
Abstract
Dietary restriction is a frequent strategy for weight loss, but adherence is difficult and returning to poor dietary habits can result in more weight gain than that previously lost. How weight loss due to unrestricted intake of a healthy diet affects the response to resumption of poor dietary habits is less studied. Moreover, whether this response differs between the sexes and if the insulin-like growth factor (IGF) system, sex dependent and involved in metabolic control, participates is unknown. Mice received rodent chow (6% Kcal from fat) or a high-fat diet (HFD, 62% Kcal from fat) for 4 months, chow for 3 months plus 1 month of HFD, or HFD for 2 months, chow for 1 month then HFD for 1 month. Males and females gained weight on HFD and lost weight when returned to chow at different rates (p < 0.001), but weight gain after resumption of HFD intake was not affected by previous weight loss in either sex. Glucose metabolism was more affected by HFD, as well as the re-exposure to HFD after weight loss, in males. This was associated with increases in hypothalamic mRNA levels of IGF2 (p < 0.01) and IGF binding protein (IGFBP) 2 (p < 0.05), factors involved in glucose metabolism, again only in males. Likewise, IGF2 increased IGFBP2 mRNA levels only in hypothalamic astrocytes from males (p < 0.05). In conclusion, the metabolic responses to dietary changes were less severe and more delayed in females and the IGF system might be involved in some of the sex specific observations.
Similar content being viewed by others


Introduction
Obesity is in part due to poor dietary habits, including increased consumption of diets rich in fat. Caloric restriction is one of the most frequent methods for weight loss; however, there is often a “rebound effect”, or an increased weight gain once unhealthy dietary habits are resumed1,2. This weight gain may be due to impairment of the neural circuitry that controls metabolism and body weight3. Other factors reported to contribute to this phenomenon include changes in adipocyte metabolism4, alterations in gut hormone levels, and a decrease in the resting energy expenditure rate or modifications in the reward system5. Improved dietary habits that include exclusion of foods rich in saturated fats and an increased intake of healthy foods, for example the Mediterranean diet, have been shown to improve weight loss and overall health even better than restrictive diets6,7. However, how weight loss due to this type of regimen affects the metabolic response to resumption of poor dietary habits is not known.
The insulin-like growth factor (IGF) system participates in a myriad of functions in the organism8,9,10,11, including metabolism12,13,14 as the two ligands of this system, IGF1 and IGF2, are anabolic hormones15 due to their structural similarity with insulin and sharing its hypoglycemic effects16. The main source of circulating IGFs is the liver17, and although they can cross the blood–brain barrier18, they are also produced centrally by neurons19,20, astrocytes21 and other glial cells21,22. Likewise, the six IGF-binding proteins (IGFBPs) that bind IGFs modifying their half-live, distribution and functions23 are also produced centrally, with IGFBP2 being one of the most prominently produced.
Sex differences in the IGF system24,25,26 are involved in the differences in growth27, and possibly metabolic responses, of males and females. In both humans and mice, females appear to be more protected against diet-induced obesity and its comorbidities than males, at least during younger ages28,29,30, with sex steroids participating in this protection31,32,33. Nutritional status alters the circulating IGF system in humans34,35,36 and in mice both systemically and centrally37 in a time- and sex-dependent manner, with some parameters normalizing after weight loss in children with obesity38. We recently reported that after 8 weeks on a high fat diet (HFD) circulating IGF2 levels are higher in mice of both sexes37, but after 12 weeks they remained elevated only in females, whereas IGFBP2 levels were decreased in males39. Hypothalamic IGF2 expression is also regulated by diet37,40 and both IGF2 and IGFBP2 are reportedly involved in glucose metabolism41,42,43, as is central44 and peripheral45,46 IGF1. Thus, the IGF system could participate in the differences between males and females in their responses to dietary change and weight gain.
The hypothalamus is responsible for the integration of metabolic signals and the regulation of feeding behavior47,48, with astrocytes in this brain area actively participating in metabolic control mechanisms49, including glucose and lipid sensing50. Moreover, astrocytes are an important source of IGF1 in the brain21,22,51; thus, part of their metabolic actions could involve production of IGFs and possibly other members of this family such as IGFBP2.
The aims of this study were to evaluate the potential sex differences, both centrally and peripherally, in the metabolic susceptibility to resumption of a HFD after weight loss in mice and the possible implications of the IGF system. We hypothesized that male and female mice have different metabolic responses to dietary changes, with the central and circulating IGF systems also being differentially regulated.
Results
Body weight and fat mass
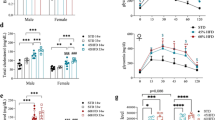
The significant results of the statistical analysis of body weight are shown in Supplementary Table 1. Body weight throughout the study (Fig. 1) changed over time and was modulated by sex and the diet, with an interaction between these factors.

Body weight throughout the study in male and female mice exposed to chow, a high-fat diet (HFD) for 4 months (HHHH), a chow diet for 3 months and an additional month on HFD (CCCH), or HFD for 2 months followed by chow for 1 month and then HFD for the last month (HHCH). #, Different from chow in the same sex; $, different from HFD in the same sex; &, different from CCCH in the same sex; @, different between sexes on the same diet. n = 9.
Body weight was greater in males than females at all study points regardless of dietary regimen. Body weight was affected by sex and diet from week 1 to week 12. All groups on the HFD weighed more than those on chow, with this reaching significance in males from week 1 (F(3,35) = 10.5, p < 0.001) onward and in females after 4 weeks (F(3,35) = 5.4, p < 0.01).When HHCH mice were switched from HFD to chow, males required one week (F(3,34) = 32.7, p < 0.001) and females two weeks (F(3,35) = 11.6, p < 0.001) for those switched to chow to weigh less than those continuing on HFD. Males also responded more rapidly to the re-exposition to HFD in the HHCH group than females, requiring one week (F(3,34) = 20.1, p < 0.001) to achieve significant weight gain, while females required 2 weeks in the HHCH (F(3,35) = 12.4, p < 0.001) and 3 weeks in CCCH (F(3,35) = 12.6, p < 0.001) groups. Moreover, HHCH mice reached the body weight of the HHHH mice at week 14, whereas CCCH mice reached them at week 15 in both males and females.
The significant results of the statistical analysis of weight gain at each point of dietary change and final fat mass are shown in Supplementary Table 2. The percent change in body weight from baseline to the end of month 2 (Fig. 2A) was influenced by sex and diet, with an interaction between these factors. The HFD induced weight gain in both males (F(3,35) = 37.1, p < 0.001) and females (F(3,35) = 13.1, p < 0.001), but this weight gain was greater in males (p < 0.001). The percent weight gain from the end of month 2 to the end of month 3 (Fig. 2B) was influenced by sex and diet, with an interaction between these factors. HHCH mice of both sexes lost weight on the chow diet (M: F(3,35) = 135.9, p < 0.001, F: F(3,35) = 46.2, p < 0.001) and this decrease was greater in males than females (F(1,17) = 12.3, p < 0.01). From the end of month 2 to the end of month 3, HHHH females continued to gain more weight than chow mice, while HHHH males did not (Fig. 2C). During the last month of the study, both sex and diet affected weight gain. The percent weight gain from the end of month 3 to the end of the month 4 (Fig. 2D) did not differ between CCCH and HHCH mice, although they gained relatively more weight than the chow and HHHH mice (males: F(3,34) = 117.8, p < 0.001; females: F(3,35) = 32.4, p < 0.001). During the last month of the study, HHHH females gained relatively more weight than males of the same group.

