
Abstract
In this study, we isolated a total of 238 culturable putative bacterial endophytes from four Pinus species (Pinus densiflora, P. koraiensis, P. rigida, and P. thunbergii) across 18 sampling sites in Korea. The samples were cultured in de Man Rogosa Sharpe and humic acid-vitamin agar media. These selective media were used to isolate lactic acid bacteria and Actinobacteria, respectively. Analysis using 16S ribosomal DNA sequencing grouped the isolated putative bacterial endophytes into 107 operational taxonomic units (OTUs) belonging to 48 genera. Gamma-proteobacteria were the most abundant bacteria in each sampling site and three tissues (needle, stem and root). The highest OTU richness and diversity indices were observed in the roots, followed by stem and needle tissues. Total metabolites extracted from three isolates (two isolates of Escherichia coli and Serratia marcescens) showed significant nematicidal activity against the pine wood nematode (Bursaphelenchus xylophilus). Our findings demonstrated the potential use of bacterial endophytes from pine trees as alternative biocontrol agents against pine wood nematodes.
Similar content being viewed by others


Introduction
Endophytes have received significant research interest and show great potential as biocontrol agents (BCAs)1,2,3,4,5. In recent decades, emerging technologies have increasingly relied on the utilization of endophytes as natural and nontoxic sources of pesticides and fertilizers6. The broad applications of BEs make them commercially, economically, and scientifically important7. Endophytes, including bacteria and fungi, produce bioactive compounds by exploiting the conditions of their specific habitat (i.e. internal living tissues of plants). Intimate and permanent associations between endophytes and host plants can be formed without causing havoc to plant tissues. Accordingly, decades of research has demonstrated that endophytes can not only promote plant growth, but also exert protective effects against abiotic and biotic stresses8.
Some studies have focused on endophytic Actinobacteria because of their ability to produce metabolites with diverse functions9,10,11. Lactic acid bacteria (LAB), phylum Firmicutes, are widely used in various industrial applications as starters for food fermentation, probiotics, and BCA12. However, studies on endophytic LAB are limited, whereas a large number of LAB was isolated from withered leaves or plant tissues damaged by insects13,14. Some researchers have been unable to isolate endophytic LAB from standing live crops15,16,17. LAB are present in low numbers in the natural plant environment18.
Chemical methods for controlling plant parasitic nematodes have been extensively applied because of their non-selectivity. However, despite their effectiveness and availability, chemical nematicides have been reconsidered due to the disadvantages, such as high risk for environmental hazard10,19,20. Avermectin extracted from Streptomyces avermitilis, an Actinobacteria species, exhibits significant nematicidal activity21. The genus Streptomyces exerts nematicidal activity against parasitic nematodes22,23. Currently, avermectin and avermectin derivatives are used as trunk-injection agents to control the pine wood nematode (PWN, Bursaphelenchus xylophilus)24,25. PWN, the causal agent of pine wilt disease (PWD), has been initially reported in Canada and USA, infects trees across Japan, China, eastern Asian, and western Europe25,26. In addition, PWN caused serious damage in Korea24,27 where Pinus desiflora, P. koraiensis, and P. thunbergii are natural hosts of PWN, whereas P. rigida, a native species found in North America, is resistant24,28. Biocontrol against PWN using endophytes is not yet fully developed, but an interrelationship between nematodes and their associated bacteria is evident29. Therefore, BEs exhibit potential use as BCAs against PWN.
We hypothesize that the characterized putative BEs might be used to control for PWN. We investigated the distribution and diversity of putative BEs from four Pinus species grown under multiple ecological conditions across Korea. Endophyte community significantly differs among species, sampling site, and tissues of pine trees. We isolated large numbers of putative BEs that do not belong to Actinobacteria and LAB. We also screened the isolated putative BEs for nematicidal activity against PWN. Screening results showed that Escherichia coli and Serratia marcescens exhibited significant nematicidal activity and can thus potentially be used as BCAs against PWN. Here, we tested the hypothesis that the hosts may possess endophytes that can be a practical and effective choice as pest control agents.
Results
Isolation and identification of putative bacterial endophytes
Putative BEs were isolated from four Pinus species across 18 sampling sites in Korea (Supplementary Table S1). HV medium was used to isolate endophytic Actinobacteria. The analysis identified 116 endophytic isolates belonging to 74 OTUs and 26 genera (Supplementary Table S2). However, only 24 Actinobacteria corresponding to 12 OTUs were isolated. Other bacteria included 92 isolates representing 62 OTUs.
We attempted to isolate endophytic LAB using MRS medium. Culturing identified 131 isolates belonging to 54 OTUs and 33 genera (Supplementary Table S2). Only three isolates of LAB belonging to 2 OTUs were isolated from the root of P. densiflora in Jejudo (Pd 2), namely, Lactococcus lactis subsp. lactis, Leuconostoc mesenteroides, and Leuconostoc sp.
Taken together, we identified a total of 238 bacterial isolates belonging to 107 OTUs and 48 genera by culturing samples in MRS and HV media. Most OTUs showed more than 99% similarity with reference strains.
