
Abstract
The ubiquitous distribution of the classic endocannabinoid system (cannabinoid receptors CB1 and CB2) has been demonstrated within the monkey nervous system, including the retina. Transient receptor potential vanilloid type 1 (TRPV1) is a cannabinoid-like non-selective cation channel receptor that is present in the retina and binds to endovannilloids and endocannabinoids, like anandamide, 2-arachidonoylglycerol and N-arachidonoyl dopamine. Retinal expression patterns of TRPV1 are available for rodents and data in higher mammals like humans and monkeys are scarce. We therefore thoroughly examined the expression and localization of TRPV1 in the retina, at various eccentricities, of the vervet (Chlorocebus sabeus) monkey, using Western blots and immunohistochemistry. Our results demonstrate that TRPV1 is found mainly in the outer and inner plexiform layers, and in the retinal ganglion cell (RGC) layer with a higher density in the periphery. Co-immunolabeling of TRPV1 with parvalbumin, a primate horizontal cell marker, revealed a clear overlap of expression throughout the entire cell structure with most prominent staining in the cell body membrane and synaptic terminals. Furthermore, double labeling of TRPV1 and syntaxin was found throughout amacrine cells in the inner plexiform layer. Finally, double staining of TRPV1 and Brn3a allowed us to confirm its previously reported expression in the cell bodies and dendrites of RGCs. The presence of TRPV1 in the horizontal pathway suggests a function of this receptor in lateral inhibition between photoreceptors through the horizontal cells, and between bipolar cells through amacrine cells.
Similar content being viewed by others


Introduction
The transient receptor potential vanilloid type 1 (TRPV1) is a non-selective cation channel receptor that is expressed in numerous cell types throughout the entire body. TRPV1 is found in microglial cells in the brain and in the retina of several animal species1,2,3. In the retina, TRPV1 has been studied mostly in rats, rabbits and fishes, but in primates and humans, data are scarce. For example, in the goldfish retina, TRPV1 is expressed in three anatomically distinct amacrine cell types4 and, in rats, it is localized in the ganglion cell layer (GCL), the optic nerve head, and in the optic nerve itself5. Moreover, in the same species, TRPV1 is located in small bundles of RGC axons in the peripheral retina and is densely expressed in large axon bundles close to the optic nerve6. TRPV1 was localized in rabbit Müller cells7 but not in rats5. In genetically manipulated TRPV1Cre:Ai9 and TRPV1Cre:Ai3 mice8,9, TRPV1 was found in RGCs but not amacrine cells10. As for the primate retina, only one study has located TRPV1 receptor in the GCL, but the exact retinal locus was not reported, neither the complete specific retinal cell types expressing it. This receptor was also found in the RGC dendrites, with diffuse labeling in the soma. In the human retina, TRPV1 was slightly more diffuse in the RGC cytoplasm compared to the macaque11. Differences in the expression of TRPV1 are probably due to the lack of specificity of the antibodies12,13 and the interspecies’ differences.
In TRPV1 knockout mice (trpv1−/−) and with pharmacological blockade of TRPV1 in normal mice, an accelerated degeneration of RGCs occurs in a model of high intraocular pressure (IOP)14. Moreover, in trpv1−/− RGCs lacked compensatory transient increase in spontaneous action potentials and necessitated higher depolarization to reach firing threshold when exposed to high IOP11,15. It was also reported that in microglia, TRPV1 was involved in the modulation and regulation of Ca2+ activity. Indeed, TRPV1 activation was linked with a strong conductance of Ca2+ that is related to the apoptosis of neurons and glial cells2,6,16,17,18. Furthermore, endogenous cannabinoids like anandamide (AEA) and 2-arachidonoylglycerol (2-AG), ligands of TRPV1, might play a protective role against ischemic injury and excitotoxicity11,15,19,20. TRPV1 has also several modulatory functions through multimodal integration of chemical stimuli such as capsaicin and endocannabinoids21,22,23,24,25.
