
Abstract
Soil organic carbon (SOC) dynamics after slash-burn agriculture are poorly understood in African tropical forest, though recent studies have revealed C4 grass invasion as a forest understory influences SOC dynamics after deforestation. This study aimed to quantify the relative SOC contribution of C4 and C3 plants separately through the sequential fallow periods of forest (cropland, or 4–7, 20–30, or >50 years of fallow forest) in the tropical forest of eastern Cameroon. We evaluated the SOC stock and natural 13C abundance for each layer. The SOC stock was largest in 4–7 years fallow forest (136.6 ± 8.8 Mg C ha−1; 100 cm depth, and C4:C3 = 58:42), and decreased with increasing fallow period. SOC from C4 plants was larger in the 4–7 and 20–30 years fallow forests (57.2–60.4 ± 5.8 Mg C ha−1; 100 cm depth), while it clearly decreased in >50 years fallow forest (35.0 ± 4.1 Mg C ha−1; 100 cm depth), resulting in the smallest SOC in this mature forest (106.4 ± 12.9 Mg C ha−1; 100 cm depth). These findings indicate that C4 grass understories contributed to the SOC restoration during early fallow succession in the tropical forest of eastern Cameroon.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
The sustainability of human societies depends on the wise use of natural resources, and soil ecosystems play an important role in this management1. To achieve the sustainable development goals (SDGs) related to food security, climate change, and sustainable use of terrestrial ecosystems, approved by the UN General Assembly in 2015, it is critical to improve the traditional management of soil resources, based on the understanding of its importance and efficiency. In tropical Africa, the traditional slash-burn agriculture (shifting cultivation) causes substantial variation of soil organic carbon (SOC) stock, which is an important indicator not only of soil fertility but also of climate change2,3,4. In general, cultivation after the slash-burn management decreases the SOC stock and soil fertility over time, whereas fallow management, which generally requires several decades or more for reforestation, restores the SOC stock and soil fertility. For the local farmers in the African tropical forest zone, natural reforestation for long periods is likely to be a useful and single management option to restore SOC5. Conversion from natural forest (or woodland) to cropland has been drastically increasing under population increase during the past century throughout tropical Africa6,7,8, causing shorter fallow periods, such as from >15–60 years in the past to <15 years in the present9, and subsequent soil degradation9,10. It is therefore necessary to evaluate the SOC variations of different periods of fallow sites, based on detailed mechanisms of SOC variations in African tropical forest to allow determination of the optimal future fallow period.
Restoration of SOC during the fallow period is mainly attributed to input of litter fall by growing trees and/or understory vegetation, such as herbs and shrubs11. Such restoration vegetation should belong to the natural vegetation succession from herbaceous, shrubs, and pioneer tree species at the early fallow stage to climax tree species at the later fallow stage. However, the effect of understories on SOC stock variations during the above natural fallow succession period remains unclear, especially in humid Africa12,13. Many studies have assessed the effect of fallow management, such as from cropland to bush or forest, on SOC stock in tropical areas, and most studies observed that fallow management clearly increased the SOC, especially in the dry to sub-humid tropics3,14. In contrast, Kotto-same et al.12 observed that there was little SOC stock fluctuation during the various fallow stages after cultivation in humid southern Cameroon, although they found an increase of aboveground biomass in forest with a longer fallow period.
Recently, many studies have evaluated the SOC dynamics separately from invaded grass-derived C (i.e., C4-plant grass) and forest-derived C (i.e., C3-plant forest), to discuss the effect of grass vegetation on SOC dynamics in humid tropical areas15,16,17. Yonekura et al.18,19 observed that C4 grass invasion after deforestation clearly increased the SOC stock, based on the increase in grass-derived SOC, using natural 13C measurements in a humid tropical area of Indonesia. Navarrete et al.20 also reported that conversion from forest to pasture in Colombia increased the SOC, mainly derived from C4 grass, under low-grazing-pressure pasture. They emphasised the importance of grass management practices, which affects the amount of C input by litter, to consider the accurate SOC dynamics in the humid tropics.
Although there is little quantitative information on the effect of grass invasion on SOC restoration during fallow succession in tropical Africa, C4 grass is one of the most popular forest understories, especially in the first stage of fallow succession after slash-burn management. Moreover, C4 grasses such as Pennisetum setaceum and Imperata cylindrica (invasive species) are widely distributed in the forest–savanna transition zone of Africa and continue to extend into the deforested zone following commercial logging21,22,23. This suggests that C4 grass invasion as a primary forest understory in the first stage of fallow succession should substantially contribute to the restoration of SOC stock; however, there is little information of the time-course SOC composition through the sequential fallow period, especially in African tropical reforested areas3,24,25.
In this study, we hypothesised that C4 grass invasion contributes to the restoration of the SOC stock, especially in the early stage of fallow succession in humid tropical Africa. To explore the importance of C4 grass understories to SOC stock variations at each fallow stage, we investigated the SOC stock in sites with different fallow periods. Our main objective was to assess the impact of C4 grass and C3 forest residues on the SOC contents and SOC dynamics, and separately quantify the relative SOC contribution of C4 grass and C3 forest through the sequential fallow periods in the tropical forest of eastern Cameroon. In this study, we combined physical SOM fractionation and 13C natural abundance to quantitatively evaluate the progressive incorporation of newly deposited C (derived from C4 and/or C3 plants) into the different fractions26,27,28.
