
Abstract
Fin development and regeneration are complex biological processes that are highly relevant in teleost fish. They share genetic factors, signaling pathways and cellular properties to coordinate formation of regularly shaped extremities. Especially correct tissue structure defined by extracellular matrix (ECM) formation is essential. Gene expression and protein localization studies demonstrated expression of fndc3a (fibronectin domain containing protein 3a) in both developing and regenerating caudal fins of zebrafish (Danio rerio). We established a hypomorphic fndc3a mutant line (fndc3awue1/wue1) via CRISPR/Cas9, exhibiting phenotypic malformations and changed gene expression patterns during early stages of median fin fold development. These developmental effects are mostly temporary, but result in a fraction of adults with permanent tail fin deformations. In addition, caudal fin regeneration in adult fndc3awue1/wue1 mutants is hampered by interference with actinotrichia formation and epidermal cell organization. Investigation of the ECM implies that loss of epidermal tissue structure is a common cause for both of the observed defects. Our results thereby provide a molecular link between these developmental processes and foreshadow Fndc3a as a novel temporal regulator of epidermal cell properties during extremity development and regeneration in zebrafish.
Similar content being viewed by others

Introduction
A wide number of conserved genetic and structural features have been identified regulating fin development in ray finned fish species, like zebrafish (Danio rerio), and imply shared mechanisms throughout evolution1. The embryonic development of pectoral fins in fish species is assumed to resemble limb development in higher vertebrates, with common molecular signals arising from a structure called the apical ectodermal ridge (AER)2,3. Only recently differences between fin and limb AER have been reported and hint at a fin specific cellular process3,4. Moreover, there are also eminent developmental differences between paired fins (pectoral fins) and unpaired fins (caudal, anal and dorsal fins). All unpaired fins arise from a common developmental precursor structure, called the median fin fold (mff), which is exclusively found in teleosts5. An increasing number of molecular processes and distinct genes are still being identified by forward and reverse genetic screens in zebrafish, revealing a complex network of factors necessary for correct median fin fold development and function6,7. Already more than 30 years ago changes of epidermal cell shape and modulation of the extracellular matrix (ECM) have been described as one of the essential factors for correct median fin fold and caudal fin morphogenesis in zebrafish8. Recently, the Wnt signaling pathway has been shown crucial for regulation of epithelial cell morphology by modulating laminin levels and thereby orchestrating correct ECM patterning in growing fins9. These early cellular steps of caudal fin development are prerequisites for subsequent processes; i.e. mesoderm cell migration, cell differentiation, fin growth, development of cartilage and bone and the gradual resorption of the median fin fold during juvenile stages10. Particularly fin rays are essential structural elements formed by the assembly of actinotrichia and lepidotrichia at the basal membrane of epidermal cells11,12. While lepidotrichia are segmented and calcified bone rays, actinotrichia are non-calcified fibers with a characteristic brush-shaped structure. Actinotrichia fibers display a transverse striation which can be observed in several fish species via electron microscopy suggesting these fibrils to be hyperpolymerized collagen13. Two collagens, Col2a1 and Col1a1, as well as actinodin proteins, encoded by and1, and2, and3 and and4, are essential for actinotrichia formation at the fin tips during fin development and regeneration14,15,16.
It is well accepted that a number of conserved molecular mechanisms are shared between extremity development during embryogenesis and fin regeneration in adult fish, a developmental process enabling complete replacement of lost tissues17,18. For both processes correct epidermal cell function, epithelial cell structure, and actinotrichia fiber assembly are essential to correctly build all skeletal and mesenchymal fin elements14,19,20. Structural factors, especially ECM proteins like integrins and laminins, have been implied in regulating Wnt signaling during regeneration and in correct assembly of the teleost fin6,9,21. Although a large number of involved “molecular players” have been described to date, not all factors necessary for correct ECM assembly in the regenerating caudal fin or correct median fin fold development and function have been elucidated yet.
FNDC3A protein has initially been described to be overexpressed in human odontoblasts22 and consists of up to nine fibronectin type III domains, which are a common feature of a large number of extracellular proteins acting by modulation of different signaling pathways23,24. Functional experiments in Symplastic spermatids (sys) knockout mice indicated that FNDC3A is essential for cell adhesion between spermatids and Sertoli cells, resulting in sterile males25. Further developmental functions of FNDC3A in vertebrates are still unknown and an association to extremity development in mammals has only been described recently by the Mouse Organogenesis Cell Atlas, showing expression of Fndc3a in epithelial cells of the limb AER26. The purpose of this study was to investigate potential functions of Fndc3a during vertebrate extremity deployment and regeneration in zebrafish (Danio rerio).
Results
Phylogenetic and syntheny analyses showed that the FNDC3A gene is highly conserved throughout vertebrate evolution and orthologues are not duplicated in ray-finned fish species (data not shown). In the zebrafish genome fndc3a is located on chromosome 15 and encodes in 29 exons for two different transcripts that are highly similar, with corresponding proteins of 1247 and 1217aa that only differ in a 30aa stretch at the N-terminus (ENSEMBL Zv9: 3,066,162-3,114,443 reverse strand; ENSDARG00000067569; ZFIN ID: ZDB-GENE-030131-7015; GenBank: XM_021466300.1, XM_021466301). Zebrafish Fndc3a protein (UniProt: A0A140LGL5) consists of one transmembrane domain located at the C-terminus, 9 fibronectin type III domains and one signal peptide located at the N-terminus. Amino acid alignment resulted in an up to 57% amino acid identity with 95% coverage, indicating a high level of conservation between human and zebrafish proteins. Furthermore, two fndc3a paralogues can be identified in the zebrafish genome: fndc3ba (chromosome 2; ENSDARG00000078179; ZFIN ID: ZDB-GENE-070510-1) and fndc3bb (chromosome 24; ENSDARG00000062023; ZFIN ID: ZDB-GENE-070510-2). Both genes share highest sequence similarities with FNDC3B and form a distinct subgroup aside from FNDC3A gens. Amino acid alignment comparison of both zebrafish paralogous to human FNDC3B show for Fndc3ba up to 68%% amino acid identity by 98% coverage, while Fndc3bb shows up to 56% by 98% coverage. Both zebrafish proteins show typical FNDC3 protein domain structure, by displaying one transmembrane and 9 fibronectin type III domains. Syntheny analyses furthermore indicated the location of both genes within two distinct duplicated genomic regions on zebrafish chromosomes 2 and 24. Both regions share up to 8 additional duplicated genes flanking zebrafish fndc3b genes, which are also located within the human FNDC3B locus. All three gene family members have not been functionally investigated in zebrafish yet.
Expression of fndc3a during early zebrafish development
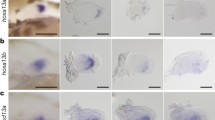
Earliest expression of fndc3a can be detected via RT-PCR and RNA-seq during blastula stages and indicate maternal transcripts of fndc3a (data not shown). To resolve the spatiotemporal expression of fndc3a during zebrafish development, we performed RNA in-situ hybridization experiments (Fig. 1). fndc3a transcripts were detected in a broad pattern and in number of different tissues, but showed cell type restricted expression within the tail bud region and the ventral median fin fold from 14 hpf onwards (hpf = hours post-fertilization; Fig. 1A,B; for visualization also of weak expression within the tailbud cells embryos shown in B are longer stained with NBT/BCIP). Expression in the tail bud region is changing during the next hours of development and could be detected apart from the median fin fold, in the cloaca, and in cells of the chordo neural hinge region (Fig. 1B). From 14 hpf onwards fndc3a was additionally present in distinct brain regions, the notochord, somites, pectoral fins and the caudal median fin fold, implying a rather broad and ubiquitous expression throughout zebrafish embryo development (Figs 1A and S4A).

Localization of fndc3a RNA and protein during embryonic zebrafish development. (A,B) Expression of fndc3a mRNA was detected in the tail bud and the median fin fold from 14 hpf onwards. fndc3a is rather broadly expressed during embryogenesis, but was highly expressed in caudal and pectoral fins, somites, notochord cells and distinct brain regions. (C,D) Detection of Fndc3a protein via immunofluorescence indicated similar regional localization as fndc3a mRNA in 22–48 hpf embryos. Furthermore, it showed intracellular accumulation of Fndc3a at notochord cells, at somite boundaries and epidermal cells at this stage. In-situ stained embryos shown in (A,B) differ in proteinase K incubation and NBT/BCIP staining times to visualize weak fndc3a expression in different tissues and stages. Dashed lines in (B) indicate planes of the corresponding numbered sections 1–5, in (C) notochord boundary and in (D) fin fold border. Fire LUT in (C,D) shows pseudo-colored images of Fndc3a immunofluorescence and marks regions of high and low intensities (highest to lowest signal: yellow, red, blue, black). cnh: chordo neural hinge; cl: cloaca; le: lateral edge; mc: mesencephalon; mff: median fin fold; mhb: midbrain hindbrain boundary (marked with chevron); nk: neural keel; no: notochord; nt: neural tube; pf: pectoral fin; sb: somite boundary; so: somites; tb: tail bud; rc: rhombencephalon. Scale bars: 100 µm, except higher magnification in (C): 20 µm.
