
Abstract
Herbivorous insects can escape the strong pressure of parasitoids by switching to feeding on new host plants. Parasitoids can adapt to this change but at the cost of changing their preferences and performance. For gregarious parasitoids, fitness changes are not always observable in the F1 generation but only in the F2 generation. Here, with the model species and gregarious parasitoid Anaphes flavipes, we examined fitness changes in the F1 generation under pressure from the simulation of host switching, and by a new two-generation approach, we determined the impact of these changes on fitness in the F2 generation. We showed that the parasitoid preference for host plants depends on hatched or oviposited learning in relation to the possibility of parasitoid decisions between different host plants. Interestingly, we showed that after simulation of parasitoids following host switching, in the new environment of a fictitious host plant, parasitoids reduced the fictitious host. At the same time, parasitoids also reduced fertility because in fictitious hosts, they are not able to complete larval development. However, from a two-generation approach, the distribution of parasitoid offspring into both native and fictitious hosts caused lower parasitoid clutch size in native hosts and higher individual offspring fertility in the F2 generation.
Similar content being viewed by others


Introduction
Natural enemies keep herbivores at densities lower than those that would deplete host plants1. Parasitoids are important natural enemies that maintain the natural balance of ecosystems2,3, with all insects in a given habitat potentially being hosts for parasitoids4. However, herbivorous insects, as the most vulnerable hosts4, are not passive participants; they evolve mechanical, physiological and immune defences against parasitoids2 and are in a constant evolutionary arms race. Regardless of the defensive mechanisms the host invents, parasitoids evolve to overcome them5,6.
In addition to direct defence, herbivorous insects can escape parasitoids by switching to a novel host plant and obtaining enemy free space7. Herbivores usually experience lower performance when colonizing a novel host plant8, but the associated fitness reduction may be compensated by lower predation and parasitism on this plant9,10. Although locating hosts in a new microhabitat is not easy for parasitoids, there are known cases of this switching11,12. These new habitats pose several challenges for parasitoids13, that might reduce their fitness, either directly (e.g. encounter of a plant toxin or metabolite by the juvenile stage) or indirectly (e.g. effect of decreased host size and quality on the parasitoid)14. In addition to possible changes in parasitoid fitness related to the new host plant of their host8,15, the new environment may present not only new predators of the parasitized hosts16 but also new potential host species for parasitoids17.
In this changing environment, it is critical whether the parasitoid’s reproductive strategy is solitary or gregarious: solitary parasitoids respond to these changes primarily by choosing the sex of the offspring developing in the host18, whereas gregarious parasitoids can choose both sex ratio and clutch size19,20. Female solitary parasitoids usually lay one egg or sometimes multiple eggs in a single host, but only one offspring completes development. In contrast, multiple offspring may complete development within a single host in gregarious parasitoids21,22. In parasitoids, the gregarious strategy has evolved numerous times (independently at least 43 times in 26 different families of Hymenoptera) from the ancestral solitary strategy23. The emergence of the gregarious strategy has been explained by “gregariousness based on non-fighting larval phenotypes”24, where loss of predatory larvae led to the origin of the gregarious strategy under very strict conditions25,26,27. Under this assumption, however, any interactions between solitary and gregarious larvae would result in a highly asymmetric result24, because when sharing the same host, tolerant gregarious larvae would be killed by predatory solitary larvae26. Thus, newer theoretical models demonstrate a much more likely evolution of gregarious strategy based on larval mobility, "gregariousness based on non-searching larval phenotypes", rather than on the lack of fighting larvae reviewed by24,25. For example, Mayhew & van Alphen28 show two closely related species with predatory larvae with solitary (Aphaereta genevensis Fischer, 1966) and gregarious (Aphaereta pallipes (Say, 1829)) strategies, where the results of superparasitization showed that the gregarious strategy is possible by differences in larval behaviour. Pexton & Mayhew29 report that there is a study of Rosenheim27 where the origin of a gregarious strategy depends on the larval tolerance, in the context of same-sex offspring developing in the same host, as female offspring are more closely related in parasitoids30. Thus, more than one change in larval phenotype is responsible for the emergence of a gregarious strategy from a solitary one: is the loss of aggression while retaining mobility and the other is the retention of aggression but with reduced mobility29.
It would seem that the solitary strategy is preferable as more parasitoid species adopt this reproductive strategy31. However, as mentioned above, the gregarious strategy gives an advantage to its holders in the form of multiple offspring that are able to complete development in a single host. This allows gregarious parasitoids to respond flexibly to changing environmental conditions, such as different host population densities20,32, host characteristics33, or presence of a predator of host20. For example, the population density of solitary parasitoids decreases with lower host densities, because only one individual develops in one host34, whereas gregarious parasitoids increase clutch size at lower host densities and thus maintain stable population density20. However, females of gregarious parasitoids must be able to choose the suitable clutch size not only according to environmental conditions35, but also according to whether their host does not accept food after parasitization (idiobiont) or whether it continues accepting food, increasing its the body size and the amount of resources for the parasitoid's offspring (koinobiont)36. In some cases of hemolymph-feeding koinobionts, such as species of genera Cotesia Cameron, 1891 and Microplitis Foerster, 1862, large part of the host remains intact after the parasitoid hatches37. These solitary parasitoids can evolve gregariously provided they have sufficient host resources to develop offspring without reducing their fitness36.