Results
Soil organic carbon stock in each fraction
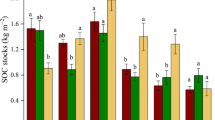
Figure 1 presents the variations of SOC stock in each fraction and soil layer under the different fallow stages, and Table 1 presents the total C contents of each fraction. The SOC was mainly present in the Clay + silt fraction throughout the soil profile, and most of the SOC was observed at 0–40 cm depth. The SOC contents were largest in the surface layer (0–5 cm depth) and decreased with soil depth in all treatment plots. The SOC stock in the M-POM and m-POM in the upper layer (0–10 cm depth) was clearly larger in Cropland (4.6 ± 1.2 (standard error) and 3.7 ± 0.7 Mg C ha−1, respectively) than in Young-F (1.1 ± 0.2 and 0.9 ± 0.1 Mg C ha−1, respectively) and Old-F (1.1 ± 0.2 and 0.6 ± 0.2 Mg C ha−1, respectively). However, these differences were not observed in each deeper layer (<10 cm depth).

Soil organic carbon (SOC) stocks (Mg C ha−1 per 5 cm soil depth) in M-POM (250–2000 μm), m-POM (53–250 μm), and Clay + silt (<53 μm) fractions under cropland and different fallow period sites for the soil profile (100 cm depth) in eastern Cameroon.
Visual inspection showed that most coarse or fine char materials in M-POM and m-POM samples were only in Cropland soils at 0–10 cm depth and Fallow-F soils at 0–5 cm depth. In contrast, most debris of M-POM and m-POM in Young-F and Old-F seemed to be fresh plant debris and not to be black in colour. For the upper layer (0–10 cm), total SOC stock was clearly larger in Fallow-F (33.4 ± 2.5 Mg C ha−1) and Cropland soils (31.2 ± 3.0 Mg C ha−1) than in Old-F (21.0 ± 3.6 Mg C ha−1), and Young-F (24.6 ± 2.7 Mg C ha−1) soils (Table 2). For the soil profile (100 cm depth), total SOC stock decreased in the order Fallow-F (136.6 ± 8.8 Mg C ha−1) > Cropland (122.2 ± 8.3 Mg C ha−1) and Young-F (119.1 ± 10.6 Mg C ha−1) > Old-F (106.4 ± 12.9 Mg C ha−1). This order was the same for SOC stock at 0–40 cm soil depth (Table 2). The total SOC stock in Fallow-F was the largest for most soil layers, except for the 0–10 cm layer in Cropland and the 20–40 cm layer in Young-F. The total SOC stock in Old-F was the smallest for all soil layers throughout the soil profile.
Natural abundance of 13C values in each fraction
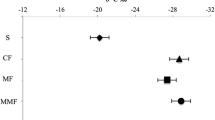
Figure 2 presents the variations of natural abundance of 13C values of SOC in each fraction through the soil profile under different fallow stages. The 13C values in M-POM showed no clear difference between the sites, and these were within −27.4‰ to −28.3‰ in the surface layer (0–5 cm) and increased with soil depth to −24.3‰ to −25.4‰ in the deeper layer (60–100 cm depth). In the case of m-POM, 13C values were clearly larger in Fallow-F (−24.5‰ ± 0.8‰) and Young-F (−24.0‰ ± 0.5‰) than in Cropland (−26.6‰ ± 0.3‰) and Old-F (−26.5‰ ± 0.5‰) only at 5–10 cm depth, while there was no clear difference for the sites in the other soil layers.

Fluctuation of δ13C (‰) values of soil organic carbon (SOC) in M-POM (250–2000 μm), m-POM (53–250 μm), and Clay + silt (<53 μm) fractions under cropland and different fallow period sites through the soil profile in eastern Cameroon. Bars indicate the standard error (N = 4). *Indicates the significant differences for the land management of each soil depth, according to ANOVA (P < 0.05).
The 13C values in Clay + silt were clearly different between the sites at 5–40 cm soil depth, while there was no clear difference in the surface layer (0–5 cm depth) and deeper layers (40–100 cm depth). The 13C values in Clay + silt decreased in the order Young-F > Fallow-F > Cropland and Old-F at 5–40 cm depth (P < 0.05). For example, 13C values in Clay + silt at 5–10, 10–20, and 20–40 cm depth of Young-F were −21.6‰ ± 0.2‰, −20.3‰ ± 0.4‰, and −20.7‰ ± 0.2‰, respectively, whereas they were −25.2‰ ± 0.6‰, −23.5‰ ± 0.2‰, and −22.9‰ ± 0.7‰ for Old-F, respectively. In addition, 13C values in Old-F and Cropland similarly fluctuated thorough the soil profile.