Detection of Fndc3a protein localization was performed via immunofluorescence with a human FNDC3A antibody. Consistent with RNA in-situ hybridization this experiment showed similar regional localization of Fndc3a, e.g. in cloaca, the median fin fold region and the chordo neuronal hinge (22–24 hpf: Fig. 1C and 48 hpf: Fig. 1D). Specific Fndc3a protein localization could also be observed, as spots in notochord cells as well as in chevron shaped stripes between somite boundaries (higher magnification images in Fig. 1C). Moreover, this experiment clarified the cellular localization of Fndc3a at the cell membrane of epidermal cells during early stages of median fin fold development (higher magnification images Fig. 1C,D).
Generation and phenotypic investigation of fndc3a wue1/wue1 mutants
To further investigate fndc3a function during zebrafish development, we established a mutant line via the CRISPR/Cas9 system27,28. sgRNAs were designed to target the evolutionary highly conserved third fibronectin III domain of Fndc3a (Fig. 2A), as incomplete genome information about the 5′end of fndc3a at the initiation of the project prevented the identification of a distinct start codon for sgRNA targeting. The established fndc3awue1/wue1 mutant line has a 5 bp substitution leading to a premature Stop codon in exon 13 of fndc3a (Zv9: ENSDARE00000690608; fndc3awue1 line; ZFIN ID: ZDB-ALT-170417-3; Fig. S1A). The alteration was validated by sequencing of genomic DNA and cDNA (Figs 2A and S1) and could be detected continuously in subsequent inbred generations (data not shown). The three most likely, computationally predicted off-site targets were sequenced and no off-site sequence alterations were detected in fndc3awue1/+ or fndc3awue1/wue1 mutants (Fig. S1B).

Generation and phenotype of fndc3awue1/wue1 zebrafish mutants. (A) The CRISPR/Cas9 system was used to target exon 13 in the zebrafish fndc3a gene coding for the third fibronectin type III domain (nucleotides marked in light blue indicate sgRNA target sequence and in red the region of mutated sequence). (B) fndc3awue1/wue1 mutants showed straightened tail buds (22 hpf; n = 19/40), kinked tails (48 hpf; n = 27/100), and caudal fin deformations (120 hpf; n = 9/41) during the first days of embryonic development. (C) A fraction of adult fndc3awue1/wue1 mutants displayed weak (n = 15/71) to strong (n = 6/71) caudal fin phenotypes and tail malformations. (D) qPCR quantification of relative fndc3a expression levels in genotypic different groups of embryos indicated reduction of fndc3a transcripts in fndc3awue1/+ and fndc3awue1/wue1 (ΔΔCt calculation; significance levels of a 2-sided paired student t-test are given). Investigation of protein domains shown in A has been performed via the SMART database (Simple Modular Architecture Research Tool; http://smart.embl-heidelberg.de)69. Black arrows indicate developmental malformations. Scale bars for whole embryos: 250 µm; scale bars for tail magnifications: 100 µm.
Visual inspection of fndc3awue1/wue1 mutants showed a tail bud and median fin fold phenotype in homozygous embryos that could first be observed 22 hpf (Fig. 2B). Almost half of fndc3awue1/wue1 embryos exhibited straightened tail buds and reduced ventral fin fold structures (22 hpf). During subsequent stages of development fndc3awue1/wue1 mutant embryos displayed kinked tails (48 hpf) and caudal fin fold malformations (120 hpf). Due to the observed variability and the expectation of a stronger, more consistent phenotype after cellular stress, we further assessed the temperature sensitivity of the fndc3awue1/wue1 phenotype29,30. The experiment revealed that raised temperatures indeed result in an increased number and severity of the caudal fin phenotype in mutants (Fig. S2).
Of adult fndc3awue1/wue1 fish approximately 30% displayed alterations of the caudal fin shape and the posterior body part. The observed phenotype ranged from minor fin shape changes in the majority of affected fish, up to axis shortening and stronger caudal fin deformations (Fig. 2C). We did not observe changes in phenotype severity or appearance rates in subsequent, homozygous generations, excluding a potential stronger maternal zygotic effect in fndc3awue1/wue1 or mitigation of the phenotype. Also, contrary to the mouse knock-out lines the fndc3awue1/wue1 line did not show reduced fertility rates.
The observation of variable adult phenotypes, incomplete phenotypic penetrance and temperature sensitivity of the embryonic phenotype led us to the assumption that fndc3awue1/wue1 mutants are hypomorphic. We tested this hypothesis by qPCR experiments, which revealed only partial loss of fndc3a transcripts due to nonsense mediated decay. We quantified fndc3a mRNA expression in three independent embryo groups with two primer pairs targeting different mRNA regions (Fig. 2D; primer sequences are given in Table S1). Relative to wildtype control fish fndc3a transcripts in fndc3awue1/+ and fndc3awue1/wue1 mutants were reduced to approximately 60% and 30% relative expression, respectively. Moreover, strong reduction of Fndc3a protein levels in mutants could be detected by immunofluorescence analyses on fndc3awue1/wue1 mutants (Fig. S3).
To further validate the fndc3a phenotype we aimed to introduce a larger genomic deletion or multiple sequence alterations within the fndc3a locus by application of an additional sgRNA targeting exon 18. The corresponding gRNA was synthesized and co-injected with exon 13 sgRNA. The resulting transient phenotype indicated more severe tail fin malformations, i.e. strong caudal fin reduction and tail curling, when compared to fndc3awue1/wue1 mutants (Fig. S4). This double fndc3a CRISPR phenotype could however not be maintained and next generation crossings did not result in a stable line. Nevertheless, it supported and confirmed our observations of the fndc3awue1/wue1 mutant and pointed at a noticeable influence of Fndc3a on caudal fin development. No alterations in the development of pectoral fins could be detected in either fndc3awue1/wue1 or the transient double CRISPR mutants (Figs S4 and S5).
Reduction of Fndc3a function results in median fin fold defects
To elucidate the molecular causes underlying the early changes in caudal fin development of fndc3awue1/wue1 mutants we analyzed expression patterns of different, well established markers, expressed in the median fin fold tissues (Table 1; Fig. 3).

Normal development of the ventral median fin fold is altered in fndc3awue1/wue1 mutants. (A) Investigation of fras1, hmcn1, hmcn2, bmp1a, and fbln1 expression in the tail bud 20–22 hpf indicated loss of mesenchymal cells in the median fin fold in fndc3awue1/wue1 mutants (arrows; fras1: 9/32; hmcn1: 11/26; hmcn2: 24/43; bmp1a: 22/45; fbln1: 16/32). (B) Expression of mesenchymal markers in posterior fins of 48 hpf old embryos was slightly changed following loss of fndc3a (fras1: 10/24; hmcn2: 12/25; bmp1a: 8/21; fbln1: 9/24). (C) Ventral fin folds in 48 hpf old embryos did not show loss of mesenchymal markers after fndc3awue1/wue1 mutation. (D) Expression of and1, an essential marker for actinotrichia formation, in fndc3awue1/wue1 in the ventral fin fold was partly lost in 22 hpf embryos (12/25), but recovered in 48 hpf old embryos to a reduced expression level (control n = 16; weaker expression in fndc3awue1/wue1 embryos n = 10/18). Dashed lines in (A,D) indicate planes of shown sections. Dashed lines in (B,C) indicate fin boundaries. Scale bars: 50 µm
All investigated early median fin fold genes were still expressed in the fndc3awue1/wue1 embryos but showed altered patterns or intensity (Table 2). Affected regions in mutants correlated with the regions of investigated fndc3a expression. Observation of further mesodermal markers in fndc3awue1/wue1 mutants 20–22 hpf additionally confirmed misplaced myod expression in ventral positions and indicated changes in chordo neural hinge cells by reduced shha, ta(ntl), and fgf8 expression (Fig. S6), hinting at a structural or steric effect on tail development after Fndc3a reduction. Moreover, gene expression analysis indicated that fndc3awue1/wue1 embryos develop only temporal defects during early median fin fold and start to recover normal gene expression patterns in median fin fold cells 48 hpf. In summary, the in-situ experiments indicated, that most prominently apical cells expressing fras1 and hmcn1 located at the ventral median fin fold were influenced by reduced levels of Fndc3a during the first two days of zebrafish development.