For both koinobionts and idiobionts, the change in clutch size, largely affect offspring body sizes, because the body size of the offspring in many cases depends on the amount of food obtained during larval development23,38 and the body size therefore decreases with higher clutch size39,40,41. At the same time, the offspring body size determines their future fertility in the F2 generation40,41,42. In general, the population density not only of parasitoids but also of other insects is determined by the number of individuals of a given species, i.e. it can be predicted from the fertility of females43,44. For gregarious parasitoids, a two-generation approach has been proposed, which provides more accurate results about parasitoid population dynamics by involving changes in fertility in the F1 and F2 generations due to different clutch sizes45.
In this study, using a two-generation approach, we examine the response of the gregarious parasitoid Anaphes flavipes (Förster, 1841) (Hymenoptera: Mymaridae) to the simulation of host switching to a fictitious host plant. Using a two-generation approach, we determined the consequences of changes in clutch size on A. flavipes fitness in the F2 generation. The idiobiont gregarious parasitoid A. flavipes is a suitable model for studying these questions because mated females parasitize the host in the egg stage, which represents the ultimate food supply for their offspring have a choice of 35 possible clutch size combinations (1 to 7 offspring laid in one host in any sex ratio)41,46. Using a two-generation approach, some combinations of clutch size are more advantageous than others47,48 because they ensure higher fertility in the F2 generation45. In addition, knowledge of the factors that affect the fertility and thus the population dynamics of A. flavipes will be helpful in the use of this wasp in biological control against economically important pests of the genus Oulema49,50.
Result
No-choice test of host plants
The offspring sex ratio was not affected by the substrate on which host eggs were offered for parasitization (usual host plant versus filter paper: GLMM-b, χ2(1) = 0.017, p = 0.896; fictitious host plant versus filter paper: GLMM-b, χ2(1) = 0.618, p = 0.432; fictitious host plant versus usual host plant GLMM-b, χ2(1) = 0.954, p = 0.329). The proportion of parasitized hosts was higher on filter paper than on usual host plants (GLM-b, χ2(1) = 13.457, p < 0.001) and fictitious host plants (GLM-b, χ2(1) = 8.752, p = 0.003). The proportion of parasitized hosts between usual and fictitious host plants was not statistically significant (GLM-b, χ2(1) = 0.515, p = 0.473). The clutch size was higher on the filter paper than on the fictitious host plant (GLMM-p, χ2(1) = 3.317, p = 0.069). The clutch size between usual host plants and fictitious host plants (GLMM-p, χ2(1) = 1.334, p = 0.248) or filter paper (GLMM-p, χ2(1) = 0.126, p = 0.723) was not statistically significant. Overall, wasps laid more offspring on filter paper than on usual (LM, F(1,38) = 5.048, p = 0.031) and fictitious (LM, F(1,38) = 6.293, p = 0.017) plants. The difference between usual and fictitious plants was not significant (LM, F(1,38) = 0.081, p = 0.777) (Suppl. Mat. 1).
Choice test of host plants
The host plant in choice tests affects the reproductive characteristics of A. flavipes. The offspring sex ratio was not affected by the host plant (usual host plant versus filter paper: GLMM-b, χ2(1) = 1.454, p = 0.228; fictitious host plant versus filter paper: GLMM-b, χ2(1) = 3.611, p = 0.057; fictitious host plant versus usual host plant: GLMM-b, χ2(1) = 0.795, p = 0.373). The proportion of parasitized hosts was higher on the usual host plant than on the fictitious host plant (GLM-b, χ2(1) = 11.397, p = 0.001) and filter paper (GLM-b, χ2(1) = 4.416, p = 0.036). The proportion of parasitized hosts between fictitious host plants and filter paper was not statistically significant (GLM-b, χ2(1) = 1.452, p = 0.228). The clutch size was not statistically significant for usual host plants compared to fictitious host plants (GLMM-p, χ2(1) = 0.103, p = 0.748) or filter paper (GLMM-p, χ2(1) = 0.29, p = 0.591) or for fictitious host plants compared to filter paper (GLMM-p, χ2(1) = 0.028, p = 0.867) (Suppl. Mat. 2).
Effect of host plant choice tests versus no-choice tests
The type of test, “no-choice” and “choice”, affected the reproductive characteristics of A. flavipes. The total number of offspring by one female (LM, F(1,78) = 25.783, p < 0.001; Fig. 1a), the proportion of parasitized hosts (GLM-b, χ2(1) = 25.519, p < 0.001; Fig. 1b) and clutch size (GLM-p, χ2(1) = 19.002, p < 0.001; Fig. 1c) were higher if the wasps were hosted on one type of host plant (no-choice test) (Suppl. Mat. 1, 2). The offspring sex ratio was not affected between different types of tests (GLMM-b, χ2(1) = 1.427, p = 0.232; Fig. 1d).

Effect of host egg substrate (mixed, usual host plant (crop), fictitious host plant (broad-leaved dock), control substrate (filter paper)) on reproductive strategy of A. flavipes: (a) offspring sex ratio (grey: females, blank: males), (b) parasitized host egg rate (grey: parasitized), (c) clutch size (i.e. offspring per parasitized host egg), and (d) total number of offspring per female. Mean values and 95% confidence interval.