From all the reported studies, there is not a precise localization of TRPV1 in the various components of the retinal mosaic and there are also contradictory results that might be attributed to interspecies’ differences and specificity of the antibody used. Moreover, studies on monkeys are scarce and the complete profile of cells expressing TRPV1 is lacking. In this study, we have thoroughly mapped TRPV1 expression in all cell types of the entire retina (from the center to the periphery) of the vervet monkey using Western blots and immunohistochemistry. Our results demonstrated that TRPV1 was mainly expressed in the horizontal system of the retina (horizontal and amacrine cells) that adds to the previously reported data by our laboratory on the endocannabinoid receptor expression in the vertical retinal system (from photoreceptors to ganglion cells). We also propose a model regarding the putative role of TRPV1 in a two-stage lateral inhibition, one at the receptor level through horizontal cells and the other at the bipolar cells’ level through amacrine cells, that might contribute to perceptual contrast enhancement.
Materials and methods
Tissue preparation
The eyes were taken from 5 adult vervet monkeys (Chlorocebus sabeus; aged 3–4 years) and were part of ongoing research projects approved by the University of Montreal and the Behavioural Science Foundation Animal Care Committee. All methods were performed in accordance with the ARRIVE guidelines and the regulations of the Canadian Council on Animal Care. No animals were specially sacrificed for this study. The description of the in vivo animal preparation has been described in previous publications26,27,28,29, and will be described briefly here. The animals were deeply anaesthetized with a lethal dose of sodium pentobarbital (25 mg/kg, intravenous) and perfused through the heart with a solution of phosphate buffered saline (PBS; pH 7.4), followed by 4% paraformaldehyde (PFA) solution in PBS. The eyes were then removed and the anterior segment of the eye, namely the cornea, iris, ciliary body, and lens, as well as the vitreous humour were cut away. The eyecups were bathed in 4% PFA (prepared in 0.1 M PBS, pH 7.4) and left in the solution. The retina was dissected free from the eyecup in a PBS medium. It was laid flat, and by brushing gently with filter paper, the vitreous body was removed. Samples of the retina were taken at the center, middle, and periphery. Each sample was then cryoprotected in 30% sucrose overnight and embedded in Shandon embedding media at − 65 °C. The blocks were cut in 20 μm sections at -18 °C with a Leica CM3050S cryostat and mounted onto gelatinized subbed glass slides. After air drying, the slides were stored at − 80 °C until further processing.
Immunofluorescence
Single- and double-labeling of the retina were performed according to previously published methods26,27,28. Briefly, the sections were rinsed 3 × 5 min in 0.1 M Tris buffer, pH 7.4/0.03% Triton and blocked for 90 min in a blocking solution (10% normal donkey serum (NDS), 0.1 M Tris buffer/0.5% Triton). Sections were then incubated with primary antibodies prepared in blocking solution overnight at room temperature. The TRPV1 antibody (#NB100-1617, Novus Biologicals, Littleton, CO, USA) was also used conjointly with one of these known specific retinal cell type markers, namely protein kinase C (PKC), syntaxin, parvalbumin (PV), and Brain-specific homeobox/POU domain protein 3A (Brn3a) (see Table 1). The next day, sections were washed for 10 min and 2 × 5 min in 0.1 M Tris/0.03% Triton. They were then blocked in 10% NDS, 0.1 M Tris /0.5% Triton for 60 min and incubated with secondary antibodies for one hour (Alexa 488 donkey anti-mouse or Alexa 647 donkey anti-mouse and Alexa 647 donkey anti-rabbit or Alexa 488 donkey anti-rabbit at 1:200, all prepared in blocking solution). Sections were counterstained with Sytox Orange Nucleic Acid Stain (1:10,000; Molecular Probes, Inc., Eugene, OR, USA) to properly align retinal sections for confocal analysis, and washed again in Tris buffer, and coverslipped with Fluoromount-G™ Mounting Medium (SouthernBiotech, Birmingham, AL, USA).