Diversity, distribution, and relative abundance of putative endophytes
Phylogenetic analysis of 238 bacterial isolates revealed the relationship between the different species of BEs (Supplementary Fig. S1). The distribution of isolates showed significant differences between the MRS and HV culture media (Fig. 1). The majority of isolates that were culturable in HV media belonged to the phyla Actinobacteria, Firmicutes, and Proteobacteria (class: Alpha-, Beta-, and Gamma-proteobacteria) (Fig. 1A). On the other hand, the majority of isolates that were culturable in MRS belonged to Firmicutes and Gamma-proteobacteria. All representative isolates were clustered into the following three phyla: Actinobacteria, Firmicutes (including LAB), and Proteobacteria (three classes: Alpha-, Beta-, and Gamma-proteobacteria). Gamma-proteobacteria (73%) constituted the majority of the isolates, whereas Beta-proteobacteria (5%) were the least represented (Fig. 1B). Other, such as Actinobacteria, accounted for 10% of all isolates. Isolates belonging to Alpha-proteobacteria (6%) and Firmicutes (6%) showed similar relative abundance.

Distribution and relative abundances of putative bacterial endophytes isolated from four Pinus species in Korea. (A) Number of endophytes isolated using MRS and HV media. (B) Pie chart showing the relative abundances of endophytes at the phylum level. (C) Pie chart showing the relative abundances of endophytes at the genus level.
The relative abundance of the 48 genera was as follows (Fig. 1C): Enterobacter (15%), Serratia (11%), Raoultella (7%), Lelliottia (6%), Pseudomonas (6%), Microbacterium (4%), and Klebsiella (4%). Forty-one genera comprised less than 3% of the isolates. Gamma-proteobacteria were the most abundant class (phylum) representing 23 genera, whereas Actinobacteria, Firmicutes, Alpha-proteobacteria, and Beta-proteobacteria were represented by 5, 7, 8, and 5 genera, respectively (Fig. 2).

Pie charts represent the relative abundance of each genus of putative bacterial endophytes isolated from four Pinus species in Korea. (A) Actinobacteria. (B) Firmicutes. (C) Alpha-proteobacteria. (D) Beta-proteobacteria. (E) Gamma-proteobacteria.
At the OTU level, Achromobacter sp. was the most abundant (33.3%, 7 isolates) in Beta-proteobacteria, which comprise a total of 10 OTUs with 21 isolates (Fig. 3). Bacillus sp. was the most abundant (23.0%, 6 isolates) in Firmicutes (12 OTUs with 26 isolates). Microbacterium paraoxydans was the most abundant (25.6%, 12 isolates) in Actinobacteria, which consist of 12 OTUs with 47 isolates. Rhizobium sp. (24.0%, 6 isolates) was the most abundant in Alpha-proteobacteria, which consist of 13 OTUs with 25 isolates. Enterobacter sp. (17.1%, 53 isolates) comprised the majority of Gamma-proteobacteria, which comprise 60 OTUs with 311 isolates.

Pie charts represent the relative abundances of isolates in each phylum (class) of putative bacterial endophytes isolated from four Pinus species in Korea. (A) Actinobacteria. (B) Firmicutes. (C) Alpha-proteobacteria. (D) Beta-proteobacteria. (E) Gamma-proteobacteria.
Distribution of putative bacterial endophytes in different tissues of pine trees
Furthermore, we analysed the biological diversity and distribution of BEs in three different tissues (needle, stem, and root) of four pine tree species. The largest proportion of BEs was isolated from root tissues using HV media, whereas the largest proportion of BEs isolated from needle tissues using MRS media (Fig. 4A). Regarding the relative proportion of bacterial phyla/class, Gamma-proteobacteria accounted for the largest proportion of class in each tissue, especially found in the needle (Fig. 4B). Firmicutes (14.1%) and Beta-proteobacteria (1.7%) were the most and least abundant, respectively. The highest percentage of Actinobacteria was found in the root (18.9%). Similarly, the large proportion of Alpha-proteobacteria (8.8%) was observed in the stem. Overall, the composition of root and needle endophytes seems to be significantly different, while there was no difference in other comparisons based on Pearson chi-square test (***P < 0.001). In summary, the largest number of isolates was isolated from root (43%), followed by stem (30%) and needle tissues (27%) (Fig. 4C).

Relative abundances of putative bacterial endophytes in three different tissues (needle, stem, and root) of four Pinus species in Korea. (A) Relative proportion of endophytes isolated using MRS and HV media in three different tissues. (B) Relative proportion of endophytes at the phylum (class) level in three different tissues. (C) Pie chart showing the relative proportion of endophytes in three different tissues. Pearson chi-square test (χ2) was conducted to compare the endophyte composition between three different tissues (***P < 0.001).
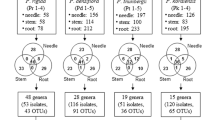
Figure 5 describes the distribution of 238 isolates in three tissues of four different Pinus species. We identified 92, 94, 60, and 94 isolates in P. densiflora, P. koraiensis, P. rigida, and P. thunbergii, respectively. No identical BEs were isolated from all tissues in P. koraiensis and P. rigida. No identical BEs were identified in both the needle and stem tissues of P. densiflora. Overall, root tissues exclusively harboured 29 OTUs (87 isolates; the highest OTU richness), followed by 20 OTUs (51 isolates) in stem tissues, and 13 OTUs (47 isolates) in the needle. Some isolates were simultaneously present in two tissues, while 14 isolates were found in all three tissues. Subsequently, the biological diversity of BEs was analysed for each tissue. The highest OTU richness were found in the root (68 OTUs, followed by stem (59 OTUs), and needle tissues (45 OUTs Fig. 6A). Nevertheless, the bacterial OTUs in the root tissues were unevenly distributed, lowest diversity of OTU (H′ = 1.885) and evenness (E = 0.446) found in root. The highest level of diversity and evenness was found in stem tissues (H′ = 2.278, E = 0.978; Fig. 6B,C).