Effect of fictitious host
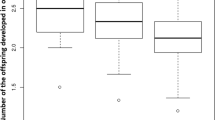
The offspring sex ratio (GLMM-b, F(1,25) = 0.448, p = 0.503) and the proportion of parasitized hosts (GLM-b, χ2(1) = 1.282, p = 0.258) were not statistically significant between wasps with 6 fictitous and 6 native hosts and wasps with 6 native hosts offered for parasitization, but fertility (LM, df = 25, χ2(1) = 18.676, p < 0.001; Fig. 2a) and clutch size (LMM, χ2(1) = 16.363, p < 0.001; Fig. 2b) were higher for wasps with 6 native hosts offered for parasitization. When comparing wasps with 6 native and 6 fictitous host eggs with wasps with 12 host eggs offered for parasitization, the fertility (LM, F(1,37) = 40.526, p < 0.001; Fig. 2a) was higher for wasps with 12 native host eggs offered for parasitization. The differences in offspring sex ratio (GLMM-b, χ2(1) = 1.573, p = 0.21), the proportion of parasitized hosts (GLM-b, χ2(1) = 0.427, p = 0.513) and clutch size (GLMM-p, χ2(1) = 1.53, p = 0.216; Fig. 2b) were not statistically significant. The rate of emergence for fictitous hosts with parasitoids was 22%, and that for control fictitous hosts without parasitoids was 94% (Suppl. Mat. 3).

(a) Clutch size (i.e. offspring per parasitized host egg) and (b) total number of offspring per female A. flavipes under different conditions: fictitious host plant with 50% rate of an unsuitable host (broad-leaved dock, 6 eggs of Oulema sp., 6 eggs of Gastrophysa viridula); usual host plant, low host density (crop, 6 eggs of Oulema sp.); usual host plant, high host density (crop, 12 eggs of Oulema sp.). Mean values and 95% confidence interval.
Discussion
In general, parasitoids show remarkable differences in their reproductive strategies, with their success depending on many factors such as ovigeny, fecundity, life span and many other51. In this study, we focused on one of the factors that affects the successful reproduction of parasitoids52,53, namely the localization of a suitable host in relation to a distinct host plant. A considerable number of studies show that parasitoids are influenced by plant defenses, either by changes in plant volatiles that hosts can use to defend themselves against parasitation or, conversely, these toxins degrade host quality for parasitoids54,55,56. However, there is a need to examine the effect of plants, herbivores and parasitoids in broader (“tritrophic”) context, and to uncover many direct and indirect interactions that will vary in strength and significance57. Thus, here, we presented the effect of host plants and the effect of a fictitious host that the parasitoid may encounter in the new environment of the fictitious plant on the fitness of parasitoids from a novel two-generation approach45. Using the model species A. flavipes, in Fig. 3, we show the hypothetical importance of this approach for studying parasitoid fitness. For example, in a situation where founders would refuse the host on the fictitious host plant, the changes in fertility in relation to a different type of host plant may not be observed in the F1 generation but will appear in the F2 generation.

A hypothetical scenario showing the importance of a two-generation approach; the females have host eggs for parasitation offered on the usual host plant (crop), fictitious host plant (Rumex) and control substrate (filter paper). (a) the female, which accepts the all host eggs offered on the different host plans (real scenario (Suppl. Mat. 4)) and (b) the female, which accepts the only host eggs offered on the usual host plant may have the same fertility in F1 generation, but using different clutch size, they obtain different fertility in F2 generation (fertility was determined according to Samková et al.45). (*The fertility values (shown in parentheses); The offspring sex ratio of A. flavipes is 3:1 (male:female)46; females are shown as  (left oriented) and males as
(left oriented) and males as  (right oriented)).
(right oriented)).
Recognition of the host plant by the parasitoid is fixed before adult emergence58,59, immediately following emergence60, or during oviposition61. In our no-choice tests, females of A. flavipes parasitized more host eggs on the substrate on which they hatched (control substrate; Fig. 4c) than on the usual and fictitious host plant (Fig. 4a,b), which would indicate that females learn about the host plant before or shortly after hatching. This finding may also be related to the fact that A. flavipes females hatch with a final number of own eggs (pro-ovigeny type)62 that can be laid immediately after hatching46. Based on the fact that synovigenic species are longer lived than proovigenic species63, it can be assumed that synovigenic species have a longer time to recognize the host plant. However, the influence of the ovaries in relation to host plant recognition is debatable because the imprinting of the host plant after the emergence of the parasitoids and, in relation to this, their higher fitness have been confirmed in quite a number of studies64,65 and these studies included both synovigenic (Trichogramma brassicae Bezdenko, 1968)63 and proovigenic (Cotesia Cameron 1891)63 parasitoid species.

The relationship between different fitness characteristics in F1 generation and fertility in F2 generation for four groups of females with different type of substrate on which the host eggs were offered; the 12 host eggs were offered on the (1) Non-choice test: (a) usual host plant (crop); (b) fictitious host plant (Rumex); (c) control substrate (filter paper) and (2) Choice test: (d) combination on all (usual, fictitious host plant and control substrate). (*The fertility values (shown in parentheses); To simplify the model the male offspring are not shown).
Interestingly, if females had a choice between the usual host plant, fictitious host plant and the control substrate, they chose to parasitize more host eggs on the usual host plant than on the learned substrate during emergence (control substrate) (Fig. 4d). This would indicate that the females can recognize the usual host plant if they have a choice between substrates, i.e., learning during oviposition61.
When comparing the results of the choice test and the no-choice test, females with a choice between substrates on which host eggs are offered for parasitization always have significantly lower values of fitness characteristics—the number of parasitized hosts, the number of offspring and the clutch size (Fig. 4). Although many factors are involved in fitness in the broader sense, one of the main indicators is still undoubtedly the number of offspring, especially when fitness is measured by maternal fertility according to the traditional approach43,44. However, the two-generation approach involves a change in fertility between the F1 and F2 generations caused by different clutch sizes45. In this study, in the choice test (Fig. 4d), the females had fewer offspring than females in the no-choice test (Fig. 4a–c) but also a smaller clutch size, thus ensuring a larger offspring body size and higher fertility, which increased by approximately 13 percent in the F2 generation (Fig. 4d). Other changes in the fertility of the F2 generation were not obvious (Fig. 4); however, for interest, the seemingly lower fertility in the F1 generation in relation to the fictitious host plant may ensure higher fertility in the F2 generation (Fig. 4b compared to 4a).