Percent weight gain compared to baseline to the end of month 2 (A), weight gain from the end of month 2 to the end of month 3 (B), weight gain from baseline to the end of month 3 (C), weight gain from the end of month 3 to the end of month 4 (D), percent total weight gain from baseline to the end of the study (E) and final body weight (F), as well as the percentage of retroperitoneal adipose tissue (G) and inguinal adipose tissue (H) in male and female mice fed with chow or a high-fat diet (HFD) for 4 months (HHHH), a chow diet for 3 months and an additional month on a HFD (CCCH), or HFD for 2 months followed by chow for 1 month and then HFD for the last month (HHCH). ***p < 0.001. a: effect of the sex, b: effect of the diet. n = 9.
Total weight gain relative to baseline (Fig. 2E) and final body weight (Fig. 2F) were dependent on sex, being higher in males than females, and diet, with an overall increase in the HHHH, CCCH and HHCH groups, with no apparent differences between the regimens of HFD intake.
The relative amounts of retroperitoneal adipose tissue (Fig. 2G) and inguinal adipose tissue (Fig. 2H) were similarly affected by the dietary regimens, with an overall increase in all groups that consumed HFD.
Energy intake
The significant results of the statistical analysis of weekly energy intake are shown in Supplementary Table 3 and although the statistical analysis was done simultaneously on all groups, for clarity data for males (Fig. 3A) and females (Fig. 3B) are represented separately.

Weekly energy intake throughout the study in male (A) and female (B) mice ingesting chow or a high-fat diet (HFD) for 4 months (HHHH), chow for 3 months and an additional month on HFD (CCCH), or HFD for 2 months followed by chow for 1 month and then HFD for the last month (HHCH). #, Different from chow in the same sex; $, different from HFD in the same sex; &, different from CCCH in the same sex. n = 3 (number of cages per group).
Energy intake was altered by time and diet from week 1 to week 15, with an interaction between time and diet. No effect of sex was found, possibly due to the high variability in female groups consuming HFD. When males were first exposed to the HFD there was an increase in energy intake (F(3,11) = 28.4, p < 0.001 and F(3,11) = 24.6, p < 0.001) that returned to control levels after two weeks and then fluctuated throughout the study. When HHCH mice returned to a chow diet, energy intake was rapidly reduced (F(3,11) = 14.4, p = 0.001) and then returned to control levels. When HHCH mice were again exposed to a HFD at the end of month 3, there was no difference in their energy intake from the CCCH group that received HFD for the first time. In females, when HHCH returned to a chow diet they also rapidly reduced energy intake (F(3,11) = 4.2, p < 0.05) that normalized during the following month. The increase in energy intake during the last month of the study did not differ between the HHCH and CCCH groups.
The global energy intake parameters are shown in Table 1. There was no difference in the total number of Kcal consumed per mouse. When energy consumption was adjusted for body weight, females consumed more Kcal than males, with no dietary effects. From baseline to the end of month 2, energy intake was higher in mice consuming HFD, while from the end of month 2 to the end of month 3 the mice switched from HFD to chow ingested fewer Kcal compared to the rest of the groups. From the end of month 3 to the end of month 4 there was an overall increase in the CCCH and HHCH groups. Mean energy efficiency (during the entire study), represented as the weight gained in grams per calories consumed, was higher in males and increased in all groups that consumed HFD in both sexes.
Glucose metabolism
Time affected glycemia levels during the GTT (Fig. 4A), with interactions between time and sex, time and diet, sex and diet, and time, sex and diet (see Supplementary Table 4 for statistical analysis).

Glycemia during the glucose tolerance test (GTT; A), the area under the curve (AUC; B) in the GTT, peripheral insulin levels (C) and glycemia at sacrifice (D) and Homeostatic Model Assessment for Insulin Resistance (HOMA-IR; E) in male and female mice fed with chow or a high-fat diet (HFD) for 4 months (HHHH), chow diet for 3 months and an additional month on a HFD (CCCH), or HFD for 2 months followed by chow for 1 month and then HFD for the last month (HHCH). ***p < 0.001; a: effect of the sex, b: effect of the diet. #, Different from chow; @, differences between sexes on the same diet. n = 6.
Consuming HFD, regardless of the regimen, affected glycemia in a sex and time-dependent manner. Basal glycemia was higher in CCCH and HHCH mice compared to the chow group in both sexes (M: F(3,22) = 8.4, p = 0.001; F: F(3,23) = 4.0, p < 0.05), whereas in HHHH mice it was only elevated in males. Glycemia was higher in male HHHH (F(1,10) = 16.2, p < 0.01), CCCH (F(1,11) = 10.5, p < 0.01) and HHCH (F(1,11) = 5.0, p < 0.05) compared to females of the same groups. At 30 min all groups had an increase in glycemia, whereas at 60 min glycemia returned to baseline in chow fed mice, remaining elevated in the HHHH, CCCH and HHCH groups, with this elevation being higher in males than females. At 90 min glycemia was elevated in HHHH, CCCH and HHCH males (F(3,22) = 18.9, p < 0.001), whereas it was only elevated in HHHH females (F(3,23) = 4.2, p < 0.05). At this time glycemia was higher in CCCH (F(1,11) = 6.7, p < 0.05) and HHCH (F(1,11) = 15.3, p < 0.01) males compared to females of the same groups. At 120 min glycemia remained elevated in HHHH, CCCH and HHCH males (F(3,22) = 25.4, p < 0.001), whereas in females it was only elevated in the HHHH group (F(3,23) = 4.1, p < 0.05), with CCCH and HHCH males continuing to have higher glycemia than females on the same diet regimen (F(1,11) = 16.0, p < 0.01 and F(1,11) = 31.3, p < 0.001, respectively). Mice in the HHHH, CCCH or HHCH groups had a higher AUC (Fig. 4B) than those consuming chow in both males (F(3,22) = 46.5, p < 0.001) and females (F(3,23) = 6.1, p < 0.01). Males had a higher AUC than females on the CCCH (F(1,11) = 10.0, p = 0.01) and HHCH (F(1,11) = 13.3, p < 0.01) dietary regimens.
At the end of the study, circulating insulin levels (Fig. 4C) were higher in males than females, and increased by HFD intake regardless of regimen. Glycemia levels at sacrifice (Fig. 4D) and the HOMA index (Fig. 4E), which was increased by HFD consumption, were also higher in males than females.
Plasma levels of leptin and members of the IGF system
Significant results of the statistical analysis of circulating levels of members of the IGF system and leptin are shown in Supplementary Table 5. Circulating free IGF1 levels (Fig. 5A) were not altered by sex or diet, while there was an overall effect of HFD to increase total IGF1 levels (Fig. 5B). Females had higher circulating IGF2 levels (Fig. 5C) than males, with no effect of diet. IGFBP2 levels (Fig. 5D) were unaffected by sex or diet. Plasma leptin levels (Fig. 5E) were increased by HFD intake (M: F(3,34) = 13.8, p < 0.001; F: F(3,34) = 7.4, p = 0.001) with males having higher levels than females in CCCH (F(1,17) = 8.9, p < 0.01) and HHCH (F(1,17) = 11.5, p < 0.01) mice.

