
Abstract
Hepatopathies can cause major metabolic abnormalities in humans and animals. This study examined differences in serum metabolomic parameters and patterns in left-over serum samples from dogs with either congenital portosystemic shunts (cPSS, n = 24) or high serum liver enzyme activities (HLEA, n = 25) compared to control dogs (n = 64). A validated targeted proton nuclear magnetic resonance spectroscopy platform was used to assess 123 parameters. Principal component analysis of the serum metabolome demonstrated distinct clustering among individuals in each group, with the cluster of HLEA being broader compared to the other groups, presumably due to the wider spectrum of hepatic diseases represented in these samples. While younger and older adult control dogs had very similar metabolomic patterns and clusters, there were changes in many metabolites in the hepatopathy groups. Higher phenylalanine and tyrosine concentrations, lower branched-chained amino acids (BCAAs) concentrations, and altered fatty acid parameters were seen in cPSS dogs compared to controls. In contrast, dogs with HLEA had increased concentrations of BCAAs, phenylalanine, and various lipoproteins. Machine learning based solely on the metabolomics data showed excellent group classification, potentially identifying a novel tool to differentiate hepatopathies. The observed changes in metabolic parameters could provide invaluable insight into the pathophysiology, diagnosis, and prognosis of hepatopathies.
Similar content being viewed by others

Introduction
The liver plays a central role in the metabolism of proteins, carbohydrates, lipids, fatty acids, and amino acids involved in both anabolic and catabolic processes. Therefore, both primary liver and systemic diseases that secondarily affect the liver can result in major metabolic abnormalities that impact quality of life and life expectancy1,2. Various congenital hepatic diseases, such as inborn errors of metabolism and structural anomalies, are recognized in humans3,4 and dogs5. Congenital portosystemic shunts (cPSS) are commonly seen vascular liver anomalies in young dogs6 and cats7, but occur very rarely in children8,9. In addition, there are many acquired acute and chronic hepatic diseases in humans and animals related to infectious, inflammatory, degenerative, vascular, neoplastic, drug- or toxin-related processes, as well as hepatopathies of idiopathic origin, which are associated with high serum liver enzyme activities10,11,12,13. Of major human public health concerns are acute and chronic hepatitis virus A to E infections and alcohol-related liver diseases14, which are not recognized in dogs15. However, viral hepatopathy in dogs can be induced by canine hepatitis adenovirus type 1, although this is now exceedingly rare due to general vaccination strategies16. While the majority of hepatitides in dogs are classified as idiopathic11, bacterial, parasitic, toxic, and immune-mediated causes are also recognized17,18.
A variety of diagnostic screening tests are applied to clinically detect hepatopathies and their consequences in human and veterinary medicine, including serum hepatic enzyme activities and liver function parameters, followed by liver imaging, biopsy, and other specific tests2,12. While methods to assess certain classes of specific metabolites in health and disease have been used for more than half a century, techniques that permit more comprehensive analyses of biological samples with metabolomic analyses have only become available in this millennium. Advanced technologies such as nuclear magnetic resonance (NMR) spectroscopy and mass spectrometry (e.g., gas chromatography-mass spectrometry [MS], liquid chromatography-MS) permit the precise identification and quantification of large numbers of metabolites, thereby informing on the impact of physiological and pathological processes on the metabolome and metabolic pathways19,20. There are several studies in human patients indicating characteristic metabolic abnormalities in patients with different hepatopathies21,22.
Metabolomic investigations of hepatopathies in veterinary medicine are still very limited23,24. Only a few studies have evaluated serum metabolomics with specific MS methods in dogs with liver diseases25,26. Moreover, determining of distinct groups of metabolites such as amino acids in dogs with cPSS has been reported27. Here, we utilize proton (1H) NMR spectroscopy to assess and compare the serum metabolome of dogs with cPSS, dogs with high serum liver enzyme activities (HLEA), and dogs with blood test results in the normal reference intervals. Our specific goals were to determine, if (1) specific metabolic abnormalities could be detected in serum from dogs with different hepatopathies by 1H NMR spectroscopy, (2) serum metabolomic patterns differ between the two hepatopathies as well as compared to control dogs, and (3) machine learning models would be capable of correctly differentiating groups solely based upon metabolomics data.
Methods
Samples and groups
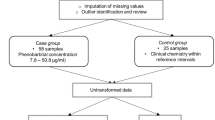
Left-over serum samples submitted for routine testing to a veterinary diagnostic laboratory (Laboklin GmbH & Co. KG, Bad Kissingen, Germany) between September 2020 and June 2021 from dogs with cPSS, HLEA, and apparently healthy younger (≤ 3 years [yr]) and older adult (> 3 yr) dogs with blood test results within the reference intervals were included in this study.
For sample selection, the laboratory’s electronic database was searched for:
-
cPSS group: samples with high serum bile acid (BA) concentrations (> 40 µmol/L, reference interval 0–15 µmol/L) and no or only slightly elevated age-related serum liver enzyme activities from dogs ≤ 3 years of age28.
-
HLEA group: samples with markedly increased serum alanine transaminase (ALT) activity (> 450 U/L, reference interval 0–55 U/L, representative of a moderate to marked increase29) without laboratory evidence of other diseases.
-
Control groups: samples with serum chemistry panel and complete blood count (CBC) results in the reference intervals (age-dependent28) were selected for control dogs ≤ 3 yr (Controls ≤ 3 yr) and > 3 yr of age (Controls > 3 yr), which were later combined because of only very minor metabolomic differences between them.
Available information on breed, age, sex, neutering status, and other data received from submission forms, the medical consult service, as well as blood test results were gathered and reviewed. For all groups, dogs were excluded if there was information available on concurrent diseases (e.g., infectious diseases, hyperadrenocorticism, cancer, or others) based upon submission forms and other laboratory tests performed at Laboklin. Furthermore, attending veterinary clinicians who submitted serum samples from dogs with high serum BA were contacted, and additional clinical information (including clinical signs and/or abdominal imaging or surgery) was obtained by Laboklin’s medical consult service (C.A.I.) to support the diagnosis of cPSS and exclude other diseases based upon the provided clinical information.
The laboratory’s inventory of frozen samples was screened for left-over serum samples with a residual volume of ≥ 300 µL. These serum samples were originally submitted to the laboratory after centrifugation and removal of clotted blood and were delivered at room temperature with a transport time of ≤ 1 day (except two samples which were kept and arrived chilled within two days). Samples with observed hemolysis, lipemia, and/or severe icterus were excluded. Frozen serum samples were thawed, aliquoted (1.8 ml CryoPure tubes, Sarstedt AG & Co. KG, Nürnbrecht, Germany) and refrozen at -80 °C until shipment for metabolomic analysis within ≤ 8 months. Serum chemistry analyses (Cobas 8000 c701 analyzer, Roche Diagnostics, Mannheim, Germany) were performed on thawed samples, if not already undertaken during routine testing. Results of CBC (ADVIA 2120i, Siemens Healthcare GmbH, Erlangen, Germany or Sysmex XT2000i, Sysmex Deutschland GmbH, Norderstedt, Germany) were reviewed if available.
Serum metabolomic analyses
Serum samples were shipped frozen overnight on ice packs to PetMeta Labs Oy, Helsinki, Finland. Metabolomic analysis was conducted using a targeted 1H NMR method optimized and validated for canine serum and plasma30. A similar method has been described and largely used for human serum and plasma31,32. Details of the method are published elsewhere 30,31,32. Briefly, the highly automated method utilizes a Bruker AVANCE III HD 500 MHz spectrometer equipped with a 5 mm triple-channel (1H, 13C, 15N) z-gradient Prodigy probe head and a cooled high-throughput sample changer SampleJet (Bruker Corp., Billerica, Massachusetts, USA). NMR analyses were conducted at Nightingale Health Oyj, Helsinki, Finland. Submitted samples were lightly mixed and centrifuged to remove any potential precipitates33. Each sample was transferred into a NMR tube (Bruker Corp., Billerica, Massachusetts, USA) and carefully mixed with sodium phosphate buffer in an automated process using a JANUS Automated Workstation equipped with an 8-tip dispense arm with Varispan (PerkinElmer Inc., Waltham, Massachusetts, USA)31. Two NMR spectra (so called LIPO and LMWM windows) were acquired automatically with standardized parameters from each sample. The LIPO window shows a conventional water-suppressed 1H NMR spectrum with broad and overlapping resonances arising mainly from proteins, lipoproteins, and lipids33. The LMWM window suppresses the broad signals of most macromolecules and lipids with T2-relaxation-filtering, improving the detection of low-molecular-weight metabolites33. The acquired NMR spectra are automatically processed using in-house scripts optimized for canine samples30, including background control, baseline removal, and signal alignments31. Absolute concentrations of the metabolic measures were obtained with a proprietary software with integrated quality control32. The quantification was based on regression modeling32 and results in the quantification of 123 measurands, including detailed lipoprotein analyses with lipoprotein subclass concentrations and compositions, amino acid concentrations and several amino acid ratios, fatty acids in both molar and relative units, and concentration of triglycerides, cholesterols, glycolysis-related metabolites, albumin, and creatinine30.