Venn diagram showing the distribution of putative bacterial endophytes in three different tissues (needle, stem, and root) of four Pinus species in Korea. The distribution of 238 isolates was described in three tissues of four different Pinus species.

Operational taxonomic unit (OTU) diversity of putative bacterial endophytes in three different tissues (needle, stem, and root) of four Pinus species across 18 sampling sites in Korea. (A) Bar graph representing the overall OTU richness in three different tissues and across the different sampling sites. (B) Bar graph representing the overall OTU diversity (Shannon’s index, H′) in three different tissues and across the different sampling sites. (C) Bar graph representing the overall OTU evenness in three different tissues across the different sampling sites. Dots indicate the richness, diversity, and evenness of different tissues in each site.
Distribution of putative endophytes in different sampling sites
Samples isolated from P. densiflora and P. thunbergii were collected from 5 sampling sites, named Pd 1 to 5 and Pt 1 to 5, respectively. Samples from P. koraiensis and P. rigida were collected from 4 sampling sites, designated Pk 1 to 4 and Pr 1 to 4, respectively.
There were significant differences in the distribution of OTUs among the 18 sampling sites, indicating high OTU richness. The highest OTU richness was found in Pk 4 (25 OTUs), and more than 20 OTUs were identified in Pr 3, Pd 2, and Pt 4. Pr 1 showed the lowest OTU richness among the sampling sites (3 OTUs) (Fig. 6A). Likewise, biological diversity was analyzed using Shannon index (H′). The highest H′ value was found in Pt 5 (H′ = 2.945), The lowest index was detected in Pr 1 (H′ = 1.098) (Fig. 6B). Furthermore, the highest OTU evenness (E) values were observed in Pr 1 (E = 1) and Pd 5 (E = 0.943), while the lowest evenness was detected in Pd 4 (E = 0.438; Fig. 6C).
More BEs were isolated from Pr 2, Pr 3, Pd 2, Pd 5, Pt 2, Pt 4, Pt 5, and Pk 4 using HV media than using MRS media (Fig. 7A). The distribution of BEs isolated from different tissues in each sampling site was analyzed (Fig. 7B). In the majority of sampling sites, the largest number of BEs was isolated from root tissue, followed by stem and needle tissues. The largest number of BEs was from needle tissues in Pr 2, Pt 5, and Pk 2, while the largest number of isolates was from stem tissues in Pr 3 and Pt 2. Gamma-proteobacteria accounted for the largest proportion of isolates obtained across all sampling sites, except for Pr 1 (Fig. 7C). The largest proportion of Gamma-proteobacteria (100%) was found in Pd 3, while the largest proportion of Firmicutes (66.7%) was found in Pr 1. The highest proportion of Actinobacteria was found in Pr 2 (28.6%) and Pt 4 (27.5%). The highest proportion of Alpha-proteobacteria and Beta-proteobacteria were observed in Pd 2.

Numbers and relative proportions of putative bacterial endophytes isolated from three different tissues (needle, stem, and root) of four Pinus species across 18 sampling sites in Korea. (A) Bar graph representing the total number of isolates using MRS and HV media across sampling sites. (B) Bar graph representing total number of isolates across sampling sites for each different tissue. (C) Bar graph representing the relative proportion of isolates at the phylum (class) level.
Nematicidal activity of total metabolites against pine wood nematode
Total metabolites extracted from BEs using ethyl acetate (EtOAc) were used to test nematicidal activity against PWN. Eight-day-old adult nematodes were used for the screening. Linearization of PWN was observed after 12 h of treatment. Primary screening identified 17 isolates that showed strong nematicidal activity, corresponding to greater than 50% mortality of nematodes after 12 h of treatment (data not shown). Among 17 isolates, 76% were Proteobacteria, while others belonged to 3 Firmicutes and 1 Actinobacteria. In particular, we identified 3 isolates from 2 different species, Escherichia coli (M131, 67% mortality; M132, 63% mortality) and Serratia marcescens (M44, 60% mortality) (Fig. 8). Three isolates showed significantly higher nematicidal activity compared to control 1 and 2 at a level of P < 0.001.

Nematicidal activity of ethyl acetate extracts isolated from the three selected putative bacterial endophytes. Nematodes (Bursaphelenchus xylophilus) were monitored after 12 h of treatment. Bar graph shows the percentage of dead nematodes at 1,000 ppm treatment. Data are the mean ± standard error of four biological replicates with six technical replicates (n = 24). Control 1 = double distilled (DD) water. Control 2 = 3% DMSO in 100% methanol (solvent control). Control 3 = Abamectin (1,000 ppm). Serratia marcescens (M44), Escherichia coli (M131), and Escherichia coli (M132). Significant difference at ***P < 0.001 vs. control 2. Scale bars = 100 μm.
