male-and female-biased gene expression of olfactory-related genes in the antennae of Asian corn borer, Ostrinia furnacalis (Guenée)(Lepidoptera: Crambidae). PloS one. 10, e0128550 (2015).
Eyres, I. et al. Differential gene expression according to race and host plant in the pea aphid. Molecular Ecology. 25, 4197–4215 (2016).
de la Paz Celorio-Mancera, M. et al. Chemosensory proteins, major salivary factors in caterpillar mandibular glands. Insect biochemistry and molecular biology 42, 796–805 (2012).
Qi, C., Jin, L. & Hong, P. A simple artificial diet for mass rearing of some noctuid species. Entomological knowledge 37, 8–10 (2000).
Gong, L. et al. Homologs to Cry toxin receptor genes in a de novo transcriptome and their altered expression in resistant Spodoptera litura larvae. Journal of invertebrate pathology 129, 1–6 (2015).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. methods 25, 402–408 (2001).
Hummelbrunner, L. A. & Isman, M. B. Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm, Spodoptera litura (Lep., Noctuidae). Journal of Agricultural and Food Chemistry 49, 715–720 (2001).
Kamaraj, C., Rahuman, A. A. & Bagavan, A. Antifeedant and larvicidal effects of plant extracts against Spodoptera litura (F.), Aedes aegypti L. and Culex quinquefasciatus Say. Parasitology research. 103, 325–331 (2008).
Koul, O., Shankar, J. & Kapil, R. The effect of neem allelochemicals on nutritional physiology of larval Spodoptera litura. Entomologia Experimentalis et Applicata 79, 43–50 (1996).
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry. 72, 248–254 (1976).
Cui, S.-Y. & Xu, W.-H. Molecular characterization and functional distribution of N-ethylmaleimide-sensitive factor in Helicoverpa armigera. Peptides 27, 1226–1234 (2006).
Vogt, R. G., Riddiford, L. M. & Prestwich, G. D. Kinetic properties of a sex pheromone-degrading enzyme: the sensillar esterase of Antheraea polyphemus. Proceedings of the National Academy of Sciences 82, 8827–8831 (1985).
Stamnes, M. A. et al. The cyclophilin homolog ninaA is a tissue-specific integral membrane protein required for the proper synthesis of a subset of Drosophila rhodopsins. Cell 65, 219–227 (1991).
Gu, S.-H. et al. Sex pheromone recognition and immunolocalization of three pheromone binding proteins in the black cutworm moth Agrotis ipsilon. Insect biochemistry and molecular biology 43, 237–251 (2013).
Zhang, Y.-N. et al. Identification and Expression Profiles of Sex Pheromone Biosynthesis and Transport Related Genes in Spodoptera litura. PloS one. 10, e0140019 (2015).
Xie, W. et al. Tissue-specific transcriptome profiling of Plutella xylostella third instar larval midgut. Int J Biol Sci 8, 1142–1155 (2012).
Gong, D.-P. et al. Identification and expression pattern of the chemosensory protein gene family in the silkworm, Bombyx mori. Insect biochemistry and molecular biology 37, 266–277 (2007).
Wang, Z.-y. et al. The categories of tobacco storage pests and their natural enemies in Guangxi Province [J]. Guangxi Agricultural Sciences 3, 014 (2007).
Mittapalli, O., Wise, I. L. & Shukle, R. H. Characterization of a serine carboxypeptidase in the salivary glands and fat body of the orange wheat blossom midge, Sitodiplosis mosellana (Diptera: Cecidomyiidae). Insect biochemistry and molecular biology 36, 154–160 (2006).
Deng, Xl et al. The discovery of a novel antagonist–Manduca sexta allatotropin analogue–as an insect midgut active ion transport inhibitor. Pest management science 72, 2176–2180 (2016).
Li, F. et al. Effects of phoxim on nutrient metabolism and insulin signaling pathway in silkworm midgut. Chemosphere 146, 478–485 (2016).
Simpson, R. et al. Expressed sequence tags from the midgut of Epiphyas postvittana (Walker)(Lepidoptera: Tortricidae). Insect molecular biology 16, 675–690 (2007).