Statistical analyses
Univariate analyses
Univariate statistical analysis was performed using the SPSS Statistics (version 26; IBM Corp., Armonk, New York, USA) and MS Office Excel (Microsoft Corp., Redmond, Washington, USA) software programs. All continuous data were assessed for normal distribution. Differences in age, routine serum clinical chemistry results, and metabolomics data were evaluated using a Kruskal–Wallis test34, and for serum BA concentrations using a Mann–Whitney U test35. Kruskal–Wallis tests for metabolomic analyses were adjusted with Bonferroni correction36. Sex and neutering status between groups were evaluated using chi-square tests. The level of significance was set at p < 0.05.
Multivariate analysis
Concentrations of metabolomics data missing at random were imputed by the median of the corresponding variable, whereas concentrations below the detection limit were imputed with a zero value. Principal component analysis (PCA), partial least squares-discriminant analysis (PLS-DA), the hierarchical cluster heatmap, and hierarchical cluster analysis (HCA) of the serum metabolomics data were performed using MetaboAnalyst 5.037 using auto-scaled variables. A hierarchical cluster heatmap with samples and parameters clustered was created to visualize changes in the most discriminative parameters identified by variable importance in projection (VIP) scores in PLS-DA (selection of top 20 variables). Hierarchical cluster analyses were performed using the Ward clustering algorithm and the Euclidean distance measure38. Machine learning methods were performed with WEKA 3.9539. Models applied here were simple logistic regression40, support vector machines41, k-nearest neighbors (KNN) algorithm42, Multilayer Perceptron (MLP) Classifier43, Random Forest44, and multinomial naïve Bayes45. The default settings of the parameters for the respective WEKA implementation were used for all machine learning methods. Machine learning models were tested using 10-fold full cross-validation for each model.
Briefly, PCA is an unsupervised pattern recognition method that is used for dimension reduction. The first two principal components explain the most possible variance of the data. Scores plots derived by PCA reveal clustering or separation of individual data points (samples), thereby enabling determination of similarities46. In contrast, PLS-DA is a supervised statistical analysis that uses prior knowledge of group assignments and thereby maximizes the separation of groups. Partial least squares-discriminant analysis is based on PLS-regression using a dummy variable encoding the class membership47. Hierarchical cluster analysis determines sample clustering by merging samples into homogeneous clusters based on homogeneity of measured serum metabolites.
Univariate analysis and PCA of the serum metabolomics data revealed only minor differences between the younger and older adult Control groups. Thus, both Control groups were combined in the further statistical analyses of the serum metabolomics data in this study to simplify analyses and increase the statistical power for machine learning approaches.
Ethics declaration
The use of left-over blood samples for research purposes is stated in the general terms and conditions of Laboklin GmbH & Co. KG and was approved by the government in Würzburg, Lower Franconia, Bavaria, Germany (RUF-55.2.2-2532-1-86-5).
Results
Samples, demographics, and routine laboratory test results
The serum samples selected fulfilled the respective group’s entry criteria for dogs with either high serum BA concentrations (cPSS) or high liver enzyme activities (HLEA) or originating from younger and older adult dogs with serum chemistry and CBC results within age-dependent reference intervals and thus no laboratory evidence of liver or other disease (Control groups) (Supplementary Table S1). While every effort was made to select cases based upon the laboratory and clinical information available, the animals were not examined by the authors.
A total of 113 left-over canine serum samples were included and analyzed by 1H NMR spectroscopy in this study, including 24 cPSS serum samples and 25 HLEA serum samples. The Control groups of dogs ≤ 3 yr and > 3 yr included 32 serum samples each. Serum samples were mostly from Germany (95%) and rarely other European countries (Czech Republic, Italy, Luxembourg, and Sweden). As designed, dogs with cPSS and Controls ≤ 3 yr were of a similar age, as were the HLEA and Control group > 3 yr (Fig. 1a, Supplementary Table S1). Mixed breed dogs accounted for the largest proportion enrolled in any group. No breed predilection and no sex differences were observed between groups, except a higher number of neutered dogs with HLEA (Supplementary Table S1).

(a) Age, (b) alanine transaminase activities (ALT), and (c) bile acid (BA) concentrations in serum samples from dogs with cPSS (n = 24), HLEA (n = 25), and younger and older adult control dogs (n = 32, for each). Bile acid concentrations were determined in all dogs with cPSS and in 24 of 32 controls dogs of ≤ 3 yr. Boxes indicate the lower to upper quartile (25th–75th percentile), and median value. Whiskers extend to minimum and maximum values. Outliers are shown as individual open circles or red stars. Grey boxes indicate reference intervals. Lines above figures reflect significant differences between specific groups (*p < 0.05; **p < 0.001).
As determined by the inclusion criteria, dogs with HLEA showed significantly higher serum ALT activities than all other groups (Fig. 1b; Supplementary Table S1). Serum BA concentrations were highly elevated in dogs with cPSS compared to Controls ≤ 3 yr, which were only available for these two groups (Fig. 1c; Supplementary Table S1).
Metabolomic analyses
The 1H NMR spectroscopy platform for metabolic profiling studies was recently validated for 123 metabolic parameters with 8,247 canine serum and plasma samples30. In the present study, all 105 serum metabolite concentrations were identifiable and measurable. Furthermore, 11 relative concentrations of fatty acids and seven selected amino acid ratios could be calculated, totaling 123 parameters as in the recently published validation study for canine samples30. Medians for all parameters for younger as well as older adult control dogs fell in the previously established serum reference intervals for dogs of all ages with 90% confidence intervals (CIs)30, except for the median concentrations of glutamine and L-VLDL-triglycerides, which were slightly lower in the Control group ≤ 3 yr. Moreover, for a few parameters, 25th or 75th percentiles of the Control groups in this study fell slightly below (concentrations of glutamine, citrate, acetate, glycoprotein acetyls [GlycA], and large very-low-density lipoprotein [L-VLDL]-triglycerides) or slightly above (concentration of glycine, glycine/branched-chain amino acid [BCAA] ratio, glycine/valine ratio, relative concentration of linoleic acid, relative concentration of omega-6 fatty acids, omega-6/omega-3 fatty acid ratio, high-density lipoprotein [HDL] particle size, and concentration of small low-density lipoprotein [S-LDL]-triglycerides) the published serum reference intervals for dogs of all ages30 (Supplementary Table S2).
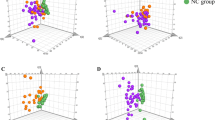
Because of the expected age difference of dogs with cPSS (younger) and HLEA (older), two age-dependent Control groups with dogs that were younger or older than 3 years of age were initially set up. However, univariate testing revealed only one slight difference between the two Control groups for citrate concentrations (p = 0.049), with the younger Control group having slightly lower median serum concentrations and the 25th percentile below the reference interval (Supplementary Table S2). Moreover, PCA demonstrated near-complete overlap between the clusters of the younger and older adult Control groups (Fig. 2a). Therefore, the two Control groups were combined in the subsequent statistical analyses of the metabolomics data.

(a) Scores plot of principal component analysis (PCA) showing overlap between serum samples from dogs of the younger (≤ 3 yr, green) and older adult (> 3 yr, red) control dogs, indicating near-complete overlap (n = 32, for both). (b) Scores plot of PCA of the serum metabolome from dogs with cPSS (blue, n = 24) and HLEA (green, n = 25) and the combined Control group (red, n = 64). Shaded circles represent 95% confidence intervals, while colored dots illustrate individual samples. The axes are labelled by the first and second principal component (PC 1 and 2, respectively) with the percentages of variance of the data explained by that principal component in parentheses.
Univariate testing of serum metabolomics data showed significant differences in 100 of 123 parameters between dogs with cPSS, HLEA, and/or the combined Control group (Supplementary Table S2).