Discussion
PWD is a common destructive disease in pine tree typically caused by PWN. To prevent progression of the disease, nematicidal compounds such as abamectin and avermectin have been extensively used. These nematicidal compounds were originally isolated from Streptomyces avermitilis30,31, suggesting that environmental bacteria could be a good source of a myriad of bioactive metabolites. Previous studies have demonstrated that Actinobacteria and LAB have nematicidal activity30,31,32. Thus, the goal of the present study was to isolate culturable putatively endophytic Actinobacteria and LAB from different tissues (needle, stem and root) of pine trees using MRS and HV media in search for possible biocontrol agents for PWN.
We identified a total of 24 (12 OTUs) and 3 (3 OTUs) isolates of culturable Actinobacteria and LAB, respectively. The majority of the isolated BEs (211 isolates, 92 OTUs) did not belong to Actinobacteria and LAB. In addition, we identified 219 unique isolates representing 60 OTUs using MRS/HV media. These findings expanded the set of the culturable BEs in pine trees isolated using different culture media.
Actinobacteria were predominantly isolated from various sources such as air, water, and plants33. Here, the majority of the 24 Actinobacteria were found in root tissues (72%) corroborating the results in the previous studies34,35. Species belonging to Actinobacteria are the predominant microbiota found in plant roots attributing to the close associations between root and soil bacteria. Moreover, epidermal breakage caused by differentiation of branch roots might facilitate the entry and colonization of Actinobacteria into root tissues. In cereal crops, more number of Actinobacteria exist in the rhizosphere36,37,38. Actinobacteria are well-known to be in association with the rhizosphere and in colonizing plant internal tissues39,40. Among the 6 identified genera of Actinobacteria in this study, Microbacterium has been shown to be the dominant genus. Interestingly, we isolated Microbacterium paraoxydans, which has been reported to promote plant growth41.
LAB are present inside living plants because of their acid-tolerance and anaerobic properties18. LAB have been isolated from plant surfaces and damaged plant tissues13,16. Similarly, many isolates have been cultivated from stem tissues of lodgepole pine trees (Pinus contorta)42. The functions of LAB in plants and the exact reasons for their abundance in damaged plants are still not clear. Under suitable conditions, it has been shown that lactic fermentation occurs spontaneously in harvested vegetables or fruits43, in which LAB play a significant role. Previous studies have demonstrated that damaged plants can secrete specific chemical signals, including organic acids such as oxaloacetate and malic acid that can attract beneficial bacteria to mediate plant defense responses44,45. A similar mechanism is likely to occur for LAB-mediated plant protection. LAB can produce antagonistic compounds and provide an acidic environment that can protect plants from pathogenic microorganisms43. In this study, three LAB OTUs were isolated from the roots of P. densiflora obtained from Jejudo. Among 18 sampling sites, the Jejudo sampling site showed the lowest soil pH (5.3) condition and high organic contents (50.6 g/kg) which possibly explain the occurrence of these LAB isolates in association with pine trees in this area.
Microbes in the environment are attracted towards fruits and flowers46, by plant substance, and dispersed to other tissues47. The migration of BEs involves whole plant from the root to needle48, thereby posing a possibility of bacterial migration to and from any of the three tissues (needle, stem and root). Our results showed that 14 BE isolates were shared among all three tissues. Root and stem tissues were found to share a high number of BEs (17 isolates), whereas most BEs were exclusively found in a single tissue type. Our findings indicated that all BE phyla were distributed among the stem and root. The highest proportions of Firmicutes and Gamma-proteobacteria were found in the needle. Actinobacteria were more abundant in the root than in the needle and stem, exemplifying its reported high abundance in soil49. In addition, the proportion of Alpha-proteobacteria was higher in the stem than in the root. Most of the BEs in this study were isolated using HV medium (pH 7.2), however a portion of Gamma-proteobacteria (50%) and Firmicutes (3%) were isolated under acidic medium (MRS, pH 6.2) suggesting that these isolated BEs might be tolerant to acidic condition and can uptake carbon and nitrogen from humic acid. In general, the diversity and relative abundance of BEs are influenced by various factors, including the host plants, environment, and cultivation method34,50.
Many endophytes exert preventive effects against plant diseases and can colonize plants for long-term and stable periods. Bacillus pumilus and B. cereus isolated from pine trees in China have been shown to exhibit strong nematicidal activity against PWN51. Furthermore, a novel bacterial strain has been identified that could secrete serine/neutral proteases that degrade the integument of nematode, which cause leakage of nematode contents resulting to the death of the parasitic nematodes52. In search for alternative sources of nematicidal compounds for PWN, we have tested secondary metabolites from the isolated pine tree BEs. We have found two strains of Escherichia coli, and Serratia marcescens that exhibited significant nematicidal activity. In a previous study, S. marcescens isolated from infected Pinus pinaster showed strong nematicidal activity of its total metabolite53.
