
Abstract
Within mosquito vector populations, infectious mosquitoes are the ones completing the transmission of pathogens to susceptible hosts and they are, consequently, of great epidemiological interest. Mosquito infection by malaria parasites has been shown to affect several traits of mosquito physiology and behavior, and could interplay with the efficacy of control tools. In this study, we evaluated, in pyrethroid resistant Anopheles gambiae, the effect of mosquito infection with the human malaria parasite Plasmodium falciparum on the efficacy of nets treated with either the insecticide deltamethrin or the repellent DEET, measuring (i) mosquito success to pass through the net, (ii) blood-feeding on a host and (iii) chemicals-induced mortality. Infection of mosquitoes at non-infectious stage did not affect their success to pass through the net, to blood-feed, nor chemicals-induced mortality. At infectious stage, depending on replicates, infected mosquitoes had higher mortality rates than uninfected mosquitoes, with stronger effect in presence of DEET. This data evidenced a cost of infection on mosquito survival at transmissible stages of infection, which could have significant consequences for both malaria epidemiology and vector control. This stresses the need for understanding the combined effects of insecticide resistance and infection on the efficacy on control tools.
Similar content being viewed by others

Introduction
Mosquito-borne diseases are considerable public health issues, mostly affecting populations in developing countries1,2. To reduce their incidence, controlling vector mosquitoes and limiting their contact with human hosts remains the most effective strategy3. One promising approach for achieving this goal would be to selectively target, within mosquito populations, individuals that are the most dangerous for humans4. Particularly, mosquitoes carrying transmissible forms of pathogens in their salivary glands (i.e. infectious mosquitoes), are of great epidemiological relevance. Yet, the implementation of control means that could specifically reach infectious mosquitoes is dependent upon an extensive knowledge of the effects of mosquito infection on its behavior and physiology. Infection by pathogens has been shown to affect mosquito phenotypic traits5. Malaria-infected Anopheles mosquitoes generally display increase attraction, biting and feeding rate6,7,8,9,10. Interestingly, these changes are often concomitant with the presence of transmissible stages of the pathogen and may contribute to increase the number of contacts between human hosts and infectious vectors11. These effects may lead to substantial epidemiological consequences, with transmission rates of mosquito borne pathogens being potentially much higher than expected12. On the other hand, infection by malaria parasites may induce fitness costs on mosquitoes, reducing their survival. This can be especially true when infection is associated with other biotic and abiotic stresses13,14,15. Consequently, all the behavioral and physiological changes associated with infection could directly impact the efficacy of control means.
To date, conventional control tools mostly rely on the use of insecticide-treated nets (ITNs), although their efficacy could be threatened by the increase of insecticide resistance mechanisms in mosquito populations16. Besides, repellents such as DEET (N, N-diéthyl-3-méthylbenzamide) offer a great potential both in cutaneous application, spatial use or net impregnation, and their use may become complementary with ITNs17,18,19. Despite epidemiological relevance, the potential effect of infection on the efficacy of control means has received little attention. The effect of Plasmodium infection on the efficacy of control tools against Anopheles vectors has demonstrated contrasted results; while some observations suggest that it does not affect repellency and knockdown effects induced by permethrin20,21, a recent study showed that infection reduces personal protection offered by ITNs22. Another study also reported that infection can partially restore susceptibility to insecticide among mosquitoes carrying resistance alleles15. Besides, DEET- induced repellency was not found to be altered in Anopheles mosquitoes infected with Plasmodium20. Infection by Plasmodium could then lead mosquitoes to overcome the deterrent effects of control means, and/or induce cumulating fitness costs for the vector together with insecticides or repellents. Altogether, these results highlight the need to better understand the effects of mosquito infection on the efficacy of control tools.
In the present study, we evaluated the impact of infection by Plasmodium falciparum on the efficacy of treated nets, the most common control tools against malaria mosquitoes. Two compounds were selected: the pyrethroid insecticide deltamethrin, which is broadly used in net impregnation due to its disengagement and lethal effects on mosquitoes associated with a low toxicity to humans23, and DEET, the “gold standard” of insect repellents, which acts both as a spatial and contact repellent, and offer as well insecticidal properties24,25. In laboratory-controlled experiments, we measured Anopheles gambiae success to find a hole and to pass through a net treated with either deltamethrin or DEET, subsequent blood-feeding on a host, and chemicals-induced mortality.