Western blotting
Western blot analysis was performed according to previously published methods28. Briefly, frozen retinal tissue was homogenized by hand using a sterile pestle in a radioimmunoprecipitation assay buffer (150 mM NaCl, 20 mM Tris, pH 8.0, 1% NP-40, 0.1% sodium dodecyl sulfate (SDS), 1 mM EDTA), supplemented with protease inhibitors (Pierce Protease Inhibitor Tablets #88266, Thermo Fisher Scientific, Rockford, IL, USA) and phenylmethylsulfonyl fluoride (0.2 mg/ml; Roche Applied Science, Laval, QC, Canada). At 4 °C for 10 min, the samples were then centrifuged, and the supernatant was extracted and stored at − 20 °C until further processing. Protein content was equalized using a Thermo Scientific Pierce BCA Protein Assay Kit (Fischer Scientific, Ottawa, ON, Canada). Ten micrograms of protein homogenate per sample were loaded for electrophoresis in a 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel. Proteins were then transferred to a polyvinylidene difluoride (PVDF) membrane using the Trans-Blot Turbo Transfer System (Bio-Rad, Hercules, CA, USA). Upon recovering and washing the membrane in TBST (5 M NaCl, 1 M pH 8 Tris, 50% Tween-20) five times for five minutes each time, the membranes were blocked for one hour in 5% skim milk in TBST. Thereafter, the membrane was left incubating overnight with the rabbit anti-TRPV1 primary antibody in blocking solution in a 1:500 dilution at 4ºC. On the following day, the membrane was washed five times for five minutes in TBST before and after a 2-h incubation with horseradish peroxidase (HRP) conjugated donkey anti-rabbit secondary antibody in blocking solution in a 1:5,000 dilution at room temperature. Enhanced chemiluminescence (ECL) Clarity Western blot substrate was used for protein detection (Bio-Rad, Hercules, CA, USA). Proteins were visualized using the ChemiDoc Imaging Software (Bio-Rad, Hercules, CA, USA). For the control condition, the same protocol was ran simultaneously as described, excepting that the anti-TRPV1 primary antibody was preincubated with its blocking peptide (BP; #NB100-1617PEP, Novus Biologicals, Littleton, CO, USA) in a 1:10 dilution for 1 h.
Confocal microscopy
Immunofluorescence images were taken according to published methods28,29. Using a Leica TCS SP2 confocal laser-scanning microscope (Leica Microsystems, Exton, PA) or an Olympus FV3000 confocal laser-scanning microscope (Olympus Canada, Richmond Hill, ON, USA), images were obtained sequentially from the green, blue or far-red channels on optical slices of less than 0.9 μm of thickness. All photomicrograph adjustments, including size, color, brightness, and contrast were done with Adobe Photoshop (CC, Adobe Systems, San Jose, CA) equally for all images for each condition, and then exported to Adobe InDesign (CC, Adobe Systems, San Jose, CA), where the final figure layout was completed.
Optical density measurements
Confocal micrographs were first converted to an 8-bit grayscale image setting to visualize TRPV1 immunolabeling throughout the retinal layers. Using the public domain software Fiji (ImageJ, version 2.0.0, NIH Image, Bethesda, Maryland), mean gray values for all images to be quantified were measured. The gray spectrum values were generated from the pixel intensity (arbitrary value 0–255, 0 representing black and no signal, and 255 representing white and very strong signal).
Statistical analysis
A one-way analysis of variance (ANOVA) was conducted to evaluate the differences in the mean relative optical densities of TRPV1 through the six different layers of the retina (N = 4 measurements). For each layer, a one-way ANOVA was also performed to compare TRPV1 labeling intensity through three distinct retinal eccentricities. Scheffe’s test was used for all post-hoc comparisons.
Results
TRPV1 antibody specificity
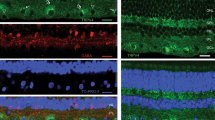
Western blot analysis demonstrates binding specificity of the TRPV1 antibody. A protein band is detected around 100 kDa, in accordance with the previously reported molecular weight for TRPV114. The signal is abolished when samples were incubated with its blocking peptide (Fig. 1A). We also tested for antibody specificity using immunohistochemistry. Retinal sections immunolabeled for TRPV1 showed expression in the outer plexiform (OPL), the inner plexiform (IPL), and ganglion cell layers (GCL) (Fig. 1B). Pre-incubation with the corresponding TRPV1 blocking peptide completely abolished antibody signal (Fig. 1C). The OPL and the GCL exhibited higher labeling intensity as compared to the photoreceptor (PRL) and the outer nuclear (ONL) layers (P < 0.05) (Fig. 1D).