Circulating levels of free insulin-like growth factor (IGF) 1 (A), total IGF1 (B), IGF2 (C), IGF binding protein (IGFBP) 2 (D) and leptin (E) in mice fed chow or a high-fat diet (HFD) for 4 months (HHHH), chow diet for 3 months and an additional month on a HFD (CCCH), or HFD for 2 months followed by chow for 1 month and then HFD for the last month (HHCH). ***p < 0.001. a: effect of the sex, b: effect of the diet. NS non-significant. n = 9.
The hypothalamic IGF system
Relative hypothalamic IGF1 mRNA levels (Fig. 6A, statistics Supplementary Table 5) were higher in females than males and decreased in CCCH mice of both sexes. IGF2 mRNA levels (Fig. 6B) were also higher in females than males on a chow diet (F(1,10) = 5.9, p < 0.05). The response to the diet differed between the sexes with male CCCH mice having higher IGF2 mRNA levels than chow and HHHH mice (F(3,22) = 3.0, p = 0.05). A similar effect was found on IGFBP2 mRNA levels (Fig. 6C), with chow females having higher IGFBP2 mRNA levels than males (F(1,10) = 7.4, p < 0.05) and with this binding protein increasing in CCCH males (F(3,22) = 3.3, p < 0.05). As previously shown24,37,39, relative hypothalamic mRNA levels of IGF2 and IGFBP2 were positively correlated (r = 0.933, p < 0.001; Fig. 6D).

Relative mRNA levels of insulin-like growth factor (IGF) 1 (A), IGF2 (B), IGF binding protein (IGFBP)2 (C), neuropeptide Y (NPY; E), Agouti-related protein (AgRP; F), and proopiomelanocortin (POMC; G) in the hypothalamus, as well as the correlation of hypothalamic IGF2 and IGFBP2 mRNA levels (D), in mice fed chow or a high-fat diet (HFD) for 4 months (HHHH), a chow diet for 3 months and an additional month on a HFD (CCCH), or HFD for 2 months followed by chow for 1 month and then HFD for the last month (HHCH). *p < 0.05, ***p < 0.001. a: effect of the sex, b: effect of the diet. n = 6.
Hypothalamic metabolic neuropeptides
Hypothalamic NPY mRNA levels (Fig. 6E, statistics Supplementary Table 5) were influenced by sex and diet, with an interaction between these factors. Females had higher NPY mRNA levels than males when on a chow diet (F(1,10) = 5.5, p < 0.05), but lower levels than males in the HHHH (F(1,9) = 7.8, p < 0.05) and HHCH (F(1,11) = 4.8, p = 0.05) groups. Decreased NPY mRNA levels were found in the HHHH, CCCH and HHCH groups in both males (F(3,21) = 30.6, p < 0.001) and females (F(3,22) = 15.0, p < 0.001). Relative AgRP mRNA levels (Fig. 6F) were higher in females than in males on chow (F(1,11) = 17.3, p < 0.01), as well as on the CCCH dietary regimen (F(1,10) = 8.9, p < 0.05). The HHHH, CCCH and HHCH groups had lower AgRP mRNA levels than chow-fed mice in both males (F(3,22) = 11.2, p < 0.001) and females (F(3,22) = 36.2, p < 0.001). Females had overall higher POMC mRNA levels (Fig. 6G) than males, with no effect of diet in either sex.
IGF2 effects in astrocyte cultures
As a positive correlation between hypothalamic IGF2 and IGFBP2 were found here and previously24,37,39, we analyzed whether hypothalamic astrocytes produce IGFBP2 in response to IGF2. As IGFs are mitogenic52,53,54,55, we analyzed cell number (Fig. 7A), which was affected by sex, with a general effect of IGF2 and the IGF2 dose (statistical analysis can be found in Supplementary Table 6). Indeed, IGF2 increased cell number in both sexes. IGF2 treatment did not alter relative IGF1 mRNA levels (Fig. 7B), nor those of IGF2R (Fig. 7C), but there was an effect of sex in both cases. In male astrocytes, IGF2 increased IGFBP2 mRNA levels (Fig. 7D) at higher concentrations (F(1,12) = 4.5, p < 0.05).

Cell number (A) and the relative IGF1 (B), IGF2 receptor (IGF2R, C) and IGFBP2 (D) mRNA levels in hypothalamic astrocyte cultures from male and female rats after 24 h of IGF2 treatment at 10, 25, 100 or 200 ng/ml. **p < 0.01. a: effect of the sex; #, general effect of IGF2; δ, effect of the dose of IGF2. n = 3.
Discussion
Here we show that not only are there sex differences in weight gain, energy intake and energy efficiency after HFD intake, as previously reported28,29,56,57,58,59,60, as well as in weight loss in response to a healthy diet61,62, but that the response to resumption of a HFD after weight loss on an unrestricted healthy diet also differs between the sexes. Indeed, laboratory chow diets have been developed as a healthy nutritional choice for rodents, due to its energy content, the low amount of fats (especially saturated fats), the inclusion of complex carbohydrates and fiber, as well as the required vitamins and minerals63. A main difference between the sexes is that males modulate their Kcal intake on a HFD, returning to control levels, while females continue to have an elevated energy intake. Although both sexes drastically reduced energy intake when returned to the healthier chow diet after consuming HFD, overweight females required more time to achieve significant weight loss and lost less weight than males. Dissimilarities were also found in insulin resistance, with males being more affected as previously reported64, as well as in the central IGF system, which was also modulated in males. These results are in agreement with the observations that women are reported to be more efficient in defending body weight and glucose parameters than men65. Indeed, the slower changes in body weight in females after both increased and decreased caloric intake could involve the protective role of estrogens32,66.
As previously shown, on a HFD females consumed more kilocalories per gram of body weight compared to males59 and males had a higher energy efficiency57 and gained weight more rapidly. Diets rich in fat are more palatable than chow, and female rats consume palatable foods independently of their satiety state67, while males more readily adjust their energy consumption to their hunger state; however, both males and females are reported to eat standard chow according to their hunger state68. The difference in palatability of the diets could also explain the drastic reduction in energy intake when the mice were given chow after HFD.
Sex differences in the hypothalamic circuitry that regulates homeostatic food intake and energy expenditure69 most likely underlie some of the metabolic dissimilarities between males and females observed here. Male mice re-exposed to HFD and those given HFD for the first time initially consumed more kilocalories than HHHH mice. This was not observed in females. It is possible that in females effects on food intake were not found due to the high variability in intake over time, especially in the HFD-exposed mice and differences in the estrous cycle stage could be involved in this variability. Indeed, female mice were only synchronized and the estrous stage analyzed at the end of the study; thus, females could be at one of the four different stages of the estrous cycles, which are known to differentially affect to food intake70.