In summary, endophytic Actinobacteria and LAB have been isolated from pine trees albeit with low abundance compared to other phyla and species, respectively. Putative endophytic bacterial community from pine trees noticeably differs in all three tissues, and most of the bacteria can be distinguished as tissue-specific and differ from one sampling site to another. The isolated Escherichia coli strains and Serratia marcescens could potentially be used as biocontrol agents for PWN. These efforts provide a better understanding of the composition of pine tree bacterial endophytes and also an empirical baseline to explore beneficial BEs for biocontrol of PWN. Further studies investigating the mechanisms of action of BEs’ metabolites as well as the identification and purification of active compound are necessary to develop strategies for controlling PWN.
Methods
Sample preparation
Pine tree samples (needle, stem and root tissues) were collected from four Pinus species (P. densiflora, P. koraiensis, P. rigida and P. thunbergii) across 18 sampling sites in Korea from June to August 2016 (Supplementary Table S1). Pine tree samples were collected from five (P. densiflora and P. thunbergii) to four (P. koraiensis and P. rigida) sampling sites. Tissue samples were collected from six trees (biological replicates) from each sampling site. Each tissue sample was collected from two opposite sides of the tree. Young needle samples (2-years old) were collected using sterilized blades. Stem samples were collected at one meter above the ground using a sterilized increment borer. Tertiary root tissues were collected without uprooting at a depth of 10 to 25 cm below the ground. Samples were individually placed in clean zip bags and stored at 4 °C until further analysis.
All tissue samples (needle, stem and root tissues) were dissected and weighed (1 g) separately. The samples were washed thrice in sterilized reverse osmosis (RO) treated water and sonicated for 20 s to remove loam and organic substance54,55,56. Samples were rinsed in sterile 0.1% Tween-20 for 5 min, 70% ethanol for 5 min, and 4% sodium hypochlorite (NaOCl) solution for 5 min. Next, samples were washed ten times in RO water to remove the chemicals and further soaked in sterile 10% (w/v) sodium bicarbonate (NaHCO3) for 10 min to inhibit the growth of fungal endophytes. The samples were then washed ten times in RO water again. To make sure that the sample surface was sterile, samples were rolled on tryptic soy agar (TSA) and nutrient agar (NA) plates (Thermo Fisher Scientific, Waltham, MA, USA), and the Petri dishes were monitored for up to 15 days at 28 °C. Each sample was ground in 10 ml of sterile phosphate buffer (10 mM, pH 7.2), shaken at 110 rpm for 30 min at 28 °C, and subsequently stored at 4 °C.
Isolation of putatively endophytic Actinobacteria and lactic acid bacteria
Phosphate buffer was serially diluted for five to ten dilutions. Each dilution (500 µL) was added to 5 mL of HV broth (MB Cell, LA, CA, USA)57 with shaking at 150 rpm for up to 2 weeks at 30 °C. The liquid culture (100 µL) was plated on HV agar and incubated at 30 °C for up to 3 weeks. After sub-culturing, pure colonies on the TSA plates were differentiated based on shape, size, colour, texture, form, height and edge. Finally, individual colonies were stored in glycerol stocks.
The procedure for isolation of putatively endophytic LAB was identical to the method employed for isolating endophytic Actinobacteria but using MRS medium (Thermo Fisher Scientific). Each dilution (500 µL) of phosphate buffer was add to 5 mL of MRS broth and shaken at 150 rpm at 30 °C for up to 1 week. After spreading the turbid liquid medium on MRS agar plates, the plates were incubated at 30 °C for up to 1 week for the selection of the different colonies.
Molecular identification
The 16S ribosomal DNA (rDNA) region was amplified from all BEs using universal primers (27 F 5′-AGAGTTTGATCCTGGCTCAG-3′; 1492 R 5′-GGTTACCTTGTTACGACTT-3′) according to the Bionics protocol (Seoul, Korea)58. Amplified sequences were analysed using Geneious version v. 10.1.3 software (Biomatters, Auckland, New Zealand)59,60 and validated via NCBI BLAST search (Supplementary Dataset 1). MEGA v. 7.0 was used to build the phylogenetic tree using the neighbour-joining method and a bootstrap test with 1,000 replications (http://www.megasoftware.net)61.
Extraction of total metabolites from putative bacterial endophytes
Total metabolites were isolated from tryptic soya broth (TSB) cultures using ethyl acetate (EtOAc)62,63. Endophytes were cultured at 30 °C overnight in TSB with shaking at 150 rpm. TSB cultures were added with an equal volume of EtOAc, sonicated for 1 h, and shaken at 120 rpm overnight. The mixture was allowed to stand for 2 h. The top clear phase was transferred to a new flask. EtOAc was evaporated using a rotary evaporator at 50 °C to concentrate the crude extracts. The crude extracts were dissolved in 3% dimethyl sulfoxide (DMSO) in 100% methanol.
Data analysis
Relative abundance was estimated as the percentage of the number of isolates belonging to a particular OTU or phylum divided by the total number of isolates. OTU richness (S) represents the number of OTU recovered from a specific sampling site or tree tissue. OTU evenness (E) was calculated using the following equation: E = H/Hmax, where Hmax is the maximum value of Shannon’s diversity index (H). H was calculated using the following equation: H = −Σ (Pi × In Pi), where Pi is the relative proportion of OTU i in a sampling site or tree tissue64. Graphs were generated and statistical analysis of two-way analysis of variance (ANOVA) (Tukey) was performed to test for significant differences between treatments using GraphPad Prism 7 (La Jolla, CA, USA). Pearson chi-square test (χ2) was performed using R programming (https://www.R-project.org/). OTUs were used to calculate for OTU richness, diversity and the related graphs.