Characterization of the TRPV1 antibody in the monkey retina. Western blot analysis of total protein samples from a vervet monkey retina, showing detection of the expected protein band at 100 kDa (A). The band was not detected when the antibody was pre-incubated with its corresponding TRPV1 blocking peptide (BP) at the ratio 1:10 (A). All lanes contained 10 μg of total protein. Immunohistochemistry on vervet retinal tissue with the antibody raised against TRPV1 revealed a unique staining profile (B). When the TRPV1 antibody was pre-incubated with its corresponding BP, it revealed an absence of staining in the vervet retinas (C). Quantitative analysis of the TRPV1 fluorescent signal through the six different retinal layers reveals a relatively higher signal intensity in the OPL and the GCL layers compared to the photoreceptor (PRL) and the outer nuclear (ONL) layers (P < 0.05) (D). TRPV1 transient receptor potential vanilloid type 1, BP blocking peptide, LC loading control, PRL photoreceptor layer, ONL outer nuclear layer, OPL outer plexiform layer, INL inner nuclear layer, IPL inner plexiform layer, GCL ganglion cell layer. Scale bar = 75 μm. Mean optical density ratios ± standard error of the mean (SEM) are depicted.
TRPV1 immunoreactivity throughout the vervet retina
TRPV1-immunoreactivity (IR) was present throughout the retina: center (Fig. 2A), middle (Fig. 2B), and periphery (Fig. 2C). A strong TRPV1 signal was localized in the lateral pathway, particularly in the IPL and OPL. Layer-specific differences were found on the relative distribution of TRPV1 through the three distinct retinal eccentricities, with the IPL and the GCL layers displaying higher signal intensity in the central retina, and the OPL and the ONL layers showing stronger peripherical TRPV1 labeling (Fig. 2D).

Labeling pattern of TRPV1-IR throughout the vervet retina. Confocal micrographs taken from the center (A), middle (B), and peripheral retina (C). Quantitative analysis of the TRPV1 fluorescent signal through the three distinct retinal eccentricities shows relatively higher central signal intensity in the IPL and the RGC layers, and greater peripherical labeling in the OPL and the ONL layers (P < 0.05) (D). PRL photoreceptor layer, ONL outer nuclear layer, OPL outer plexiform layer, INL inner nuclear layer, IPL inner plexiform layer, GCL ganglion cell layer, GCFL ganglion cell fiber layer. Scale bar = 75 µm. Mean optical density ratios ± standard error of the mean (SEM) are depicted.
Double-labeling immunohistochemistry
In order to verify the retinal cell type expression, double immunostaining was carried out for TRPV1 and a molecular marker for specific primate retinal cells. A consistent staining pattern across all five monkey retinas was found for each double staining. Although labeling was observed in all retinal layers, TRPV1-IR was most prominent in the horizontal pathway, namely horizontal and amacrine cells.
Distribution of TRPV1 in the various cells of the retina
TRPV1 is found in retinal ganglion cells
The widespread TRPV1-positive cell bodies in the ganglion cell layer suggests specific ganglion cell labeling (Fig. 3A–C). We verified this result with double-labeling of TRPV1-IR with Brn3a-IR, that characterizes ganglion cells in the retina of mammals, including our species (vervet monkey)26,28. Most cell bodies Brn3a-positive were also double-labeled for TRPV1 immunoreactivity. This included all the ganglion cell fibers. Most prominent staining appears to be in the central retina.

Double-label immunofluorescence illustrating immunolabeling of TRPV1 in ganglion cells, horizontal cells, and amacrine cells. (A-C) The antibody against Brn3a labels the nucleus of ganglion cells in the monkey retina, and these cells were strongly TRPV1 immunoreactive. Arrows indicate Brn3a-positive ganglion cells that are TRPV1 immunoreactive. Each protein expression is presented alone in grayscale: Brn3a (A) and TRPV1 (B); then, the two are presented merged (C), Brn3a in green and TRPV1 in red). (D-I) TRPV1 is expressed in horizontal cell synaptic terminals (arrows in OPL) and cell bodies (arrowheads in the INL). Each expression is presented alone in grayscale: PV (D,G), TRPV1 (E,H); then, the two are presented merged (F,J). (J–L) Arrows show the expression of TRPV1 in the amacrine cell bodies (INL). Each expression is presented alone in grayscale: syntaxin-positive amacrine cells (J) and TRPV1 (K); then, the two are presented merged (L). INL inner nuclear layer, IPL inner plexiform layer, OPL outer plexiform layer, GCL ganglion cell layer. Scale = 75 μm for (A–C) and (J–L), 30 μm for (D–F) and 10 μm for (G–I).