The PCA revealed clustering of canine samples from cPSS, HLEA, and combined Controls based on the serum metabolome (Fig. 2b). The total variance of the first two principal components contributed 61% in the PCA model for all three groups (PC 1 = 40.3%, PC 2 = 20.7%). Both the cPSS and the combined Control group showed tight clustering, while the HLEA group clustered more broadly, overlapping with the other two clusters. Many variables were found to influence the projection and separation of the groups as reflected in marginalized metabolites in the PCA’s loadings plot (Supplementary Fig. S1). To maximize the separation of the groups observed by PCA, PLS-DA was applied. In the PLS-DA model for the three groups, the first two components contributed to a total variance of 47.3%. The three groups showed robust clustering, which partially overlapped, particularly with the larger HLEA cluster (Fig. 3a). The PLS-DA model was validated by a 10-fold cross-validation with R2, Q2, and accuracy as displayed in Supplementary Fig. S2. All figures show a robust model with four components being selected as optimal number of components based on the Q2 criterion. Furthermore, a permutation test with 1000 permutations was performed. The results displayed in Supplementary Fig. S3 demonstrate that the model is not overfitting the data. None of the 1000 permutations achieved the same separation distance defined by the ratio of the between-group sum of the squares and the within-group sum of the squares (B/W-ratio)48. The variables most responsible for separation in the PLS-DA were ranked according to the VIP scores. The metabolites identified as most discriminating between the three groups were phenylalanine, tyrosine, S-HDL esterified cholesterol and S-HDL cholesterol concentrations, and relative concentrations of palmitic acid, arachidonic acid, and saturated fatty acids, followed by others (Fig. 3b).

(a) Scores plot of partial least squares-discriminant analysis (PLS-DA) based on metabolomics data between serum samples of dogs with cPSS (blue, n = 24), HLEA (green, n = 25) and combined Controls (red, n = 64). Shaded circles represent 95% confidence intervals, while colored dots illustrate individual samples. The axes are labelled by the first and second components with the percentages of variance of the data explained by that component in parentheses. (b) Variable importance in projection (VIP) scores of component 1 of the PLS-DA identifies the top 20 discriminating parameters in descending order of importance from the serum metabolomics data of dogs with cPSS (n = 24), HLEA (n = 25) and the combined Control group (n = 64). The colored legend on the right indicates the relative abundance of variables, with red and blue indicating high and low values, respectively, while beige illustrates neutral values.
Among the altered metabolites between groups were multiple amino acids, with some having high discriminatory power. For instance, dogs with cPSS showed higher serum concentrations of the aromatic amino acids (AAAs) phenylalanine and tyrosine, as well as of histidine, compared to the Control group. Moreover, dogs with cPSS had significantly lower concentrations of total BCAAs, particularly leucine and valine (but not isoleucine), while dogs with HLEA had higher concentrations of total BCAAs, particularly isoleucine and valine (but not leucine) compared to the Control group (Fig. 4a, Supplementary Table S2). Phenylalanine and tyrosine were found to be relevant parameters for group separation in PLS-DA (Fig. 3b). Phenylalanine concentrations were elevated in dogs with both hepatopathies (Fig. 4b). While being a relevant marker in PLS-DA to distinguish between the three groups, there was no significant difference by univariate testing between dogs with cPSS or HLEA. However, serum tyrosine concentrations were only high in dogs with cPSS (Fig. 4c).

Concentrations (mmol/L) of total branched-chain amino acids (BCAAs) (a), phenylalanine (b), tyrosine (c), percentage of saturated fatty acids (d), percentage of arachidonic acid (e), and concentrations (mmol/L) of glycoprotein acetyls (GlycA) (f) in serum samples from dogs with cPSS (n = 24), HLEA (n = 25), and the combined Control group (n = 64). Boxes indicate the lower to upper quartile (25th–75th percentile) and median value. Whiskers extend to minimum and maximum values. Outliers are shown as individual open circles or red stars. Grey boxes indicate reference intervals. Lines above figures reflect significant differences between specific groups (*p < 0.05; **p < 0.001).
Several serum lipoprotein concentrations as well as the lipoprotein compositions were altered in dogs with cPSS and HLEA compared to the Control group (Supplementary Table S2, Supplementary Fig. S1). In particular, concentrations of small-HDL particles, lipids, cholesterol, esterified cholesterol, and phospholipids were found to have a discriminative power in PLS-DA (Fig. 3b) and were decreased in dogs with cPSS and, to a lesser extent, in the HLEA group. Similarly, serum concentrations of large-HDL particles, lipids, cholesterol, and esterified cholesterol were decreased in dogs with cPSS, but normal in the HLEA group (except large-HDL particles), and therefore were discriminatory parameters in PLS-DA (Fig. 3b). In contrast, dogs with HLEA showed increased concentrations of VLDL and LDL particles. Finally, total cholesterol concentrations were higher in dogs with HLEA, while total cholesterol concentrations were lower in dogs with cPSS compared to the Control group. Moreover, serum total triglyceride concentrations were trending to be increased in dogs with HLEA compared to the Control group (p > 0.05) (Supplementary Table S2).
Absolute fatty acid concentrations were generally decreased in serum from dogs with cPSS (except docosahexaenoic acid) and mostly increased in dogs with HLEA compared to the Control group (Supplementary Table S2). Moreover, the relative quantities of some serum fatty acids were altered in dogs of either disease state compared to the Control group. For instance, dogs with cPSS showed lower relative concentrations of polyunsaturated fatty acids, such as the omega-6-fatty acid arachidonic acid and docosapentaenoic acid, compared to the Control group (Supplementary Table S2). In contrast, the relative concentrations of saturated fatty acids (Fig. 4d), specifically palmitic acid, were increased in dogs with cPSS. Both relative concentrations of arachidonic and palmitic acid were identified as relevant parameters for group separation by PLS-DA (Fig. 3b). In contrast, dogs with HLEA had higher relative concentrations of stearic acid and decreased relative concentrations of arachidonic acid (Supplementary Table S2, Fig. 4e). The decrease in the relative concentration of arachidonic acid was more apparent in dogs with cPSS (Fig. 4e).
The inflammatory biomarker GlycA, measured by 1H NMR spectroscopy, was increased in dogs with HLEA compared to Controls, while dogs with cPSS had serum GlycA concentrations within the reference interval (Fig. 4f).
In addition, serum acetate and lactate concentrations were increased in both hepatopathies (Supplementary Table S2). Citrate concentrations were low and high in dogs with cPSS and HLEA, respectively. However, the citrate and lactate changes were marginal and mostly in the established reference interval30. Serum concentrations of glucose and pyruvate did not differ between the groups (Supplementary Table S2).
In a hierarchical cluster heatmap of the above-mentioned parameters identified by VIP scores in PLS-DA, the pattern and clustering for the cPSS samples was distinct, while the samples of dogs with HLEA and the Control group were intermixed and showed less intragroup homogeneity (Fig. 5).

Hierarchical cluster heatmap (for samples and variables) of serum metabolomic results using the top 20 parameters identified by partial least squares-discriminant analysis (PLS-DA) variable importance in projection (VIP) scores. Each column represents one serum sample with group marking colored at the top: blue cPSS (n = 24), green HLEA (n = 25), and red Control group (n = 64). The colored legend on the right indicates the relative metabolite concentrations, with different red and blue intensities indicating high and low values, respectively. Horizontal and vertical black lines depict clustering of samples and parameters.
Hierarchical cluster analysis using the Ward clustering algorithm and the Euclidean distance measure was used to classify samples into homogenous clusters based on similarities of their serum metabolomic results (Fig. 6). Most of the samples from dogs with cPSS and Controls could be assigned to two distinct clusters, indicating within-group similarities. In contrast, only 40% of HLEA samples formed a separate cluster, and many HLEA samples assigned to clusters of control or cPSS samples.

Dendrogram of hierarchical cluster analysis of serum metabolomic results from dogs with cPSS (blue, n = 24), HLEA (green, n = 25), and the combined Control group (red, n = 64). Each number on the x-axis reflects a serum sample. The y-axis shows the similarity levels expressed as Euclidean distances. Horizontal and vertical lines depict clustering of samples and differences of the distances, respectively.
Several machine learning methods were applied to predict the correct assignment of samples based solely on their serum metabolomics data and without considering any routine clinical pathology results. Depending on the model used, a correct classification was achieved in 83–94% of samples using 10-fold cross-validation (Supplementary Table S3). For instance, the simple logistic regression model40 was able to classify 106 of 113 samples correctly based solely on the serum metabolomic analysis (Table 1).