To validate specific effects on median fin fold development in fndc3awue1/wue1 mutants Morpholino knockdown and rescue experiments were conducted (Figs S7 and S8). Phenotypic investigation of fndc3a morphants clarified that similar median fin fold malformations and comparable changes in median fin fold gene expression were induced in a dosage dependent manner in the first 48 h of development (Fig. S7B–D). Notably, injection of fndc3a Morpholino into fndc3awue1/wue1 embryos resulted in an enhanced number of embryos showing tail phenotypes and supports the hypothesis of a hypomorphic mutation (Fig. S7B,C). Rescue and overexpression experiments were performed by injection of full-length human FNDC3A mRNA (Fig. S8). The tail phenotype and median fin fold gene expression in fndc3awue1/wue1 mutants could be rescued after moderate RNA supplementation [25 ng/µl]. Injection of FNDC3A RNA at this concentration into wildtype embryos did not result in a raised number of tail malformations (Fig. S8B). Noteworthy, injection of higher RNA doses [50 ng/µl] resulted in a raised number of prominent tail deformations, especially in fndc3awue1/wue1 mutants and to a smaller extent also in injected wildtype embryos. These experiments indicate a narrow threshold level for Fndc3a to fulfill its function in early median fin fold cells and support the observed fndc3awue1/wue1 phenotype.
fndc3a wue1/wue1 mutants show actinotrichia breakdown and defects in basal epidermal cells
Since our expression and knock-down observations implied a function of Fndc3a during early median fin fold establishment we subsequently investigated a potential link to epidermal cells and to associated structures, like actinotrichia fibers, in fndc3awue1/wue1. Actinotrichia were visualized by either differential interference contrast (DIC) microscopy (Fig. 4A) or immunofluorescent staining of Col2a, a structural component of actinotrichia (Fig. 4B). This experiment revealed that actinotrichia fibers were still present in fndc3awue1/wue1 mutants, but displayed obvious structural alterations and signs of breakdown in the caudal fin. While wildtype fish at 52 hpf showed radiant symmetrical arrangement of Col2a in the developing caudal fin, fndc3awue1/wue1 embryos partly lacked these structures and depicted unstructured, crumbled collagen fibers in the fin mesenchyme. High levels of remaining Col2a in fndc3awue1/wue1 mutants could be detected in apical cells at the fin border, which were visible as distinct cells in DIC microscopy.

The fndc3awue1/wue1 mutation results in structural defects in epidermal cells during fin development. Visualization of actinotrichia by either differential interference contrast microscopy (A) or immunofluorescence staining of Col2a (B) showed loss of mature actinotrichia in fndc3awue1/wue1 mutants 52 hpf (control n = 0/17; fndc3awue1/wue1 n = 19/26; white arrows indicate lost actinotrichia and Col2a accumulation in apical cells of the median fin fold). (C) Investigation of fndc3awue1/wue1 mutants showed reduced median fin fold width and reduced TP63 positive epidermal cell number in the ventral median fin. (D,E) Quantification of median fin fold width and TP63 positive cells in 24 and 48 hpf embryos (Mann-Whitney U test; p < 0.05; two-tailed, significant U value changes are indicated with asterisks). (F) Ultrastructural analysis of dorsal and ventral fins of control and fndc3awue1/wue1 mutants revealed breakdown of actinotrichia fibers and cellular malformations in cells of the basal epidermal layer in the ventral fin folds of 52 hpf old embryos (circles indicate misplaced fibers; dashed lines indicate the basal membrane; yellow brackets indicate outer epidermal cells and green brackets indicate basal epidermal cells; black asterisks indicate cavities). a: actinotrichia; bel: basal epidermal layer; epi: epidermis; mff: median fin fold; m: mesenchymal cell; N: nucleus; oel: outer epidermal layer.
Potential cellular reasons for actinotrichia breakdown and Col2 misallocation can be numerous. Based on the expression timing and localization of fndc3a RNA and Fndc3a protein during median fin fold development and the reduced function, we assumed that disturbance of epidermal cell structure might be causative for the observed phenotype and that the normal cellular organization within the developing mff is lost8,16. Indeed, investigation of TP63, an epithelial stem cell marker, expressed in all epidermal cells of the basal layer37,38 (Fig. 4C), showed a reduced number of epidermal cells in the ventral fin fold, lack of ventral fin fold structures and reduced median fin fold width at this position (arrows in Fig. 4C; quantification in Fig. 4D) in 24 hpf fndc3awue1/wue1 mutants. At 48 hpf fndc3awue1/wue1 mutants then developed outgrowing ventral median fin folds and regained ventral structures. Although these were still shorter in length they did not display a significant lower number of epidermal cells in comparison to control embryos (Fig. 4C; quantification in Fig. 4E), indicating cellular recovery of the median fin fold structure at this time. Additionally, fndc3awue1/wue1 mutants displayed aggregation of epidermal cells at the fin fold border, which was not observed in control embryos (Fig. 4D10). Consistent with the phenotypic investigation, in pectoral fins no alteration in Col2a or TP63 immunofluorescence staining could be detected (Fig. S4C).
Transmission Electron Microscopy (TEM) further clarified the cellular consequences of Fndc3a reduction in the epidermal cell layer. While fndc3awue1/wue1 mutant embryos showed only a slight reduction of actinotrichia in dorsal median fin folds (lower row Fig. 4F), an almost complete loss of actinotrichia in ventral median fin folds could be observed. Only small remains, showing the characteristic stratification, were still present at the basal membrane, in addition to misplaced collagen fibers not attached to the basal membrane (circles Fig. 4F). While mesenchymal cells, the outer and basal epidermal cell layer were present, the sub-cellular structure of epidermal cells was altered after fndc3a reduction. Cavities between the outer and basal epidermal layers (asterisks in Fig. 4F), as well as changes in the ECM surrounding these cells could be detected. This observation strongly implies that the disruption of actinotrichia fibers and the described effects on TP63 positive cells after Fndc3a reduction are due to cellular defects in the basal epidermal cell layer.
Fin regeneration is also temporally influenced in fndc3a wue1/wue1 mutants
Besides their function during fin development, actinotrichia and epidermal cells possess essential roles during regeneration of adult caudal fins after amputation16,39. Similar to their role in fin development, Col2a and Col1a were identified to be necessary factors of actinotrichia formation also during regeneration40, implying comparable underlying cellular processes. We therefore hypothesized that reduction of Fndc3a function might also interfere with fin regeneration.
Regeneration experiments on adult caudal fins confirmed fndc3a expression and localization of Fndc3a protein prominently in epidermal cells of regenerates between 2 and 8 days past amputation (dpa; Fig. 5A). Investigation of fin regeneration in fndc3awue1/wue1 compared to wildtype fish led to two remarkable observations. First, regenerates looked opaque and disorganized, and aberrant cells attached to the epidermal layer were eminent between 4 dpa and 8 dpa in fndc3awue1/wue1 mutants (arrows in Figs 5B and S9). Second, the apical fin borders of regenerates looked disorganized and clumped in fndc3awue1/wue1 mutants (arrows in Fig. 5B).

Interference with Fndc3a function during fin regeneration results in epidermal cells defects. (A) Expression of fndc3a and localization of Fndc3a in regenerates could be detected in epidermal cell layers. (B) Phenotypical and histological investigations of fin regeneration in fndc3awue1/wue1 mutants indicated abnormal regeneration by showing unorganized fin borders at the regenerative front and aberrant epidermal cells (arrows; abnormal regenerate phenotypes, control: 3/19; fndc3awue1/wue1: 13/19). (C) Col2a protein localization visualized via immunofluorescence on 8 dpa regenerates showed accumulation of collagen in abnormal cells of fndc3awue1/wue1 mutants (white arrows; n = 6 for each group). (D) TP63 staining in regenerates of fndc3awue1/wue1 mutants 4 and 8 dpa indicated disorganization of epidermal cells (arrows indicate epidermal thickening, cavities within the regenerate and abnormal cells; n = 4 for each group). (E) TEM analysis showed accumulation of electron dense material in the Golgi apparatus of abnormal cells attached to the epidermal layer (arrow). (F) TEM analyses of actinotrichia in fin regenerates 4 dpa revealed loss of these fibers located near the basal membrane of epidermal cells in fndc3a depleted fish (arrows). Arrowheads indicate amputation site. a: actinotrichia; epi: epidermis; dashed lines indicate basal membrane boundary.