Results
An. gambiae mosquitoes of 3 to 5 days old carrying pyrethroid-resistance alleles were experimentally infected with P. falciparum-containing blood collected in human participants in Bouaké, Ivory Coast. Prevalence of infection varied between 15 and 92% across 14 experimental infections, with a mean total prevalence of 69.77% ± 5.88. Dissection at six to seven days post-infective blood meal showed oocysts numbers per female ranging from 1 to 351 across experimental infections. A subset of mosquitoes exposed to inactivated P. falciparum- containing blood (and then considered as non-infectious) was also tested for the presence/absence of P. falciparum. None of these 11 tested mosquitoes were found positive for the presence of the parasite. Behavioral experiments were performed on mosquitoes at 6–8 days post blood-meal (6–8 dpbm), when infected individuals were carrying non-transmissible stages in their midgut (i.e. oocysts), and at 12–14 days post blood-meal (12–14 dpbm), when infected individuals were carrying transmissible stages of the parasite in their salivary glands (i.e. sporozoites). The pyrethroid insecticide deltamethrin and the synthetic repellent DEET were tested for their efficacy to reduce mosquito success to find a hole and to pass through a treated net, blood-feeding on a host after passing through the net, as well as chemical-induced mortality. For each behavioral replicate and chemical, four groups of mosquitoes were tested simultaneously with the following treatments: (i) unexposed to infection - control net (NI-Control), (ii) unexposed to infection - treated net (NI-Delta or NI-DEET), (iii) exposed to infection - control net (I-Control) and iv) exposed to infection - treated net (I-Delta or I-DEET). For deltamethrin, 1,053 mosquitoes were tested across 3 behavioral replicates at 6–8 dpbm and 3 at 12–14 dpbm using blood samples from 4 donors. For DEET, 1,687 mosquitoes were tested across 5 behavioral replicates at 6–8 dpbm and 8 at 12–14 dpbm using blood samples from 10 donors.
Effect of mosquito infection on efficacy of deltamethrin-impregnated nets
Passing rate was not affected by infection at 6–8 dpbm nor at 12–14 dpbm (Table 1, Fig. 1A). Deltamethrin significantly reduced mosquito passing rate both at 6–8 dpbm (mean passing rate = 75.42% ± 7.71 in NI-Control, 38.80% ± 10.42 in NI-Delta, 65.13% ± 18.49 in I-Control and 49.58% ± 11.19 in I-Delta) and at 12–14 dpbm (72.08% ± 13.04 in NI-Control, 48.79% ± 6.90 in NI-Delta, 76.85% ± 14.93 in I-Control and 56.07% ± 6.28 in I-Delta) (Table 1; Fig. 1A).

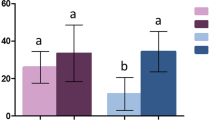
Passing rate through deltamethrin-impregnated nets (A), blood-feeding (B) and mortality associated (C). Blood-feeding was calculated for females that successfully passed through the impregnated net. Hatched bars show infected females, and full bars show non – infected ones. I = Infected, NI = Non-Infected. Results are presented as mean ± standard error (s.e). Different letters indicate significant differences (post hoc chi-squared tests with a Tukey correction).
Blood-feeding success was not significantly affected by mosquito infection at 6–8 dpbm, but was significantly reduced by deltamethrin treatment (mean blood-feeding = 92.24% ± 3.60 in NI-Control, 68.88% ± 12.50 in NI-Delta, 96.26% ± 1.97 in I-Control and 73.74% ± 4.40 in I-Delta). At 12–14 dpbm, a significant interaction was found between infection and treatment regarding this trait. Blood-feeding was significantly reduced by deltamethrin treatment (86.87% ± 8.44 in NI-Control, 47.27% ± 6.94 in NI-Delta, 98.55% ± 1.44 in I-Control and 46.55% ± 7.65 in I-Delta). However, paired comparisons showed only significant differences in blood-feeding due to the deltamethrin treatment and no difference between infected and uninfected females, neither upon control nets (P = 0.16) nor upon deltamethrin-treated nets (P = 0.92) (Table 1; Fig. 1B).