Discussion
While measurements of singular metabolic parameters have been previously used to clinically assess patients in human and veterinary medicine, more recent comprehensive analytic test methods are now available to assess the metabolome in health and disease19,20. For example, while amino acid analyzers or gas chromatography can be used individually or in combination to identify and quantify distinct groups of low-molecular weight metabolites, 1H NMR spectroscopy can assess a broad range of metabolites without the need for tedious sample preparation steps like derivatization and only requires a very small sample volume (≤ 100µL). In this study, we have applied this novel metabolomics platform to identify metabolic abnormalities in serum samples from dogs with either cPSS (n = 24) or HLEA (n = 25) for comparison with dogs with blood test results within age-dependent reference intervals (n = 64). Hierarchical cluster analysis indicates within-group similarities in serum metabolomic results for samples from both the cPSS and Control groups. However, samples from dogs with HLEA were more randomly distributed within the cPSS and the Control group clusters, showing that the serum metabolomes of dogs with HLEA are more heterogeneous (Fig. 6). Notably, the broader clusters of HLEA observed by PCA (Fig. 2b) and PLS-DA (Fig. 3a) likely reflect the variety of hepatic diseases that may be included in this group (no histopathological results available). Moreover, dogs with cPSS have hepatic atrophy, while the liver of dogs with HLEA may show inflammation, destruction, and regeneration12.
The 1H NMR spectroscopy platform used here quantifies 105 metabolites and calculates 18 fractions and ratios of amino acids and/or fatty acids in canine serum samples. For the younger and older adult Control groups, the medians and the 25th and 75th percentiles of serum metabolomic parameters fell largely into the recently published normal serum reference intervals for dogs of all ages30. The minor differences may relate to the smaller number of samples and different sample handling practices and demographics compared to the validation study30. The serum metabolomic results for both Control groups (≤ 3 yr versus > 3 yr) showed only minor differences and near-complete overlapping of clusters in PCA and thus were combined in further analyses (Fig. 2a).
Most serum metabolite concentrations measured by 1H NMR spectroscopy showed differences between the three groups, and distinct metabolomic patterns differentiated the two hepatic conditions from each other, as well as from controls. Among the 123 metabolomic parameters measured, 100 were significantly different in dogs with cPSS, HLEA, and/or the combined Control group. Overall, many abnormalities appeared marginal compared to the established large reference intervals30. Interestingly, most lipid-related metabolic abnormalities moved in the opposite direction for cPSS and HLEA. Other parameters, for instance, phenylalanine concentrations, moved in varying degrees in the same direction for both hepatopathies. Finally, as these canine hepatopathies reflect similarities to human conditions, our findings may also be insightful for differentiating hepatopathies in humans. Applying pattern recognition methods and successfully classifying distinct diseases solely by metabolites could be an alternative to otherwise invasive diagnostic approaches, especially with hepatopathies.
The metabolomic changes observed in this study measured by 1H NMR spectroscopy in canine hepatopathies confirm and expand prior metabolomic studies, which were carried out using smaller sample sizes and primarily focused on other specific groups of metabolites25,26. One study identified 50 metabolites with significant differences, however, no metabolite concentrations and only measured peak heights were made available25. As age-matched control samples were missing in prior investigations25,26, our findings that there are no major differences in metabolites or metabolic patterns in a comparative study of younger and older adult control dogs will aid interpretation of prior and future studies.
Machine learning models, solely utilizing the serum metabolomic parameters, correctly assigned a major proportion of samples to the groups of control dogs and dogs with either cPSS or HLEA (Table 1, Supplementary Table S3). In human patients with non-alcoholic fatty liver disease, non-alcoholic steatohepatitis, or non-alcoholic steatohepatitis associated cirrhosis, machine learning models were applied to differentiate these diseases and showed good accuracy49. In the future, these methods could be further explored for diagnosis and monitoring of patients with various hepatopathies. As we focused on dogs with hepatopathies in this study, we did not include the assessment of serum samples from dogs with other disorders. Such analyses will be needed to better categorize disease states by multivariate analyses, including machine learning. In addition, the broad clusters of the HLEA group in PCA and PLS-DA indicate that the study of additional specific hepatopathies and severity states are required to determine if metabolomic cluster analyses and machine learning can differentiate them with or without routine diagnostic analysis. Due to the use of left-over samples, the information on clinical features, diet, and fasting/feeding status of the dogs was limited. Therefore, further prospective studies with clinically and histopathologically well defined groups, preferably with follow-up during and after treatment, are needed to confirm our findings.
Below, selective changes in specific metabolite groups in dogs with cPSS and HLEA are discussed:
Amino acid abnormalities
In contrast to dedicated amino acid high-performance liquid chromatography (HPLC) analyzers which require more sample processing, the 1H NMR spectroscopy platform used here measures only nine amino acids. Still, those measured (e.g., BCAAs, phenylalanine, and tyrosine) are of particular interest when assessing hepatopathies. However, other metabolomic methods able to quantify more serum amino acids revealed that alterations in cysteine, serine, methionine, and threonine were additional distinguishing parameters of cPSS and hepatitis in small groups of dogs25. As serum serine and methionine concentrations changed during medical treatment in dogs with cPSS27, they could also be considered for further studies in hepatopathies. Indeed, the BCAAs (leucine, isoleucine, and valine) and AAAs (phenylalanine and tyrosine) concentrations were altered in both hepatopathy groups of this study. Both groups showed high serum concentrations of phenylalanine, and dogs with cPSS also showed high concentrations of tyrosine (Fig. 4b and 4c) and histidine. In contrast, concentrations of total BCAAs were low in dogs with cPSS, while dogs with HLEA had high concentrations of total BCAAs compared to the Control group (Fig. 4a). Similar changes in the amino acid panel of dogs with cPSS have previously been reported using other methods25,27,50,51. While, increased phenylalanine concentrations have been previously found in dogs with either hepatitis or liver neoplasia, the degree of elevation could not differentiate the two conditions52. Likewise, in our study, the difference between serum phenylalanine concentrations in dogs with cPSS and HLEA did not reach significance by univariate analysis (Fig. 4b), but phenylalanine was still a relevant marker in PLS-DA to distinguish between the two hepatopathies.
Accordingly, the BCAA to AAA ratio was reduced, which also occurs in humans with liver cirrhosis53,54,55,56,57. The increases of tyrosine and phenylalanine in our canine study may reflect impaired hepatic metabolism and shunting of blood, as both are mainly catabolized in the liver54,58. The reduction in BCAA concentrations with hepatopathies may be due to several factors, including malnutrition, portosystemic shunting, increased gluconeogenesis, and conversion of ammonia into glutamine in the skeletal muscle56,57. The altered BCAA and AAA concentrations may contribute to hepatic encephalopathy59,60,61 and to decreased albumin synthesis62. These amino acid changes may be guiding the supplementation and restriction of amino acids in canine hepatopathies. A reduction in phenylalanine and tyrosine, and a rise in valine concentrations were achieved in dogs after shunt surgery, indicating the potential for metabolic reversibility27.
Abnormalities of glycolysis related metabolites
The increased serum lactate concentrations in dogs with cPSS and HLEA observed in this study were marginal and values stayed mostly in the established reference intervals30. Humans with liver cirrhosis63 or fulminant liver failure show elevated lactate concentrations64,65, potentially related to impaired hepatic lactate clearance with reduction of hepatic gluconeogenesis and increased glycolysis66,67. In people with non-alcoholic fatty liver disease, increased plasma levels of citrate, an intermediary metabolic substrate, were observed and associated with increased breakdown of fatty acids68. Also, human patients with fibrosis due to chronic hepatitis C69 and liver cirrhosis70 showed increased citrate concentrations. Interestingly, dogs with cPSS had slightly diminished citrate concentrations, which deserves further examination in the future. Serum acetate concentrations were increased in both hepatopathies, while only dogs in the HLEA group showed increased fatty acid concentrations. The liver hardly metabolizes any free acetate, which is instead generated during accelerated ketogenesis from fatty acid oxidation71 and higher levels could be associated with a ketogenic situation. However, glucose and glycolytic parameters, e.g., lactate, pyruvate, and their ratio may be affected by sample handling and storage as they are sensitive to post collection metabolic activity and thus values may not properly reflect the in vivo situation72.
Finally, in a small study of serum from dogs with cPSS and chronic hepatitis, an alternative metabolomics platform (gas chromatography-MS) suggested that a slight increase in the serum xylitol peak height might be one of the main metabolic determinants in differentiating hepatopathies from healthy controls25. However, in a human metabolomics study, decreased xylitol serum concentrations were found in patients with hepatitis B73. Xylitol, which was not measured in this 1H NMR study, is an intermediate product of uronic acid pathway and is needed for the production of xylulose. In contrast to plants, concentrations of xylitol in mammals seem to be low due to faster metabolism74. The biological importance of xylitol in liver disease remains unknown.