Interpretation of our results in the natural environment will be difficult because in nature, the offspring of switched hosts can use toxins from new host plants for their own protection66,67. In our experiments, the native host consumed only their native host plants; however, interestingly, with the simulation of switched hosts, the wasps with choice between the host plants had a lower fitness. It can be assumed that lower parasitoid fitness can be compensated by possible lower predation of fictitious hosts68. The switched host usually escapes the strong pressure of predators and parasitoids on their native plant9,10. Although under certain conditions, the predator avoids parasitized prey69, generally the parasitoid fitness is reduced more than predator fitness by intraguild predation70,71. Thus, for parasitoids, specialization on switched hosts should be stronger and more advantageous than for predators12.
Following host switching in a new environment, parasitoids can adapt to new potential host species4. In the second part of this study, we simulate the impact of a fictitious host on the fitness of parasitoids when they follow a switched host into a new environment. In the new environment of the fictitious host plant, the A. flavipes females parasitize the native host as well as the fictitious host. By parasitism, they reduce the fertility of the native and fictitious hosts and, concurrently, partly their own fertility because they were not able to complete larval development in the fictitious host (same as in our previous study Samková et al.45). Here, we show that the fertility of switched parasitoids is lower (females have 6 native and 6 fictitious hosts on the fictitious host plant; Fig. 5c) than the fertility of parasitoids on the usual host plant, which has high (12 native hosts; Fig. 5a) or low (6 native hosts; Fig. 5b) host population density. However, from a two-generation approach, if switched parasitoid females distribute their offspring between both the native and fictitious hosts, the clutch size decreases, and the offspring in the native host obtain a larger body size and higher fertility in the F2 generation (Fig. 5; similarly as in Samková et al.45). It can be assumed that in the new environment of a new host plant, among other factors16,55, parasitoid fitness will be reduced by parasitism of fictitious hosts in which the parasitoid is unable to complete larval development, but using this ovipositing behaviour, gregarious parasitoids increase the fertility of offspring developed in native hosts (Fig. 5; Samková et al.45). In addition to higher fertility, larger female offspring also obtain more females in the F3 generation offspring compared to smaller females72, and it can also be assumed that there is higher flight efficiency40,73, longevity74 or longer egg-laying time75. Additionally, by parasitizing the fictitious host, parasitoids gain the opportunity for specialization on new potential host species.

The relationship between different fitness characteristics in F1 generation and fertility in F2 generation for three groups of females in relation to occurrence of fictitious host in new environment of fictitious host plant; (a) females with 12 available native host on their native host plant (crop); (b) females with 6 available native host on their native host plant (crop) and (b) females with 6 available native host and 6 available fictitious host on the fictitious host plant (Rumex). (*The fertility values (shown in parentheses); The offspring sex ratio of A. flavipes is 3:1 (male:female)46; females are shown as  (left oriented) and males as
(left oriented) and males as  (right oriented).
(right oriented).
Material and methods
Parasitic wasps
Anaphes flavipes was reared from host eggs (Oulema spp.) collected in cereal fields in Prague (50.136° N, 14.363° E) from the end of April until the end of June in 2019. The parasitized host eggs were stored in Petri dishes with only moistened filter papers without host plants until adult wasps emerged. These “wild” wasps were used as an initial population from which the next generations of parasitoids were reared in an environmental chamber at 22 ± 2 °C with 40–60% relative humidity and continuous illumination. Subsequent generations of females and males were used for experiments. All wasps used in the experiment were naive to the host plant, that is, they were kept in Petri dishes without a host plant only on filter paper. Mated females (not older than 24 h postemergence) were placed in Petri dishes with host eggs. The females were not fed before the start of the experiment or during the experiment, and they had free access to water (modified from Samkova et al.20,41,45,72).
Host species
The fictitious host species Gastrophysa viridula (DeGeer, 1775) (Coleoptera: Chrysomelidae) and the habitual/common hosts of the Oulema species complex (including two very ecologically close species, O. duftschmidi (Redtenbacher, 1874) and O. melanopus (Linnaeus, 1758) (Coleoptera: Chrysomelidae)) were used; these species were used identically as in our previous studies e.g.20,41 and in other studies46,76 because they were determined only on the basis of genital preparation77. In the current study, the host culture was established from adults collected in Prague (50.136° N, 14.363° E) and in Police n/Met (50.527° N, 16.245° E). The adults were kept in plastic boxes with moistened filter papers, were fed of their native host plant (G. viridula were fed by R. obtusifolius and Oulema spp. were fed by T. aestivum), and had unlimited access to water. They were allowed to lay their eggs on leaves at 22 ± 2 °C, a relative humidity of 40–60% and a 16:8-h L:D cycle. We used host eggs no older than 24 h (modified from Samkova et al.20,41,45,72).
Host plant
The substrates on which host eggs were offered for parasitization were as follows: the usual host plant was Triticum aestivum L. (Poaceae), and Rumex obtusifolius L. (Polygonaceae) was used as the fictitious host plant. The usual host plant is a typical plant on which the host Oulema spp. occurs naturally in Europe and North America77, and the parasitoid Anaphes flavipes also occurs in this environment46. In the Czech Republic, R. obtusifolius, a plant host of G. viridula, which is ecologically and phylogenetically close to Oulema spp. is found near the land of this native plant(pers. obs.), so this plant was chosen as a fictitious plant because there is a possibility that the parasitoid of A. flavipes will encounter this plant. The host plants were obtained at localities in Prague (50.1367° N, 14.3638° E), and they were used within three hours of collection; prior to their use, they were stored in water. The host eggs were offered to wasps for parasitization on one (no-choice experiment) or two pieces (choice experiment) of the same leaf size. As a control, the host eggs were laid loosely on filter paper.