Horizontal cells express TRPV1
Since TRPV1-IR is co-localized with PV-immunoreactive cells, it indicates that horizontal cells are indeed TRPV1-positive. The PV-immunoreactivity is generally linked to similar staining of 2 specific classes of horizontal cells in the monkey retina (arrows), namely H1 and H2, as reported by Röhrenbeck et al. (1987). Obvious co-localization was observed in both horizontal cell types in the retina of vervet monkeys30 (Fig. 3D–I).
TRPV1 is localized in amacrine cells
To evaluate TRPV1-IR expression in amacrine cells, we used the monoclonal antibody HPC-1 that recognizes syntaxin in these cells. All amacrine cells showed expression of TRPV1-IR (Fig. 3J–L) even though variations in the intensity of immunolabeling were noticed.
TRPV1 is absent in rod bipolar cells
Cell fibers in the proximal INL that were TRPV1-immunoreactive looked like PKC-immunoreactive rod bipolar cell axons in the central retina. However, no PKC-IR rod bipolar cells, including their cell bodies, axons, and axon terminals, were co-localized with TRPV1 labeling, confirming that TRPV1 was absent in rod bipolar cells (Fig. 4A–C).

Double-label immunofluorescence showing no signal for TRPV1 in rod bipolar cells and Müller cells. TRPV1 is neither expressed in the cell bodies (small arrows in the upper INL) of bipolar cells nor their synaptic terminals at the edge of the OPL (arrowheads). Each protein expression is presented alone in grayscale: PKC (A) and TRPV1 (B); then, the two are presented merged (C), PKC in green and TRPV1 in red. Long arrows show specifically that the Müller cell bodies do not express TRPV1. Each protein expression is presented alone in grayscale: glutamine synthetase (D) and TRPV1 (E); then, the two are presented merged (F), glutamine synthetase in green and TRPV1 in red. Scale = 75 μm for (A–F).
TRPV1 is not expressed in the retinal glia (Müller cells)
In order to assess the expression of TRPV1 in Müller cells, we used double labeling of TRPV1 and glutamine synthetase (GS), a specific marker of the soma and processes of Müller cells31,32,33. Müller cells cover the entire retina and their extensions spread out into the GCL to form characteristic end-feet. GS-positive cell bodies and processes of Müller cells did not show TRPV1 immunoreactivity (Fig. 4D–F).
Discussion
Our anatomical results indicate the existence of TRPV1 in the horizontal pathway of the vervet monkey retina, extending further the results obtained by others14,34. Moreover, TRPV1 spans the whole vervet monkey retina, from its center to the periphery. While TRPV1 has been localized in a few retinal cells among different species14,34, this study is the first to show its precise expression in other retinal layers besides the GCL. Moreover, our results clarify the expression and localization of TRPV1 in the retina, mostly in the inhibitory cells of the lateral pathway. This localization extends further our understanding of the distribution of the endocannabinoid receptors in the monkey retina. The presence of TRPV1 in the horizontal pathway suggests its unique role in retinal function.
We compared TRPV1 labeling with that of the other cannabinoid receptors, CB1R, CB2R, and GPR55 previously described in our laboratory in the vervet monkey retina26,31,35. These four receptor types appear to be differentially expressed in the monkey retina. CB1R is observed in the central portion of the retina, from the photoreceptors (cones with heavy staining in the outer segments and pedicles, and rods with little staining confined into their spherules), the horizontal cells, the bipolar cells, the amacrine cells, and the ganglion cells. CB2R is mainly restricted to the retinal glia, the Müller cells. GPR55 is exclusively confined in rods, with a larger signal in its inner segments. As for TRPV1, it is mainly expressed in the horizontal pathway, the horizontal and amacrine cells, but also in ganglion cells. Figure 5 illustrates schematically these data for all retinal cell types.

Mapping of the receptors CB1R, CB2R, GPR55, and TRPV1 in the monkey retina. These receptors are differently expressed in the retina of monkeys. These results are compiled from several published articles26,31,35. CB1R is represented in green, CB2R in magenta, GPR55 in blue, and TRPV1 in red. OS, photoreceptors outer segments; IS, photoreceptors inner segments; ONL, outer nuclear layer; ONL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer.