Subsequent histological investigation via H&E and immunofluorescence staining of these regenerates confirmed that the aberrant cells were loosely attached cells that depicted high levels of Col2a at the regenerative front (arrows in Fig. 5B,C). TEM analysis further clarified that these cells were still attached to the epidermal cell layer and incorporated electron dense material in their Golgi apparatus and in intracellular vesicles (arrow in Fig. 5E). TP63 staining for epidermal cells in 4 to 8 dpa regenerates then showed interference with normal regenerate structure and disturbance of epidermal cell layers in fndc3awue1/wue1 mutants (Fig. 5D). It also showed that actinotrichia fibers of fndc3awue1/wue1 mutants only depicted loose collagen filaments adjacent to the membrane layer of epidermal cells and were lacking the prominent compact structure of bundles of stratified actinotrichia fibers in wildtype fish (Fig. 5F).
Furthermore, consistent with the hypomorphic fndc3awue1/wue1 phenotype during fin development, the observed effects could be enhanced by keeping fish at a raised temperature of 32 °C during regeneration (Fig. S8). Although prominent cellular abnormalities were detected throughout the first days past amputation, a significant difference in regenerate length appeared only at 6 dpa. No significant differences in overall tail length growth could be detected between mutant and wildtype fish at all other investigated stages (Fig. S10). All regenerating fins grew to their former size after ~14 dpa (data not shown), but small changes in fin morphology, e.g. cooped fin rays and loss of interray tissues, could be observed in fndc3awue1/wue1 mutants even 6 weeks past amputation (Fig. S9C). All this led to the assumption that a potential reason for the observed weak and transient effects is an altered epidermal cell organization in regenerates after reduced Fndc3a function, similar to the processes observed in the median fin fold during early development.
fndc3a wue1/wue1 mutants display ECM alterations in median fin folds and in caudal fin regenerates
Correct assembly of actinotrichia and correct caudal fin morphology are greatly dependent on the extracellular composition and the cell shape of surrounding epidermal cells during development and regeneration. These cellular characteristics within the median fin fold determine correct signaling, e.g. by Wnt signals or cell behavior18,41. Fibronectin domain containing proteins like Fndc3a have been linked to functions during ECM assembly and maintenance42. Thus, we assumed that Fndc3a function might provoke cell shape or ECM alterations in epidermal cells.
To follow-up on this we analyzed F-actin and β-catenin localization in the median fin folds of wildtype and fndc3awue1/wue1 mutants (Fig. 6). Cell boundaries of wildtype embryos 22 hpf displayed a dense, stereotypical assembly of epidermal cells in the ventral median fin fold (Fig. 6A,B). Reduction of Fndc3a resulted in clustering of these cells, altered cell shapes (arrows Fig. 6A) and appearance of cavities (dashed lines Fig. 6A). The cavities are zones within the fin folds showing no F-actin and no nuclear staining, suggesting cell free spaces. β-catenin in wildtype embryos 24 hpf displayed homogeneous membranous localization in cells of the ventral median fin fold and nuclear localization in epithelial cells at the fin border. In fndc3awue1/wue1 mutants however uniform β-catenin localization was impaired and instead appeared as accumulations of protein in a speckled fashion within the cytoplasm (arrows in Fig. 6B). Nuclear localization of β-catenin in fndc3awue1/wue1 mutants was still clearly detected at the apical fin border (arrowheads in Fig. 6B).

Correct ECM structure in the median fin fold and regenerating caudal fins is hampered in fndc3awue1/wue1 mutants. (A,B) F-actin in the median fin fold was visualized by phalloidin staining and localization of β-catenin by immunofluorescence (n = 6 for each group, 22–24 hpf). Cellular organization of ventral median fin fold cells and ECM matrix was symmetrically structured in control embryos and showed nuclear localization of active Wnt signals in apical cells (white arrowheads in B). fndc3awue1/wue1 mutants depicted cellular alterations and unstructured ECM assembly by showing irregular cell shapes (arrows in A), cavities within the fin fold (dashed lines in A) and speckled accumulation of β-catenin between cells (arrows in B). Nuclear localization of β-catenin in apical cells was maintained (arrowheads in B). (C,D) Fin regenerates of fndc3awue1/wue1 mutants stained for F-actin showed regenerate abnormalities (arrows in C), irregular regenerate borders (dashed lines in C) and cellular cavities (dashed lines in D; n = 4 for each group). (E,F) Fin regenerates of fndc3awue1/wue1 mutants stained for β-catenin depicted divergent ECM assembly (arrows in E), appearance of abnormal cells loosely attached to the regenerate (arrows in F) and cavities (dashed lines in F; n = 3 for each group). Images either show maximum intensity projections (30 to 40 single z-slices; z-distance: 1.5 µm) or a representative higher resolution single z slice.
Comparison of wildtype and fndc3awue1/wue1 mutants clarified that similar distinctive features were also observed during caudal fin regeneration. Irregular regenerate borders and cellular alterations of fndc3awue1/wue1 mutants could be detected with both F-actin (dashed line and arrows Fig. 6C) and β-catenin staining (arrows in Fig. 6E). High resolution confocal microscopy of single z slices clarified, that the exhibited fragmented structures in F-actin staining in 2 dpa regenerates are cavities within the blastema which could also be seen with β-catenin staining 4 dpa (dashed lines Fig. 6D,F). Furthermore, it revealed β-catenin negative, detached cells outside of the regenerate (arrow in Fig. 6F). In summary, our observations in fndc3awue1/wue1 mutants indicate a partial loss of cellular structure and adhesion within the blastema during caudal fin regeneration in adults as well as within the median fin fold during early embryonic development.
Discussion
fndc3a expression in developing zebrafish embryos was first detected as maternal transcripts during blastula stages. During segmentation period stages (14 hpf) fndc3a expression was most prominent in the tail bud and later in apical cells of the ventral median fin fold, the pectoral fins, the notochord and in cells of the chordo neural hinge. Fndc3a protein localization showed comparable patterns and was subsequently detected in the cell membrane of ectoderm derived cells and in notochord cells. Spatiotemporally similar expression patterns in the zebrafish median fin fold have been described for other prominent genes also essential for vertebrate extremity development, e.g. dlx5a and tp6338,43,44. Comparison of zebrafish fndc3a during extremity development to mouse Fndc3a expression shows a partially similar pattern, i.e. early in the AER of the limb bud and later in interdigital regions of feet (Gene expression database; http://www.informatics.jax.org/expression.shtml), and was validated by single cell sequencing in the Mouse Organogenesis Cell Atlas (MOCA)26. Although expression of Fndc3a was detected in the AER of mice E10.5 and E11.5, mouse Fndc3a knockout result in male infertility, while defects in limb development were not described25.
The CRISPR/Cas9 generated fndc3awue1 mutation in zebrafish, presented in our study, resulted in a premature Stop codon within exon 13. The mutation did not lead to complete nonsense mediated decay and full loss-of-function phenotype, as residual fndc3a mRNA expression could be detected and the observed phenotypes are rather weak and partly transient. Our data implies a potential residual expression level and only diminished function of Fndc3a in these mutants. Although tissue specific effects can also accrue when changed protein levels are interfering with signaling thresholds, signaling equilibriums or tissue structure. Morpholino knockdown and RNA overexpression experiments in wild type and in fndc3awue1/wue1 mutants, respectively, displayed on the one hand a consistent phenotype of the mutant and further a partial rescue of the induced mutation. On the other hand, the experiments resulted also in an increased severity after Morpholino and RNA injection into fndc3awue1/wue1, implying that the induced mutation does not abolish the function of the gene completely, but rather leads to a reduced function and has to be considered a hypomorphic mutation. RNA injections also showed, that at higher concentrations of FNDC3A RNA [50 ng/µl] changed fndc3a levels result in tail deformations. Interestingly, in mutants the phenotype penetrance was even enhanced at these higher concentrations. This observation hints to a narrow threshold level of functional Fndc3a at the mff to guarantee its normal development and indicates alternative or compensating cellular processes for mff development in the mutants. Our results cannot exclude a compensation mechanism for fin development and regeneration in fndc3awue1/wue1 mutants, which despite of reduced Fndc3a function would be able to recover cellular functions and ultimately lead to a rather normally developed caudal fin during larval development and in adult fish. Compensation could either be facilitated by residing functional Fndc3a proteins, as indicated by remaining transcripts in mutants, by other fibronectin homologues, or by activation of known signaling networks during fin development and regeneration18. Especially tissues, like neuronal tissues, notochord and pectoral fins, in which fndc3a is strongly expressed but which display no defects in mutants should be investigated for compensatory mechanisms in close detail to reveal the involved factors for this process. Potential compensating factors might be determined by looking at other orthologues of the fndc3 gene family, e.g. fndc3ba and fndc3bb, although expression patterns as well as function of these genes have not been investigated yet. Dorsophila miles to go mutants develop after complete depletion of the FNDC3 orthologue severe developmental defects, pupal lethality, neuromuscular junction defects and indicate a rather severe embryonic phenotype after complete loss of all FNDC3 functions45. Our experiments in zebrafish targeted one vertebrate FNDC3 orthologue and therefore might resemble only a fraction of the full FNDC3 loss-of-function phenotype.