Estimated SOC stock derived from C3 and C4 plants
Figure 3 presents the distribution of SOC derived from C3 and C4 plant in each fraction at 0–40 cm soil depth, and Table 2 shows the calculated SOC stock derived from C3 and C4 plants for each specific soil layer. In M-POM, most SOC was derived from C3 plants (92–98% at 0–5 cm depth and 68.5–77.8% at 20–40 cm depth). SOC derived from C3 plants in M-POM at 0–10 cm depth was clearly larger in Cropland (4.2 ± 0.9 Mg C ha−1) than in Young-F (1.0 ± 0.1 Mg C ha−1) and Old-F (1.0 ± 0.2 Mg C ha−1). In m-POM, the ratio of SOC derived from C3 plants decreased in Fallow-F and Young-F at 10–40 cm depths (e.g. 60.3–66.1%), whereas it was clearly high in Old-F (71.2–85.5%), and intermediate in Cropland (62.8–72.9%). SOC derived from C4 plants in m-POM at 10–40 cm was clearly larger in Young-F (0.6 ± 0.1 Mg C ha−1) than in Old-F (0.2 ± 0.1 Mg C ha−1). In Clay + silt, there was a larger contribution of C4-derived SOC especially in sub-surface layers (5–40 cm), whereas there was no clear effect in the surface layer (0–5 cm depth). The ratio of C4-derived SOC to the total SOC in Clay + silt at 5–40 cm depth was 35.4%, 48.0%, 55.0%, and 33.2% in Cropland, Fallow-F, Young-F, and Old-F, respectively. Within the upper layer (0–40 cm depth), SOC stock derived from C4 plants in Clay + silt was larger in Young-F (34.9 ± 3.3 Mg C ha−1) and Fallow-F (34.3 ± 1.9 Mg C ha−1) than in Old-F (18.1 ± 2.3 Mg C ha−1), while SOC stock derived from C3 plants in Clay + silt was larger in Fallow-F (45.8 ± 3.2 Mg C ha−1) and Cropland (42.4 ± 3.1 Mg C ha−1) than in Young-F (34.7 ± 4.2 Mg C ha−1).

Distribution of soil organic carbon (SOC) derived from tropical forests (C3: filled bar) and grassland (C4: slash bar) in M-POM (250–2000 μm), m-POM (53–250 μm), and Clay + silt (<53 μm) fractions under cropland and different fallow period sites at each soil depth in eastern Cameroon.
Within the soil profile (100 cm depth: Table 2), SOC stock derived from C3 plants was clearly larger in Cropland (82.0 ± 5.2 Mg C ha−1) and Fallow-F (79.4 ± 5.5 Mg C ha−1) than in Young-F (58.7 ± 7.2 Mg C ha−1). In contrast, SOC stock derived from C4 plants was clearly larger in Young-F (60.4 ± 5.8 Mg C ha−1) and Fallow-F (57.2 ± 3.8 Mg C ha−1) than in Cropland (42.2 ± 3.5 Mg C ha−1) and Old-F (35.0 ± 4.1 Mg C ha−1).
Discussion
Total SOC stock at both 100 cm depth and 0–40 cm depth decreased in the order Fallow-F > Cropland and Young-F > Old-F in the present study. This indicates that SOC stock peaked in the Fallow-F (i.e., the first stage of fallow succession), and then started to decrease with forest growth and maturity (i.e., middle to final stage of fallow succession) in this area. However, it should be noted that these results were obtained from the comparison of different fallow period sites, rather than from long-term, fixed-point observation. On the basis of the contribution of different C sources in Table 2, both C4- and C3-derived SOC was largest in the Fallow-F soil compared with other sites, indicating the significant contribution of both C4- and C3-derived SOC to the high SOC in Fallow-F. Because C4-derived SOC clearly increased from Cropland to Fallow-F, C4 grass invasion as a forest understory in Fallow-F is likely to have contributed to the substantial increase of SOC at this early fallow stage. In general, under the humid tropical conditions, the amount of OM incorporated into the soil annually is larger under the grass vegetation rather than under forest vegetation14. This is because forest fixes the C into the aboveground biomass especially in the growing and intermediate stages, such as Fallow-F and Young-F, whereas most C fixed by annual grass, including its root biomass, is incorporated into the soil every year3,29. Tonucci et al.28 also found a significant contribution of C4 pasture grass to SOC stock in agroforestry systems in Brazilian Cerrado. In addition, high N availability of Fallow-F might also stimulate the growth of C4 underforestry, resulting in larger C4 derived SOC30. Therefore, in the present study, invasion of C4 grass as forest understories is likely to have contributed to the increment of SOC stock, especially in the early fallow succession stage, with the largest SOC stock observed in Fallow-F.
In addition, added OM by slash-burn management (mostly C3-derived SOC) is likely to also contribute to the largest SOC stock in Fallow-F. This is because, considering the similar 13C values between the Cropland and Old-F in whole fractions through the soil profile (Fig. 2), the C source of SOC was not critically changed from Old-F to Cropland, i.e., the main C source is C3 plants. This indicates that the difference of SOC stock between the Cropland and Old-F must be mainly caused by applied OM after slash-burn management, such as burned forest biomass. This is consistent with the finding of larger C3-derived SOC in M-POM (mostly char material) in Cropland than that of Old-F. In addition, we also found that C3-derived SOC in each fraction of Cropland for the 0–40 cm depth was roughly similar to that of Fallow-F, whereas C3-derived SOC in M-POM and Clay + silt fractions for this depth clearly decreased from Fallow-F to Young-F (Fig. 3 and Table 2). This indicates that added OM by slash-burn management, including char, remained until the Fallow-F stage, though it began to be pulverized and/or lost in the Young-F stage. Zimmermann et al.31 estimated the average turnover time of purified wood C to be 67 years in a typical tropical savanna in Australia. Because of the more humid conditions in the present study, the coarse char material should be pulverized during a much shorter period, such as 67 years. Dong et al.32 also observed that biochar C loss was ca. 40% after 5 years in dry temperate cropland in China. Because we also found the decrease of C3-derived SOC in the M-POM and m-POM in the upper layer (0–10 cm depth) of Young-F and Old-F compared with those of Cropland, coarse char materials are likely to reduce in size and become part of the Clay + silt fraction during this period (i.e., from Cropland to Young-F)33. Lopez-Martin et al.34 observed the rapid decrease of coarse pyrogenic OM 7 years after forest fire event in Southern Spain, and indicated that the lack of mineral-pyrogenic OM association may have allowed rapid degradation of coarse pyrogenic OM or loss due to transport. On the basis of these results, in the present study, applied burned biomass (including char) should mostly remain in soil ca. 10–20 years after slash-burn management (such as the Cropland and Fallow-F stages in this study) under humid tropical climate conditions, possibly because of its purified structure (char) by fire35,36,37. Thus, in this study, added OM from slash-burn management was considered to still remain in Fallow-F, and it is likely to also contribute to the largest SOC stock in Fallow-F.