Importantly, TRPV1 is mainly expressed in the horizontal pathway that contains the inhibitory GABAergic horizontal cells that links photoreceptors and amacrines that connect bipolar cells. The network of TRPV1 channels in horizontal cells, amacrine cells, and RGCs, is likely to be involved in the endocannabinoid and endovanilloid signaling in the retina, which are neuromodulation processes of retinal output4,21. The cannabinoid receptors in the vervet monkey retina have been ascribed specific roles in relation to their specific expression. CB1R and CB2R are involved in the modulation of both photopic and scotopic ERG responses36 and, GPR55 mediates exclusively scotopic ERG responses27. The presence of TRPV1 in inhibitory horizontal pathway might be responsible for modulating information entering the vertical pathway.
Hypothetical function of TRPV1 in the monkey retina
Horizontal cells (HC) are known to have a modulatory effect on the cone photoreceptors that they connect at the level of their pedicles37. It has been suggested that HC can exert their influence through the modulation of the release of the inhibitory neurotransmitter GABA38. We propose here another modulatory function of HC, this time, through the activation of the endocannabinoid system present in the photoreceptors (NAPE-PLD for AEA and DAGL for 2-AG). For example, the stimulation of a cone photoreceptor by a discrete small spot evokes the release of endocannabinoids, which in turn activates the TRPV1 receptors on the post-synaptic HC membrane. The Ca++ channels are then activated and there is a large calcium influx through the HC membrane. The influx of Ca++ depolarizes the HC and stimulates the release of inhibitory GABA in the synaptic cleft. The adjacent cone photoreceptors connected through the HC dendrites are therefore inhibited, which might result in the increased activation of the stimulated cone photoreceptor (Fig. 6). This mechanism involving the endocannabinoid system could be responsible for a first stage lateral inhibition process that influences the cone recipient bipolar cells, and a second stage inhibitory process that involves the amacrine cells (AC). AC connect the bipolar cell dendrites that synapse onto the retinal ganglion cells (RGC) somata. AC modulate the RGC input through a mechanism of action similar to HC since they also contain TRPV1 receptors, as well as GABA and glycine. The presence of the endocannabinoid system in the lateral retina suggests a two-stage inhibitory mechanism that fine-tunes the information processed by retinal ganglion cells.

Schematic illustration of the hypothetical mechanism of action of TRPV1 in horizontal cells of the monkey retina following stimulation of a cone. The different steps are highlighted by numbers. (1) When a cone is stimulated (C1), endocannabinoids (eCBs), particularly AEA and 2-AG, are produced in the synaptic cleft. (2) TRPV1 receptors are activated by eCBs in horizontal cells. (3) The calcium influx through the membrane causes depolarization of the horizontal cell. (4) GABA release in the synaptic cleft. (5) GABA binds to GABA receptors on the presynaptic neighboring cone pedicle (5). This mechanism of lateral inhibition could also potentially modulate the information transmitted to the bipolar cells and retinal ganglion cells. S spot stimulus, C1 stimulated cone 1, C2 adjacent cone 2, H horizontal cell, eCB endocannabinoid, GABAr Gamma-amino-butyric acid receptors, TRPV1 Transient receptor potential vanilloid type 1.
Data availability
The data generated and analyzed during the current study are available from the corresponding author on reasonable request.
References
Sappington, R. M. & Calkins, D. J. Contribution of TRPV1 to microglia-derived IL-6 and NFkappaB translocation with elevated hydrostatic pressure. Investig. Ophthalmol. Vis. Sci. 49, 3004–3017. https://doi.org/10.1167/iovs.07-1355 (2008).
Kim, S. R., Kim, S. U., Oh, U. & Jin, B. K. Transient receptor potential vanilloid subtype 1 mediates microglial cell death in vivo and in vitro via Ca2+-mediated mitochondrial damage and cytochrome c release. J. Immunol. 177, 4322–4329 (2006).
Katsura, H. et al. Activation of Src-family kinases in spinal microglia contributes to mechanical hypersensitivity after nerve injury. J. Neurosci. 26, 8680–8690. https://doi.org/10.1523/JNEUROSCI.1771-06.2006 (2006).
Zimov, S. & Yazulla, S. Vanilloid receptor 1 (TRPV1/VR1) co-localizes with fatty acid amide hydrolase (FAAH) in retinal amacrine cells. Vis. Neurosci. 24, 581–591. https://doi.org/10.1017/S095252380707054X (2007).
Leonelli, M., Martins, D. O., Kihara, A. H. & Britto, L. R. Ontogenetic expression of the vanilloid receptors TRPV1 and TRPV2 in the rat retina. Int. J. Dev. Neurosci. 27, 709–718. https://doi.org/10.1016/j.ijdevneu.2009.07.003 (2009).