Despite some sex differences, mice gained and lost weight after the dietary changes with no difficulty and with no apparent difference on weight gain between mice re-exposed or exposed to HFD for the first time. Likewise, the percentage of adipose tissue and circulating leptin levels increased with HFD in both sexes, regardless of the HFD regimen. One might speculate that some effects of HFD intake might require more time, with a greater weight gain and longer development of secondary complications required to cause more long-lasting and profound effects on the metabolic control system such that it is not totally normalized with weight loss. Indeed, the time an individual is obese is important for the rebound effect on adiposity in humans71. However, we observed no rebound effect, which could be because here mice were never on a restrictive diet, which can alter hedonic behaviors that can influence ingestion when exposed to palatable foods; this may also underlie some differences with other interventional studies.
Alterations in glucose metabolism were observed in response to long-term HFD intake, as previously reported59,60,72, with no differences in glycemia or the AUC being observed between the HHHH, CCCH and HHCH groups. However, male HHCH mice had higher insulin and HOMA-IR than HHHH mice. This indicates that glucose metabolism was not totally normalized with weight loss and/or it became more susceptible to further dietary assaults, as HHCH male mice required higher levels of insulin to maintain normal glycemia. In females no differences were found between HHHH and HHCH mice groups. Affectation of glucose tolerance occurred as early as one month after HFD intake in male mice, while females ingesting a HFD for only one month had lower insulin and HOMA levels compared to both HHHH continuous-HFD intake and HHCH mice, again indicating that females are more resistant to these modifications. Basal glycemia at sacrifice was not affected by diet, whereas basal glycemia in the GTT was higher in the groups that consumed HFD, which might be explained by the differences in the duration of fasting (12 h before sacrifices and 6 h before GTT) may explain this discrepancy.
Sex differences in the circulating IGF system have been reported in humans35,36,73 and rodents25,74. Here we observed higher peripheral levels of IGF2 in females compared to males, but no sex differences in total or free IGF1 levels, as previously reported74. This is in agreement, some studies showing no sex differences in total IGF125 or free IGF1 levels through development74, but not with others24,39. Circulating total IGF1 levels increased in mice of both sexes with HFD-induced weight gain, as in previous studies75,76; however, no dietary changes in free IGF1, IGF2 or IGFBP2 levels were found here. Some studies report an increase38,77 and others a reduction78 in circulating IGF2 levels in patients with obesity, with these differences possibly being due to the degree and/or duration of obesity. IGFBP2, with reduced circulating levels in patients with obesity34,38,79 and animal models of obesity80, is purported to be protective against obesity and diabetes onset42,43. However, no changes in circulating IGFBP2 levels were observed here. This apparent discrepancy with our previous studies could be associated with the lack of dietary effect on basal glycemia in the study presented here.
Increased central IGF1 improves peripheral glucose metabolism and increases insulin sensitivity44. Moreover, increased hypothalamic IGF1 levels could be involved in protection against HFD-induced metabolic damage. Hypothalamic IGF1 mRNA levels increased in male mice after 7 weeks of HFD intake81, but here they were reduced after 4 weeks (CCCH) in both sexes, while after 4 months of HFD no changes were found. This indicates dynamic changes in this growth factor in response to metabolic modifications that deserve further investigation. Elevated circulating IGF2 is reported to exert a negative feedback effect on IGF1 levels82 and the reduction in IGF1 mRNA levels in CCCH mice could be a consequence, at least in part, of increased IGF2 production. However, when we analyzed whether astrocytes, one of the main sources of these factors centrally, might be involved in this control, we saw no effect of IGF2 on IGF1 mRNA levels, suggesting that other signals and/or cell types are involved.
Hypothalamic IGF2 mRNA levels were increased in CCCH males, with no changes found in females. However, hypothalamic IGF2 mRNA was previously reported to increase after 2 months of HFD intake in females37. This differential timing in IGF2 modifications could be associated with differences between the sexes in the onset of alterations in glucose metabolism83. In agreement, both glycemia and the hypothalamic IGF2 mRNA levels increase after more extended exposure to HFD in females37. IGFBP2 mRNA levels were also increased exclusively in the hypothalamus of male CCCH mice. As IGFBP2 is reported to exert a protective role against obesity and diabetes onset42,43, and glucose metabolism is more rapidly affected in males after HFD intake57,72,84, one might speculate that the novel HFD exposure results in activation of protective mechanisms. Hypothalamic IGF2 and IGFBP2 mRNA levels were positively correlated, as reported previously24,37,39. In male astrocytes, IGF2 increased IGFBP2 mRNA levels suggesting that these glial cells might be involved in the central association of these factors. This binding protein has a higher preference for binding IGF2 over that of IGF185 and may act as a regulator of this ligand86. Thus, the increase in IGFBP2 mRNA could be to regulate IGF2 levels87.
Females had higher mRNA levels of NPY, AgRP and POMC than males, as previously reported88,89. The levels of NPY and AgRP mRNA were decreased as early as one month after HFD intake in both sexes, a previously reported phenomenon interpreted as homeostatic response to reduce food intake90,91 and indeed, a reduction in energy intake was observed after the initial hyperphagic period, especially in males. Other studies indicate no change in these orexigenic neuropeptides after two weeks of HFD in male mice92 or after longer periods (4 to 8 weeks)93. We previously reported that expression of both orexigenic neuropeptides returned to control levels after one month of a chow diet and weight loss in both sexes39, indicating that the plasticity of this system in response to diet changes and weight loss was maintained.
No effects of diet on hypothalamic POMC mRNA levels were observed here. Hypothalamic POMC mRNA levels have been reported to increase in male mice after 781 and 11 weeks of HFD94, or to not change in male mice after 16 weeks95 or in female mice after 20 weeks96 of HFD. The composition of the HFD could explain part of these differences.
In conclusion, males and females have different metabolic responses and the results reported here reinforce this concept, demonstrating dissimilarities in the response to different dietary changes. The weight gain in response to the re-exposition to HFD did not differ from that observed in mice ingesting HFD for the first time, indicating that greater weight gain and/or a longer period of being overweight may be necessary to observe a difference. However, it is also possible that the employment of a healthy diet to lose weight instead of food restriction underlies this observation and this deserves further investigation as it could be important for weight loss counseling. Moreover, we might speculate that following a restricted diet instead would lower resting metabolic rate and return to HFD would result in a rebound effect. Glucose metabolism was more affected by reestablishment of poor dietary habits in males, again demonstrating sex differences in this parameter, with the IGF system possibly being involved in this process, as it was differentially modulated in males and females and this clearly deserves further studies.
Methods
Ethical statement
This study was performed in accordance with the European Communities Council Directive (2010/63/UE) and the Royal Decree 53/2013 pertaining to the protection of experimental animals. It was also approved by the Ethical Committee of Animal Experimentation of the Hospital Puerta de Hierro of Madrid and the Animal Welfare Organ of the Community of Madrid. Mice were maintained at 22 ± 2 °C with free access to tap water in a stable 12-h light–dark cycle. The study design and reporting of the results adheres to the ARRIVE guidelines (https://arriveguidelines.org).
Animals and diets
Seventy-two six-week-old C57BL/6J mice (36 males and 36 females) were purchased from Charles River Laboratories (Sant Cugat del Vallès, Barcelona, Spain). Upon their arrival, animals were weighed, and 3 mice of the same sex were randomly placed in each cage. They had free access to standard rodent chow and tap water while being allowed to acclimate to their new environment for one week.