From Fallow-F to Young-F succession, we found that the C3-derived SOC clearly decreased, whereas C4-derived SOC remained stable (Table 2). This suggests the mechanism of decomposition of C3-derived SOC as explained above, and the existence of substantial C4 grass as a forest understory even in Young-F. In addition, we found that C4-derived SOC abruptly decreased from Young-F to Old-F succession, especially in Clay + silt (from 34.9 to 18.1 Mg C ha−1 at 0–40 cm depth, P < 0.05), whereas C3-derived SOC increased in Clay + silt (from 34.7 to 38.6 Mg C ha−1 at 0–40 cm depth). This indicates the disappearance of C4 grass understory in Old-F, possibly due to the crown closure and little insolation38,39.
Based on our field observations, there were few forest understories containing mainly C3 plants in Old-F, compared with that in Fallow-F and Young-F, for which the forest understory contained a higher amount of C4 grass. Thus, forest understories must strongly control the SOC stock in the middle to final stage of fallow succession in African tropical forest, and SOC stock was largest in the early fallow stage such as the 4–7-year fallow period, because of the added OM both of char by slash-burn management (C3-derived SOC) and of grass litter in forest understories (C4-derived SOC). Conversely, SOC stock was smallest in the final fallow stage (little C4 grass understories) such as after 50 years of fallow, i.e., in mature forest40,41. However, considering the amount of aboveground biomass C in each site, total C in Old-F (106.4 Mg C ha−1 in soil and 276.7 Mg C ha−1 in aboveground biomass) should be substantially larger than that in fallow-F (136.6 Mg C ha−1 in soil and 104.4 Mg C ha−1 in aboveground biomass). Therefore, to achieve the C sequestration to mitigate the climate change at ecosystem level (SOC + aboveground-C + belowground-C + deadwood-C + litter-C), a longer fallow period is likely to contribute to C sequestration in the tropical forest ecosystem42,43,44. Silatsa et al.13 also observed the significant contribution of aboveground biomass (cacao tree) to the C storage of agroforestry ecosystems in southern Cameroon.
There was a small amount of SOC present as M-POM and m-POM in Old-F and Young-F even in the surface layer, whereas SOC as M-POM and m-POM in Cropland was clearly larger and was mostly composed of C3 charred biomass rather than fresh plant debris (based on our visual observation). These results may indicate the rapid decomposition of normal plant debris and therefore applied OM, whichever from C4 or C3 plants, should be rapidly mineralised and moved into the Clay + silt fraction45,46. Yonekura et al.27 suggested the rapid mineralisation of applied litter under the humid tropical conditions, resulting in difficulties of C sequestration. In addition, because coarse char material in the M-POM and m-POM fractions in Cropland and Fallow-F was reduced in size and may move into the Clay + silt fraction in Young-F and Old-F, the Clay + silt-size char material is likely to have an influence on SOC stock and its stability for decomposition in Young-F and Old-F by association with mineral and/or OM. Thus, further study is necessary to assess the relationship between the particle char material and SOC stability in the Clay + silt fraction24,47.
Conclusion
We evaluated the dynamics and the contributions of C4 grass and C3 forest to SOC stock during the fallow succession in eastern Cameroon by measuring SOC at sites with different fallow periods. It was found that the SOC stock was largest in Fallow-F. The finding that C4-derived C was the main contributor to restoration of the SOC in Fallow-F and Young-F sites (i.e., a 4–30 year fallow period) indicated that forest understories, particularly C4 grass, were important to restore the SOC stock during the early fallow period in African tropical forest of eastern Cameroon. In addition, OM added by slash-burn management still remained and contributed to larger SOC stock in Fallow-F. Based on these results, we put forward the hypothesis that a shorter fallow period, such as Fallow-F, might be sufficient to restore the soil fertility (or SOC restoration) for sustainable slash-burn agriculture in this area. Moreover, we also found that a longer fallow period is likely to contribute to C sequestration and climate change mitigation, because of the enhanced aboveground biomass C of mature forest. Further study is necessary to assess the above hypothesis, reveal the optimal fallow period for sustainable slash-burn agriculture in this area, and determine whether a shorter fallow period (such as Fallow-F in this study) in the tropical forest of eastern Cameroon can supply sufficient nutrients (such as N, P, and other minerals) to subsequent crop cultivation, including determination of an optimal cultivation period.
Methods
Site description and soil sampling
The study area was located in the Diang district of Bertour region (4°34ʹ56″N, 13°16ʹ00″E) in the eastern part of Cameroon, which contains the dominant geographic feature of the Cameroon Plateau. Oxisols are generally distributed throughout this region48,49, which stands at an elevation of approximately 650–700 m. The climate of the region is tropical monsoon with a mean annual temperature of 23.5 °C, and annual rainfall of 1350–1550 mm based on our former field survey50. This area usually experiences two rainy seasons, a short season from September to November and a long season from March to July.