Lipid metabolism abnormalities
In contrast to other metabolomic analyzers, the 1H NMR platform can quantify a larger number of lipids, lipoproteins, and fatty acids. However, as 1H NMR spectroscopy has not been utilized extensively to date in dogs, little has been published on the impact of various canine hepatopathies on these serum metabolite concentrations. It has been previously shown that cholesterol concentrations can be low in cPSS and other hepatopathies with impaired liver function10, as was also observed in the cPSS group of our study, albeit values were still in the reference interval. In contrast, many diseases, including chronic hepatitis75, cholangitis/cholangiohepatitis76, and cholelithiasis77, which are associated with elevated serum liver enzyme activities, can show hypercholesterinemia and hypertriglyceridemia. Although the specific causes of hepatic disease of dogs in the HLEA group were not determined in this study (no liver histopathology), dogs within this group had high serum cholesterol concentrations. Hypercholesterinemia in liver disease in humans and animals is related to cholestasis10,13, as well as to increased hepatic synthesis and decreased lecithin-cholesterol acyltransferase13. Moreover, while concentrations of most lipoproteins were also elevated in dogs with HLEA, they remained normal or low in dogs with cPSS compared to the Control group. However, concentrations of small-HDL particles, as well as cholesterol, lipids, and phospholipids within these particles, were lower in dogs with HLEA and even lower in dogs with cPSS compared to the Control group. Being the smallest lipoproteins, HDL are secreted by the liver and intestine and transport cholesterol back to the liver78. Low HDL cholesterol levels are seen in human patients with liver diseases such as acute viral hepatitis and severe cholestasis79, as well as liver cirrhosis, and can arise from multiple causes80,81,82. Lower HDL cholesterol concentrations were also associated with decreased survival time in patients with liver cirrhosis and acute gastrointestinal bleeding81 and also predicted the development of complications associated with liver cirrhosis82. Indeed, lower HDL concentrations were suggestive of more severe chronic liver failure in human patients83. As small-HDL and large-HDL particles, and some of their associated lipids and cholesterols had discriminative power in our PLS-DA (Fig. 3b), further studies are warranted to evaluate not only the discriminatory power between groups but also to study the association of those parameters with severity and recovery or progression of canine hepatopathies.
It is advantageous to assess individual fatty acids by both absolute concentrations and their concentrations relative to total fatty acid concentrations to better reflect changes of individual fatty acids32. While the absolute concentrations of individual free and esterified fatty acids uniquely determined by the 1H NMR platform here reflect the changes in total concentrations of triglycerides and cholesterol, the changes in relative concentrations of individual fatty acids vary. Total fatty acid concentrations and nearly all individual fatty acid concentrations (except absolute concentrations of arachidonic, docosapentaenoic, and docosahexaenoic acid) were elevated in dogs with HLEA compared to the Control group. In contrast, dogs with cPSS showed reduced concentrations of total and all individual fatty acids (except docosahexaenoic acid) compared to the Control group. In a study of hepatectomized dogs, relative serum concentrations of arachidonic acid were reduced and associated with impaired arachidonic acid synthesis84. Likewise, humans with various liver diseases have reduced arachidonic and polyunsaturated fatty acids, such as linoleic or linolenic acid, likely due to impaired liver metabolism and shunting79,85,86. While there was no observed difference in relative concentrations of linoleic acid in any group in the present study, dogs with cPSS and HLEA showed lower relative concentrations of arachidonic acid, which was most apparent in dogs with cPSS (Fig. 4e).
Inflammatory biomarker
Glycoprotein acetyls represent a heterogenous signal of N-acetylglucosamine residues within certain acute-phase proteins, mainly α1-acid glycoprotein, α1-antitrypsin, α1-antichymotrypsin, haptoglobin, and transferrin87,88. Indeed, measurement of GlycA by 1H NMR spectroscopy serves as an inflammatory biomarker87,88,89,90, and in humans serum concentrations of GlycA correlate with increases in other inflammatory biomarkers, such as high-sensitivity C-reactive protein, fibrinogen, interleukin-6, and serum amyloid A concentrations87,89,90,91. Several diseases have been associated with increased GlycA concentrations, including type 2 diabetes, cardiovascular diseases, autoimmune diseases, obesity, and others92,93,94,95,96.
Normal ranges of serum GlycA concentrations have been assessed by 1H NMR spectroscopy in dogs30 and were increased in phenobarbital-treated dogs97. In addition, serum α1-acid glycoprotein concentrations were elevated in dogs with acute hepatopathies98. Furthermore, in one study99 serum haptoglobin concentrations were found to be high and low in dogs with hepatitides and end-stage liver disease, respectively. However, in another study serum haptoglobin concentrations remained unchanged in dogs with hepatitis or cPSS100. While no acute-phase protein concentrations were measured in our study, the serum GlycA concentrations were normal and high in dogs with cPSS and HLEA, respectively (Fig. 4f). This is consistent with the different disease etiology as cPSS arises from a non-inflammatory vascular anomaly, while the HLEA group likely included dogs with inflammatory hepatopathies.
Limitations
As mentioned throughout the manuscript, this study has several limitations based on its use of left-over samples submitted to a veterinary reference laboratory. Dogs in the two hepatopathy groups as well as control dogs were carefully selected based upon routine blood test results and other information gathered from submission forms. For the cPSS group, clinical information gathered from the attending veterinarians was also assessed. However, the animals were not directly examined by the authors and histopathological diagnoses were not available to further define the origin and underlying etiology of the elevated serum liver enzyme activities in the HLEA group. Serum bile acid concentrations were not available for dogs with HLEA, as this parameter would be confounded by hyperbilirubinemia in many cases. Dogs with HLEA could also have acquired portosystemic shunts. While dogs in the Control group showed routine clinical chemistry and hematological test results within the reference intervals, occult diseases or diseases unable to be detected via routine blood test results cannot be fully excluded. However, routine blood and metabolomic test results of the control dogs were tightly grouped and well within the published normal reference intervals.
While guidelines and general veterinary practice recommend collection of blood samples from fasted dogs for diagnostic testing, the fasting status of the dogs could not be definitively determined. Typically dogs with cPSS are inappetent101 and thus likely reflect fasting samples. Furthermore, postprandial bile acid results are not needed when high fasting serum bile acid concentrations were found on first sample for the diagnosis of cPSS. Finally, we only included samples with high bile acid concentrations > 40 µmol/L, which would be indicative of cPSS in both fasting and postprandial samples.
Samples are typically sent overnight to reference laboratories from veterinary practices (laboratory’s courier service). The stability of metabolites in canine serum samples was demonstrated in the validation study of the 1H NMR method30. However, some metabolite concentrations can change within minutes to hours of collection72 and different travel times might have caused some variation in metabolite values.
Despite these limitations, the two hepatopathy and Control groups clearly differed by routine as well as metabolomic parameters. As major differences were noted, further studies using more standardized sample acquisition from fasted dogs with more expedient serum separation and freezing are warranted. In the future, this technique may allow further differentiation of these groups into subgroups based upon liver histopathology or into specific types of cPSS.
Conclusion
Targeted metabolomic 1H NMR analysis revealed major metabolomic abnormalities in the serum of dogs with high liver enzyme activities and dogs with congenital portosystemic shunts. Among altered metabolites were amino acids, such as BCAAs, phenylalanine and tyrosine, but also fatty acids, lipids, GlycA, and others. Identification of altered biomarkers and metabolic pathways may improve our understanding of the pathophysiology and be helpful for diagnostics of hepatopathies in dogs. Potentially, further studies evaluating the effect of treatment on serum metabolomics might identify additional parameters to be used as biomarkers in therapy management and prognosis. Serum metabolomic patterns of both canine hepatopathy groups varied markedly and machine learning models showed great potential for classifying hepatopathies based solely on the metabolomics data. While our study used left-over samples, additional studies with distinct hepatopathies beyond cPSS and histopathological diagnoses are warranted to confirm and expand our promising findings and determine if singular parameters such as phenylalanine or tyrosine might be used in routine diagnostics to support laboratory differentiation between liver diseases, thereby advancing precision medicine.
Data availability
The datasets generated and analyzed during this study are available from the corresponding author upon reasonable request.
References
Tennant, B. C. & Center, S. A. Hepatic function. In Clinical Biochemistry of Domestic Animals 6th edn (ed. Kaneko, J. J.) 379–412 (Elsevier Saunders, 2008).
Boyer, T. D. et al. (eds) Zakim and Boyer’s Hepatology. A Textbook of Liver Disease 6th edn. (Saunders Elsevier, 2012).
Andres, J. M. Neonatal hepatobiliary disorders. Clin. Perinatol. 23, 321–352 (1996).
Rocha, G. et al. Disorders of the neonatal liver and bile ducts. Acta Med. Port. 23, 767–776 (2010).
Watson, P. Canine breed-specific hepatopathies. Vet. Clin. N. Am. Small Anim. Pract. 47, 665–682. https://doi.org/10.1016/j.cvsm.2016.11.013 (2017).