Mortality was not significantly affected by infection neither at 6–8 dpbm nor at 12–14 dpbm. Mortality was significantly increased in deltamethrin treatment both at 6–8 dpbm (mean mortality = 3.34% ± 1.86 in NI-Control, 18.56% ± 7.81 in NI-Delta, 6.72% ± 0.42 in I-Control and 20.19% ± 6.34 in I-Delta) and at 12–14 dpbm (8.17% ± 4.40 in NI-Control, 41.77% ± 4.47 in NI-Delta, 3.66% ± 1.90 in I-Control and 36.29% ± 15.87 in I-Delta) (Table 1; Fig. 1C).
Effect of mosquito infection on efficacy of DEET-impregnated nets
Infection did not significantly affect passing rate, neither at 6–8 dpbm nor at 12–14 dpbm. Passing rate was only significantly reduced in DEET treatment both at 6–8 dpbm (mean passing rate = 70.39% ± 11.82 in NI-Control, 36.04% ± 11.65 in NI-DEET, 83.78% ± 7.70 in I-Control and 29.97% ± 12.87 in I-DEET) and at 12–14 dpbm (81.52% ± 7.60 in NI-Control, 26.61% ± 5.60 in NI-DEET, 70.56% ± 8.89 in I-Control and 35.86% ± 7.33 in I-DEET) (Table 2; Fig. 2A).

Passing rate through DEET-impregnated nets (A), blood-feeding (B) and mortality (C). Blood-feeding was calculated for females that successfully passed through the impregnated net. Hatched bars show infected females, and full bars show non – infected ones. I = Infected, NI = Non-Infected. Results are presented as mean ± standard error (s.e). Different letters indicate significant differences (post hoc chi-squared tests with a Tukey correction).
At 6–8 dpbm, blood-feeding was not significantly affected by infection, but was significantly reduced by DEET treatment (mean blood-feeding = 91.13% ± 5.88 in NI-Control, 72.57% ± 16.11 in NI-DEET, 91.06% ± 4.06 in I-Control and 68.57% ± 18.29 in I-DEET). At 12–14 dpbm, blood-feeding was significantly reduced both by DEET treatment and by infection. Paired comparisons showed however no statistical difference between non-infected females and infected when exposed to control nets (86.17% ± 4.11 in NI-Control and 69.17% ± 11.83 in I-Control, P = 0.063), and when exposed to DEET-treated nets (46.19% ± 13.46 in NI-DEET and 40.45% ± 12.17 in I-DEET, P = 0.098) (Table 2; Fig. 2B).
At 6–8 dpbm, mortality was not affected by infection nor by treatment (mean mortality = 3.31% ± 1.83 in NI-Control, 2.07% ± 2.07 in I-Control, 6.30% ± 4.06 in NI-DEET and 6.23% ± 3.70 in I-DEET). At 12–14 dpbm, infection showed significant effect on mortality, irrespectively of net treatment. While paired comparisons showed that infectious females displayed higher mortality when exposed to DEET that non-infected ones (11.79% ± 5.07 in NI-DEET and 34.83% ± 7.80 in I-DEET, P = 0.023), no difference was observed between infected females exposed to control net and non-infected ones (10.78% ± 4.82 in NI-Control and 21.44% ± 10.41 in I-Control, P = 0.20) (Table 2; Fig. 2C). This suggest that infection-induced mortality is higher when infectious females face DEET-impregnated nets. In order to assess whether mortality affects females before or after blood-meal, we compared mortality between non-infected and infected females among both blood-fed females and unfed ones. Infection did not significantly affect mortality among blood-fed females (X² = 0.046, Df = 1, P = 0.83). However, it did show significant impact among the unfed group (X² = 9.58, Df = 1, P = 0.0020), suggesting that mortality occurred during host seeking.
Discussion
The results presented showed no effect of Plasmodium infection in mosquitoes on their ability to pass through a hole of a deltamethrin-impregnated net, to blood-feed after passing through the net, nor on their mortality. This is consistent with previous observations that did not find any impact of Plasmodium infection on pyrethroid efficacy20,26. However, it seems to contradict the recent published results that showed decreases in the efficacy of ITNs against infectious mosquitoes22. Some differences in the design of the assays may be responsible for these divergences. First, the chemicals used are different. They belong to two distinct classes of pyrethroids and are known to induce different effects on mosquito nervous system27, deltamethrin (used in the present study) being less irritant after contact than permethrin (used in the previous study)23. Then, we used a homozygous kdr An. gambiae strain whereas the previous study used an An. arabiensis strain partially resistant to pyrethroids. The use of specific laboratory strains under different rearing conditions could have impacted the observed results. Moreover, An. arabiensis has been shown to be more irritated by pyrethroids than An. gambiae when both strain carry insecticide resistance alleles28, and kdr mutation is also known to decrease the irritant effects of pyrethroids28,29. The observed effects of infection on pyrethroids efficacy in the previous study22 may then be impacted by the generalized insecticide resistance in vector populations30. The present data offer a complementary view on the effects on infection on control tools efficacy in a kdr homozygous population. Also, our results highlight that, although mosquitoes were carrying pyrethroid resistant alleles, deltamethrin remain effective to some extent to prevent them entering the net and to induce mortality.