In 2011, we collected soil samples from cropland and different fallow stage sites as follows: (1) cropland (Cropland), (2) short fallow forest (4–7 years; Fallow-F), (3) young forest (20–30 years; Young-F), and (4) old and matured forest (>50 years; Old-F), where the young and old forest developed after slash-burn cultivation. The detailed soil characteristics of Young-F and Old-F have previously been explained in Sugihara et al.51. In the Cropland site, local farmers generally cultivate cassava, banana, and groundnuts for 3–5 years, after opening and burning the mature forest such as Old-F. According to the interview with the local farmers, we selected the Cropland sites that had been in cultivation for 2–4 years, and sites that were mature forest (Old-F). In this region, to create the cultivation area, local farmer slashes only small- or middle-sized trees, but not the large trees (because of its difficulty by hand), and burns the material, and thus big trees still remain in the Cropland site even after slash-burn cultivation12. The Fallow-F, Young-F, and Old-F sites were divided by the period of fallow, which were representative of each natural fallow succession stages in this region, based on an interview with the local farmers. Our vegetation surveys (30 m × 30 m for each site, four replications) revealed no clear differences in the forest tree species composition between Young-F and Old-F, and the dominant vegetation was Albizia zygia, which predominates in the tree layer and Myrianthus arboreus. Estimated aboveground biomass in Young-F and Old-F were 221.1 Mg ha−1 (standard error, 84.0) and 276.7 Mg ha−1 (standard error, 44.7), respectively. In case of Fallow-F, the forest tree species composition was different from that of Young-F and Old-F, mainly comprising Musanga cecropioides, Ficus exasperata, and Macaranga spinosa, which are the typical pioneer species in early forest succession in this region. The estimated aboveground biomass in Fallow-F was 104.4 Mg ha−1 (standard error, 27.5). In addition, in the Fallow-F and Young-F sites, there were also C4 grass forest understories, such as P. setaceum and I. cylindrica, though there were few C4 grass forest understories in Old-F, possibly due to the crown closure and low-light conditions.
We made four pits (four replications) for each site (i.e., a total of 16 pits), and collected soil samples from each soil layer (0–5, 5–10, 10–20, 20–40, 40–60, and 60–100 cm). All sampling sites, except for one pit in Old-F, were within 10 km of each other (Supplemental Fig. 1). Soil samples were air dried, sieved to 2.0 mm, and then held at room temperature until used. All visible plant material larger than 2.0 mm sieve size was removed manually. We also collected two 100 cc soil cores at each soil depth for determination of the soil bulk density, to calculate the soil C stock in the soil profiles. Each of the experimental sites was relatively flat and had a flat area covering more than 1 km2. None of the pits contained gravel, and the clayey soils were classified as Typic Kandiudoxes48. Table S1 provides details of the physicochemical properties of soils under each site. The soil pH (H2O) was measured with a glass electrode (pH/ion meter 225, Iwaki Glass, Japan) at a soil:solution ratio of 1:5 (w:v) after shaking for 1 h. Particle size distribution was evaluated by a pipette method after removing OM with 5% H2O2 at 80 °C. To determine the cation-exchange capacity (CEC), we washed the residual soil with ethanol after ammonium acetate extraction and then extracted the remaining NH4+ with 10% NaCl, and determined the NH4+ concentration. Most physicochemical properties were not significantly different between the sites; however, clay contents of surface layers (0–5 and 5–10 cm depth) were smaller in Cropland than in Old-F and Young-F.
Soil fractionation
The soil samples were manually fractionated by particle size, according to a procedure from Cambardella & Elliot52, as described by Sugihara et al.51. Briefly, 50 g of each soil sample was placed into a 250-mL plastic container and mixed with 150 mL of 0.05 mol L−1 NaCl solution. The soil suspension was then shaken for 16 h in a reciprocal shaker at 150 rpm, after which it was passed through a 0.25-mm sieve and rinsed with distilled water until a clear solution was obtained. Next, the sieved material was continuously passed through a 0.053-mm sieve and rinsed with distilled water until a clear solution was obtained. The material retained on each sieve was dried at 60 °C for 24 h as macro-POM (M-POM; 0.25–2.0 mm) and micro-POM (m-POM; 0.053–0.25 mm), containing the soil’s macro- and micro-particles, respectively; both also contained some plant debris. The soil slurry that passed through the 0.053-mm sieve was dried at 60 °C for 48 h to represent the clay + silt fraction (Clay + silt; <0.053 mm), which contained the mineral-associated OM. Generally, the M-POM fraction is defined as easily mineralisable OM, and the Clay + silt fraction includes residual or stable OM53. In the present study, M-POM mainly contained plant debris and its char, whereas m-POM mainly contained small particles of ash. The recovery ratio for this procedure was 93–99% of total mass and 93–101% of total carbon (TC).
Soil analysis
All the fractionated samples were completely ground to powder using a tungsten mortar and pestle. The TC content for whole soil and for fractionated soil samples were determined with a CN analyser (Vario Max CN, Elementar, Hanau, Germany).
For stable C isotope determinations, oven-dried fractionated samples were analysed for δ13C values using a stable isotope ratio mass spectrometer (IsoPrime100, Elementar UK Ltd, Cheadle, UK) operating in continuous flow mode, coupled with a CN analyser (Vario Max CN, Elementar, Hanau, Germany). Carbon isotope ratios are presented in δ-notation:
where Rsample is the 13C/12C ratio of the sample, and Rstd is the 13C/12C ratio of the international standard PDB54. SOC content derived from the C4 grass plants (Pennisetum spp., and Imperata spp.) or from the C3 forest plants (Albizia spp., and Myrianthus spp.) was estimated based on the equations18,20,55:
where δ is the δ13C of a given samples, δT is a composite sample of the C3 forest plants (−28.5‰) and δG is a composite sample of C4 grass (−14.6‰). Both δT and δG were average (N = 5) of plant litter for C4 and C3 vegetation, and both values were consistent with the previous study in eastern Cameroon22 and therefore, in this study, we assumed that the added OM of C3 and C4 plants had the above δ13C value, respectively.
