
Abstract
Behçet’s disease (BD) is reportedly associated with polymorphisms of the ubiquitin-associated domain containing 2 (UBAC2) gene in Turkish, Italian, and Chinese populations. Here we investigated whether UBAC2 polymorphisms were associated with BD in a Japanese population. Using data from 611 Japanese BD patients and 737 Japanese controls who participated in our previous genome-wide association study, we analyzed the 58 genotyped single-nucleotide polymorphisms (SNPs) in the region 100 kb upstream and downstream of UBAC2. We also performed imputation analysis in the region, with 562 imputed SNPs included in the statistical analyses. Association testing revealed that the T allele of rs9517723 in the lncRNA LOC107984558 was significantly associated with ocular and central nervous system (CNS) lesions and showed the strongest association under the recessive model (TT vs. CT+CC: ocular lesion, Pc = 0.0099, OR = 1.56; CNS lesion, Pc = 0.0052, OR = 3.42). Expression analysis revealed that rs9517723 TT homozygotes showed significantly increased UBAC2 expression (P < 0.05). Our findings suggest that enhanced UBAC2 expression associated with the homozygous risk allele (TT) of rs9517723 could induce overactivation of ubiquitination-related pathway, resulting in the development of ocular and CNS lesions in BD.
Similar content being viewed by others

Introduction
Behçet’s disease (BD) is a rare, chronic, systemic, inflammatory disorder characterized by recurrent ocular symptoms, oral and genital ulcers, and skin lesions. In some cases, BD is associated with inflammation throughout the body, for example, in the joints, vascular system, lungs, gastrointestinal tract, central nervous system, and epididymis1. BD has been diagnosed worldwide, although it is most commonly found in Mongoloid populations, and rarely in Caucasian populations, and it shows a particularly high prevalence in countries along the ancient Silk Route from Japan to the Middle East and the Mediterranean basin2.
Although the etiology of BD remains unclear, several external environmental factors appear to trigger BD in individuals of a particular genetic background. The human leucocyte antigen (HLA) HLA-B*51 is the major susceptibility gene responsible for BD3,4,5. However, HLA-B*51-negative patients can also develop BD, suggesting the involvement of other genetic factors.
BD is also reportedly associated with single-nucleotide polymorphisms (SNPs) of the ubiquitin-associated domain containing 2 (UBAC2) gene region on chromosome 13q32.3. Fei et al. performed a genome-wide association study (GWAS) using pooled DNA in a Turkish population, and initially reported that BD was potentially associated with a UBAC2 intronic SNP (rs9513584)6. Their further investigation of 14 SNPs in UBAC2 confirmed the association between UBAC2 and BD in the Turkish and Italian populations, and revealed that the intronic SNP rs7999348 was the most likely candidate among the genotyped SNPs7. Subsequently, Hou et al. performed a replication study with 25 SNPs in UBAC2, and reported that the promoter SNP rs3825427 showed the most significant association with BD in a Han Chinese population8. Overall, these findings suggest that UBAC2 variants are an important risk factor for BD susceptibility in multiple populations. However, the association between UBAC2 variants and BD has not yet been assessed in a Japanese population.
The aim of the present study was to investigate whether genetic variants in the UBAC2 region are associated with BD in a Japanese population. We performed a comprehensive association analysis of SNPs in the UBAC2 region among Japanese patients with BD.
Results
Comprehensive allelic association analysis
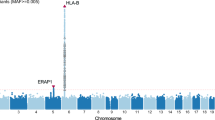
Our allelic association analysis in a Japanese population included a total of 620 SNPs (58 genotyped and 562 imputed). Of these SNPs, 100 SNPs showed an association with BD with a P value of <0.05 (Fig. 1). The strongest association was observed for rs9517723 located on LOC107984558, which showed an increased frequency of its major allele (T) in cases compared to controls (P = 0.0024; odds ratio (OR) = 1.27). However, this increase did not reach significance after correcting for multiple testing (Pc = 0.13) (Table 1). While the SNPs rs3825427, rs9513584 and rs7999348 were previously reported to be associated with BD, we found no such associations in our present population.

In-depth SNP analysis of the UBAC2 region. The lead SNP (rs9517723) is depicted as a purple diamond. The color coding of all other SNPs indicates linkage disequilibrium (LD) with the lead SNP: red, r2 ≥ 0.8; yellow, 0.6 ≤ r2 < 0.8; green, 0.4 ≤ r2 < 0.6; cyan, 0.2 ≤ r2 < 0.4; blue, r2 < 0.2; and gray, r2 is unknown. The left y axis represents the −log10 P values for allelic association with Behçet’s disease, and the right y axis represents the estimated recombination rate. The horizontal red line indicates the significance level of P = 0.05. Gene annotations are shown below the figure. The plot was created using LocusZoom.