The experiment testing the interactions between DEET impregnation on nets and infection status of mosquitoes showed that the presence of oocysts in mosquito midgut did not influence passing rate through the net, blood-feeding, nor mortality. In the same way, when mosquitoes harbored sporozoites, passing rate was not affected by infection under our experimental design. However, mosquitoes harboring sporozoites displayed an overall increased mortality, especially in presence of DEET. This effect of infectious parasite stages on mosquito mortality was not observed in the deltamethrin dataset, in presence and absence of the chemical. It is worth noting that a high variability was observed between replicates in the DEET dataset, so the increased mortality was not evidenced in each assay. This could be an effect of the variation in the number of mosquitoes released in the different tunnels between replicates, of blood donor, which has been shown to modulate the impact of infection on mosquito fitness13, and of parasite isolates.
The increased mortality observed in infectious mosquitoes in the DEET dataset, more markedly in presence of DEET, suggests that infection by Plasmodium could generate a fitness cost, affecting their survival. This is consistent with previous studies showing deleterious effects of infection in the Anopheles – Plasmodium system31,32,33. Although the cost of Plasmodium infection on mosquitoes is under debate for long time34,35, it appears that this cost might be expressed or amplified when associated to other stresses13,14 or to fitness costly mutations such as resistance alleles36. A stress induced by DEET could then enhance the cost of infection. Interestingly, the increased mortality was observed only in non-blood-fed females, suggesting that it occurred during host-seeking. This behavior is costly for mosquitoes37, and might interact synergistically with infection. Accordingly, infectious females were previously shown to display higher feeding-associated mortality, which could be a direct cost of the effort to host-seek38.
Our results show that, infection with Plasmodium does not affect ability of kdr resistant mosquitoes to find a hole and to pass through a net impregnated with deltamethrin. However, in some cases, infection increases mosquito mortality, especially in presence of DEET. Interestingly, when this fitness cost is expressed, it is observable only in females that harbor sporozoite stages of the parasite. This may lead to important epidemiological consequences. Yet, insecticide-resistant and infectious mosquitoes are the priority targets in the fight against mosquito-borne diseases. However, if insecticide resistant mosquitoes are more likely to die when infectious and if mortality occurs during host-seeking, transmission rates might be lower than expected. Moreover, due to their higher probability to die, those mosquitoes may be reached more easily by control tools. Altogether these observations reinforce the idea of targeting specifically infectious mosquitoes in the fight against mosquito borne diseases39. This could be achieved by using both lower doses of chemicals that only target infectious mosquitoes and biological control agents that act against late-life stages40,41. Here, DEET appears as a candidate to affect foraging behavior of epidemiologically relevant infectious females. This is strengthened by the presence a cost of DEET exposure on life-history traits in Anopheles gambiae females that we previously identified42. Nonetheless, the fact that the increased mortality we observe varied depending on replicates or studies suggests complex interactions between environment, control tools, infection and insecticide resistance. Understanding these interplays is a keystone for successfully targeting within a vector population individuals that are the more susceptible to transmit pathogens.
Methods
Mosquito strain and P. falciparum experimental infection of mosquitoes by Direct Membrane Feeding Assays (DMFA)
Experiments were performed on Anopheles gambiae. Pyrethroid resistance is widespread among malaria vector populations worldwide30 and the frequency of resistant alleles is particularly high in the study area43. Therefore, we used a KdrKis strain, fixed for the kdr-west allele, which confers resistance to pyrethroids and DDT. The colony was obtained by introgression into the Kisumu genome the kdr-west allele obtained from pyrethroid resistant mosquitoes in Kou Valley, Burkina Faso44. Individuals were reared at 24–28 °C and 55–80% relative humidity, with a light: dark photoperiod of 12:12 h. Larvae were reared in groups of about 300 in 1 L of distilled water and were fed TetraMin® fish food. Adults were fed a 10% honey solution for sugar intake and were maintained on guinea pigs for blood intake.