Statistical analysis
All statistical analyses were performed with SYSTAT 13.5 (SYSTAT Software, Richmond, CA, USA). All data are expressed on a dry-weight basis. To compare the δ13C values obtained in each fraction, we conducted a one-way analysis of variance (ANOVA) to assess the significance of differences between the experimental sites. When ANOVA indicated significant differences, mean comparisons were performed with the Tukey multiple comparison test. We also conducted the same multiple analysis to the total SOC, and calculated C derived from C3 and C4 between the experimental sites. In all cases, P < 0.05 was considered significant.
References
Keesstra, D. S. et al. The significance of soils and soil science towards realization of the United Nations Sustainable Development Goals. SOIL 2, 111–128, https://doi.org/10.5194/soil-2-111-2016 (2016).
Palm, C. A. et al. Management of organic matter in the tropics: translating theory into practice. Nutrient Cycling in Agroecosystems 61, 63–75, https://doi.org/10.1023/A:1013318210809 (2001).
Don, A., Schumacher, J. & Freibauer, A. Impact of tropical land-use change on soil organic carbon stocks-a meta-analysis. Global Change Biology 17, 1658–1670, https://doi.org/10.1111/j.1365-2486.2010.02336.x (2011).
Minasny, B. et al. Soil carbon 4 per mille. Geoderma 292, 59–86, https://doi.org/10.1016/j.geoderma.2017.01.002 (2017).
Nhantumbo, A. B. J. C., Ledin, S. & Du Preez, C. C. Organic matter recovery in sandy soils under bush fallow in southern Mozambique. Nutrient Cycling in Agroecosystems 83, 153–161, https://doi.org/10.1007/s10705-008-9208-9 (2009).
Walker, S. M. & Desanker, P. V. The impact of land use on soil carbon in Miombo Woodlands of Malawi. Forest Ecology and Management 203, 345–360, https://doi.org/10.1016/j.foreco.2004.08.004 (2004).
Mapanda, F. et al. Short-term responses of selected soil properties to clearing and cropping of miombo woodlands in central Zimbabwe. Soil and Tillage Research 129, 75–82, https://doi.org/10.1016/j.still.2013.01.008 (2013).
Heinimann, A. et al. A global view of shifting cultivation: Recent, current, and future extent. PLoS One 12(9), e0184479, https://doi.org/10.1371/journal.pone.0184479 (2017).
Zhang, Q., Justice, C. O. & Desanker, P. V. Impact of simulated shifting cultivation on deforestation and the carbon stocks of the forests of central Africa. Agriculture, Ecosystems and Environment 90, 203–209, https://doi.org/10.1016/S0167-8809(01)00332-2 (2002).
Vagen, T. G., Lal, R. & Singh, B. R. Soil carbon sequestration in sub-Saharan. Africa: a review. Land Degradation and Development 61(1), 53–71, https://doi.org/10.1002/ldr.644 (2005).
Adiku, S. G. K. et al. Short-term effects of crop rotation, residue management, and soil water on carbon mineralization in a tropical cropping system. Plant and Soil 311, 29–38, https://doi.org/10.1007/s11104-008-9652-y (2008).
Kotto-Same, J., Woomer, P. L., Appolinaire, M. & Louis, Z. Carbon dynamics in slash-and-burn agriculture and land use alternatives of the humid forest zone in Cameroon. Agriculture, Ecosystems and Environment 65, 245–256, https://doi.org/10.1016/S0167-8809(97)00060-1 (1997).
Silatsa, F. B. T. et al. Modeling carbon stock dynamics under fallow and cocoa agroforest systems in the shifting agricultural landscape of Central Cameroon. Agroforest Systems 91, 993–1006, https://doi.org/10.1007/s10457-016-9973-4 (2016).
Mureva, A. et al. Soil organic carbon increases in Semi-arid regions while it decreases in humid regions due to Woody-Plant encroachment of grasslands in South Africa. Scientific Reports 8, 15506, https://doi.org/10.1038/s41598-018-33701-7 (2018).
Spiotta, E. M., Silver, W. L., Swanston, C. W. & Ostertag, R. Soil organic matter dynamics during 80 years of reforestation of tropical pasture. Global Change Biology 15, 1584–1597, https://doi.org/10.1111/j.1365-2486.2008.01805.x (2009).
Zhang, K., Dang, H., Zhang, Q. & Cheng, X. Soil carbon dynamics following land-use change varied with temperature and precipitation gradients: evidence from stable isotopes. Global Change Biology 21, 2762–2882, https://doi.org/10.1111/gcb.12886 (2015).
Chiti, T. et al. Impact of woody encroachment on soil organic carbon storage in the Lope National Park, Gabon. Biotropica 49(1), 9–12, https://doi.org/10.1111/btp.12369 (2017).
Yonekura, Y. et al. Changes in soil carbon stock after deforestation and subsequent establishment of “Imperata” grassland in the Asian humid tropics. Plant and Soil 329, 495–507, https://doi.org/10.1007/s11104-009-0175-y (2010).