LD analysis
We observed long-range LD across the UBAC2 gene region. The strongest signal was for rs9517723, which exhibited a strong or moderate LD (r2 ≥ 0.2) with many of the other 99 SNPs that showed a significant P value of < 0.05 (Fig. 1). However, rs9517723 exhibited very moderate or low LD (r2 < 0.2) with many of the remaining 520 SNPs with P values of >0.05. Among the 100 SNPs showing a significant P value of <0.05, we performed stepwise regression analysis to test the independence of multiple possible associations in the region. Conditioning by rs9517723 eliminated the association of the other 99 SNPs (P > 0.05), indicating that rs9517723 could account for most of the association of these SNPs with BD in the Japanese population. On the other hand, rs9517723 was in very moderate or low LD with the previously reported BD-associated SNPs (rs3825427: r2 = 0.07; rs9513584: r2 = 0.15; rs7999348: r2 = 0.10).
Association between rs9517723 and clinical symptoms of BD
We also analyzed the relationships between rs9517723 and clinical symptoms of BD (Table 2). An allelic association test in the Japanese population revealed that the T allele of rs9517723 was significantly associated with increased risk of ocular and central nervous system (CNS) lesions, with a stronger association with CNS lesions: ocular lesion, Pc = 0.018, OR = 1.36; CNS lesion, Pc = 0.0066, OR = 2.78. Table 3 shows the genotypic association results for rs9517723. In both additive and recessive models, we found an association with the disease at P < 0.05 among all entire Japanese patients; however, this association did not reach significance after correction (Pc > 0.05). On the other hand, the T allele of rs9517723 was significantly associated with increased risk of ocular and CNS lesions under the additive and recessive models. The OR was stronger among patients with CNS lesions, and the associations with both ocular and CNS lesions were stronger in the recessive model (ocular lesion: Pc = 0.0099, OR = 1.56; CNS lesion: Pc = 0.0052, OR = 3.42) than in the additive model (ocular lesion, Pc = 0.023, OR = 1.34; CNS lesion, Pc = 0.0078, OR = 2.94). Moreover, the T allele of rs9517723 was associated with 1.33-fold to 1.42-fold increased risks of oral ulcer, skin lesion, genital ulcer, and arthritis; however, these risks were not significant (P < 0.05, Pc > 0.05).
Expression analysis
The SNP rs9517723 is located on the first exon of LOC107984558, which encodes a long non-coding RNA (lncRNA) and is located between the protein coding genes, UBAC2 (43 kb downstream) and TM9SF2 (68 kb upstream) (Fig. 1). Through a variety of mechanisms, lncRNA can regulate gene expression in cis or in trans9, 10. Thus, we investigated whether rs9517723 affected the expression level of UBAC2 and/or TM9SF2. UBAC2 expression was significantly increased in the rs9517723 TT homozygotes (TT vs. CT, P = 0.020; TT vs. CC+CT, P = 0.019) (Fig. 2a). The rs9517723 genotype was not associated with TM9SF2 expression level (Fig. 2b).

Expression analysis of UBAC2 (a) and TM9SF2 (b) stratified by rs9517723 genotype.
Discussion
In our present study, we aimed to assess whether genetic variants in the UBAC2 region affected BD development in a Japanese population. We performed comprehensive association analysis of SNPs in the region 100 kb upstream and 100 kb downstream of UBAC2 among Japanese patients with BD. This study is the first comprehensive investigation of the UBAC2 region for association with BD. We found that SNP rs9517723 in the lncRNA LOC107984558 was recessively associated with the risks of ocular and CNS lesions (showing a stronger association with CNS lesions) but not with BD itself in the Japanese population. This suggests that rs9517723 contributes to the development of ocular and CNS lesions, especially CNS lesions, with a recessive effect.