List of terms used in the experiments:
1. Native host = Oulema species; native host for parasitoids A. flavipes.
2. Fictitious host = Gastrophysa viridula; non-native host for A. flavipes.
3. Usual host plant = Triticum aestivum; native host plants; it is native host plant for Oulema species respectively for native host of A. flavipes.
4. Fictitious host plant = Rumex obtusifolius = non-native host plants; it is native host plants for G. viridula respectively for non-native host of A. flavipes.
Laboratory experiments
All laboratory experiments were performed in Petri dishes (8.5 cm) in a thermal cabinet at 22 ± 2 °C and 40–60% relative humidity under a 16:8 h (L:D) photoperiod. Individual parasitized host eggs were moved to 1.5-ml plastic tubes on the 9th or 10th day after parasitization and stored at the same temperature in a thermal cabinet. The number and sex ratio of the wasps that emerged from each parasitized host egg were measured (modified from Samkova et al.20,41,45,72).
Experimental design
-
1.
No-choice test of host plants
Twelve host eggs of Oulema sp. were offered to each of the 60 female wasps for 8 h in three groups: the host eggs were offered (1) on the usual host plant–crop (T. aestivum) (n = 20); (2) on the fictitious host plant (R. obtusifolius) (n = 20) and (3) on filter paper (n = 20).
-
2.
Choice test of host plants
Each female (n = 20) had 12 host eggs available for parasitization for 8 h on the three different substrates in the same Petri dish (4 host eggs on the usual host plant–crop (T. aestivum), 4 host eggs on the fictitious host plant (R. obtusifolius) and 4 host eggs on filter paper).
-
3.
Effect of fictitious host
Each female had host eggs offered for parasitization in three groups: (1) simulation of switched host to the fictitious host plant (6 native and 6 fictitious host eggs on the fictitious host plant; n = 15); (2) simulation of native environment with a low population density of host (6 native host eggs on the usual host plant; n = 11); and (3) simulation of native environment with a high population density of host (12 native host eggs on the usual host plant; n = 19). For fictitious hosts, we measured the rate of emergence in (1) experiments with native and fictitious hosts (n = 114), and for controls, we measured the rate of emergence for fictitious hosts without parasitoids (n = 100).
Simulation of novel two-generation reproductive approach
In our previous study, we have shown that with higher clutch size the body size of the A. flavipes offspring decreases and at the same time, that the offspring body size determines their fertility41. Therefore, in the study by Samková et al.45 we proposed a two-generation model to see intergenerational changes in fertility when clutch size changes in the F1 generation. In this study, we applied this model to the groups of experiments where there was a change in clutch size in the F1 generation, respectively a change in offspring body size—Figs. 3, 4 and 5. In these figures, the distribution of offspring in host eggs (number and sex ratio) was determined using the frequencies of each clutch size in real data (Suppl. Mat. 4). Fertility of hypothetical group in Fig. 3 was extrapolared by formula “fertility = 20—sqrt(clutch size)“, which fitted means of all measured fertility levels (rounded to the nearest integer).
The sex ratio of offspring did not differ significantly between groups (see Results), hence the 3:1 ratio used by Anderson & Paschke46 in the figures (Figs. 3, 4 and 5).
Statistical data processing
All statistical analyses were performed in R 4.0.3 (R foundation for statistical computing 202078). The models used were specific for each dependent variable. For the number of offspring per dish, we used linear models (LM), and for the rate of parasitized eggs, we used generalized linear models for binomial distribution (GLM-b). The offspring sex ratio was analysed by means of mixed-effect generalized linear models for binomial distribution (GLMM-b), and clutch size was analysed either by mixed effect linear models (LMM) or by mixed-effect generalized linear models for Poisson distribution (GLMM-p, data transformation: x-1) depending on the distribution of a particular dataset. An ID specific for each dish was used as a random factor in all mixed-effect models. The mixed-effect models were built in the R package lme479 and analysed by their comparison with simpler models (see below) by means of ANOVA.
Basic models always included a single fixed factor. The factors were egg substrate (Triticum/Rumex/control filter paper), substrates per dish (single substrate/mixed), or presence of leaf bug eggs (yes/no). Basic mixed-effect models were compared with null models that included the random factor only. Models assessing interactive effects between factors (e.g., different effects of substates in single-substrate and mixed dishes) always included two factors and their interaction. Mixed-effect models with interactions were compared with a model including the same two factors but not the interaction.
We calculated 95% confidence intervals for normally distributed data in R 4.0.3 (R foundation for statistical computing 202078). For binomial and Poisson-distributed data, 95% confidence intervals were calculated using the R packages Hmisc80 and DescTools81, respectively.
Ethical approval
The use of plants in the present study complies with international, national and/or institutional guidelines. The hosts plants were obtained at the same localities as hosts and parasitoids species localities in the Czech Republic. The plant samples used in the experiment are stored as dry material at the Department of Plant Protection, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Kamýcká 129, CZ-165 00, Prague 6 – Suchdol, Czech Republic: Triticum aestivum L. (Poaceae) and Rumex obtusifolius L. (Polygonaceae), CZ, Praha—Suchdol, hand collection, 1.v.2019–30.vi.2019, A. Samkova lgt., 50.1367° N, 14.3638° E, A. Samkova det. These same plant species (determined by A. Samkova) from the same host and parasitoid locations (50.136° N, 14.363° E; 50.527° N, 16.245° E) were also used for host feeding.