Yonekura, Y. et al. Dynamics of soil carbon following destruction of tropical rainforest and the subsequent establishment of Imperata grassland in Indonesian Borneo using stable carbon isotopes. Global Change Biology 18, 2606–2616, https://doi.org/10.1111/j.1365-2486.2012.02722.x (2012).
Navarrete, D., Sitch, S., Aragao, L. E. & Pedroni, L. Conversion from forests to pasture in the Colombian Amazon leads to contrasting soil carbon dynamics depending on land management practices. Global Change Biology 22, 3503–3517, https://doi.org/10.1111/gcb.13266 (2016).
Mertens, B. & Lambin, E. F. Land-cover-change trajectories in southern Cameroon. Annals of the Association of American Geographers 90, 467–494, https://doi.org/10.1111/0004-5608.00205 (2000).
Guillet, B. et al. Agreement between floristic and soil organic carbon isotope (13C/12C, 14C) indicators of forest invasion of savannas during the last century in Cameroon. Journal of Tropical Ecology 17(6), 809–832, https://doi.org/10.1111/0004-5608.00205 (2001).
Sankaran, M. et al. Determinants of woody cover in African savannas. Nature 438, 846–849, https://doi.org/10.1111/0004-5608.00205 (2005).
Peltzer, D. A. et al. Effects of biological invasion on forest carbon sequestration. Global Change Biology 16, 732–746, https://doi.org/10.1111/0004-5608.00205 (2010).
Sanchez, C. A. & Lindsell, J. A. The role of remnant trees in carbon sequestration, vegetation structure and tree diversity of early succession regrowing fallows in eastern Siera Leone. African Journal of Ecology 55(2), 188–197, https://doi.org/10.1111/aje.12340 (2017).
Cadisch, G. et al. Carbon turnover (δ13C) and nitrogen mineralization potential of particulate light soil organic matter after rainforest clearing. Soil Biology and Biochemistry 28, 1555–1567, https://doi.org/10.1016/S0038-0717(96)00264-7 (1996).
Yonekura, Y. et al. Soil organic matter dynamics in density and particle-size fractions following destruction of tropical rainforest and the subsequent establishment of Imperata grassland in Indonesian Borneo using stable carbon isotopes. Plant and Soil 372, 683–699, https://doi.org/10.1007/s11104-013-1763-4 (2013).
Tonucci, R. G., Nair, V. D., Nair, R. & Garcia, R. Grass vs. tree origin of soil organic carbon under different land-use systems in the Brazilian Cerrado. Plant and Soil 419, 281–292, https://doi.org/10.1007/s11104-017-3347-1 (2017).
Chen, C. R., Condron, L. M. & Xu, Z. H. Impacts of grassland afforestation with coniferous trees on soil phosphorus dynamics and associated microbial processes: A review. Forest Ecology and Management 255, 396–409, https://doi.org/10.1016/j.foreco.2007.10.040 (2008).
Craig, M. E., Pearson, S. M. & Fraterrigo, J. M. Grass invasion effects on forest soil carbon depend on landscape-level land use patterns. Ecology 96, 2265–2279, https://doi.org/10.1890/14-1770.1 (2015).
Zimmermann, M. et al. Rapid degradation of pyrogenic carbon. Global Change Biology 18, 3306–3316, https://doi.org/10.1111/j.1365-2486.2012.02796.x (2012).
Dong, X., Li, G., Lin, Q. & Zhao, X. Quantity and quality changes of biochar aged for 5 years in soil under field conditions. Catena 159, 136–143, https://doi.org/10.1016/j.catena.2017.08.008 (2017).
Singh, N. et al. Transformation and stabilization of pyrogenic organic matter in a temperate forest field experiment. Global Change Biology 20, 1629–1642, https://doi.org/10.1111/gcb.12459 (2014).
Lopez-Martin, M., Gonzalez-Vila, F. J. & Knicker, H. Distribution of black carbon and black nitrogen in physical soil fractionations from soils seven years after an intense forest fire and their role as C sink. Science and the Total Environment 637, 1187–1196, https://doi.org/10.1016/j.scitotenv.2018.05.084 (2018).
Glaser, B., Lehmann, J. & Zech, W. Ameliorating physical and chemical properties of highly weathered soils in the tropics with charcoal – a review. Biology and Fertility of Soils 35, 219–230, https://doi.org/10.1007/s00374-002-0466-4 (2002).
Santin, C., Doerr, S. H., Preston, C. M. & Gonzalez-Rodriguez, G. Pyrogenic organic matter production from wildfires: a missing sink in the global carbon cycle. Global Change Biology 21, 1621–1633, https://doi.org/10.1111/gcb.12800 (2015).
Luo, Y., Yu, Z., Zhang, K., Xu, J. & Brookes, P. C. The properties and functions of biochars in forest ecosystems. Journal of Soils and Sediments 16, 2005–2020, https://doi.org/10.1007/s11368-016-1483-5 (2016).
Montgomery, R. A. & Chanzdon, R. L. Forest structure, canopy architecture, and light transmittance in tropical wet forests. Ecology 82(10), 2707–2718 (2001). 10.1890/0012-9658(2001)082[2707:FSCAAL]2.0.CO;2.
Dupuy, J. M. & Chazdon, R. L. Interacting effects of canopy gap, understory vegetation and leaf litter on tree seedling recruitment and composition in tropical secondary forests. Forest Ecology and Management 255, 3716–3725, https://doi.org/10.1016/j.foreco.2008.03.021 (2008).