Nematicidal activity test
PWN was obtained from National Institute of Forest Science (Seoul, Korea). PWNs were fed with the fungus Botrytis cinerea, and the plate was incubated at 25 °C in the dark for 8 days65. PWN was harvested using Baermann funnel technique66. PWN was suspended in sterile water, counted, and separated in 96-well polystyrene plates (100 nematodes in 90 µL of water). Crude extracts (0.1 mg in 10 µL, final concentration = 1,000 ppm) were added to the wells of a 96-well polystyrene plate, and the plate was incubated at 25 °C in dark for 12 h. The following treatments were used as controls: control 1, double distilled (DD) water; control 2, 3% DMSO in methanol; positive control, abamectin at 1,000 ppm. Dead nematodes were observed under a microscope and counted. The percentages of dead nematodes were calculated for descriptive analysis. According to the Schneider-Orelli formula, the mortality of nematodes can be corrected by ignoring the mortality in the control67: Corrected mortality (%) = (mortality % in treatment − mortality % in negative control)/(1 − mortality % in negative control).
References
Park, Y.-H. et al. Screening and characterization of endophytic fungi of Panax ginseng Meyer for biocontrol activity against ginseng pathogens. Biol. Control 91, 71–81 (2015).
Park, Y. H., Kim, Y., Mishra, R. C. & Bae, H. Fungal endophytes inhabiting mountain-cultivated ginseng (Panax ginseng Meyer): Diversity and biocontrol activity against ginseng pathogens. Sci. Rep. 7, 1–10 (2017).
Park, Y. H. et al. Endophytic Trichoderma citrinoviride isolated from mountain-cultivated ginseng (Panax ginseng) has great potential as a biocontrol agent against ginseng pathogens. J. Ginseng Res. 3, 1–13 (2019).
Eljounaidi, K., Lee, S. K. & Bae, H. Bacterial endophytes as potential biocontrol agents of vascular wilt diseases – review and future prospects. Biol. Control 103, 62–68 (2016).
Khan Chowdhury, M. D. E. et al. Composition, diversity and bioactivity of culturable bacterial endophytes in mountain-cultivated ginseng in Korea. Sci. Rep. 7, 10098 (2017).
Berg, G., Rybakova, D., Grube, M. & Köberl, M. The plant microbiome explored: implications for experimental botany. J. Exp. Bot. 67, 995–1002 (2016).
Backman, P. A. & Sikora, R. A. Endophytes: An emerging tool for biological control. Biol. Control 46, 1–3 (2008).
Cao, L., Qiu, Z., You, J., Tan, H. & Zhou, S. Isolation and characterization of endophytic streptomycete antagonists of fusarium wilt pathogen from surface-sterilized banana roots. FEMS Microbiol. Lett. 247, 147–152 (2005).
Dinesh, R., Srinivasan, V., TE, S., Anandaraj, M. & Srambikkal, H. Endophytic actinobacteria: diversity, secondary metabolism and mechanisms to unsilence biosynthetic gene clusters. Crit. Rev. Microbiol. 43, 546–566 (2017).
Passari, A. K. et al. Insights into the functionality of endophytic actinobacteria with a focus on their biosynthetic potential and secondary metabolites production. Sci. Rep. 7, 1–17 (2017).
Hug, J. J., Bader, C. D., Remškar, M., Cirnski, K. & Müller, R. Concepts and methods to access novel antibiotics from actinomycetes. Antibiot. 7, 44 (2018).
Mazzoli, R., Bosco, F., Mizrahi, I., Bayer, E. A. & Pessione, E. Towards lactic acid bacteria-based biorefineries. Biotechnol. Adv. 32, 1216–1236 (2014).
Stirling, A. C. & Whittenbury, R. Sources of the lactic acid bacteria occurring in silage. J. Appl. Bacteriol. 26, 86–90 (1963).
Anacarso, I., Bassoli, L., Sabia, C., Iseppi, R. & Condò, C. Isolation and identification of lactic acid bacteria from plants and other vegetable matrices and microbial recombination with Enterococcus spp. Am. Res. Thoughts 1, 1503–1515 (2015).
Kroulik, J. T., Burkey, L. A. & Wiseman, H. G. The microbial populations of the green plant and of the cut forage prior to ensiling. J. Dairy Sci. 38, 256–262 (1955).
Fenton, M. P. An investigation into the sources of lactic acid bacteria in grass silage. J. Appl. Bacteriol. 62, 181–188 (1987).
Duar, R. M. et al. Lifestyles in transition: evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 41, S27–S48 (2017).
Mundt, J. O. & Hammer, J. L. Lactobacilli on plants. Appl. Environ. Microbiol. 16, 1326–30 (1968).
Akhtar, M. & Mahmood, I. Control of plant-parasitic nematodes with organic and inorganic amendments in agricultural soil. Appl. Soil Ecol. 4, 243–247 (1996).