UBAC2 encodes an ubiquitination-related structural domain that is implicated in ubiquitination and proteasomal degradation. BD is reportedly associated with other ubiquitination-related genes, including ubiquitin associated and SH3 domain containing B (UBASH3B)6, small ubiquitin-like modifier 4 (SUMO4)11,12,13, and ubiquitin-conjugating enzyme E2Q family-like 1 (UBE2QL1)14, suggesting that the ubiquitination-related pathway may be involved in BD pathophysiology. In our present study, we found that the homozygous risk allele (TT) of the lncRNA LOC107984558 rs9517723 was significantly correlated with enhanced UBAC2 expression. Sawalha et al. also showed that UBAC2 expression was significantly increased in the homozygous risk allele of a BD-associated SNP (the GG genotype of rs7999348)7. While their study did not assess associations between the allele and clinical symptoms of BD7, their findings and our present results suggest that the enhanced UBAC2 expression associated with these homozygous risk alleles could induce overactivation of the ubiquitination-related pathway, resulting in the development of ocular and CNS lesions or of BD itself. On the other hand, Hou et al. showed that the risk allele of their identified BD-associated SNP (the T allele of rs3825427) led to downregulated UBAC2 expression8, suggesting that decreased UBAC2 expression also contributes to BD risk. These contradictory findings indicate a need for further functional studies to clarify how UBAC2 contributes to BD pathophysiology.
LncRNAs are defined as non-coding RNA transcripts of more than 200 nucleotides in length. Recent evidence shows that lncRNAs play key functional roles in diverse biological processes, including chromatin remodeling, transcriptional, posttranscriptional, and epigenetic regulation9, 10, 15. They also contribute to the pathophysiology of various diseases, including cancers and neurological, autoimmune, and ocular diseases15,16,17,18,19. Moreover, genetic variants in lncRNAs can modulate the structure and expression of localized lncRNAs, leading to functional alterations of their interacting partners20,21,22. The available data regarding lncRNAs may support our present findings that rs9517723 in the lncRNA LOC107984558 leads to enhanced UBAC2 expression, contributing to BD pathophysiology. In general, lncRNA expression is more tissue-specific than the expression of protein-coding genes, and most lncRNAs are highly expressed in the CNS with low expression in other tissues23,24,25. This characteristic expression pattern of lncRNAs may explain why rs9517723 in the lncRNA LOC107984558 is more strongly associated with CNS lesions than with other BD symptoms. However, we did not assess the expression pattern of LOC107984558 in our current study, nor did we find public databases containing LOC107984558 expression data.
In conclusion, our present findings indicate that rs9517723 in the lncRNA LOC107984558 was significantly associated with increased risks of ocular and CNS lesions within a Japanese population. Our results further suggest that rs9517723 is associated with enhanced UBAC2 expression, which contributes to the development of those lesions. Further validation studies in other ethnic populations are needed to confirm these findings. Additionally, further expression analyses using RNA isolated from cells of ocular and CNS lesions in patients with BD are needed to more clearly elucidate the effect of 9517723 on UBAC2 expression. In the future, rs9517723 may serve as a useful genetic marker for BD diagnosis, especially in cases with CNS lesions.
Methods
Subjects
Our previous GWAS enrolled 611 unrelated Japanese individuals with BD, and 737 unrelated Japanese controls26. Here we used genotype data from that study, specifically for the 58 SNPs found from 100 kb upstream to 100 kb downstream of the UBAC2. All 58 SNPs satisfied the following quality control criteria: a call rate >98%, Hardy-Weinberg equilibrium (HWE) P > 0.001, and minor allele frequency >1%. The Japanese patients were diagnosed with BD according to the standard criteria27 proposed by the Japan Behçet’s Disease Research Committee. All control participants were healthy volunteers, who were unrelated to each other or to the patients. All participants gave their written informed consent. The study methodology adhered to the tenets of the Declaration of Helsinki, and was approved by the Ethics Committee of Yokohama City University School of Medicine.
Imputation analysis of the UBAC2 gene region
To evaluate potential associations with un-genotyped SNPs within the region encompassing the UBAC2 gene, we performed imputation analysis. The genotypes of our Japanese GWAS set were imputed based on the 58 genotyped SNPs using MACH v1.0 (http://www.sph.umich.edu/csg/abecasis/MACH/index.html)28, 29. The reference panel comprised the 1000 Genomes Phase 3 datasets of 315 East Asian samples, which included a set of Japanese samples from Tokyo (JPT, N = 104), Han Chinese samples from Beijing (CHB, N = 103), and Southern Han Chinese samples (CHS, N = 108)) (http://www.1000genomes.org/)30. All imputed SNPs were filtered with the following quality control settings: HWE P > 0.001, minor allele frequency >1%, and a squared correlation between imputed and true genotypes (Rsq) of >0.90. A total of 562 imputed SNPs were included in further analysis.