Sappington, R. M., Sidorova, T., Long, D. J. & Calkins, D. J. TRPV1: contribution to retinal ganglion cell apoptosis and increased intracellular Ca2+ with exposure to hydrostatic pressure. Investig. Ophthalmol. Vis. Sci. 50, 717–728. https://doi.org/10.1167/iovs.08-2321 (2009).
Martínez-García, M. C. et al. Differential expression and localization of transient receptor potential vanilloid 1 in rabbit and human eyes. Histol. Histopathol. 28, 1507–1516. https://doi.org/10.14670/HH-28.1507 (2013).
Mishra, S. K. & Hoon, M. A. Ablation of TrpV1 neurons reveals their selective role in thermal pain sensation. Mol. Cell. Neurosci. 43, 157–163 (2010).
Jo, A. O. et al. Mouse retinal ganglion cell signalling is dynamically modulated through parallel anterograde activation of cannabinoid and vanilloid pathways. J. Physiol. 595, 6499–6516 (2017).
Lakk, M. et al. Polymodal TRPV1 and TRPV4 sensors colocalize but do not functionally interact in a subpopulation of mouse retinal ganglion cells. Front. Cell. Neurosci. 12, 353 (2018).
Sappington, R. M. et al. Activation of transient receptor potential vanilloid-1 (TRPV1) influences how retinal ganglion cell neurons respond to pressure-related stress. Channels (Austin) 9, 102–113. https://doi.org/10.1080/19336950.2015.1009272 (2015).
Gilliam, J. C. & Wensel, T. G. TRP channel gene expression in the mouse retina. Vis. Res. 51, 2440–2452 (2011).
Molnar, T. et al. Store-operated channels regulate intracellular calcium in mammalian rods. J Physiol. 590, 3465–3481 (2012).
Sappington, R. M. et al. Activation of transient receptor potential vanilloid-1 (TRPV1) influences how retinal ganglion cell neurons respond to pressure-related stress. Channels 9, 102–113 (2015).
Ward, N. J., Ho, K. W., Lambert, W. S., Weitlauf, C. & Calkins, D. J. Absence of transient receptor potential vanilloid-1 accelerates stress-induced axonopathy in the optic projection. J Neurosci 34, 3161–3170. https://doi.org/10.1523/JNEUROSCI.4089-13.2014 (2014).
Agopyan, N., Head, J., Yu, S. & Simon, S. A. TRPV1 receptors mediate particulate matter-induced apoptosis. Am. J. Physiol. Lung Cell Mol. Physiol. 286, L563-572. https://doi.org/10.1152/ajplung.00299.2003 (2004).
Reilly, C. A. et al. Calcium-dependent and independent mechanisms of capsaicin receptor (TRPV1)-mediated cytokine production and cell death in human bronchial epithelial cells. J. Biochem. Mol. Toxicol. 19, 266–275. https://doi.org/10.1002/jbt.20084 (2005).
Aarts, M. M. & Tymianski, M. TRPMs and neuronal cell death. Pflugers Arch. 451, 243–249. https://doi.org/10.1007/s00424-005-1439-x (2005).
Kim, S. R. et al. Roles of transient receptor potential vanilloid subtype 1 and cannabinoid type 1 receptors in the brain: neuroprotection versus neurotoxicity. Mol. Neurobiol. 35, 245–254 (2007).
Yazulla, S. Endocannabinoids in the retina: from marijuana to neuroprotection. Prog. Retin Eye Res. 27, 501–526. https://doi.org/10.1016/j.preteyeres.2008.07.002 (2008).
Ryskamp, D. A., Redmon, S., Jo, A. O. & Križaj, D. TRPV1 and endocannabinoids: emerging molecular signals that modulate mammalian vision. Cells 3, 914–938. https://doi.org/10.3390/cells3030914 (2014).
Caterina, M. J. et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389, 816–824. https://doi.org/10.1038/39807 (1997).
Van Der Stelt, M. & Di Marzo, V. Endovanilloids. Putative endogenous ligands of transient receptor potential vanilloid 1 channels. Eur. J. Biochem. 271, 1827–1834. https://doi.org/10.1111/j.1432-1033.2004.04081.x (2004).