DMFAs were carried out according to procedures previously described by Ouédraogo et al.45. Screening for P. falciparum infectious human carriers were conducted in primary school groups in Bouaké, Ivory Coast. Ethical approval for the use of experimental animals and human participants was obtained from the Ministry of Health and Fight against AIDS in Ivory Coast through the National Ethic Committee N° 063/MSHP/CNER-kp. All human participants were enrolled after receipt of written informed consent from their legal guardians. All methods were performed in accordance with the relevant guidelines and regulations.
After collection of gametocyte-containing blood, plasma was removed and replaced with European naive AB serum. Batches of 3–5 days old mosquito females were allowed to feed either infectious blood or heat-treated (non-infectious) blood46. After one hour of exposure, unfed females were discarded and only fully fed mosquitoes were kept and maintained in the same conditions as during the rearing. Fed females were given the opportunity to oviposit. A total of 14 infectious blood samples from distinct blood donors were used.
Six to seven days after blood meal, infection rate among exposed mosquitoes was assessed in a subset of 10 females per infectious blood sample. Midguts were dissected in 0.4% mercurochrome (Sigma-Aldrich) and examined by light microscopy for the presence and number of oocysts.
Nets impregnation and behavioral assays in tunnel tests
Pieces of polyester nets of 25 × 25 cm side were treated with either the pyrethroid insecticide deltamethrin or the synthetic repellent DEET at a dose of 25 mg/m² and 500 mg/m² respectively. Concentrations were selected to induce 25 to 75% of mean blood-feeding inhibition in order to allow for observable differences of treatment efficacy between groups of mosquitoes according to infection status. The dose used for deltamethrin also corresponds to the recommended dose for conventional net treatment47,48. Deltamethrin and DEET were respectively dissolved in acetone and ethanol and tested independently, using their respective solvents as controls.
Behavioral experiments were performed using tunnel tests. The equipment consists of four square glass tunnels each divided in two chambers29,49. In one chamber, a male guinea pig from the colony used for mosquito rearing was placed as a bait. The two tunnel compartments were separated by a cardboard frame covered with the impregnated net (25 × 25 cm). Nets were deliberately holed by nine 1 cm diameter equidistant holes to give opportunity for mosquitoes to pass from the release chamber to the second chamber containing the guinea pig. For each behavioral replicate and chemical, four groups of mosquitoes were tested simultaneously in four different tunnels with the following conditions: (i) unexposed to infection - control net (NI-Control), (ii) unexposed to infection - treated net (NI-Delta or NI-DEET), (iii) exposed to infection - control net (I-Control) and (iv) exposed to infection - treated net (I-Delta or I-DEET). The same number of females (from 10 to 120) were released in the four tunnels. Treatments were randomly attributed to each of the four tunnels at each replicate. Each behavioral replicate corresponds to an experimental infection from one given donor. For each chemical, part of the females were tested at 6–8 days post blood-meal (6–8 dpbm), when infected individuals carried oocysts in their midgut (i.e. immature, non-infectious stage of the parasite). The other part was tested at 12–14 days post blood-meal (12–14 dpbm), when infected individuals carried sporozoites in their salivary glands (i.e. mature, infectious stages of the parasite). Exposure lasted eight hours, in order to mimic one night of sleeping under a treated net, after which the position of each female mosquito in the tunnel compartments was recorded, as well as mortality at the end of the test, and blood-feeding status.
Determination of infection status
After each behavioral assay, all females exposed to infectious blood meal and a subset of females exposed to non-infectious blood were tested for their infection status. They were conserved and stored individually in 100 µl of DNAzol® reagent (Molecular Research Center, Inc, Cincinnati, OH, USA). The entire insect was used for females killed 6–8 dpbm. Only the cephalothoraxes were used for females killed 12–14 dpbm; this allows to detect infected mosquitoes at days 6 to 8 and exclusively the presence of sporozoites stages at days 12 to 14. DNA extraction from individual mosquitoes was performed using DNAzol® according to the manufacturer’s instructions. P. falciparum detection was then carried out by qPCR50,51. Females were considered positive for P. falciparum when the Cq (quantification cycle) ranged from 25 to 35 and when the Tm (primer melting temperature) ranged from 75.5 to 80.