Ewing, G. O., Suter, P. F. & Bailey, C. S. Hepatic insufficiency associated with congenital anomalies of the portal vein in dogs. J. Am. Anim. Hosp. Assoc. 10, 463–467 (1974).
Ruland, K. et al. Portosystemic shunts in cats—evaluation of six cases and a review of the literature. BMTW 122, 211–218 (2009).
Sokollik, C., Bandsma, R. H. J., Gana, J. C., van den Heuvel, M. & Ling, S. C. Congenital portosystemic shunt: characterization of a multisystem disease. J. Pediatr. Gastroenterol. Nutr. 56, 675–681. https://doi.org/10.1097/mpg.0b013e31828b3750 (2013).
Baiges, A. et al. Congenital extrahepatic portosystemic shunts (Abernethy Malformation): an international observational study. Hepatology 71, 658–669. https://doi.org/10.1002/hep.30817 (2020).
Chapman, S. E. & Hostutler, R. A. A laboratory diagnostic approach to hepatobiliary disease in small animals. Vet. Clin. N. Am. Small Anim. Pract. 43, 1209–1225. https://doi.org/10.1016/j.cvsm.2013.07.005 (2013).
Poldervaart, J. H., Favier, R. P., Penning, L. C., van den Ingh, T. S. G. A. M. & Rothuizen, J. Primary hepatitis in dogs: a retrospective review (2002–2006). J. Vet. Intern. Med. 23, 72–80. https://doi.org/10.1111/j.1939-1676.2008.0215.x (2009).
Cocker, S. & Richter, K. Diagnostic evaluation of the liver. In Textbook of Veterinary Internal Medicine: Diseases of the Dog and the Cat 8th edn (eds Ettinger, S. J. et al.) 1611–1621 (Elsevier Saunders, 2017).
Poynard, T. & Imbert-Bismut, F. Laboratory testing for liver disease. In Zakim and Boyer’s Hepatology: A Textbook of Liver Disease 6th edn (eds Boyer, T. D. et al.) 201–215 (Saunders Elsevier, 2012).
Blachier, M., Leleu, H., Peck-Radosavljevic, M., Valla, D.-C. & Roudot-Thoraval, F. The burden of liver disease in Europe: a review of available epidemiological data. J. Hepatol. 58, 593–608. https://doi.org/10.1016/j.jhep.2012.12.005 (2013).
Cerquetella, M. et al. Chronic hepatitis in man and in dog: a comparative update. Rev. Esp. Enferm. Dig. 104, 203–209. https://doi.org/10.4321/S1130-01082012000400006 (2012).
Decaro, N. et al. Infectious canine hepatitis: an “old” disease reemerging in Italy. Res. Vet. Sci. 83, 269–273. https://doi.org/10.1016/j.rvsc.2006.11.009 (2007).
van den Ingh, T. S. G. A. M., van Winkle, T., Cullen, J. M., Charles, J. A. & Desmet, V. J. Morphological classification of parenchymal disorders of the canine and feline liver: 2. Hepatocellular death, hepatitis and cirrhosis. In WSAVA Standards for Clinical and Histological Diagnosis of Canine and Feline Liver Diseases 85–101 (Saunders Elsevier, 2006).
Webb, C. B. Canine inflammatory/Infectious hepatic disease. In Textbook of Veterinary Internal Medicine: Diseases of the Dog and the Cat 8th edn (eds Ettinger, S. J. et al.) 1628–1633 (Elsevier Saunders, 2017).
Dunn, W. B., Broadhurst, D. I., Atherton, H. J., Goodacre, R. & Griffin, J. L. Systems level studies of mammalian metabolomes: the roles of mass spectrometry and nuclear magnetic resonance spectroscopy. Chem. Soc. Rev. 40, 387–426. https://doi.org/10.1039/b906712b (2011).
Hollywood, K., Brison, D. R. & Goodacre, R. Metabolomics: current technologies and future trends. Proteomics 6, 4716–4723. https://doi.org/10.1002/pmic.200600106 (2006).
Yu, M., Zhu, Y., Cong, Q. & Wu, C. Metabonomics research progress on liver diseases. Can. J. Gastroenterol. Hepatol. 2017, 8467192. https://doi.org/10.1155/2017/8467192 (2017).
Amathieu, R. et al. Nuclear magnetic resonance based metabolomics and liver diseases: recent advances and future clinical applications. World J. Gastroenterol. 22, 417–426. https://doi.org/10.3748/wjg.v22.i1.417 (2016).
Carlos, G., Dos Santos, F. P. & Fröehlich, P. E. Canine metabolomics advances. Metabolomics 16, 16. https://doi.org/10.1007/s11306-020-1638-7 (2020).
Tran, H., McConville, M. & Loukopoulos, P. Metabolomics in the study of spontaneous animal diseases. J. Vet. Diagn. Invest. 32, 635–647. https://doi.org/10.1177/1040638720948505 (2020).
Lawrence, Y. A. et al. Untargeted metabolomic profiling of serum from dogs with chronic hepatic disease. J. Vet. Intern. Med. 33, 1344–1352. https://doi.org/10.1111/jvim.15479 (2019).
Whitfield, P. D. et al. Metabolomics as a diagnostic tool for hepatology: validation in a naturally occurring canine model. Metabolomics 1, 215–225. https://doi.org/10.1007/s11306-005-0001-3 (2005).
Devriendt, N. et al. Plasma amino acid profiles in dogs with closed extrahepatic portosystemic shunts are only partially improved 3 months after successful gradual attenuation. J. Vet. Intern. Med. 35, 1347–1354. https://doi.org/10.1111/jvim.16135 (2021).
Harper, E. J., Hackett, R. M., Wilkinson, J. & Heaton, P. R. Age-related variations in hematologic and plasma biochemical test results in Beagles and Labrador Retrievers. J. Am. Vet. Med. Assoc. 223, 1436–1442. https://doi.org/10.2460/javma.2003.223.1436 (2003).
Center, S. A. Interpretation of liver enzymes. Vet. Clin. N. Am. Small Anim. Pract. 37, 297–333. https://doi.org/10.1016/j.cvsm.2006.11.009 (2007).
Ottka, C., Vapalahti, K., Puurunen, J., Vahtera, L. & Lohi, H. A novel canine nuclear magnetic resonance spectroscopy-based metabolomics platform: validation and sample handling. Vet. Clin. Pathol. 50, 410–426. https://doi.org/10.1111/vcp.12954 (2021).
Soininen, P., Kangas, A. J., Würtz, P., Suna, T. & Ala-Korpela, M. Quantitative serum nuclear magnetic resonance metabolomics in cardiovascular epidemiology and genetics. Circ. Cardiovasc. Genet. 8, 192–206. https://doi.org/10.1161/CIRCGENETICS.114.000216 (2015).
Würtz, P. et al. Quantitative serum nuclear magnetic resonance metabolomics in large-scale epidemiology: a primer on -omic technologies. Am. J. Epidemiol. 186, 1084–1096. https://doi.org/10.1093/aje/kwx016 (2017).
Soininen, P. et al. High-throughput serum NMR metabonomics for cost-effective holistic studies on systemic metabolism. Analyst. 134, 1781–1785. https://doi.org/10.1039/b910205a (2009).
Kruskal, W. H. & Wallis, W. A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 47, 583–621. https://doi.org/10.1080/01621459.1952.10483441 (1952).
Mann, H. B. & Whitney, D. R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Stat. 18, 50–60. https://doi.org/10.1214/aoms/1177730491 (1947).
Dunn, O. J. Multiple comparisons among means. J. Am. Stat. Assoc. 56, 52–64. https://doi.org/10.1080/01621459.1961.10482090 (1961).
MetaboAnalyst. https://www.metaboanalyst.ca/ (2021).
Rokach, L. & Maimon, O. Clustering methods. In Data Mining and Knowledge Discovery Handbook (eds Maimon, O. Z. & Rokach, L.) 321–352 (Springer, 2005).
Witten, I. H., Frank, E., Hall, M. A. & Pal, C. J. Data Mining. Practical Machine Learning Tools and Techniques. The WEKA Workbench. Online Appendix. 4th ed. (Elsevier Reference Monographs, 2011).
Sumner, M., Frank, E. & Hall, M. Speeding up logistic model tree induction. In Knowledge Discovery in Databases: PKDD 2005. 9th European Conference on Principles and Practice of Knowledge Discovery in Databases Vol. 3721 (eds Hutchison, D. et al.) 675–683 (Springer, 2005).
Crammer, K. & Singer, Y. On the algorithmic implementation of multiclass kernel-based vector machines. J. Mach. Learn. Res. 2, 265–292 (2002).