Akhtar, M. & Malik, A. Roles of organic soil amendments and soil organisms in the biological control of plant-parasitic nematodes: A review. Bioresour. Technol. 74, 35–47 (2000).
Burg, R. W. et al. Avermectins, new family of potent anthelmintic agents: producing organism and fermentation. Antimicrob. Agents Chemother. 15, 361–367 (1979).
Samac, D. A. & Kinkel, L. L. Suppression of the root-lesion nematode (Pratylenchus penetrans) in alfalfa (Medicago sativa) by Streptomyces spp. Plant Soil 235, 35–44 (2001).
Ruanpanun, P., Laatsch, H., Tangchitsomkid, N. & Lumyong, S. Nematicidal activity of fervenulin isolated from a nematicidal actinomycete, Streptomyces sp. CMU-MH021, on Meloidogyne incognita. World J. Microbiol. Biotechnol. 27, 1373–1380 (2011).
Shin, S.-C. Pine wilt disease in Korea BT - pine wilt disease. In (eds Zhao, B. G., Futai, K., Sutherland, J. R. & Takeuchi, Y.) 26–32, https://doi.org/10.1007/978-4-431-75655-2_5 (Springer Japan, 2008).
Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 51, 61–83 (2013).
Tóth, Á. Bursaphelenchus xylophilus, the pinewood nematode: Its significance and a historical review. Acta Biol. Szeged. 55, 213–217 (2011).
Kwon, T.-S., Shin, J. H., Lim, J.-H., Kim, Y.-K. & Lee, E. J. Management of pine wilt disease in Korea through preventative silvicultural control. Forest Ecol. Manag. 261, 562–569 (2011).
Park, I.-K., Kim, J., Lee, S.-G. & Shin, S.-C. Nematicidal activity of plant essential oils and components from ajowan (Trachyspermum ammi), allspice (Pimenta dioica) and litsea (Litsea cubeba) essential oils against pine wood nematode (Bursaphelenchus xylophilus). J. Nematol. 39, 275 (2007).
Proença, D. N., Grass, G. & Morais, P. V. Understanding pine wilt disease: roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. Microbiologyopen 6, 1–20 (2017).
Siddique, S., Syed, Q., Adnan, A. & Ashraf Qureshi, F. Isolation, characterization and selection of avermectin-producing Streptomyces avermitilis strains from soil samples. Jundishapur J. Microbiol. 7, 1–7 (2014).
Jayakumar, J. & Ramakrishnan, S. Evaluation of avermectin and its combination with nematicide and bioagents against root knot nematode, Meloidogyne incognita in tomato. J. Biol. Control 23, 317–319 (2009).
Anter, A. A., Amin, A. W., Ashoub, A. H. & El-Nuby, A. S. Evaluation of some rhizobacteria as induce systemic resistance or bio-control agents in controlling root-knot nematode, Meloidogyne incognita on tomato. Egypt. J. Agronematol. 13, 107–123 (2014).
Hruska, K. & Kaevska, M. Mycobacteria in water, soil, plants and air: A review. Vet. Med. 57, 623–679 (2012).
Tan, R. X. & Zou, W. X. Endophytes: a rich source of functional metabolites. Nat. Prod. Rep. 18, 448–459 (2001).
Passari, A. K., Mishra, V. K., Saikia, R., Gupta, V. K. & Singh, B. P. Isolation, abundance and phylogenetic affiliation of endophytic actinomycetes associated with medicinal plants and screening for their in vitro antimicrobial biosynthetic potential. Front. Microbiol. 6, 1–13 (2015).
El-Tarabily, K. A. et al. The potential for the biological control of cavity-spot disease of carrots, caused by Pythium coloratum, by streptomycete and non-streptomycete actinomycetes. New Phytol. 137, 495–507 (1997).
Crawford, D. L., Lynch, J. M., Whipps, J. M. & Ousley, M. A. Isolation and characterization of actinomycete antagonists of a fungal root pathogen. Appl. Environ. Microbiol. 59, 3899–3905 (1993).
Miller, H. J., Henken, G. & Veen, J. V. Variation and composition of bacterial populations in the rhizospheres of maize, wheat, and grass cultivars. Canadian Journal of Microbiology 35, 656–660 (1989).
Bukhalid, R. A. & Loria, R. Cloning and expression of a gene from Streptomyces scabies encoding a putative pathogenicity factor. J. Bacteriol. 179, 7776–7783 (1997).
Healy, F. G., Bukhalid, R. A. & Loria, R. Characterization of an insertion sequence element associated with genetically diverse plant pathogenic Streptomyces spp. J. Bacteriol. 181, 1562–1568 (1999).
Madhaiyan, M. et al. Microbacterium azadirachtae sp. nov., a plant-growth-promoting actinobacterium isolated from the rhizoplane of neem seedlings. Int. J. Syst. Evol. Microbiol. 60, 1687–1692 (2010).
Bal, A., Anand, R., Berge, O. & Chanway, C. P. Isolation and identification of diazotrophic bacteria from internal tissues of Pinus contorta and Thuja plicata. Can. J. For. Res. 42, 807–813 (2012).
Holzer, M., Mayrhuber, E., Danner, H. & Braun, R. The role of Lactobacillus buchneri in forage preservation. Trends Biotechnol. 21, 282–287 (2003).