Statistical Analysis
All statistical analyses were performed using R software52. Only females positive for the presence of P. falciparum were included in statistical analysis and hereafter mentioned as “infected” females. Control females exposed to non-infectious blood are hereafter called “non-infected”. DEET and deltamethrin datasets were analyzed independently, using the same statistical method. For each chemical, logistic regression by generalized linear model (glm, quasi-binomial distribution, logit link) was used to compare between infected and uninfected females: (i) the proportion of females that successfully passed through the impregnated net, (ii) the proportion of blood-fed females among those that passed the net, and (iii) the proportion of dead females. Analysis were performed separately for females at 6–8 dpbm and for females at 12–14 dpbm. Treatment (control versus chemical), infection status (infected versus non-infected), their interaction, and replicates were coded as fixed factors. Outputs of the models were obtained using type 2 ANOVA (car package53). Results are presented as mean ± standard error (SE).
Data Availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request54.
References
WHO. Global vector control response 2017–2030 (2017).
WHO. A global brief on vector-borne diseases (2014).
Bhatt, S. et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 526, 207–211 (2015).
Koella, J. C., Lynch, P. A., Thomas, M. B. & Read, A. F. Towards evolution-proof malaria control with insecticides. Evol. Appl. 2, 469–480 (2009).
Stanczyk, N. M., Mescher, M. C. & De Moraes, C. M. Effects of malaria infection on mosquito olfaction and behavior: extrapolating data to the field. Curr. Opin. Insect Sci. 20, 7–12 (2017).
Rossignol, P. A., Ribeiro, J. M. C. & Spielman, A. Increased biting rate and reduced fertility in sporozoite-infected mosquitoes. Am. J. Trop. Med. Hyg. 35, 277–279 (1986).
Wekesa, J. W., Copeland, R. S. & Mwangi, R. W. Effect of Plasmodium falciparum on blood feeding behavior of naturally infected Anopheles mosquitoes in Western Kenya. Am. J. Trop. Med. Hyg. 47, 484–488 (1992).
Koella, J. C., Sørensen, F. L. & Anderson, R. A. The malaria parasite, Plasmodium falciparum, increases the frequency of multiple feeding of its mosquito vector, Anopheles gambiae. Proc. Biol. Sci. 265, 763–8 (1998).
Smallegange, R. C. et al. Malaria infected mosquitoes express enhanced attraction to human odor. PLoS One 8, e63602 (2013).
Cator, L. J., Lynch, P. A., Read, A. F. & Thomas, M. B. Do malaria parasites manipulate mosquitoes? Trends Parasitol. 28, 466–470 (2012).
Anderson, R. A., Koella, J. C. & Hurd, H. The effect of Plasmodium yoelii nigeriensis infection on the feeding persistence of Anopheles stephensi Liston throughout the sporogonic cycle. Proc. R. Soc. B 266, 1729–1733 (1999).
Cator, L. J., Lynch, P. A., Thomas, M. B. & Read, A. F. Alterations in mosquito behaviour by malaria parasites: potential impact on force of infection. Malar. J. 13 (2014).
Sangare, I. et al. Stress dependent infection cost of the human malaria agent Plasmodium falciparum on its natural vector Anopheles coluzzii. Infect. Genet. Evol. 25, 57–65 (2014).
Aboagye-antwi, F. et al. Hydric stress-dependent effects of Plasmodium falciparum infection on the survival of wild- caught Anopheles gambiae female mosquitoes. Malar. J. 9, 1–12 (2010).
Alout, H. et al. Interplay between Plasmodium infection and resistance to insecticides in vector mosquitoes. J. Infect. Dis. 210, 1464–70 (2014).
Labbé, P., Alout, H., Djogbénou, L., Pasteur, N. & Weill, M. Evolution of resistance to insecticide in disease vectors. In Genetics and evolution of infectious diseases (ed. Michel Tibayrenc) 363–409, https://doi.org/10.1016/B978-0-12-384890-1.00014-5 (Elsevier Publishing Compagny, 2011).
Achee, N. L. et al. Spatial repellents: from discovery and development to evidence-based validation. Malar. J. 11, 1–9 (2012).
Pennetier, C. et al. New protective battle-dress impregnated against mosquito vector bites. Parasit. Vectors 3, 1–7 (2010).