Expression analysis
From our genome-wide expression (GWE) dataset, we obtained expression data for the UBAC2 and transmembrane 9 superfamily member 2 (TM9SF2) genes from 313 Japanese healthy volunteers (Meguro et al., unpublished data). The GWE analysis was performed using the Illumina HumanHT-12 v4 Expression BeadChip Kit. First, whole blood was collected from subjects in PAXgene Blood RNA tubes (Becton Dickinson, Heidelberg, Germany), and total RNA was extracted from whole blood using the PAXgene Blood RNA Kit (Qiagen) following the manufacturers’ protocols. Next, the total RNA samples were processed using the TargetAmp-Nano Labeling Kit for the Illumina Expression BeadChip (Epicentre, Wisconsin, USA) and hybridized to the BeadChips following the manufacturers’ protocols.
Statistical analysis
We performed allelic and genotypic association analyses and stepwise regression analyses, and calculated linkage disequilibrium (LD) using SNP & Variation Suite software version 8.6.0 (Golden Helix, Inc., Bozeman, MT, USA). A correlation/trend test was used to assess differences in allele and genotype frequencies between cases and controls. We generated a regional association plot for the UBAC2 region using LocusZoom (http://csg.sph.umich.edu/locuszoom/)31. Tagging SNPs were selected from the genotype data of the Japanese GWAS set (611 BD patients and 737 controls) with an r2 threshold of 0.80, using PLINK version 1.07 (http://pngu.mgh.harvard.edu/purcell/plink/)32. This identified 52 tagging SNPs, capturing all 620 SNPs (58 genotyped and 562 imputed SNPs) within the UBAC2 region. The obtained P values were corrected for multiple testing using Bonferroni correction based on the number of tagging SNPs. A corrected P (Pc) value of < 0.05 was considered significant. Differences in the expression levels of UBAC2 and TM9SF2 were analyzed using the Mann-Whitney U test.
References
Kaklamani, V. G., Vaiopoulos, G. & Kaklamanis, P. G. Behçet’s disease. Semin. Arthritis Rheum. 27, 197–217 (1998).
Ohno, S. et al. Close association of HLA-Bw51 with Behçet’s disease. Arch. Opthalmol. 100, 1455–1458 (1982).
Yabuki, K. et al. Association of MICA gene and HLA-B*5101 with Behçet’s disease in Greece. Invest. Ophthalmol. Vis. Sci. 40, 1921–1926 (1999).
Mizuki, N. et al. Analysis of microsatellite polymorphism around the HLA-B locus in Iranian patients with Behçet’s disease. Tissue Antigens. 60, 396–399 (2002).
Mizuki, N. et al. Association analysis between the MIC-A and HLA-B alleles in Japanese patients with Behçet’s disease. Arthritis Rheum. 42, 1961–1966 (1999).
Fei, Y. et al. Identification of novel genetic susceptibility loci for Behçet’s disease using a genome-wide association study. Arthritis Res. Ther. 11, R66 (2009).
Sawalha, A. H. et al. A putative functional variant within the UBAC2 gene is associated with increased risk of Behçet’s disease. Arthritis Rheum. 63, 3607–3612 (2011).
Hou, S. et al. Replication study confirms the association between UBAC2 and Behçet’s disease in two independent Chinese sets of patients and controls. Arthritis Res. Ther. 14, R70 (2012).
Mercer, T. R., Dinger, M. E. & Mattick, J. S. Long non-coding RNAs: insights into functions. Nat. Rev. Genet. 10, 155–159 (2009).
Vance, K. W. & Ponting, C. P. Transcriptional regulatory functions of nuclear long noncoding RNAs. Trends Genet. 30, 348–355 (2014).
Hou, S. et al. SUMO4 gene polymorphisms in Chinese Han patients with Behcet’s disease. Clin. Immunol. 129, 170–175 (2008).
Kamoun, M., Ben Dhifallah, I., Karray, E., Zakraoui, L. & Hamzaoui, K. Association of small ubiquitin-like modifier 4 (SUMO4) polymorphisms in a Tunisian population with Behçet’s disease. Clin. Exp. Rheumatol. 28, S45–49 (2010).
Park, G., Kim, H. S., Choe, J. Y. & Kim, S. K. SUMO4 C438T polymorphism is associated with papulopustular skin lesion in Korean patients with Behçet’s disease. Rheumatol. Int. 32, 3031–3037 (2012).