For the study, one group (Chow) of mice was fed a standard rodent chow diet (6% Kcal from fat, 17% Kcal from proteins, 77% from carbohydrates, 3.41 kcal/g, Panlab, Barcelona, Spain) during the 4 months and served as our controls. Another group (HHHH) received a HFD (62% Kcal from fat, 18% Kcal from proteins, 20% Kcal from carbohydrates, 5.1 kcal/g, LabDiet, Sodispan Research SL, Madrid, Spain) for the same period of time. A third group received chow for 3 months and then was given the HFD for the final month (CCCH). To determine the effect of “re-exposition to HFD” mice were fed with HFD for the two initial months, switched to chow for the third month, and then was re-exposed to HFD for the last month of the study (HHCH). Nine animals of each sex were included in each group (n = 9), resulting in 3 cages per sex and diet. Body weight and food intake were monitored weekly until the termination of the study. Food intake was monitored by weighing the amount of food remaining of a previously determined quantity, with the pieces that fell into the cages being taken into account. However, a caveat that should be taken into account is that due to the lower consistency of the HFD, it breaks more easily and the recovery may be less efficient. Mean energy efficiency during the entire study was calculated as the weight gained in grams per the total number of calories consumed.
Glucose tolerance test (GTT)
One week prior to the end of the study, a glucose tolerance test (GTT) was performed as described37,39. Six animals per group were randomly chosen and then fasted for 6 h but maintaining free access to tap water. Mice were weighed and intraperitoneally injected with a D-glucose solution in PBS (0.4 g/ml) at a dose of 2 mg per gram of body weight. A Freestyle Optimum Neo glucometer (Abbott, Whitney, UK) was employed for determining glycemia at 0 (basal), 30-, 60-, 90-, and 120-min post injection.
Sacrifices and tissue collection
To synchronize the estrous cycle in female mice thus avoiding additional variability, bedding from male cages was mixed in the bedding of the females’ cages 3–4 days before sacrifice. The estrous cycle stage was determined by vaginal cytology at sacrifice, with 83% of the females being sacrificed at estrous. Mice were weighed and fasted 12 h prior to sacrifice, which took place by decapitation after rapid exposure to CO2 to reduce suffering between 09:00 and 11:00 am. Glycemia levels were measured with a Freestyle Optimum Neo glucometer (Abbott). Peripheral blood was collected in tubes with a 0.5 M ethylenediaminetetraacetic acid (EDTA) solution to prevent clotting. Tubes were centrifuged at 3000 rpm for 15 min at 4 °C and plasma was aliquoted and maintained at − 80 °C.
In mice, the brain was extracted from skull after decapitation, and the hypothalamus (rostrally limited by the optic chiasm and caudally by the anterior margin of the mammillary bodies) was then dissected and frozen. Retroperitoneal visceral adipose tissue and inguinal subcutaneous white adipose tissue depots were dissected, weighed and frozen. All tissues were stored at − 80 °C until processing.
Enzyme-linked immunosorbent assays (ELISAs)
Plasma levels of free IGF1 (Ref.: AL-136, AnshLabs, Webster, TX, USA), total IGF1 (Ref.: E25; Mediagnost, Reutlingen, Germany), IGF2 (Ref.: MG200; R&D Systems, Minneapolis, Minnesota, USA), IGFBP2 (Ref.: RAB0234; Millipore, Burlington, MA, USA), insulin (Ref.: EZRMI-13K; Millipore) and leptin (Ref.: EZML-82K; Millipore) were assayed according to manufacturer’s instructions, and absorbance was read using a TECAN Infinite M200. Homeostatic Model Assessment for Insulin Resistance (HOMA-IR) was calculated according to the equation:
Quantitative real-time polymerase chain reaction (RT-qPCR)
An RNeasy Plus Mini Kit (Qiagen, Hilden, Germany) was used to isolate RNA according to the manufacturer’s instructions. The RT-PCR assays were performed as previously described57, in which 0.5 μg of RNA were used and the NZY First-Strand cDNA Synthesis Kit (NZYTech, Lisbon, Portugal) was used for retrotranscription. TaqMan probes for the genes of interest (Table 2) were employed, and a QuantStudio 3 Real-Time PCR System (Applied Biosystems, Carlsbad, CA, USA) was used for signal detection.
As the housekeeping gene, an endogenous GAPDH control (Applied Biosystems) was used, and the ΔΔCt method was applied for the analyses. In the in vivo studies, the results were normalized and expressed in percentage in comparison with the male chow group, whereas in the in vitro studies the reference group was the control treatment in astrocytes from male rats.
Primary hypothalamic astrocyte cultures
Primary astrocyte cultures were obtained from PND2 Wistar rat pups and performed as previously described97. Male and female pups were differentiated by ano-genital distance and cultured separately. Rat pups were sacrificed by decapitation, the brains extracted and the hypothalami dissected, with the meninges being carefully discarded. Astrocyte cultures were maintained in DMEM/F12 media (Gibco, Invitrogen Co., Thermo-Fisher, Waltham, Massachusetts, USA) enriched with 10% fetal bovine serum (FBS, Sigma-Merck, Darmstadt, Germany) and treated with 1% penicillin/streptomycin and anti-mycotic (Gibco), with the medium being changed every 2–3 days. When the cell confluence was approximately 70%, the flasks were placed in an Ecotron orbital shaking incubator for 16 h at 280 rpm and 37 °C to eliminate remaining oligodendrocytes and microglia. Cells were then trypsinized and counted by using a Countess II FL Automated Cell Counter (Thermo-Fisher) and seeded at 1.5 × 104 cells/cm2 in 100 mm culture plates and allowed to recover for 24 h. Cells were serum-starved for 24 h prior to the treatments. For the different experiments, astrocytes were treated for 24 h with an IGF2 (I8904, Sigma-Merck) solution in PBS at 10, 25, 100 or 200 ng/ml supplemented with 0.1% BSA, as described98, or with vehicle.
Cell number estimation by crystal violet staining
Cells were seeded at 3.5 × 104 cells per well in 24-well plates for 24 h. After serum-starvation as described above, treatments were added for 24 h. After treatment, a 1% glutaraldehyde solution in PBS was added for 15 min, rinsed and then a 0.1% crystal violet (Sigma-Merck) solution was added for 20 min and then thoroughly rinsed. When the plates were dry, a 10% acetic acid solution was added, and the dye solubilized for 5 min. The samples were added to a 96 well plate and the absorbance at 590 nm was read on a TECAN Infinite M200. The absorbance in each well was normalized to the control values.
Statistical analyses
Statistical analyses were performed with SPSS 15.0 (SPSS Inc., Chicago, IL, USA) software, whereas graphs were made with GraphPad Prism 8 software (San Diego, CA, USA). In the in vivo studies, a 2-way ANOVA with the factors of sex and diet (chow, HHHH, CCCH or HHCH) was performed, which was followed by a 1-way ANOVA when appropriate. In all experiments, weight gain and food intake over time, as well as glycemia changes during the GTT, were analyzed by a 3-way ANOVA with repeated measures with time, sex and the diet as factors. In the in vitro study, a 2-way ANOVA was used to determine the effects and interaction between the sex and the IGF2 dose (0, 10, 25, 100 or 200 ng/ml). A 2-way ANOVA with sex and the presence of IGF2 regardless of dose was also calculated. A Pearson correlation coefficient was calculated for the linear correlation between variables. In all analyses, p < 0.05 was considered significant.