A specific phenotype in the developing median fin fold of fndc3awue1/wue1 mutants i.e. loss of ventral median fin fold cells, loss of gene expression domains in the ventral fin fold, loss of actinotrichia fibers and Col2a accumulation was observed. Most prominently a number of early markers in the ventral median fin fold were reduced or altered in fndc3awue1/wue1 mutants, indicating the need of cellular Fndc3a function in this specific region and early phase of development. In contrast to more globally expressed genes linked to early caudal fin development and to actinotrichia deposition in zebrafish, e.g. pinfin mutants lacking Fras1 function or nagel mutants lacking Hmcn1 function6, the hypomorphic phenotype in fndc3awue1/wue1 mutants is milder and locally restricted corresponding to its more specific expression pattern. One cellular consequence of Fndc3a reduction was disturbance of the actinotrichia and collagen fibers assembly process during development, as indicated by altered Col2a location in fndc3awue1/wue1 mutants and by EM imaging. Collagen helix assembly is performed at the ER and is controlled by chaperones, especially Hsp47/SerpinH146. Interestingly knockdown of hsp47/serpinH1 results in a phenotype resembling fndc3awue1/wue1 i.e. actinotrichia organization failure and deformations of regenerating fin47. In comparison to published loss-of-function mutations in zebrafish collagen genes (e.g. col1a1: chi, Chihuahua; col9a1: prp, persistent plexus) which result in characteristic, severe bone growth defects, variable skeletal dysplasia and vascular plexus formation48,49, the fndc3awue1/wue1 mutation has only a mild effect. Interference also with expression domains of mesenchymal markers, e.g. myoD, suggests that Fncd3a is required for the establishment of ventral cell fates of the developing median fin fold, potentially by setting up the correct cellular structure in this region.
The process of median fin fold development is tightly regulated by modulation of epidermal cell shape and correct ECM assembly8,9. TP63 positive epidermal cells are still present in the fndc3awue1/wue1 mutants during median fin fold development 20 to 48 hpf, but show reduced numbers at ventral positions, delayed ventral fin fold growth and altered morphological properties. Fndc3a shares fibronectin domain III protein domains with other well studied factors like Fibronectin 1, which are known to interact with prominent ECM proteins50 and are essential for matrix assembly51. The observed breakup of correct epidermal cell assembly and appearance of cavities in the ECM of basal epidermal cells after reduction of Fndc3a function indicate that Fndc3a might play a similar role in correct ECM assembly and establishment of correct cellular structures in the median fin fold. Potential downstream effects like misplaced mesodermal cells, loss of actinotrichia fibers and detached epidermal cells are most likely explained by deranged ECM structure in the early epidermal cell layer after Fndc3a reduction. Mesenchymal cells for example migrate after their induction along predetermined ECM structures and actinotrichia fibers to form the fin skeleton33. Irregular skeletal formation of caudal structures, thus, might be a consequence of early developmental irregularities. Future experiments will have to clarify if binding of Fndc3a to integrins is abandoned in fndc3awue1/wue1 mutants, thereby directly influencing cell adhesion, basal epithelium establishment, and signaling52,53. This idea is supported by the similarity of fndc3awue1/wue1 fin fold defects to laminin (lmna5) and integrin (itga3) mutants6,9,21 and the occurrence of aberrant cells in the epidermal layer of regenerates in the fndc3awue1/wue1 mutants. Moreover, our experiments do not rule out a potential intracellular function of Fndc3a. Protein localization by Carrouel et al. (2008) clearly detected FNDC3A in the Golgi apparatus of human odontoblast. Reduction of Fndc3a function therefore may also result in hampered protein processing within this organelle and thereby interfere with modification of ECM proteins or membranous export.
The severe tail phenotype observed in a minority of adult fndc3awue1/wue1 fish and in transient double CRISPR injected individuals (Fig. S4) suggests a stronger phenotype after complete loss of Fndc3a and points towards interference with prominent signaling pathways, e.g. BMP and/or TGF-beta signaling. Especially the phenotypic similarity to smad5 (somitabun or piggytail)54,55, tll1 (mini fin or tolloid)7,56,57 and bmp1a (frilly fins)35 zebrafish mutants imply an interplay between Fndc3a function and these signaling pathways. The activation of serine/threonine kinase receptors of the TGF-beta superfamily results in phosphorylation of pathway specific intracellular R-SMAD proteins, i.e. SMAD1/5/9, and thereby executing their transcriptional function58. Initial experiments in fndc3awue1/wue1 mutants show that at 22 hpf active TGF-beta/BMP signals as essential patterning factors are lost at ventral median fin fold positions (Fig. 7). This observation is in accordance with investigations focusing on Fndc3b/fad104, which state direct protein-protein interaction of Fndc3b to SMAD proteins and the ability for Fndc3b to act as regulator of BMP/SMAD signaling during calvarial ossification in mice59,60. At the moment a similar interaction for Fndc3a and SMAD proteins has not been reported, but our experiments strongly indicate a connection of the fndc3awue1/wue1 phenotype to loss of SMAD transmitted BMP signals in zebrafish.

Investigation of TGF-beta/BMP signals in the median fin fold of fndc3awue1/wue1 mutants. Images show tail structures of representative control and fndc3awue1/wue1 mutant 22 hpf embryos stained for phosphorylated SMAD (pSMAD1/5/9) by immunofluorescence. Lack of pSMAD positive cells is observed in ventral median fin fold structures in fndc3awue1/wue1 mutants. All pictures display maximum intensity projections. White arrows indicate loss of pSMAD signals. Fire LUT shows pseudo-colored pSMAD1/5/9 signals. Dashed white lines indicate median fin fold borders. Scale bars: 50 µm.
Besides median fin fold development we investigated a potential function of Fndc3a during caudal fin regeneration. In accordance with median fin fold development, regeneration and blastema formation depends on correct epidermal cell assembly and a distinct ECM structure61,62. We initially detected fndc3a transcripts and Fndc3a protein in the distal blastema and in epidermal cell layers of regenerates 2 to 8 dpa. In fndc3awue1/wue1 mutants we observed several temporal effects on caudal fin regeneration i.e. detached epidermal cells, Col2a accumulation, disorganized epidermal cells layers and lack of actinotrichia. These are comparable to the observed developmental phenotype within the median fin fold and indicate ECM malformation to be causative for this effect on adult structures as well. Our regeneration experiments hint to rather minor and temporal effects of Fndc3a on regeneration during initiation and the first few days of regeneration.
In summary, investigation of fndc3a expression and function in zebrafish reveals a transient and spatially restricted role of this genetic factor during extremity development and during fin regeneration (Fig. 8). The observed effects after reduction of Fndc3a function on actinotrichia fibers and correct fin morphology are probably secondary and provoked by cellular changes. Fndc3a can influence TP63 positive epidermal cells by altering cell shape or cell adhesion during extremity development. Most likely disruption of correct ECM structure in basal epidermal cells is the consequential underlying cellular mechanism responsible for the observed fin malformations and signaling changes. Our results therefore demonstrate a cellular link between median fin fold development and caudal fin regeneration due to the necessity of correct cell shape and tissue cohesion in both processes via Fndc3a.