Remmers, E. F. et al. Genome-wide association study identifies variants in the MHC class I, IL10, and IL23R-IL12RB2 regions associated with Behçet’s disease. Nat. Genet. 42, 698–702 (2010).
Prensner, J. R. & Chinnaiyan, A. M. The emergence of lncRNAs in cancer biology. Cancer Discov. 1, 391–407 (2011).
Jain, S., Thakkar, N., Chhatai, J., Bhadra, M. P. & Bhadra, U. Long non-coding RNA: Functional agent for disease traits. RNA Biol. 1–14, doi:10.1080/15476286.2016.1172756 (2016).
Ng, S. Y., Lin, L., Soh, B. S. & Stanton, L. W. Long noncoding RNAs in development and disease of the central nervous system. Trends Genet. 29, 461–468 (2013).
Sigdel, K. R., Cheng, A., Wang, Y., Duan, L. & Zhang, Y. The Emerging Functions of Long Noncoding RNA in Immune Cells: Autoimmune Diseases. J. Immunol. Res. 2015, 848790 (2015).
Li, F., Wen, X., Zhang, H. & Fan, X. Novel Insights into the Role of Long Noncoding RNA in Ocular Diseases. Int. J. Mol. Sci. 17, 478 (2016).
Burd, C. E. et al. Expression of linear and novel circular forms of an ink4/arf-associated non-coding rna correlates with atherosclerosis risk. PLoS Genet. 32, e1001233 (2010).
Zhu, Z. et al. An insertion/deletion polymorphism within RERT-lncRNA modulates hepatocellular carcinoma risk. Cancer Res. 72, 6163–6172 (2012).
Chung, S. et al. Association of a novel long non-coding RNA in 8q24 with prostate cancer susceptibility. Cancer Sci. 102, 245–252 (2011).
Ravasi, T. et al. Experimental validation of the regulated expression of large numbers of non-coding RNAs from the mouse genome. Genome Res. 16, 11–19 (2006).
Mercer, T. R., Dinger, M. E., Sunkin, S. M., Mehler, M. F. & Mattick, J. S. Specific expression of long noncoding RNAs in the mouse brain. Proc. Natl. Acad. Sci. USA 105, 716–721 (2008).
Derrien, T. et al. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome Res. 22, 1775–1789 (2012).
Mizuki, N. et al. Genome-wide association studies identify IL23R-IL12RB2 and IL10 as Behçet’s disease susceptibility loci. Nat. Genet. 42, 703–706 (2010).
Mizushima, Y. Recent research into Behçet’s disease in Japan. Int. J. Tissue React. 10, 59–65 (1998).
Li, Y., Willer, C. J., Sanna, S. & Abecasis, G. Genotype Imputation. Annu. Rev. Genomics Hum. Genet. 10, 387–406 (2009).
Li, Y., Willer, C. J., Ding, J., Scheet, P. & Abecasis, G. R. MaCH: using sequence and genotype data to estimate haplotypes and unobserved genotypes. Genet. Epidemiol. 34, 816–834 (2010).
1000 Genomes Project Consortium, et al. An integrated map of genetic variation from 1,092 human genomes. Nature. 491, 56–65 (2012).
Pruim, R. J. et al. LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics. 26, 2336–2337 (2010).
Purcell, S. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575 (2007).
Acknowledgements
We sincerely thank all of the study participants, and all of the medical staff involved in diagnosis and sample collection.
Author information
Authors and Affiliations
Contributions
A.M., S.O. and N.M. designed the study. S.O. and N.M. contributed study samples. K.Y., A.M., M.T. and E.S. conducted the experiments and analyzed data. K.Y. and A.M. wrote the manuscript. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yamazoe, K., Meguro, A., Takeuchi, M. et al. Comprehensive analysis of the association between UBAC2 polymorphisms and Behçet’s disease in a Japanese population. Sci Rep 7, 742 (2017). https://doi.org/10.1038/s41598-017-00877-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-00877-3
- Springer Nature Limited
This article is cited by
-
Association between UBAC2 gene polymorphism and the risk of noise-induced hearing loss: a cross-sectional study
Environmental Science and Pollution Research (2022)
-
Association of Behçet disease with psoriasis and psoriatic arthritis
Scientific Reports (2021)
-
UBAC2 promotes bladder cancer proliferation through BCRC-3/miRNA-182-5p/p27 axis
Cell Death & Disease (2020)