Data availability
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation by contacting Dr. J.A. Chowen.
References
Valero-Perez, M. et al. Regular consumption of Lipigo((R)) promotes the reduction of body weight and improves the rebound effect of obese people undergo a comprehensive weight loss program. Nutrients 12, 1960 (2020).
Kroeger, C. M., Hoddy, K. K. & Varady, K. A. Impact of weight regain on metabolic disease risk: A review of human trials. J. Obes. 2014, 614519 (2014).
MacLean, P. S. et al. Enhanced metabolic efficiency contributes to weight regain after weight loss in obesity-prone rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 287, R1306-1315 (2004).
Zapata, R. C. et al. Adipocytes control food intake and weight regain via Vacuolar-type H(+) ATPase. Nat. Commun. 13, 5092 (2022).
Busetto, L. et al. Mechanisms of weight regain. Eur. J. Intern. Med. 93, 3–7 (2021).
Delgado-Lista, J. et al. Long-term secondary prevention of cardiovascular disease with a Mediterranean diet and a low-fat diet (CORDIOPREV): A randomised controlled trial. Lancet 399, 1876–1885 (2022).
Estruch, R. et al. Effect of a high-fat Mediterranean diet on bodyweight and waist circumference: A prespecified secondary outcomes analysis of the PREDIMED randomised controlled trial. Lancet Diabetes Endocrinol. 7, e6–e17 (2019).
Zhang, H. Y. et al. Neuroprotective effects of insulin-like growth factor-2 in 6-hydroxydopamine-induced cellular and mouse models of Parkinson’s disease. Neural Regen. Res. 18, 1099–1106 (2023).
Pereira, S. S. et al. IGF2 role in adrenocortical carcinoma biology. Endocrine 66, 326–337 (2019).
Supeno, N. E. et al. IGF-1 acts as controlling switch for long-term proliferation and maintenance of EGF/FGF-responsive striatal neural stem cells. Int. J. Med. Sci. 10, 522–531 (2013).
Yakar, S. et al. Circulating levels of IGF-1 directly regulate bone growth and density. J. Clin. Investig. 110, 771–781 (2002).
Kineman, R. D., Del Rio-Moreno, M. & Sarmento-Cabral, A. 40 YEARS of IGF1: Understanding the tissue-specific roles of IGF1/IGF1R in regulating metabolism using the Cre/loxP system. J. Mol. Endocrinol. 61, T187–T198 (2018).
Lewitt, M. S., Dent, M. S. & Hall, K. The insulin-like growth factor system in obesity, insulin resistance and type 2 diabetes mellitus. J. Clin. Med. 3, 1561–1574 (2014).
Livingstone, C. & Borai, A. Insulin-like growth factor-II: Its role in metabolic and endocrine disease. Clin. Endocrinol. (Oxf.) 80, 773–781 (2014).
Yakar, S., Werner, H. & Rosen, C. J. Insulin-like growth factors: Actions on the skeleton. J. Mol. Endocrinol. 61, T115–T137 (2018).
Chernausek, S. D. et al. Long-term treatment with recombinant insulin-like growth factor (IGF)-I in children with severe IGF-I deficiency due to growth hormone insensitivity. J. Clin. Endocrinol. Metab. 92, 902–910 (2007).
Yakar, S. et al. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proc. Natl. Acad. Sci. USA 96, 7324–7329 (1999).
Pan, W. & Kastin, A. J. Interactions of IGF-1 with the blood-brain barrier in vivo and in situ. Neuroendocrinology 72, 171–178 (2000).
Myhre, C. L. et al. Microglia express insulin-like growth factor-1 in the hippocampus of aged APP(swe)/PS1(DeltaE9) transgenic mice. Front. Cell. Neurosci. 13, 308 (2019).
Zhang, J., Moats-Staats, B. M., Ye, P. & D’Ercole, A. J. Expression of insulin-like growth factor system genes during the early postnatal neurogenesis in the mouse hippocampus. J. Neurosci. Res. 85, 1618–1627 (2007).
Suh, H. S., Zhao, M. L., Derico, L., Choi, N. & Lee, S. C. Insulin-like growth factor 1 and 2 (IGF1, IGF2) expression in human microglia: Differential regulation by inflammatory mediators. J. Neuroinflamm. 10, 37 (2013).
Pitt, J. et al. Neuroprotective astrocyte-derived insulin/insulin-like growth factor 1 stimulates endocytic processing and extracellular release of neuron-bound Abeta oligomers. Mol. Biol. Cell 28, 2623–2636 (2017).
Baxter, R. C. Insulin-like growth factor (IGF)-binding proteins: Interactions with IGFs and intrinsic bioactivities. Am. J. Physiol. Endocrinol. Metab. 278, E967-976 (2000).
Guerra-Cantera, S. et al. Short-term diet induced changes in the central and circulating IGF systems are sex specific. Front. Endocrinol. (Lausanne) 11, 513 (2020).
Peshti, V. et al. Characterization of physiological defects in adult SIRT6-/- mice. PLoS ONE 12, e0176371 (2017).
Donahue, C. P., Kosik, K. S. & Shors, T. J. Growth hormone is produced within the hippocampus where it responds to age, sex, and stress. Proc. Natl. Acad. Sci. USA 103, 6031–6036 (2006).
Borski, R. J., Tsai, W., DeMott-Friberg, R. & Barkan, A. L. Regulation of somatic growth and the somatotropic axis by gonadal steroids: Primary effect on insulin-like growth factor I gene expression and secretion. Endocrinology 137, 3253–3259 (1996).
Macotela, Y., Boucher, J., Tran, T. T. & Kahn, C. R. Sex and depot differences in adipocyte insulin sensitivity and glucose metabolism. Diabetes 58, 803–812 (2009).
Trevaskis, J. L., Meyer, E. A., Galgani, J. E. & Butler, A. A. Counterintuitive effects of double-heterozygous null melanocortin-4 receptor and leptin genes on diet-induced obesity and insulin resistance in C57BL/6J mice. Endocrinology 149, 174–184 (2008).
Corsetti, J. P., Sparks, J. D., Peterson, R. G., Smith, R. L. & Sparks, C. E. Effect of dietary fat on the development of non-insulin dependent diabetes mellitus in obese Zucker diabetic fatty male and female rats. Atherosclerosis 148, 231–241 (2000).
Dakin, R. S., Walker, B. R., Seckl, J. R., Hadoke, P. W. & Drake, A. J. Estrogens protect male mice from obesity complications and influence glucocorticoid metabolism. Int. J. Obes. (Lond.) 39, 1539–1547 (2015).
Stubbins, R. E., Holcomb, V. B., Hong, J. & Nunez, N. P. Estrogen modulates abdominal adiposity and protects female mice from obesity and impaired glucose tolerance. Eur. J. Nutr. 51, 861–870 (2012).
Clegg, D. J. et al. Estradiol-dependent decrease in the orexigenic potency of ghrelin in female rats. Diabetes 56, 1051–1058 (2007).