Model of fndc3a function during zebrafish median fin fold development and caudal fin regeneration. Expression and localization of fndc3a was detected during early phases of median fin fold development and in caudal fin regenerates. Reduced Fndc3a function resulted in prominent changes of ECM structure in epidermal cells. During development this results in impaired collagen assembly of actinotrichia fibers and loss of ventral median fin fold cells. During regeneration altered ECM structure results in actinotrichia breakdown and detached epidermal cells.
Materials and Methods
Animal maintenance
Laboratory zebrafish embryos (Danio rerio) of the AB/TU and AB/AB strain (ZDB-GENO-010924-10; ZDB-GENO-960809-7) were maintained as described by Westerfield63 under standard aquatic conditions at an average of 24 °C water temperature. Embryos were staged by morphological characteristics according to Kimmel et al.64 hpf indicate hours-post fertilization at 28.5 °C. All procedures involving experimental animals were performed in compliance with German animal welfare laws, guidelines, and policies. Generation of fndc3awue1/wue1 mutants and fin clipping was approved by the Committee on the Ethics of Animal Experiments of the University of Würzburg and the “Regierung von Unterfranken” (Permit Number: DMS-2532-2-13 and DMS-2532-2-9). The generated fndc3awue1/wue1 line was submitted to ZFIN.org (ZFIN ID: ZDB-ALT-170417-3).
CRISPR/Cas9 system and construction of short guiding RNA (sgRNA) constructs
For oligo cloning of sgRNA target sequences the previously published pDR274 vector was used27 (Addgene Plasmid #42250; sequences of primers used in this study are given in Table S1). For cas9 RNA synthesis the MLM361327 (Addgene Plasmid #42251) or the pCS2-nCas9n vector28 (Addgene Plasmid #47929) were utilized. Both vectors were purchased from Addgene (www.addgene.org; Cambridge, USA). For designing and constructing of sgRNAs the open access ZiFit Targeter software (http://zifit.partners.org/ZiFiT/) was used. Specific target sites were identified by alignments of zebrafish and human sequences. sgRNA target site (GGATTCCAGGCCAGTTATGA) is located in exon 13 of fndc3a (ENSEMBL Zv9 Transcript: ENSDART00000097261) and targets the second Fibronectin type III domain, while sgRNA target site in exon 18 (GGCGTACAGTGGTTCGGCTC) targets the third Fibronectin type III domain.
sgRNA transcription and microinjection
sgRNAs were transcribed via the MAXIscript T7 kit (Ambion/ life technologies, Darmstadt, Germany) and were puridied via phenol/chloroform extraction. Cas9 RNA was transcribed via the mMESSAGE mMACHINE kit (Ambion/ Life Technologies, Darmstadt, Germany) and subsequently cleaned via RNeasy purification kit (Qiagen, Venlo, Netherlands). If the MLM3613 vector was used for Cas9 Synthesis, polyA tail synthesis was performed with E.coli Poly(A) Polymerase (New England Biolabs, Ipswich, MA, USA) prior to purification. One cell stage zebrafish embryos were injected with solutions comprising sgRNA (25–50 ng/µl each), cas9 RNA (75–100 ng/µl), Phenol red (pH7.0; 0.05% final concentration; for visualization of injection solution) and Fluorescein isothiocyanate-dextran (Sigma-Aldrich; 1 mg/µl). Positively injected embryos were identified 24 hpf by transient green fluorescence of Fluorescein isothiocyanate-dextran and were raised for line establishment.
Whole mount RNA in-situ hybridization
RNA in situ hybridization was performed according to standard protocols65,66. Proteinase K incubation and NBT/BCIP staining times were adjusted to age of embryos and to investigated tissues. RNA probes were synthesized from cloned partial mRNA sequences of target genes using the DIG or FLU RNA Labeling Kit (Roche, Basel, Switzerland). We used a fndc3a cDNA fragment of 595 bp size (used primers: zf_fndc3a_ribo_fwd2 and zf_fndc3a_ribo_rev2) to synthesize a specific anti-sense RNA probe. Sense probes were synthesized as negative control for each anti-sense probe and were used under the same reaction conditions. Primers used for probe cloning are listed in Table S1.
Immunofluorescence and histology
Immunofluorescence was performed on embryos, sections or on whole regenerating fins according to standard protocols16,67. Primary antibodies used: Fndc3a (HPA008927; Sigma-Aldrich; dilution 1:50–100; Antibody Registry: AB_1078899), Col2a (II-II6B3; anti-mouse; The Developmental Studies Hybridoma Bank; dilution 1:500; Antibody Registry: AB_528165), β-catenin (610153; BD Biosciences; dilution 1:250; Antibody Registry: AB_397554), TP63 (ab735; Abcam; dilution 1:200; Antibody Registry: AB_305870), Phospho-Smad1 (Ser463/465)/Smad5 (Ser463/465)/Smad9 (Ser465/467) clone D5B10 (13820; anti-rabbit; Cell Signaling Technology; 1:200 dilution; Antibody Registry: AB_2493181). Except Fndc3a, all used antibodies have been used previously for experiments in zebrafish and validated protocols have been previously published. Specificity of the used Fndc3a antibody was validated by the Human Protein Atlas project for usage in human tissues (https://www.proteinatlas.org/ENSG00000102531-FNDC3A/antibody). Validation for usage in zebrafish was performed by epitope sequence analysis, antibody dilution series, and incorporation of adequate negative controls in all experiments. Cell nuclei were counterstained with Hoechst 33258 (Sigma-Aldrich; dilution 1:5000). F-Actin was stained with Acti-stain 488 phalloidin (Cytoskeleton, Inc.; PHDG1-A; dilution 1:20). Whole mount alcian blue staining was performed according to Walker and Kimmel68. H&E staining on 10 µm paraffin sections was performed according to a general histology protocol using Mayer’s Hematoxylin and Eosin.
Fin regeneration experiments
For fin amputation experiments adult zebrafish (mixed sex, age 3–5 months, 3 independent experiments) were anesthetized with tricaine (MS-222; 3-amino benzoic acid ethyl ester; final concentration ~150 mg/l) prior to amputation of ~20% length the of caudal fin. Images were taken 0, 2, 6, 8 and 10 days post-amputation (dpa) under a stereomicroscope. To increase the expected phenotype fish were kept at an increased temperature (32 °C) during the experiments. Regenerate samples for immunofluorescence, in-situ hybridization, genotyping, or histology were taken by a second amputation anterior to the first lesion site (2, 4, 6 and 8 dpa).
Image acquisition and quantification
Images were acquired depending on the experiment either with a Leica S8 APO Stereomicroscope (whole embryos), a Zeiss Imager A1 (in-situ hybridizations) or a Nikon A1+ Laser scanning confocal microscope (Immunofluorescence). Image acquisition was performed via device specific cameras/detector and corresponding software (Leica Application suite; Zeiss AxioVision; Nikon NIS-Elements). Further image analyses and quantifications were performed with ImageJ/Fiji (https://fiji.sc/). For figure arrangement CorelDraw Graphics Suite x7 software (Corel Corporation) was used.
Quantification of median fin fold width was performed with ImageJ/Fiji by length measurement of maximum intensity projections of TP63/Hoechst stained embryos 24 and 48 hpf. Each embryo was measured at three independent positions at the dorsal or ventral median fin fold. Median fin fold width was measured between the apical fin and somite borders. Quantification of epidermal cells in 24 hpf embryos was performed by automated counting of TP63 positive cells in confocal stacks (size z: 40; z-step: 2 µm) on the complete ventral and dorsal median fin folds. While in 48 hpf embryos three independent positions of the same size (ventral and dorsal median fin fold, tail tip; 8800 µm2) were investigated per sample. Used ImageJ plugins and commands: Stack/Z Project, Despeckle, Watersheed, Analyze particles (size 2–10 µm, circularity 0.5–1.0). A minimum of 12 samples for each condition was investigated.