Pennetier, C. et al. Mixture for controlling insecticide- resistant malaria vectors. Emerg. Infect. Dis. 14, 1707–1714 (2008).
Robert, L. L., Schneider, I. & Wirtz, R. A. DEET and permethrin as protectants against malaria-infected and uninfected Anopheles stephensi mosquitoes. J. Am. Mosq. Control Assoc. 7, 304–306 (1991).
Glunt, K. D., Thomas, M. B. & Read, A. F. The effects of age, exposure history and malaria infection on the susceptibility of Anopheles mosquitoes to low concentrations of pyrethroid. 6 (2011).
Thiévent, K. et al. Malaria infection in mosquitoes decreases the personal protection offered by permethrin-treated bednets. Parasit. Vectors 11, 1–10 (2018).
Siegert, P. Y., Walker, E. & Miller, J. R. Differential behavioral responses of Anopheles gambiae (Diptera: Culicidae) modulate mortality caused by pyrethroid-treated bednets. J. Econ. Entomol. 102, 2061–2071 (2009).
Licciardi, S., Herve, J., Darriet, F., Hougard, J.-M. & Corbel, V. Lethal and behavioural effects of three synthetic repellents (DEET, IR3535 and KBR 3023) on Aedes aegypti) mosquitoes in laboratory assays. Med. Vet. Entomol. 20, 288–293 (2006).
Leal, W. S. The enigmatic reception of DEET — the gold standard of insect repellents. Curr. Opin. Insect Sci. 6, 93–98 (2014).
Glunt, K. D., Thomas, M. B. & Read, A. F. The effects of age, exposure history and malaria infection on the susceptibility of Anopheles mosquitoes to low concentrations of pyrethroid. PLoS One 6, e24968 (2011).
Breckenridge, C. B. et al. Evidence for a separate mechanism of toxicity for the Type I and the Type II pyrethroid insecticides. Neurotoxicology 30, 17–31 (2009).
Kawada, H. et al. Insecticidal and repellent activities of pyrethroids to the three major pyrethroid-resistant malaria vectors in western Kenya. Parasit. Vectors 7 (2014).
Chandre, F. et al. Modifications of pyrethroid effects associated with kdr mutation in Anopheles gambiae. Med. Vet. Entomol. 14, 81–88 (2000).
WHO. World malaria report 2017. (WHO, 2017).
Gad, A. M., Maier, W. A. & Piekarski, G. Pathology of Anopheles stephensi after infection with Plasmodium berghei berghei. Parasitol. Res. 60, 249–261 (1979).
Klein, T. A., Harrison, B. A., Andre, R. G., Whitmire, R. E. & Inlao, I. Detrimental effects of Plasmodium cynomolgi infections on the longevity of Anopheles dirus. Mosq. News 42, 265–271 (1982).
Ferguson, H. & Read, A. F. Genetic and environmental determinants of malaria parasite virulence in mosquitoes. Proc. R. Soc. B 269, 1217–1224 (2002).
Ferguson, H. M. & Read, A. F. Why is the effect of malaria parasites on mosquito survival still unresolved? Trends Parasitol. 18, 256–261 (2002).
Cohuet, A., Harris, C., Robert, V. & Fontenille, D. Evolutionary forces on Anopheles: what makes a malaria vector? Trends Parasitol. 26, 130–136 (2010).
Alout, H., Dabiré, R. K., Djogbénou, L. S., Abate, L. & Corbel, V. Interactive cost of Plasmodium infection and insecticide resistance in the malaria vector Anopheles gambiae. Sci. Rep. 6, 1–11 (2016).
Lyimo, I. N. & Ferguson, H. M. Ecological and evolutionary determinants of host species choice in mosquito vectors. Trends Parasitol. 25 (2009).
Anderson, R. A., Knols, B. G. J. & Koella, J. C. Plasmodium falciparum sporozoites increase feeding-associated mortality of their mosquito hosts Anopheles gambiae s. l. Parasitology 120, 329–333 (2000).
Alout, H., Labbé, P., Chandre, F. & Cohuet, A. Malaria vector control still matters despite insecticide resistance. Trends Parasitol. 33, 610–618 (2017).
Read, A. F., Lynch, P. A. & Thomas, M. B. How to make evolution-proof insecticides for malaria control. PLoS Biol. 7, 1–10 (2009).