Barrios, V., Buno, M., Pozo, J., Munoz, M. T. & Argente, J. Insulin-like growth factor-binding protein-2 levels in pediatric patients with growth hormone deficiency, eating disorders and acute lymphoblastic leukemia. Horm. Res. 53, 221–227 (2000).
Argente, J. et al. Normative data for insulin-like growth factors (IGFs), IGF-binding proteins, and growth hormone-binding protein in a healthy Spanish pediatric population: Age- and sex-related changes. J. Clin. Endocrinol. Metab. 77, 1522–1528 (1993).
Martin-Rivada, A. et al. Pappalysins and stanniocalcins and their relationship with the peripheral IGF axis in newborns and during development. J. Clin. Endocrinol. Metab. 107, 2912–2924 (2022).
Guerra-Cantera, S. et al. Impact of long-term HFD intake on the peripheral and central IGF system in male and female mice. Metabolites 10, 462 (2020).
Argente, J. et al. Multiple endocrine abnormalities of the growth hormone and insulin-like growth factor axis in prepubertal children with exogenous obesity: Effect of short- and long-term weight reduction. J. Clin. Endocrinol. Metab. 82, 2076–2083 (1997).
Guerra-Cantera, S. et al. Sex differences in metabolic recuperation after weight loss in high fat diet-induced obese mice. Front. Endocrinol. (Lausanne) 12, 796661 (2021).
Barrand, S., Crowley, T. M., Wood-Bradley, R. J., De Jong, K. A. & Armitage, J. A. Impact of maternal high fat diet on hypothalamic transcriptome in neonatal Sprague Dawley rats. PLoS ONE 12, e0189492 (2017).
Uchimura, T. et al. An essential role for IGF2 in cartilage development and glucose metabolism during postnatal long bone growth. Development 144, 3533–3546 (2017).
Hedbacker, K. et al. Antidiabetic effects of IGFBP2, a leptin-regulated gene. Cell Metab. 11, 11–22 (2010).
Wheatcroft, S. B. et al. IGF-binding protein-2 protects against the development of obesity and insulin resistance. Diabetes 56, 285–294 (2007).
Hong, H. et al. Central IGF1 improves glucose tolerance and insulin sensitivity in mice. Nutr. Diabetes 7, 2 (2017).
Le Roith, D., Kim, H., Fernandez, A. M. & Accili, D. Inactivation of muscle insulin and IGF-I receptors and insulin responsiveness. Curr. Opin. Clin. Nutr. Metab. Care 5, 371–375 (2002).
Yakar, S. et al. Liver-specific igf-1 gene deletion leads to muscle insulin insensitivity. Diabetes 50, 1110–1118 (2001).
Chowen, J. A., Frago, L. M. & Fernandez-Alfonso, M. S. Physiological and pathophysiological roles of hypothalamic astrocytes in metabolism. J. Neuroendocrinol. 31, e12671 (2019).
Timper, K. & Bruning, J. C. Hypothalamic circuits regulating appetite and energy homeostasis: Pathways to obesity. Dis. Model. Mech. 10, 679–689 (2017).
Chowen, J. A. et al. The role of astrocytes in the hypothalamic response and adaptation to metabolic signals. Prog. Neurobiol. 144, 68–87 (2016).
Leloup, C. et al. Glucose and hypothalamic astrocytes: More than a fueling role?. Neuroscience 323, 110–120 (2016).
Chen, W., He, B., Tong, W., Zeng, J. & Zheng, P. Astrocytic insulin-like growth factor-1 protects neurons against excitotoxicity. Front. Cell. Neurosci. 13, 298 (2019).
Ferreira Mendes, J. M. et al. Effects of IGF-1 on proliferation, angiogenesis, tumor stem cell populations and activation of AKT and hedgehog pathways in oral squamous cell carcinoma. Int. J. Mol. Sci. 21, 6487 (2020).
Gao, S. et al. lncINS-IGF2 promotes cell proliferation and migration by promoting G1/S transition in lung cancer. Technol. Cancer Res. Treat. 18, 1533033818823029 (2019).
Ren, M. et al. Insulin-like growth factor-1 promotes cell cycle progression via upregulation of cyclin D1 expression through the phosphatidylinositol 3-kinase/nuclear factor-kappaB signaling pathway in FRTL thyroid cells. Acta Pharmacol. Sin. 30, 113–119 (2009).
Morrione, A. et al. Insulin-like growth factor II stimulates cell proliferation through the insulin receptor. Proc. Natl. Acad. Sci. USA 94, 3777–3782 (1997).
Ros, P. et al. Sex differences in long-term metabolic effects of maternal resveratrol intake in adult rat offspring. Endocrinology 161, bqaa090 (2020).
Freire-Regatillo, A. et al. Sex differences in the peripubertal response to a short-term, high-fat diet intake. J. Neuroendocrinol. 32, e12756 (2020).
Fontes-Oliveira, C. C., Soares Oliveira, B. M., Körner, Z., Harandi, V. M. & Durbeej, M. Effects of metformin on congenital muscular dystrophy type 1A disease progression in mice: A gender impact study. Sci. Rep. 8, 16302 (2018).
Paruthiyil, S., Hagiwara, S. I., Kundassery, K. & Bhargava, A. Sexually dimorphic metabolic responses mediated by CRF(2) receptor during nutritional stress in mice. Biol. Sex Differ. 9, 49 (2018).
Pettersson, U. S., Walden, T. B., Carlsson, P. O., Jansson, L. & Phillipson, M. Female mice are protected against high-fat diet induced metabolic syndrome and increase the regulatory T cell population in adipose tissue. PLoS ONE 7, e46057 (2012).
Crane, M. M., Jeffery, R. W. & Sherwood, N. E. Exploring gender differences in a randomized trial of weight loss maintenance. Am. J. Mens Health 11, 369–375 (2017).
Bhogal, M. S. & Langford, R. Gender differences in weight loss; Evidence from a NHS weight management service. Public Health 128, 811–813 (2014).
Warden, C. H. & Fisler, J. S. Comparisons of diets used in animal models of high-fat feeding. Cell Metab. 7, 277 (2008).
Hevener, A., Reichart, D., Janez, A. & Olefsky, J. Female rats do not exhibit free fatty acid-induced insulin resistance. Diabetes 51, 1907–1912 (2002).
Hagobian, T. A. & Evero, N. Exercise and weight loss: what is the evidence of sex differences?. Curr. Obes. Rep. 2, 86–92 (2013).
Handgraaf, S. et al. Prevention of obesity and insulin resistance by estrogens requires ERalpha activation function-2 (ERalphaAF-2), whereas ERalphaAF-1 is dispensable. Diabetes 62, 4098–4108 (2013).
Buttigieg, A. et al. Preference for high-fat diet is developed by young Swiss CD1 mice after short-term feeding and is prevented by NMDA receptor antagonists. Neurobiol. Learn. Mem. 107, 13–18 (2014).
Buczek, L., Migliaccio, J. & Petrovich, G. D. Hedonic eating: Sex differences and characterization of orexin activation and signaling. Neuroscience 436, 34–45 (2020).
Chowen, J. A., Freire-Regatillo, A. & Argente, J. Neurobiological characteristics underlying metabolic differences between males and females. Prog. Neurobiol. 176, 18–32 (2019).