Coates, B. S., Sumerford, D. V., Hellmich, R. L. & Lewis, L. C. Mining an Ostrinia nubilalis midgut expressed sequence tag (EST) library for candidate genes and single nucleotide polymorphisms (SNPs). Insect molecular biology 17, 607–620 (2008).
Pauchet, Y. et al. Pyrosequencing the Manduca sexta larval midgut transcriptome: messages for digestion, detoxification and defence. Insect molecular biology 19, 61–75 (2010).
Wu, Z. et al. Discovery of chemosensory genes in the oriental fruit fly, Bactrocera dorsalis. PloS one. 10, e0129794 (2015).
Zhang, G.-N., Hu, F., Dou, W. & Wang, J.-J. Morphology and distribution of sensilla on tarsi and ovipositors of six fruit flies (Diptera: Tephritidae). Annals of the Entomological Society of America 105, 319–327 (2012).
Yi, X. et al. BdorCSP2 is important for antifeed and oviposition-deterring activities induced by Rhodojaponin-III against Bactrocera dorsalis. PloS one. 8, e77295 (2013).
Xue, W. et al. Identification and Expression Analysis of Candidate Odorant-Binding Protein and Chemosensory Protein Genes by Antennal Transcriptome of Sitobion avenae. PloS one. 11, e0161839 (2016).
Li, Z.-Q. et al. Expression Analysis and Binding Assays in the Chemosensory Protein Gene Family Indicate Multiple Roles in Helicoverpa armigera. Journal of chemical ecology 41, 473–485 (2015).
Kitabayashi, A. N., Arai, T., Kubo, T. & Natori, S. Molecular cloning of cDNA for p10, a novel protein that increases in the regenerating legs of Periplaneta americana (American cockroach). Insect biochemistry and molecular biology 28, 785–790 (1998).
Maleszka, J., Foret, S., Saint, R. & Maleszka, R. RNAi-induced phenotypes suggest a novel role for a chemosensory protein CSP5 in the development of embryonic integument in the honeybee (Apis mellifera). Development genes and evolution 217, 189–196 (2007).
Liu, Y.-L. et al. Unique function of a chemosensory protein in the proboscis of two Helicoverpa species. Journal of Experimental Biology 217, 1821–1826 (2014).
Matsuo, T. Genes for host-plant selection in Drosophila. Journal of neurogenetics 22, 195–210 (2008).
Liu, G. X. et al. Biotype expression and insecticide response of Bemisia tabaci Chemosensory Protein‐1. Archives of insect biochemistry and physiology 85, 137–151 (2014).
Xuan, N. et al. Increased expression of CSP and CYP genes in adult silkworm females exposed to avermectins. Insect science. 22, 203–219 (2015).
Zhu, J. et al. Conserved chemosensory proteins in the proboscis and eyes of Lepidoptera. International Journal of Biological Sciences 12, 1394 (2016).
Nagnan-Le Meillour, P. et al. Chemosensory proteins from the proboscis of Mamestra brassicae. Chemical Senses. 25, 541–553 (2000).
Acknowledgements
This study was funded by Science and Technology Planning Project of Guangdong Province (2016A020210090) and Youth Science and technology training project of South China Agricultural University.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: X.Y., X.F.Z., M.Y.H., G.H.Z. Performed the experiments: X.Y., J.W.Q. Analyzed the data: X.Y., J.W.Q., M.H., G.H.Z. Wrote the paper: X.Y., X.F.Z., G.H.Z.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Yi, X., Qi, J., Zhou, X. et al. Differential expression of chemosensory-protein genes in midguts in response to diet of Spodoptera litura . Sci Rep 7, 296 (2017). https://doi.org/10.1038/s41598-017-00403-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-00403-5
- Springer Nature Limited
This article is cited by
-
De novo transcriptome of the whole-body of the gastropod mollusk Philomycus bilineatus, a pest with medical potential in China
Journal of Applied Genetics (2020)
-
Characterizing potential repelling volatiles for “push-pull” strategy against stem borer: a case study in Chilo auricilius
BMC Genomics (2019)
-
Transcriptome and gene expression analysis of three developmental stages of the coffee berry borer, Hypothenemus hampei
Scientific Reports (2019)