Cookm, R. L., Stape, J. L. & Binkley, D. B. Soil carbon dynamics following reforestation of tropical pasture. Soil Science Society of American. Journal 78, 290–296, https://doi.org/10.2136/sssaj2012.0439 (2014).
Fujisaki, K. et al. From forest to cropland and pasture systems: a critical review of soil organic carbon stock changes in Amazonia. Global Change Biology 21, 2773–2786, https://doi.org/10.1111/gcb.12906 (2015).
Liao, C. et al. Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis. New Phytologist 177, 706–714, https://doi.org/10.1111/j.1469-8137.2007.02290.x (2008).
Fonseca, W., Benayas, J. M. R. & Alice, F. E. Carbon accumulation in the biomass and soil of different aged secondary forests in the humid tropics of Costa Rica. Forest Ecology and Management 262, 1400–1408, https://doi.org/10.1016/j.foreco.2011.06.036 (2011).
Mukul, S. M., Herbohn, J. H. & Firn, J. Tropical secondary forests regenerating after shifting cultivation in the Philippines uplands are important carbon sinks. Scientific Reports 6, 22483, https://doi.org/10.1038/srep22483 (2016).
da Silva, M. L. et al. Carbon content in Amazonian Oxisols after forest conversion to pasture. Revista Brasileira De Ciencia Do Solo 33, 1603–1311, https://doi.org/10.1590/S0100-06832009000600009 (2009).
Fernandez-Ugalde, O. et al. Does phyllosilicate mineralogy explain organic matter stabilization in different particle-size fractions in a 19-year C-3/C-4 chronosequence in a temperate Cambisol? Geoderma 264, 171–178, https://doi.org/10.1016/j.geoderma.2015.10.017 (2016).
El-Naggar, A. et al. Biochar influences soil carbon pools and facilitates interactions with soil: A field investigation. Land Degradation and Development 29, 2162–2171, https://doi.org/10.1002/ldr.2896 (2018).
Soil Survey Staff. Keys to Soil Taxonomy. 12th Ed. Unites States Department of Agriculture Natural Resourcese Conservation Service, Washington, DC (2014).
Sugihara, S. et al. Effect of vegetation on soil C, N, P and other minerals in Oxisols at the forest-savanna transition zone of central Africa. Soil Science and Plant Nutrition 60, 45–59, https://doi.org/10.1080/00380768.2013.866523 (2014).
Shibata, M. et al. Effect of original vegetation on nutrient loss patterns from Oxisol cropland in forests and adjacent savannas of Cameroon. Agriculture, Ecosystems and Environment 257, 132–143, https://doi.org/10.1016/j.agee.2018.01.031 (2018).
Sugihara, S. et al. Soil phosphorus of stable fraction differentially associate with carbon in the tropical forest and savanna of eastern Cameroon. Soil Science and Plant Nutrition 63, 616–627, https://doi.org/10.1080/00380768.2017.1403841 (2017).
Cambardella, C. A. & Elliot, E. T. Particulate soil organic matter changes across a Grassland cultivation sequence. Soil Science Society of American. Journal 56, 777–783, https://doi.org/10.2136/sssaj1992.03615995005600030017x (1992).
Von Lutzow, M. et al. SOM fractionation methods; relevance to functional pools and to stabilization mechanisms. Soil Biology and Biochemistry 39, 2183–2207, https://doi.org/10.1016/j.soilbio.2007.03.007 (2007).
Bernoux, M., Cerri, C. C., Neill, C. & de Moraes, J. F. L. The use of stable carbon isotopes for estimating soil organic matter turnover rates. Geoderma 82, 43–58, https://doi.org/10.1016/S0016-7061(97)00096-7 (1998).
Balesdent, J. & Mariotti, A. Measurement of soil organic matter turnover using 13C natural abundance. In: Mass Spectrometry of Soils (eds Boutton, T. W. & Yamasaki, S.), pp. 83–111. Marcel-Dekker, New York, NY (1996).
Acknowledgements
We thank Alex Boon, PhD, from Edanz Group (www.edanzediting.com/ac) for editing a draft of this manuscript. The JST-JICA-SATREPS, Japanese Society for the Promotion of Science KAKENHI Grant Numbers #24255019, #24228007, #17H06171 and #18H02315 financially supported this work.
Author information
Authors and Affiliations
Contributions
S.S., H.T., T.K. and S.F. planned and designed the research. S.S., M.S., A.D.M. and S.F., performed the soil survey and sampling. S.S. and M.S. performed chemical analysis including the 13C analysis. S.S. analysed the data, wrote the main manuscript, and prepared figures and tables. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sugihara, S., Shibata, M., Mvondo Ze, A.D. et al. Forest understories controlled the soil organic carbon stock during the fallow period in African tropical forest: a 13C analysis. Sci Rep 9, 9835 (2019). https://doi.org/10.1038/s41598-019-46406-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-46406-2
- Springer Nature Limited
This article is cited by
-
Above- and belowground composition and diversity of subtropical plantations and their relationships with soil nutrient stocks
Plant and Soil (2024)
-
Dynamics of land use land cover and its impact on carbon stocks in Sub-Saharan Africa: an overview
Environment, Development and Sustainability (2022)
-
Fire-fallow agriculture as a sustainable cropping system for maintaining organic carbon in Maré Loyalty Island (New Caledonia, southwest Pacific)
Regional Environmental Change (2021)
-
Effects of land use change on turnover and storage of soil organic matter in a tropical forest
Plant and Soil (2020)
-
Mineralogical composition and C/N contents in soil and water among betel vineyards of coastal Odisha, India
SN Applied Sciences (2020)