References
Hairston, N. G., Smith, F. E. & Lawrence, B. S. Community structure, population control, and competition. Am. Nat. 94, 421–425 (1960).
Gross, P. Insect behavioral and morphological defenses against parasitoid. Annu. Rev. Entomol. 38, 251–273 (1993).
Tylikinais, J. M., Tscharntke, T. & Klein, A. M. Diversity, ecosystem function and stability of parasitoid—host interactions across a tropical habitat gradient. Ecology 87, 3047–3057 (2006).
Strand, M. R. & Obrycki, J. J. Host specificity of insect parasitoids and predators. Bioscience 46, 422–429 (1996).
Dawkins, R. & Krebs, J. R. Arms races between and within species. Proc. R. Soc. Lond. B. 205, 489–511 (1979).
Kraaijeveld, A. R., van Alphen, J. J. M. & Godfray, H. C. J. The coevolution of host resistance and parasitoid virulence. Parasitology 116, 29–45 (1998).
Jeffries, M. J. & Lawton, J. H. Enemy free space and the structure of ecological communities. Biol. J. Linn. Soc. 23, 269–286 (1984).
Grosman, A. H. et al. No adaptation of a herbivore to a novel host but loss of adaptation to its native host. Sci. Rep.-UK 5, 16211 (2015).
Diamond, S. E. & Kingsolver, J. G. Fitness consequences of host plant choice: A field experiment. Oikos 119, 542–550 (2010).
Meijer, K., Schilthuizen, M., Beukeboom, L. & Smit, C. A review and meta-analysis of the enemy release hypothesis in plant–herbivorous insect systems. PeerJ 4, e2778 (2016).
Forbes, A. A., Powell, T. H., Stelinski, L. L., Smith, J. J. & Feder, J. L. Sequential sympatric speciation across trophic levels. Science 323, 776–779 (2009).
Grosman, A. H., Holtz, A. M., Pallini, A., Sabelis, M. W. & Janssen, A. Parasitoids follow herbivorous insects to a novel host plant, generalist predators less so. Entomol. Exp. Appl. 162, 261–271 (2017).
Soler, R., Bezemer, T. M., Van Der Putten, W. H., Vet, L. E. & Harvey, J. A. Root herbivore effects on above-ground herbivore, parasitoid and hyperparasitoid performance via changes in plant quality. J. Anim. Ecol. 74, 1121–1130 (2005).
Ode, P. J. Plant chemistry and natural enemy fitness: Effects on herbivore and natural enemy interactions. Annu. Rev. Entomol. 51, 163–185 (2006).
Thompson, J. N. Trade-offs in larval performance on normal and novel hosts. Entomol. Exp. Appl. 80, 133–139 (1996).
Lucas, É., Coderre, D. & Brodeur, J. Intraguild predation among aphid predators: Characterization and influence of extraguild prey density. Ecology 79, 1084–1092 (1998).
Henry, L. M., May, N., Acheampong, S., Gillespie, D. R. & Roitberg, B. D. Host-adapted parasitoids in biological control: Does source matter?. Ecol. Appl. 20, 242–250 (2010).
Mackauer, M. Sexual size dimorphism in solitary parasitoid wasps: influence of host quality. Oikos 76, 265–272 (1996).
Bezemer, T. M. & Mills, N. J. Clutch size decisions of a gregarious parasitoid under laboratory and feld conditions. Anim. Behav. 66, 1119–1128 (2003).
Samková, A., Hadrava, J., Skuhrovec, J. & Janšta, P. Host population density and presence of predators as key factors influencing the number of gregarious parasitoid Anaphes flavipes offspring. Sci. Rep.-UK 9, 6081 (2019).
Schmidt, J. M. & Smith, J. J. B. Correlations between body angles and substrate curvature in the parasitoid wasp Trichogramma minutum: a possible mechanism of host radius measurement. J. Exp. Biol. 125, 271–285 (1986).
Boivin, G. & Baaren, J. The role of larval aggression and mobility in the transition between solitary and gregarious development in parasitoid wasps. Ecol. Lett. 3, 469–474 (2000).
Mayhew, P. J. The evolution of gregariousness in parasitoid wasps. P. Roy. Soc. Lond. B. Bio. 265, 383–389 (1998).
Pexton, J. J. & Mayhew, P. J. Competitive interactions between parasitoid larvae and the evolution of gregarious development. Oecologia 141, 179–190 (2004).
Harvey, P. H. & Partridge, L. Murderous mandibles and black holes in hymenopteran wasps. Nature 326, 128–129 (1987).
Godfray, H. C. J. The evolution of clutch size in parasitic wasps. Am. Nat. 129, 221–233 (1987).
Rosenheim, J. A. Single-sex broods and the evolution of nonsiblicidal parasitoid wasps. Am. Nat. 141, 90–104 (1993).
Mayhew, P. J. & van Alphen, J. J. Gregarious development in alysiine parasitoids evolved through a reduction in larval aggression. Anim. Behav. 58, 131–141 (1999).
Pexton, J. J. & Mayhew, P. J. Immobility: The key to family harmony?. Trends. Ecol. Evol. 16, 7–9 (2001).
Hamilton, W. D. Extraordinary sex ratios. Science 156, 477–488 (1967).
Mayhew, P. J. & Hardy, I. C. Nonsiblicidal behavior and the evolution of clutch size in bethylid wasps. Am. Nat. 151, 409–424 (1998).