Aha, D. W., Kibler, D. & Albert, M. K. Instance-based learning algorithms. Mach. Learn. 6, 37–66. https://doi.org/10.1007/BF00153759 (1991).
Morariu, D., Crețulescu, R. & Breazu, M. The WEKA multilayer perceptron classifier. J. Adv. Stat. IT&C Econom. Life Sci. 7 (2017).
Breiman, L. Random forests. Mach. Learn. 45, 5–32. https://doi.org/10.1023/A:1010933404324 (2001).
Hand, D. J. & Yu, K. Idiot’s Bayes: not so stupid after all?. Int. Stat. Rev. 69, 385–398. https://doi.org/10.2307/1403452 (2001).
Hotelling, H. Analysis of a complex of statistical variables into principal components. J. Educ. Psychol. 24, 417–441. https://doi.org/10.1037/h0071325 (1933).
Squillacciotti, S. Prediction oriented classification in PLS path modeling. In Handbook of Partial Least Squares (eds Esposito Vinzi, V. et al.) 219–233 (Springer, 2010).
Bijlsma, S. et al. Large-scale human metabolomics studies: a strategy for data (pre-) processing and validation. Anal. Chem. 78, 567–574. https://doi.org/10.1021/ac051495j (2006).
Masarone, M. et al. Untargeted metabolomics as a diagnostic tool in NAFLD: discrimination of steatosis, steatohepatitis and cirrhosis. Metabolomics 17, 12. https://doi.org/10.1007/s11306-020-01756-1 (2021).
Aguirre, A., Yoshimura, N., Westman, T. & Fischer, J. E. Plasma amino acids in dogs with two experimental forms of liver damage. J. Surg. Res. 16, 339–345. https://doi.org/10.1016/0022-4804(74)90053-5 (1974).
Smith, A. R. et al. Alterations in plasma and CSF amino acids, amines and metabolites in hepatic coma. Ann. Surg. 187, 343–350. https://doi.org/10.1097/00000658-197803000-00024 (1978).
Neumann, S., Welling, H. & Thuere, S. Evaluation of serum L-phenylalanine concentration as indicator of liver disease in dogs: a pilot study. J. Am. Anim. Hosp. Assoc. 43, 193–200. https://doi.org/10.5326/0430193 (2007).
Fischer, J. E. et al. Plasma amino acids in patients with hepatic encephalopathy. Effects of amino acid infusions. Am. J. Surg. 127, 40–47. https://doi.org/10.1016/0002-9610(74)90009-9 (1974).
Morgan, M. Y., Milsom, J. P. & Sherlock, S. Plasma ratio of valine, leucine and isoleucine to phenylalanine and tyrosine in liver disease. Gut 19, 1068–1073. https://doi.org/10.1136/gut.19.11.1068 (1978).
Dam, G. et al. Muscle metabolism and whole blood amino acid profile in patients with liver disease. Scand. J. Clin. Lab. Invest. 75, 674–680 (2015).
Morgan, M. Y., Marshall, A. W., Milsom, J. P. & Sherlock, S. Plasma amino-acid patterns in liver disease. Gut 23, 362–370. https://doi.org/10.1136/gut.23.5.362 (1982).
Yamato, M., Muto, Y., Yoshida, T., Kato, M. & Moriwaki, H. Clearance rate of plasma branched-chain amino acids correlates significantly with blood ammonia level in patients with liver cirrhosis. Int. Hepatol. Commun. 3, 91–96. https://doi.org/10.1016/0928-4346(94)00159-3 (1995).
Levine, R. J. & Conn, H. O. Tyrosine metabolism in patients with liver disease. J. Clin. Invest. 46, 2012–2020. https://doi.org/10.1172/JCI105690 (1967).
Holeček, M. Evidence of a vicious cycle in glutamine synthesis and breakdown in pathogenesis of hepatic encephalopathy-therapeutic perspectives. Metab. Brain Dis. 29, 9–17. https://doi.org/10.1007/s11011-013-9428-9 (2014).
Fischer, J. & Baldessarini, R. False neurotransmitters and hepatic failure. Lancet 298, 75–80. https://doi.org/10.1016/s0140-6736(71)92048-4 (1971).
Butterworth, J., Gregory, C. R. & Aronson, L. R. Selective alterations of cerebrospinal fluid amino acids in dogs with congenital portosystemic shunts. Metab. Brain Dis. 12, 299–306. https://doi.org/10.1007/BF02674674 (1997).
Katayama, K. Zinc and protein metabolism in chronic liver diseases. Nutr. Res. 74, 1–9. https://doi.org/10.1016/j.nutres.2019.11.009 (2020).
Jeppesen, J. B., Mortensen, C., Bendtsen, F. & Møller, S. Lactate metabolism in chronic liver disease. Scand. J. Clin. Lab. Invest. 73, 293–299. https://doi.org/10.3109/00365513.2013.773591 (2013).
Record, C. O., Iles, R. A., Cohen, R. D. & Williams, R. Acid-base and metabolic disturbances in fulminant hepatic failure. Gut 16, 144–149. https://doi.org/10.1136/gut.16.2.144 (1975).
Bihari, D., Gimson, A. E., Lindridge, J. & Williams, R. Lactic acidosis in fulminant hepatic failure. Some aspects of pathogenesis and prognosis. J. Hepatol. 1, 405–416. https://doi.org/10.1016/S0168-8278(85)80778-9 (1985).
Clemmesen, J. O., Høy, C.-E., Kondrup, J. & Ott, P. Splanchnic metabolism of fuel substrates in acute liver failure. J. Hepatol. 33, 941–948. https://doi.org/10.1016/s0168-8278(00)80126-9 (2000).
Murphy, N. D. et al. Liver and intestinal lactate metabolism in patients with acute hepatic failure undergoing liver transplantation. Crit. Care Med. 29, 2111–2118. https://doi.org/10.1097/00003246-200111000-00011 (2001).
van de Wier, B. et al. Elevated citrate levels in non-alcoholic fatty liver disease: the potential of citrate to promote radical production. FEBS Lett. 587, 2461–2466. https://doi.org/10.1016/j.febslet.2013.06.019 (2013).
Sands, C. J. et al. Metabolic phenotyping for enhanced mechanistic stratification of chronic hepatitis C-induced liver fibrosis. Am. J. Gastroenterol. 110, 159–169. https://doi.org/10.1038/ajg.2014.370 (2015).
Kramer, L. et al. Citrate pharmacokinetics and metabolism in cirrhotic and noncirrhotic critically ill patients. Crit. Care Med. 31, 2450–2455. https://doi.org/10.1097/01.CCM.0000084871.76568.E6 (2003).
Yamashita, H., Kaneyuki, T. & Tagawa, K. Production of acetate in the liver and its utilization in peripheral tissues. Biochim. Biophys. Acta 1532, 79–87. https://doi.org/10.1016/s1388-1981(01)00117-2 (2001).
Haverstick, D. M. & Groszbach, A. R. Specimen collection and processing. In Tietz Textbook of Clinical Chemistry and Molecular Diagnostics 5th edn (eds Burtis, C. A. & Ashwood Edward, R.) 145–162 (Saunders Elsevier, 2013).
Gao, R. et al. Serum metabolomics to identify the liver disease-specific biomarkers for the progression of hepatitis to hepatocellular carcinoma. Sci. Rep. 5, 18175. https://doi.org/10.1038/srep18175 (2015).
Ylikahri, R. Metabolic and nutritional aspects of xylitol. In: Advances in Food Research, Vol. 25. 159–180 (Elsevier, 1979).
Assawarachan, S. N., Chuchalermporn, P., Maneesaay, P. & Thengchaisri, N. Changes in serum lipid profiles among canine patients suffering from chronic hepatitis. Vet. Sci. 8, 221. https://doi.org/10.3390/vetsci8100221 (2021).
Harrison, J. L., Turek, B. J., Brown, D. C., Bradley, C. & Callahan Clark, J. Cholangitis and cholangiohepatitis in dogs: a descriptive study of 54 cases based on histopathologic diagnosis (2004–2014). J. Vet. Intern. Med. 32, 172–180. https://doi.org/10.1111/jvim.14866 (2018).
Lee, S., Kweon, O.-K. & Kim, W. H. Associations between serum leptin levels, hyperlipidemia, and cholelithiasis in dogs. PLoS ONE 12, e0187315. https://doi.org/10.1371/journal.pone.0187315 (2017).
Watson, T. D. G. & Barrie, J. Lipoprotein metabolism and hyperlipidaemia in the dog and cat: a review. J. Small Anim. Pract. 34, 479–487. https://doi.org/10.1111/j.1748-5827.1993.tb03519.x (1993).