Olofsson, L. E., Pierce, A. A. & Xu, A. W. Functional requirement of AgRP and NPY neurons in ovarian cycle-dependent regulation of food intake. Proc. Natl. Acad. Sci. USA 106, 15932–15937 (2009).
Aronoff, J. E. et al. Why do humans undergo an adiposity rebound? Exploring links with the energetic costs of brain development in childhood using MRI-based 4D measures of total cerebral blood flow. Int. J. Obes. (Lond.) 46, 1044–1050 (2022).
Underwood, E. L. & Thompson, L. T. A high-fat diet causes impairment in hippocampal memory and sex-dependent alterations in peripheral metabolism. Neural Plast. 2016, 7385314 (2016).
Waters, D. L., Yau, C. L., Montoya, G. D. & Baumgartner, R. N. Serum sex hormones, IGF-1, and IGFBP3 exert a sexually dimorphic effect on lean body mass in aging. J. Gerontol. A Biol. Sci. Med. Sci. 58, 648–652 (2003).
Frystyk, J. et al. Developmental changes in serum levels of free and total insulin-like growth factor I (IGF-I), IGF-binding protein-1 and -3, and the acid-labile subunit in rats. Endocrinology 139, 4286–4292 (1998).
Doyle, S. L. et al. IGF-1 and its receptor in esophageal cancer: Association with adenocarcinoma and visceral obesity. Am. J. Gastroenterol. 107, 196–204 (2012).
l’Allemand, D. et al. Associations between body mass, leptin, IGF-I and circulating adrenal androgens in children with obesity and premature adrenarche. Eur. J. Endocrinol. 146, 537–543 (2002).
Xuan, L. et al. Insulin-like growth factor 2 promotes adipocyte proliferation, differentiation and lipid deposition in obese type 2 diabetes. J. Transl. Sci. 6, 1–7 (2019).
Sandhu, M. S., Gibson, J. M., Heald, A. H., Dunger, D. B. & Wareham, N. J. Low circulating IGF-II concentrations predict weight gain and obesity in humans. Diabetes 52, 1403–1408 (2003).
Frystyk, J., Skjaerbaek, C., Vestbo, E., Fisker, S. & Orskov, H. Circulating levels of free insulin-like growth factors in obese subjects: The impact of type 2 diabetes. Diabetes Metab. Res. Rev. 15, 314–322 (1999).
Kang, H. S. et al. Metformin stimulates IGFBP-2 gene expression through PPARalpha in diabetic states. Sci. Rep. 6, 23665 (2016).
Baquedano, E. et al. The absence of GH signaling affects the susceptibility to high-fat diet-induced hypothalamic inflammation in male mice. Endocrinology 155, 4856–4867 (2014).
Fukuda, I. et al. Clinical features of insulin-like growth factor-II producing non-islet-cell tumor hypoglycemia. Growth Horm. IGF Res. 16, 211–216 (2006).
Garla, V. et al. Non-islet cell hypoglycemia: Case series and review of the literature. Front. Endocrinol. (Lausanne) 10, 316 (2019).
Samuel, P., Khan, M. A., Nag, S., Inagami, T. & Hussain, T. Angiotensin AT(2) receptor contributes towards gender bias in weight gain. PLoS ONE 8, e48425 (2013).
Bach, L. A. IGF-binding proteins. J. Mol. Endocrinol. 61, T11–T28 (2018).
Rogler, C. E. et al. Altered body composition and increased frequency of diverse malignancies in insulin-like growth factor-II transgenic mice. J. Biol. Chem. 269, 13779–13784 (1994).
Grimes, R. W. & Hammond, J. M. Insulin and insulin-like growth factors (IGFs) stimulate production of IGF-binding proteins by ovarian granulosa cells. Endocrinology 131, 553–558 (1992).
Ruigrok, S. R. et al. Modulation of the hypothalamic nutrient sensing pathways by sex and early-life stress. Front. Neurosci. 15, 695367 (2021).
Wang, C. et al. TAp63 contributes to sexual dimorphism in POMC neuron functions and energy homeostasis. Nat. Commun. 9, 1544 (2018).
Mazzone, C. M. et al. High-fat food biases hypothalamic and mesolimbic expression of consummatory drives. Nat. Neurosci. 23, 1253–1266 (2020).
Horvath, T. L. et al. Synaptic input organization of the melanocortin system predicts diet-induced hypothalamic reactive gliosis and obesity. Proc. Natl. Acad. Sci. USA 107, 14875–14880 (2010).
Staszkiewicz, J., Horswell, R. & Argyropoulos, G. Chronic consumption of a low-fat diet leads to increased hypothalamic agouti-related protein and reduced leptin. Nutrition 23, 665–671 (2007).
de Araujo, T. M. et al. The partial inhibition of hypothalamic IRX3 exacerbates obesity. EBioMedicine 39, 448–460 (2019).
Gamber, K. M. et al. Over-expression of leptin receptors in hypothalamic POMC neurons increases susceptibility to diet-induced obesity. PLoS ONE 7, e30485 (2012).
Ito, Y. et al. GABA type B receptor signaling in proopiomelanocortin neurons protects against obesity, insulin resistance, and hypothalamic inflammation in male mice on a high-fat diet. J. Neurosci. 33, 17166–17173 (2013).
Roepke, T. A. et al. Loss of ERalpha partially reverses the effects of maternal high-fat diet on energy homeostasis in female mice. Sci. Rep. 7, 6381 (2017).
Fuente-Martin, E. et al. Leptin regulates glutamate and glucose transporters in hypothalamic astrocytes. J. Clin. Investig. 122, 3900–3913 (2012).
Zhang, K. et al. Insulin-like growth factor 2 promotes the adipogenesis of hemangioma-derived stem cells. Exp. Ther. Med. 17, 1663–1669 (2019).
Acknowledgements
We would like to thank the entire animal care personal from the Hospital Puerta de Hierro for their excellent technical assistance.
Funding
The authors are funded by Grants from the Spanish Ministry of Science and Innovation (BFU2017-82565-C21-R2 and PU-PID2021-122653OB-I00 to JC & LF), Fondo de Investigación Sanitaria (PI1600485 and PI1900166 to JA), Fondos FEDER and Centro de Investigación Biomédica en Red Fisiopatologı́a de Obesidad y Nutrición (CIBEROBN), Instituto de Salud Carlos III (JA).
Author information
Authors and Affiliations
Contributions
Conception and design of study: L.F., J.A., and J.C. Funding acquisition: L.F., J.A., and J.C.. Animal handling: S.G.-C., M.J.-H., R.C.-P., P.R., J.G.-P., and I.P.-N. Biochemical analysis: S.G.-C., S.C., and V.B. Data analysis and discussion: S.G.-C., L.F., J.A., and J.C. Redaction of manuscript: S.G.-C., L.M.F., and J.C. Revision of manuscript: S.G.-C., L.F., M.J.-H., R.C.-P., S.C., P.R., J.G.-P., I.P.-N., V.B., J.A., and J.C.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Guerra-Cantera, S., Frago, L.M., Jiménez-Hernaiz, M. et al. The metabolic effects of resumption of a high fat diet after weight loss are sex dependent in mice. Sci Rep 13, 13227 (2023). https://doi.org/10.1038/s41598-023-40514-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-40514-w
- Springer Nature Limited