Zaviezo, T. & Mills, N. Factors influencing the evolution of clutch size in a gregarious insect parasitoid. J. Anim. Ecol. 69, 1047–1057 (2000).
Koppik, M., Tiel, A. & Hofmeister, T. S. Adaptive decision making or diferential mortality: What causes ofspring emergence in a gregarious parasitoid?. Entomol. Exp. Appl. 150, 208–216 (2014).
Visser, M. E., Van Alphen, J. J. & Hemerik, L. Adaptive superparasitism and patch time allocation in solitary parasitoids: An ESS model. J. Anim. Ecol. 61, 93–101 (1992).
Waage, J. K. & Ming, N. S. The reproductive strategy of a parasitic wasp: I. optimal progeny and sex allocation in Trichogramma evanescens. J. An. Ecol. 53, 401–415 (1984).
Harvey, J. A., Poelman, E. H. & Tanaka, T. Intrinsic inter-and intraspecific competition in parasitoid wasps. Ann. Rev. Entomol. 58, 333–351 (2013).
Harvey, J. A., Bezemer, T. M., Gols, R., Nakamatsu, Y. & Tanaka, T. Comparing the physiological effects and function of larval feeding in closely-related endoparasitoids (Braconidae: Microgastrinae). Physiol. Entomol. 33, 217–225 (2008).
Cloutier, C., Duperron, J., Tertuliano, M. & McNeil, J. N. Host instar, body size and fitness in the koinobiotic parasitoid Aphidius nigripes. Entomol. Exp. Appl. 97, 29–40 (2000).
Bai, B., Luck, R. F., Forster, L., Stephens, B. & Janssen, J. M. The effect of host size on quality attributes of the egg parasitoid Trichogramma pretiosum. Entomol. Exp. Appl. 64, 37–48 (1992).
Kazmer, D. J. & Luck, R. F. Field tests of the size-fitness hypothesis in the egg parasitoid Trichogramma Pretiosum. Ecology 76, 412–425 (1995).
Samková, A., Hadrava, J., Skuhrovec, J. & Janšta, P. Reproductive strategy as a major factor determining female body size and fertility of a gregarious parasitoid. J. Appl. Entomol. 143, 441–450 (2019).
Wei, K., Tang, Y. L., Wang, X. Y., Cao, L. M. & Yang, Z. Q. The developmental strategies and related profitability of an idiobiont ectoparasitoid Sclerodermus pupariae vary with host size. Ecol. Entomol. 39, 101–108 (2014).
May, R. M., Hassell, M. P., Anderson, M. R. & Tonkyn, D. V. Density dependence in host-parasitoid models. J. Anim. Ecol. 50, 855–865 (1981).
Hoddle, M. S., Van Driesche, R. G., Elkinton, J. S. & Sanderson, J. P. Discovery and utilization of Bemisia argentifolii patches by Eretmocerus eremicus and Encarsia formosa (Beltsville strain) in greenhouses. Entomol. Exp. Appl. 87, 15–28 (1998).
Samková, A., Raska, J., Hadrava, J. & Skuhrovec, J. An intergenerational approach for prediction of parasitoid population dynamics. BioRxiv. https://doi.org/10.1101/2021.02.22.432341 (2021).
Anderson, R. C. & Paschke, J. D. The biology and ecology of Anaphes flavipes (Hymenoptera: Mymaridae), an exotic egg parasite of the cereal leaf beetle. Ann. Entomol. Soc. Am. 61, 1–5 (1968).
Klomp, H. & Teerink, B. J. The significance of oviposition rates in the egg parasite Trichogramma embryophagum Htg. Arch. Neerl. Zool. 17, 350–375 (1967).
Waage, J. K. & Lane, J. A. The reproductive strategy of a parasitic wasp: II. Sex allocation and local mate competition in Trichogramma evanescens. J. Anim. Ecol. 53, 417–426 (1984).
Dysart, R. J., Maltby, H. L. & Brunson, M. H. Larval parasites of Oulema melanopus in Europe and their colonization in the United States. Entomophaga 18, 133–167 (1973).
Skuhrovec, J. et al. Insecticidal activity of two formulations of essential oils against the cereal leaf beetle. Acta Agr. Scand. 68, 489–495 (2018).
Jervis, M. A., Ellers, J. & Harvey, J. A. Resource acquisition, allocation, and utilization in parasitoid reproductive strategies. Annu. Rev. Entomol. 53, 361–385 (2008).
Vinson, S. B. & Iwantsch, G. F. Host suitability for insect parasitoids. Ann. Rev. Entomol. 25, 397–419 (1980).
Mackauer, M., Sequeira, R. & Otto, M. Growth and development in parasitoid wasps adaptation to variable host resources. In Vertical Food Web Interactions 191–203 (Springer, 1997).
Ode, P. J. Plant toxins and parasitoid trophic ecology. Curr. Opin. Insect sci. 32, 118–123 (2019).
Cronin, J. T. & Abrahamson, W. G. Do parasitoids diversify in response to host-plant shifts by herbivorous insects?. Ecol. Entomol. 26, 347–355 (2001).
Sarfraz, M., Dosdall, L. M. & Keddie, B. A. Host plant nutritional quality affects the performance of the parasitoid Diadegma insulare. Biol. Control. 51, 34–41 (2009).
Harvey, J. A. Factors affecting the evolution of development strategies in parasitoid wasps: The importance of functional constraints and incorporating complexity. Entomol. Exp. Appl. 117, 1–13 (2005).