Electron microscopy
Embryos or regenerates were washed with PBS and fixed (2.5% glutaraldehyde, 50 mM cacodylat pH 7.2, 50 mM KCl, and 2.5 mM MgCl2) overnight at 4 °C. Subsequently, embryos or tissues were washed five times with 50 mM cacodyl buffer (pH 7.2) and fixed for 4 h with 2% OsO4 in 50 mM cacodylat (pH 7.2) buffer. Embryos or tissues were subsequently stained with 2% uranylacetate overnight. After gradual dehydration with ethanol, they were transferred to propylenoxid and embedded in Epon (SERVA Electrophoresis GmbH, Heidelberg, Germany). Ultrathin sections were analyzed using an EM10 from Zeiss (Oberkochen, Germany).
qPCR experiments
For qPCR experiments RNA was extracted from pools of 12 embryos each (age 24 hpf; F3 fndc3awue1/+ and fndc3awue1/wue1 generation; control: AB). For each genotype three independent pools were generated and compared. For cDNA synthesis 1 µg RNA was transcribed into cDNA and further analyzed in a ViiA7 Real-Time PCR System (Thermo Fisher Scientific). Each group and primer sample was analyzed in triplicates on a single qPCR plate utilizing HOT FIREPol Eva Green Mix Plus (Solis BioDyne). Data analysis of was performed via QuantStudio Real-Time PCS Software v1.1 by ΔΔCt method. AB control samples were used as reference sample for relative comparison. Amplification of gapdh and eef1a1l1 was used as endogenous controls. Used primer pairs, amplicon sizes and targeted regions are noted in Table S1.
References
Yano, T. & Tamura, K. The making of differences between fins and limbs. J Anat 222, 100–113, https://doi.org/10.1111/j.1469-7580.2012.01491.x (2013).
Amaral, D. B. & Schneider, I. Fins into limbs: Recent insights from sarcopterygian fish. Genesis, https://doi.org/10.1002/dvg.23052 (2017).
Yano, T., Abe, G., Yokoyama, H., Kawakami, K. & Tamura, K. Mechanism of pectoral fin outgrowth in zebrafish development. Development 139, 2916–2925, https://doi.org/10.1242/dev.075572 (2012).
Masselink, W. et al. A somitic contribution to the apical ectodermal ridge is essential for fin formation. Nature 535, 542–546, https://doi.org/10.1038/nature18953 (2016).
van den Boogaart, J. G., Muller, M. & Osse, J. W. Structure and function of the median finfold in larval teleosts. The Journal of experimental biology 215, 2359–2368, https://doi.org/10.1242/jeb.065615 (2012).
Carney, T. J. et al. Genetic analysis of fin development in zebrafish identifies furin and hemicentin1 as potential novel fraser syndrome disease genes. Plos Genet 6, e1000907, https://doi.org/10.1371/journal.pgen.1000907 (2010).
van Eeden, F. J. et al. Genetic analysis of fin formation in the zebrafish, Danio rerio. Development 123, 255–262 (1996).
Dane, P. J. & Tucker, J. B. Modulation of epidermal cell shaping and extracellular matrix during caudal fin morphogenesis in the zebra fish Brachydanio rerio. J Embryol Exp Morphol 87, 145–161 (1985).
Nagendran, M. et al. Canonical Wnt signalling regulates epithelial patterning by modulating levels of laminins in zebrafish appendages. Development 142, 2080, https://doi.org/10.1242/dev.125849 (2015).
Parichy, D. M., Elizondo, M. R., Mills, M. G., Gordon, T. N. & Engeszer, R. E. Normal table of postembryonic zebrafish development: staging by externally visible anatomy of the living fish. Dev Dyn 238, 2975–3015, https://doi.org/10.1002/dvdy.22113 (2009).
Grandel, H. & Schulte-Merker, S. The development of the paired fins in the zebrafish (Danio rerio). Mech Dev 79, 99–120, https://doi.org/10.1016/s0925-4773(98)00176-2 (1998).
Zhang, J. et al. Loss of fish actinotrichia proteins and the fin-to-limb transition. Nature 466, 234–237, https://doi.org/10.1038/nature09137 (2010).
Montes, G. S., Becerra, J., Toledo, O. M., Gordilho, M. A. & Junqueira, L. C. Fine structure and histochemistry of the tail fin ray in teleosts. Histochemistry 75, 363–376, https://doi.org/10.1007/BF00496739 (1982).
Konig, D., Page, L., Chassot, B. & Jazwinska, A. Dynamics of actinotrichia regeneration in the adult zebrafish fin. Developmental biology 433, 416–432, https://doi.org/10.1016/j.ydbio.2017.07.024 (2018).
Phan, H. E., Northorp, M., Lalonde, R. L., Ngo, D. & Akimenko, M. A. Differential actinodin1 regulation in embryonic development and adult fin regeneration in Danio rerio. PLoS One 14, e0216370, https://doi.org/10.1371/journal.pone.0216370 (2019).
Duran, I., Mari-Beffa, M., Santamaria, J. A., Becerra, J. & Santos-Ruiz, L. Actinotrichia collagens and their role in fin formation. Developmental biology 354, 160–172, https://doi.org/10.1016/j.ydbio.2011.03.014 (2011).
Iovine, M. K. Conserved mechanisms regulate outgrowth in zebrafish fins. Nature chemical biology 3, 613–618, https://doi.org/10.1038/nchembio.2007.36 (2007).
Wehner, D. & Weidinger, G. Signaling networks organizing regenerative growth of the zebrafish fin. Trends in genetics: TIG 31, 336–343, https://doi.org/10.1016/j.tig.2015.03.012 (2015).
Mari-Beffa, M., Carmona, M. C. & Becerra, J. Elastoidin turn-over during tail fin regeneration in teleosts. A morphometric and radioautographic study. Anat Embryol (Berl) 180, 465–470 (1989).
Pfefferli, C. & Jazwinska, A. The art of fin regeneration in zebrafish. Regeneration (Oxf) 2, 72–83, https://doi.org/10.1002/reg2.33 (2015).
Webb, A. E. et al. Laminin alpha5 is essential for the formation of the zebrafish fins. Developmental biology 311, 369–382, https://doi.org/10.1016/j.ydbio.2007.08.034 (2007).
Carrouel, F. et al. HUGO (FNDC3A): a new gene overexpressed in human odontoblasts. Journal of dental research 87, 131–136 (2008).
Cheng, P. et al. Fibronectin mediates mesendodermal cell fate decisions. Development 140, 2587–2596, https://doi.org/10.1242/dev.089052 (2013).
Zhu, J. & Clark, R. A. Fibronectin at select sites binds multiple growth factors and enhances their activity: expansion of the collaborative ECM-GF paradigm. J Invest Dermatol 134, 895–901, https://doi.org/10.1038/jid.2013.484 (2014).
Obholz, K. L., Akopyan, A., Waymire, K. G. & MacGregor, G. R. FNDC3A is required for adhesion between spermatids and Sertoli cells. Developmental biology 298, 498–513, https://doi.org/10.1016/j.ydbio.2006.06.054 (2006).
Cao, J. et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 566, 496–502, https://doi.org/10.1038/s41586-019-0969-x (2019).
Hwang, W. Y. et al. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat Biotechnol 31, 227–229, https://doi.org/10.1038/nbt.2501 (2013).
Jao, L. E., Wente, S. R. & Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc Natl Acad Sci USA 110, 13904–13909, https://doi.org/10.1073/pnas.1308335110 (2013).
Johnson, S. L. & Weston, J. A. Temperature-sensitive mutations that cause stage-specific defects in Zebrafish fin regeneration. Genetics 141, 1583–1595 (1995).
Somero, G. N. Proteins and temperature. Annu Rev Physiol 57, 43–68, https://doi.org/10.1146/annurev.ph.57.030195.000355 (1995).
Gautier, P., Naranjo-Golborne, C., Taylor, M. S., Jackson, I. J. & Smyth, I. Expression of the fras1/frem gene family during zebrafish development and fin morphogenesis. Dev Dyn 237, 3295–3304, https://doi.org/10.1002/dvdy.21729 (2008).
Talbot, J. C. et al. fras1 shapes endodermal pouch 1 and stabilizes zebrafish pharyngeal skeletal development. Development 139, 2804–2813, https://doi.org/10.1242/dev.074906 (2012).
Feitosa, N. M. et al. Hemicentin 2 and Fibulin 1 are required for epidermal-dermal junction formation and fin mesenchymal cell migration during zebrafish development. Developmental biology 369, 235–248, https://doi.org/10.1016/j.ydbio.2012.06.023 (2012).
Jasuja, R. et al. bmp1 and mini fin are functionally redundant in regulating formation of the zebrafish dorsoventral axis. Mech Dev 123, 548–558, https://doi.org/10.1016/j.mod.2006.05.004 (2006).
Asharani, P. V. et al. Attenuated BMP1 function compromises osteogenesis, leading to bone fragility in humans and zebrafish. Am J Hum Genet 90, 661–674, https://doi.org/10.1016/j.ajhg.2012.02.026 (2012).
Zhang, H. Y., Lardelli, M. & Ekblom, P. Sequence of zebrafish fibulin-1 and its expression in developing heart and other embryonic organs. Dev Genes Evol 207, 340–351, https://doi.org/10.1007/s004270050121 (1997).