McNamara, F. N., Randall, A. & Gunthorpe, M. J. Effects of piperine, the pungent component of black pepper, at the human vanilloid receptor (TRPV1). Br J Pharmacol 144, 781–790. https://doi.org/10.1038/sj.bjp.0706040 (2005).
Prager-Khoutorsky, M., Khoutorsky, A. & Bourque, C. W. Unique interweaved microtubule scaffold mediates osmosensory transduction via physical interaction with TRPV1. Neuron 83, 866–878. https://doi.org/10.1016/j.neuron.2014.07.023 (2014).
Bouskila, J. et al. Expression and localization of the cannabinoid receptor type 1 and the enzyme fatty acid amide hydrolase in the retina of vervet monkeys. Neuroscience 202, 117–130 (2012).
Bouskila, J. et al. Scotopic vision in the monkey is modulated by the G protein-coupled receptor 55. Vis. Neurosci. 33, E006 (2016).
Bouskila, J., Palmour, R. M., Bouchard, J.-F. & Ptito, M. Retinal structure and function in monkeys with fetal alcohol exposure. Exp. Eye Res. 177, 55–64 (2018).
Kucera, R. et al. Expression and localization of CB1R, NAPE-PLD, and FAAH in the vervet monkey nucleus accumbens. Sci. Rep. 8, 1–12 (2018).
Röhrenbeck, J., Wässle, H. & Heizmann, C. W. Immunocytochemical labelling of horizontal cells in mammalian retina using antibodies against calcium-binding proteins. Neurosci. Lett. 77, 255–260 (1987).
Bouskila, J., Javadi, P., Casanova, C., Ptito, M. & Bouchard, J. F. Müller cells express the cannabinoid CB2 receptor in the vervet monkey retina. J. Comp. Neurol. 521, 2399–2415 (2013).
Ikeda, T. et al. Immunohistological Study of Monkey Foveal Retina. Sci. Rep. 9, 1–17 (2019).
Nishikawa, S. & Tamai, M. Müller cells in the human foveal region. Curr. Eye Res. 22, 34–41 (2001).
Zimov, S. & Yazulla, S. Localization of vanilloid receptor 1 (TRPV1/VR1)-like immunoreactivity in goldfish and zebrafish retinas: restriction to photoreceptor synaptic ribbons. J. Neurocytol. 33, 441–452 (2004).
Bouskila, J., Javadi, P., Casanova, C., Ptito, M. & Bouchard, J.-F. Rod photoreceptors express GPR55 in the adult vervet monkey retina. PLoS ONE 8, e81080 (2013).
Bouskila, J. et al. Cannabinoid receptors CB1 and CB2 modulate the electroretinographic waves in vervet monkeys. Neural Plast. https://doi.org/10.1155/2016/1253245 (2016).
Masland, R. H. The fundamental plan of the retina. Nat. Neurosci. 4, 877–886 (2001).
Kramer, R. H. & Davenport, C. M. Lateral inhibition in the vertebrate retina: the case of the missing neurotransmitter. PLoS Biol. 13, e1002322 (2015).
Barnstable, C. J., Hofstein, R. & Akagawa, K. A marker of early amacrine cell development in rat retina. Dev. Brain Res. 20, 286–290 (1985).
Acknowledgements
The authors would like to thank the Behavioral Sciences Foundation Laboratories of St-Kitts in the West Indies, for supplying the vervet monkey retinal tissues. The Natural Science and Engineering Research Council of Canada (6362-2017: MP; RGPAS 478115-2015/RGPIN 2015-06582: JFB) and the Canadian Institutes of Health Research (163014-2019:MP and JFB, PJT-156029: JFB) supported this work. JB holds a postdoctoral fellowship from “Fonds de recherche du Québec—Santé (FRQS)” and was awarded the Yvonne & Andrew Koenig Scholarship.
Author information
Authors and Affiliations
Contributions
MP and JB conceptualized the study. JB and CMF performed the experiments and analyzed the data. JB wrote the main body of the text with CMF and MP. RP prepared and provided vervet monkey retinas. MP and JFB supervised and provided equipment and funding. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bouskila, J., Micaelo-Fernandes, C., Palmour, R.M. et al. Transient receptor potential vanilloid type 1 is expressed in the horizontal pathway of the vervet monkey retina. Sci Rep 10, 12116 (2020). https://doi.org/10.1038/s41598-020-68937-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-68937-9
- Springer Nature Limited