Cortesero, A. M. & Monge, J. P. Influence of pre-emergence experience on response to host and host plant odours in the larval parasitoid Eupelmus vuilleti. Entomol. Exp. Appl. 72, 281–288 (1994).
Gandolfi, M., Mattiacci, L. & Dorn, S. Preimaginal learning determines adult response to chemical stimuli in a parasitic wasp. Proc. Roy. Soc. Lon. Series. B-Biol. Scien. 270, 2623–2629 (2003).
Kester, K. M. & Barbosa, P. Postemergence learning in the insect parasitoid, Cotesia congregata (Say) (Hymenoptera: Braconidae). J. Insect Behav. 4, 727–742 (1991).
Vet, L. E. & Groenewold, A. W. Semiochemicals and learning in parasitoids. J. Chem. Ecol. 16, 3119–3135 (1990).
Samková, A., Hadrava, J., Skuhrovec, J. & Janšta, P. Effect of adult feeding and timing of host exposure on the fertility and longevity of the parasitoid Anaphes flavipes. Entomol. Exp. Appl. 167, 932–938 (2019).
Jervis, M. A., Heimpel, G. E., Ferns, P. N., Harvey, J. A. & Kidd, N. A. Life-history strategies in parasitoid wasps: A comparative analysis of ‘ovigeny’. J. Anim. Ecol. 70, 442–458 (2001).
Bjorksten, T. A. & Hoffmann, A. A. Persistence of experience effects in the parasitoid Trichogramma nr. brassicae. Ecol. Entomol. 23, 110–117 (1998).
Lentz, A. J. & Kester, K. M. Postemergence experience affects sex ratio allocation in a gregarious insect parasitoid. J. Insect. Behav. 21, 34–45 (2008).
Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 47, 57–92 (2002).
Zvereva, E. L. & Rank, N. E. Fly parasitoid Megaselia opacicornis uses defensive secretions of the leaf beetle Chrysomela lapponica to locate its host. Oecologia 140, 516–522 (2004).
Roy, H. E., Handley, L. J. L., Schönrogge, K., Poland, R. L. & Purse, B. V. Can the enemy release hypothesis explain the success of invasive alien predators and parasitoids?. Biocontrol 56, 451–468 (2011).
Snyder, W. E. & Ives, A. R. Interactions between specialist and generalist natural enemies: Parasitoids, predators, and pea aphid biocontrol. Ecology 84, 91–107 (2003).
Polis, G. A., Myers, C. A. & Holt, R. D. The ecology and evolution of intraguild predation: Potential competitors that eat each other. Annu. Rev. Ecol. Syst. 20, 297–330 (1989).
Nakashima, Y. & Senoo, N. Avoidance of ladybird trails by an aphid parasitoid Aphidius ervi: active period and efects of prior oviposition experience. Entomol. Exp. Appl. 109, 163–166 (2003).
Samková, A., Raška, J., Hadrava, J., Skuhrovec, J. & Janšta, P. Female manipulation of offspring sex ratio in the gregarious parasitoid Anaphes flavipes from a new two-generation approach. BioRxiv https://doi.org/10.1101/2021.02.22.432331 (2021).
Visser, M. E. The importance of being large: the relationship between size and fitness in females of the parasitoid Aphaereta minuta (Hymenoptera: Braconidae). J. Anim. Ecol. 63, 963–978 (1994).
Banks, M. & Thomson, D. J. Lifetime mating success in the damselfly Coenagrion puella. Anim. Behav. 33, 1175–1183 (1985).
Ellers, J. & Jervis, M. Body size and the timing of egg production in parasitoid wasps. Oikos 102, 164–172 (2003).
Anderson, R. C. & Paschke, J. D. Additional Observations on the Biology of Anaphes flavipes (Hymenoptera: Mymaridae), with Special Reference to the Efects of Temperature and Superparasitism on Development. Ann. Entomol. Soc. Am. 62, 1316–1321 (1969).
Bezděk, J. & Baselga, A. Revision of western Palaearctic species of the Oulema melanopus group, with description of two new species from Europe (Coleoptera: Chrysomelidae: Criocerinae). Acta. Ent. Mus. Nat. Pra. 55, 273–304 (2015).
R. Core Team R. A language and environment for statistical computing. R Foundation for Statistical Computing (R Core Team, 2020).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw 67, 1–48. (2015) URL: https://CRAN.R-project.org/package=Hmisc.
Harrell, F. E. Jr, Dupont, C., et mult. al. (2020) Hmisc: Harrell Miscellaneous. R package version 4.4–2. URL: https://CRAN.R-project.org/package=Hmisc.
Signorell et mult. al. (2021). DescTools: Tools for descriptive statistics. R package version 0.99.40. URL: https://cran.r-project.org/package=DescTools.
Acknowledgements
We would like to thank the colleagues who provided us with technical equipment: Jana Mazáková, Marie Maňasová, Jana Wenzlová and Miloslav Zouhar. This work was supported by the Ministry of Agriculture of the Czech Republic, institutional support MZe-RO0418 and programme NAZV No. QK1910281 (MZe ČR) (both to JS).
Author information
Authors and Affiliations
Contributions
A.S., J.H. and J.S designed the experiment. A.S. collected the data and prepared figures 3,4,5. J.H. and J.R. analysed the data. J.R. and A.S. prepared figures 1,2. A.S., J.H., J.R. and J.S. prepared the manuscript. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Samková, A., Raška, J., Hadrava, J. et al. Effect of host switching simulation on the fitness of the gregarious parasitoid Anaphes flavipes from a novel two-generation approach. Sci Rep 11, 19473 (2021). https://doi.org/10.1038/s41598-021-98393-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-98393-y
- Springer Nature Limited