Arranz, M. I. et al. Fatty acid composition of lipoprotein lipids in hepatobiliary diseases. Eur. J. Clin. Chem. Clin. Biochem. 34, 701–709. https://doi.org/10.1515/cclm.1996.34.9.701 (1996).
Habib, A. et al. High-density lipoprotein cholesterol as an indicator of liver function and prognosis in noncholestatic cirrhotics. Clin. Gastroenterol. Hepatol. 3, 286–291. https://doi.org/10.1016/s1542-3565(04)00622-6 (2005).
Cheng, R. et al. High-density lipoprotein cholesterol is a predictor of survival in cirrhotic patients with acute gastrointestinal bleeding: a retrospective study. BMC Gastroenterol. 20, 381. https://doi.org/10.1186/s12876-020-01522-6 (2020).
Trieb, M. et al. HDL-related biomarkers are robust predictors of survival in patients with chronic liver failure. J. Hepatol. 73, 113–120. https://doi.org/10.1016/j.jhep.2020.01.026 (2020).
Amathieu, R. et al. Metabolomic approach by 1H NMR spectroscopy of serum for the assessment of chronic liver failure in patients with cirrhosis. J. Proteome Res. 10, 3239–3245. https://doi.org/10.1021/pr200265z (2011).
Caren, R. & Corbo, L. The origin of plasma arachidonic acid in dogs. Metabolism 17, 1043–1050. https://doi.org/10.1016/0026-0495(68)90012-7 (1968).
Abdel-Ghaffar, T. Y., Amin, M., Abdel Rasheed, M. & Fouad, H. H. Essential fatty acids status in infants and children with chronic liver disease. J. Clin. Biochem. Nutr. 33, 1–6. https://doi.org/10.3164/jcbn.33.1 (2003).
Cabré, E. & Gassull, M. A. Polyunsaturated fatty acid deficiency in liver diseases: pathophysiological and clinical significance. Nutrition 12, 542–548. https://doi.org/10.1016/s0899-9007(96)00122-0 (1996).
Otvos, J. D. et al. GlycA: a composite nuclear magnetic resonance biomarker of systemic inflammation. Clin. Chem. 61, 714–723. https://doi.org/10.1373/clinchem.2014.232918 (2015).
Bell, J. D., Brown, J., Nicholson, J. K. & Sadler, P. J. Assignment of resonances for ‘acute-phase’ glycoproteins in high resolution proton NMR spectra of human blood plasma. FEBS Lett. 215, 311–315. https://doi.org/10.1016/0014-5793(87)80168-0 (1987).
Gruppen, E. G., Connelly, M. A., Otvos, J. D., Bakker, S. J. L. & Dullaart, R. P. F. A novel protein glycan biomarker and LCAT activity in metabolic syndrome. Eur. J. Clin. Invest. 45, 850–859. https://doi.org/10.1111/eci.12481 (2015).
Dullaart, R. P. F., Gruppen, E. G., Connelly, M. A., Otvos, J. D. & Lefrandt, J. D. GlycA, a biomarker of inflammatory glycoproteins, is more closely related to the leptin/adiponectin ratio than to glucose tolerance status. Clin. Biochem. 48, 811–814. https://doi.org/10.1016/j.clinbiochem.2015.05.001 (2015).
Dungan, K., Binkley, P. & Osei, K. GlycA is a novel marker of inflammation among non-critically ill hospitalized patients with type 2 diabetes. Inflammation 38, 1357–1363. https://doi.org/10.1007/s10753-014-0107-8 (2015).
Ormseth, M. J. et al. Utility of a novel inflammatory marker, GlycA, for assessment of rheumatoid arthritis disease activity and coronary atherosclerosis. Arthritis Res. Ther. 17, 117. https://doi.org/10.1186/s13075-015-0646-x (2015).
Levine, J. A. et al. Associations of GlycA and high-sensitivity C-reactive protein with measures of lipolysis in adults with obesity. J. Clin. Lipidol. 14, 667–674. https://doi.org/10.1016/j.jacl.2020.07.012 (2020).
Gruppen, E. G. et al. GlycA, a pro-Inflammatory glycoprotein biomarker, and incident cardiovascular disease: relationship with c-reactive protein and renal function. PLoS ONE 10, e0139057. https://doi.org/10.1371/journal.pone.0139057 (2015).
Connelly, M. A. et al. GlycA, a marker of acute phase glycoproteins, and the risk of incident type 2 diabetes mellitus: PREVEND study. Clin. Chim. Acta 452, 10–17. https://doi.org/10.1016/j.cca.2015.11.001 (2016).
Fuertes-Martín, R., Correig, X., Vallvé, J.-C. & Amigó, N. Human serum/plasma glycoprotein analysis by 1H-NMR, an emerging method of inflammatory assessment. J. Clin. Med. 9, 354. https://doi.org/10.3390/jcm9020354 (2020).
Ottka, C., Weber, C., Müller, E. & Lohi, H. Serum NMR metabolomics uncovers multiple metabolic changes in phenobarbital-treated dogs. Metabolomics 17, 54. https://doi.org/10.1007/s11306-021-01803-5 (2021).
Yuki, M., Itoh, H. & Takase, K. Serum alpha-1-acid glycoprotein concentration in clinically healthy puppies and adult dogs and in dogs with various diseases. Vet. Clin. Pathol. 39, 65–71. https://doi.org/10.1111/j.1939-165X.2009.00181.x (2010).
Andersson, M., Stenström, M., Vatne, M., Sevelius, E. & Jönsson, L. Disease-related variations of the glycosylation of haptoglobin in the dog. J. Comp. Pathol. 119, 227–238. https://doi.org/10.1016/S0021-9975(98)80046-7 (1998).
Crawford, K. et al. Serum haptoglobin concentrations in dogs with liver disease. Vet. Rec. 173, 579. https://doi.org/10.1136/vr.101306 (2013).
Berent, A. C. & Tobias, K. M. Portosystemic vascular anomalies. Vet. Clin. N. Am. Small Anim. Pract. 39, 513–541. https://doi.org/10.1016/j.cvsm.2009.02.004 (2009).
Acknowledgements
We thank the veterinarians who submitted samples to Laboklin GmbH & Co. KG, thereby enabling the use of left-over blood samples for scientific purposes. We further acknowledge the Laboklin teams in Germany and Czech Republic for their assistance in obtaining medical information. We further acknowledge Dr. Leslie King for careful scientific editing of the manuscript.
Funding
Laboklin GmbH & Co. KG enabled the collection of left-over blood samples and routine blood test analyses. Metabolomic NMR analyses were provided by PetMeta Labs Oy. Bioinformatic analysis was performed independent of Laboklin and PetMeta Labs and were supported by intramural funds within the Institute for Chemistry and Bioanalytics, School of Life Sciences, University of Applied Sciences Northwestern Switzerland, Muttenz, Switzerland.
Author information
Authors and Affiliations
Contributions
Conceptualization of study by U.G., C.A.I., C.O., E.M., H.L.; serum sample collection and gathering of all clinicopathological information by C.A.I. and C.W.; transferring NMR data by C.O.; data and statistical analyses by C.A.I., F.D., and U.G.; writing manuscript by C.A.I. and U.G.; reviewing and editing manuscript by F.D., G.S., C.O., H.L., C.W., and E.M.; E.M. and H.L. secured funding. All authors have read and agreed to publish the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
This study was conducted as part of Carolin A. Imbery’s doctoral thesis at the University of Zürich, Switzerland. Carolin A. Imbery and Corinna Weber are employed, and Elisabeth Müller is the owner and executive director of Laboklin GmbH & Co. KG, Bad Kissingen, Germany. Laboklin offers a variety of diagnostic laboratory tests for animals. Claudia Ottka is employed, and Hannes Lohi is the board director of PetMeta Labs Oy, Helsinki, Finland, which provides NMR metabolomic testing for dogs. Urs Giger has been a scientific advisor to Laboklin. Frank Dieterle is the CEO of Dieterle Life Sciences Consulting, Binningen, Switzerland. Götz Schlotterbeck declares no potential conflict of interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Imbery, C.A., Dieterle, F., Ottka, C. et al. Metabolomic serum abnormalities in dogs with hepatopathies. Sci Rep 12, 5329 (2022). https://doi.org/10.1038/s41598-022-09056-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-09056-5
- Springer Nature Limited
This article is cited by
-
Changes in free amino acid profile in goats with pregnancy toxemia
Veterinary Research Communications (2023)
-
Metabolic changes associated with two endocrine abnormalities in dogs: elevated fructosamine and low thyroxine
Metabolomics (2022)