Eisenhoffer, G. T. et al. A toolbox to study epidermal cell types in zebrafish. J Cell Sci 130, 269–277, https://doi.org/10.1242/jcs.184341 (2017).
Lee, H. & Kimelman, D. A dominant-negative form of p63 is required for epidermal proliferation in zebrafish. Dev Cell 2, 607–616 (2002).
Santamaria, J. A. & Becerra, J. Tail fin regeneration in teleosts: cell-extracellular matrix interaction in blastemal differentiation. J Anat 176, 9–21 (1991).
Duran, I. et al. Collagen duplicate genes of bone and cartilage participate during regeneration of zebrafish fin skeleton. Gene Expr Patterns 19, 60–69, https://doi.org/10.1016/j.gep.2015.07.004 (2015).
Wehner, D. et al. Wnt/beta-catenin signaling defines organizing centers that orchestrate growth and differentiation of the regenerating zebrafish caudal fin. Cell reports 6, 467–481, https://doi.org/10.1016/j.celrep.2013.12.036 (2014).
Henderson, B., Nair, S., Pallas, J. & Williams, M. A. Fibronectin: a multidomain host adhesin targeted by bacterial fibronectin-binding proteins. FEMS Microbiol Rev 35, 147–200, https://doi.org/10.1111/j.1574-6976.2010.00243.x (2011).
Bakkers, J., Hild, M., Kramer, C., Furutani-Seiki, M. & Hammerschmidt, M. Zebrafish DeltaNp63 is a direct target of Bmp signaling and encodes a transcriptional repressor blocking neural specification in the ventral ectoderm. Dev Cell 2, 617–627 (2002).
Heude, E., Shaikho, S. & Ekker, M. The dlx5a/dlx6a genes play essential roles in the early development of zebrafish median fin and pectoral structures. PLoS One 9, e98505, https://doi.org/10.1371/journal.pone.0098505 (2014).
Syed, A. et al. Miles to go (mtgo) encodes FNDC3 proteins that interact with the chaperonin subunit CCT3 and are required for NMJ branching and growth in Drosophila. Developmental biology 445, 37–53, https://doi.org/10.1016/j.ydbio.2018.10.016 (2019).
Ricard-Blum, S. The collagen family. Cold Spring Harb Perspect Biol 3, a004978, https://doi.org/10.1101/cshperspect.a004978 (2011).
Bhadra, J. & Iovine, M. K. Hsp47 mediates Cx43-dependent skeletal growth and patterning in the regenerating fin. Mech Dev 138(Pt 3), 364–374, https://doi.org/10.1016/j.mod.2015.06.004 (2015).
Fisher, S., Jagadeeswaran, P. & Halpern, M. E. Radiographic analysis of zebrafish skeletal defects. Developmental biology 264, 64–76 (2003).
Huang, C. C. et al. Collagen IX is required for the integrity of collagen II fibrils and the regulation of vascular plexus formation in zebrafish caudal fins. Developmental biology 332, 360–370, https://doi.org/10.1016/j.ydbio.2009.06.003 (2009).
Akiyama, S. K. Integrins in cell adhesion and signaling. Hum Cell 9, 181–186 (1996).
Singh, P., Carraher, C. & Schwarzbauer, J. E. Assembly of fibronectin extracellular matrix. Annu Rev Cell Dev Biol 26, 397–419, https://doi.org/10.1146/annurev-cellbio-100109-104020 (2010).
Wada, N. Spatiotemporal changes in cell adhesiveness during vertebrate limb morphogenesis. Dev Dyn 240, 969–978, https://doi.org/10.1002/dvdy.22552 (2011).
Towers, M. & Tickle, C. Generation of pattern and form in the developing limb. Int J Dev Biol 53, 805–812, https://doi.org/10.1387/ijdb.072499mt (2009).
Mullins, M. C. et al. Genes establishing dorsoventral pattern formation in the zebrafish embryo: the ventral specifying genes. Development 123, 81–93 (1996).
Hild, M. et al. The smad5 mutation somitabun blocks Bmp2b signaling during early dorsoventral patterning of the zebrafish embryo. Development 126, 2149–2159 (1999).
Connors, S. A., Trout, J., Ekker, M. & Mullins, M. C. The role of tolloid/mini fin in dorsoventral pattern formation of the zebrafish embryo. Development 126, 3119–3130 (1999).
Blader, P., Rastegar, S., Fischer, N. & Strahle, U. Cleavage of the BMP-4 antagonist chordin by zebrafish tolloid. Science 278, 1937–1940, https://doi.org/10.1126/science.278.5345.1937 (1997).
Weiss, A. & Attisano, L. The TGFbeta superfamily signaling pathway. Wiley Interdiscip Rev Dev Biol 2, 47–63, https://doi.org/10.1002/wdev.86 (2013).
Nishizuka, M. et al. Disruption of the novel gene fad104 causes rapid postnatal death and attenuation of cell proliferation, adhesion, spreading and migration. Exp Cell Res 315, 809–819, https://doi.org/10.1016/j.yexcr.2008.12.013 (2009).
Kishimoto, K. et al. FAD104, a regulatory factor of adipogenesis, acts as a novel regulator of calvarial bone formation. J Biol Chem 288, 31772–31783, https://doi.org/10.1074/jbc.M113.452961 (2013).
Govindan, J. & Iovine, M. K. Dynamic remodeling of the extra cellular matrix during zebrafish fin regeneration. Gene Expr Patterns 19, 21–29, https://doi.org/10.1016/j.gep.2015.06.001 (2015).
Govindan, J., Tun, K. M. & Iovine, M. K. Cx43-Dependent Skeletal Phenotypes Are Mediated by Interactions between the Hapln1a-ECM and Sema3d during Fin Regeneration. PLoS One 11, e0148202, https://doi.org/10.1371/journal.pone.0148202 (2016).
Westerfield, M. The zebrafish book. A guide for the laboratory use of zebrafish (Danio rerio). Vol. 4th ed. (Univ. of Oregon Press, 2000).
Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B. & Schilling, T. F. Stages of embryonic development of the zebrafish. Dev Dyn 203, 253–310, https://doi.org/10.1002/aja.1002030302 (1995).
Hauptmann, G. & Gerster, T. Two-color whole-mount in situ hybridization to vertebrate and Drosophila embryos. Trends in genetics: TIG 10, 266 (1994).
Thisse, C. & Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nature protocols 3, 59–69, https://doi.org/10.1038/nprot.2007.514 (2008).
Inoue, D. & Wittbrodt, J. One for all–a highly efficient and versatile method for fluorescent immunostaining in fish embryos. PLoS One 6, e19713, https://doi.org/10.1371/journal.pone.0019713 (2011).
Walker, M. B. & Kimmel, C. B. A two-color acid-free cartilage and bone stain for zebrafish larvae. Biotechnic & histochemistry: official publication of the Biological Stain Commission 82, 23–28, https://doi.org/10.1080/10520290701333558 (2007).
Letunic, I. & Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res 46, D493–D496, https://doi.org/10.1093/nar/gkx922 (2018).
Acknowledgements
The authors thank Anja Kirschbauer for professional fish keeping support, Tabea Röder for in-situ probe synthesis, Manfred Schartl for sharing aquatic equipment, as well as Claudia Gehrig and Daniela Bunsen from the Electron Microscopy Core Unit of the Biocenter at the University of Würzburg, for preparation of ultrathin sections and support with the electron microscopy analysis. We are indebted to all our colleagues for fruitful scientific discussions. A prepublication version of the manuscript was made available to the scientific community on bioRxiv before publication in this journal. This publication was funded by the German Research Foundation (DFG) and the University of Würzburg in the funding program Open Access Publishing.
Author information
Authors and Affiliations
Contributions
D.L., S.G. and E.K. designed the experiments. D.L., I.K., S.K., M.M. and M.O. conducted the experiments. D.L. and E.K. wrote the manuscript with input from all authors.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liedtke, D., Orth, M., Meissler, M. et al. ECM alterations in Fndc3a (Fibronectin Domain Containing Protein 3A) deficient zebrafish cause temporal fin development and regeneration defects. Sci Rep 9, 13383 (2019). https://doi.org/10.1038/s41598-019-50055-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-50055-w
- Springer Nature Limited
This article is cited by
-
Genome-wide transcriptome analysis reveals equine embryonic stem cell-derived tenocytes resemble fetal, not adult tenocytes
Stem Cell Research & Therapy (2020)