Schilling, O. et al. Transcriptional and metabolic responses of Bacillus subtilis to the availability of organic acids: transcription regulation is important but not sufficient to account for metabolic adaptation. Appl. Environ. Microbiol. 73, 499–507 (2007).
Rudrappa, T., Czymmek, K. J., Pare, P. W. & Bais, H. P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 148, 1547–1556 (2008).
Compant, S., Mitter, B., Colli-Mull, J. G., Gangl, H. & Sessitsch, A. Endophytes of grapevine flowers, berries, and seeds: identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microb. Ecol. 62, 188–197 (2011).
Compant, S., Clément, C. & Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 42, 669–678 (2010).
Chi, F. et al. Ascending migration of endophytic rhizobia, from roots to leaves, inside rice plants and assessment of benefits to rice growth physiology. Appl. Environ. Microbiol. 71, 7271–7278 (2005).
Berdy, J. Are actinomycetes exhausted as a source of secondary metabolites? Russ. Biotechnol. 7, 3–26 (1995).
Ulrich, K., Ulrich, A. & Ewald, D. Diversity of endophytic bacterial communities in poplar grown under field conditions. FEMS Microbiol. Ecol. 63, 169–180 (2008).
Liangliang, L. I., Tan, J. & Chen, F. The screening and identification of two bacterial strains with nematicidal activity against Bursaphelenchus xylophilus. J. Nanjing For. Univ. 41, 37–41 (2017).
Huang, X. W., Niu, Q. H., Zhou, W. & Zhang, K. Q. Bacillus nematocida sp. nov., a novel bacterial strain with nematotoxic activity isolated from soil in Yunnan, China. Syst. Appl. Microbiol. 28, 323–327 (2005).
Paiva, G. et al. Nematicidal bacteria associated to pinewood nematode produce extracellular proteases. PLoS One 8, 1–11 (2013).
Chaudhry, V. & Patil, P. B. Genomic investigation reveals evolution and lifestyle adaptation of endophytic Staphylococcus epidermidis. Sci. Rep. 6, 19263 (2016).
Kaewkla, O. & Franco, C. M. M. Rational approaches to improving the isolation of endophytic actinobacteria from australian native trees. Microb. Ecol. 65, 384–393 (2013).
Ma, L. et al. Phylogenetic diversity of bacterial endophytes of Panax notoginseng with antagonistic characteristics towards pathogens of root-rot disease complex. Antonie van Leeuwenhoek, Inter. J. G. Mol. Microb. 103, 299–312 (2013).
Taechowisan, T., Peberdy, J. F. & Lumyong, S. Isolation of endophytic actinomycetes from selected plants and their antifungal activity. World J. Microbiol. Biotechnol. 19, 381–385 (2003).
Ji, S. H. et al. Biocontrol activity of Bacillus amyloliquefaciens CNU114001 against fungal plant diseases. Mycobiol. 41, 234–242 (2013).
Kerkhof, L. J. et al. Profiling bacterial communities by MinION sequencing of ribosomal operons. Microbiome 5, 116 (2017).
Snesrud, E. et al. The birth and demise of the ISApl1-mcr-1-ISApl1 composite transposon: the vehicle for transferable colistin resistance. MBio 9, e02381–17 (2018).
Kumar, S., Stecher, G. & Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874 (2016).
Vinale, F. et al. Major secondary metabolites produced by two commercial Trichoderma strains active against different phytopathogens. Lett. Appl. Microbiol. 43, 143–148 (2006).
Paul, N. C., Ji, S. H., Deng, J. X. & Yu, S. H. Assemblages of endophytic bacteria in chili pepper (Capsicum annuum L.) and their antifungal activity against phytopathogens in vitro. Plant Omics 6, 441 (2013).
Ricotta, C. Bridging the gap between ecological diversity indices and measures of biodiversity with Shannon’s entropy: comment to Izsák and Papp. Ecol. Model. 152, 1–3 (2002).
Liu, G., Lai, D., Liu, Q. Z., Zhou, L. & Liu, Z. L. Identification of nematicidal constituents of Notopterygium incisum rhizomes against Bursaphelenchus xylophilus and Meloidogyne incognita. Molecules 21, 1276 (2016).
Czyzewska, M. M. et al. Dietary acetylenic oxylipin falcarinol differentially modulates GABAA receptors. J. Nat. Prod. 77, 2671–2677 (2014).
Guo, Q. et al. A nematicidal tannin from Punica granatum L. rind and its physiological effect on pine wood nematode (Bursaphelenchus xylophilus). Pest. Biochem. Physiol. 135, 64–68 (2017).
Acknowledgements
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03030357).
Author information
Authors and Affiliations
Contributions
Y.L. performed most of the experiments; L.N.P., H.K., J.H., B.S.H., S.K.K. and S.-C.P. analyzed the data; Y.L., L.N.P. and H.B. wrote the article with contribution of all the authors; H.B. supervised the experiments.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, Y., Ponpandian, L.N., Kim, H. et al. Distribution and diversity of bacterial endophytes from four Pinus species and their efficacy as biocontrol agents for devastating pine wood nematodes. Sci Rep 9, 12461 (2019). https://doi.org/10.1038/s41598-019-48739-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-48739-4
- Springer Nature Limited