Lynch, P. A., Grimm, U., Thomas, M. B. & Read, A. F. Prospective malaria control using entomopathogenic fungi: comparative evaluation of impact on transmission and selection for resistance. Malar. J. 11, 1–15 (2012).
Mulatier, M. et al. Effect of DEET-multiple exposures on behavior and life history traits in the malaria mosquito Anopheles gambiae (s.s.). Parasit. Vectors 11 (2018).
Camara, S. et al. Mapping insecticide resistance in Anopheles gambiae (s.l.) from Côte d’Ivoire. Parasit. Vectors 11 (2018).
Djégbe, I. Modifications physiologiques et comportementales induites par la résistance aux insecticides chez les vecteurs de Paludisme au Bénin (2013).
Ouédraogo, A. L. et al. A protocol for membrane feeding assays to determine the infectiousness of P. falciparum naturally infected individuals to Anopheles gambiae. Malar. World J. 4, 17–20 (2013).
Sangare, I. et al. Studying fitness cost of Plasmodium falciparum infection in malaria vectors: validation of an appropriate negative control. Malar. J. 12, 1 (2013).
Horn, K., Boecker, T., Grofmeyer, D., Nentwig, G. & Stoecker, R. Technology for conversion of conventional mosquito nets in the field into long lasting bed nets. In Proceedings of the Fifth International Conference on Urban Pests 287–294 (2005).
WHO. Insecticide-treated mosquito net interventions (2003).
Elissa, N. & Curtis, C. F. Evaluation of different formulations of deltamethrin in comparison with permethrin for impregnation of netting. Pestic. Sci. 44, 363–367 (1995).
Marie, A. et al. Evaluation of a real-time quantitative PCR to measure the wild Plasmodium falciparum infectivity rate in salivary glands of Anopheles gambiae. Malar. J. 12, 1–7 (2013).
Boissière, A. et al. Application of a qPCR Assay in the investigation of susceptibility to malaria infection of the M and S molecular forms of An. gambiae s. s. in Cameroon. PLoS One 8, e54820 (2013).
R Development Core Team. R: A language and environment for statistical computing (2008).
Fox, J. & Weisberg, S. An {R} companion to applied regression (2011).
Mulatier, M. Interaction entre l’état physiologique du moustique et le mode d’action des répulsifs chez les vecteurs de pathogènes. (Université de Montpellier, 2018).
Acknowledgements
The authors acknowledge all the participants involved in the study as well as the local authorities for their support. We are also very grateful to the IPR staff in Bouaké for technical assistance and to Félix Zoumènou and Awa Soro for administrative support. The manuscript has been previously published as a chapter of the PhD thesis of the first author. This work was supported by The French National Research Program for Environmental and Occupational Health of Anses [grant number EST-2016/1/39], by the Languedoc-Roussillon (LR)/Occitanie region and EU-FEDER under the name “Chercheur(se)s d′Avenir”, by the LabEx CeMEB, under an ANR “Investissements d’avenir” program [ANR-10-LABX-04-01] and by the French Initiative 5%—Expertise France [Grant Number No. 15SANIN213]. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
A.C., C.P., L.P.A.A., S.C., L.D. and M.M. designed the study. S.C., M.M., A.C. and D.C. performed data acquisition. M.M., A.P., T.L., D.C. and A.C. performed statistical analysis. M.M., S.C. and A.C. drafted the manuscript. C.P., A.C., T.L., N.M., R.D., F.C., A.K., L.P.A.A. and S.A. were responsible for financial and technical support. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mulatier, M., Camara, S., Koffi, A. et al. Efficacy of vector control tools against malaria-infected mosquitoes. Sci Rep 9, 6664 (2019). https://doi.org/10.1038/s41598-019-43195-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-43195-6
- Springer Nature Limited
This article is cited by
-
Attractive targeted sugar bait: the pyrrole insecticide chlorfenapyr and the anti-malarial pharmaceutical artemether–lumefantrine arrest Plasmodium falciparum development inside wild pyrethroid-resistant Anopheles gambiae s.s. mosquitoes
Malaria Journal (2023)
-
Compensatory mechanisms in resistant Anopheles gambiae AcerKis and KdrKis neurons modulate insecticide-based mosquito control
Communications Biology (2021)
-
Behavioural and Electrophysiological Responses of Female Anopheles gambiae Mosquitoes to Volatiles from a Mango Bait
Journal of Chemical Ecology (2020)