
Abstract
The objective of this study was to evaluate the impact of dietary zinc supplementation in pre-weaned dairy calves on the phenotypic antimicrobial resistance (AMR) of fecal commensal bacteria. A repository of fecal specimens from a random sample of calves block-randomized into placebo (n = 39) and zinc sulfate (n = 28) groups collected over a zinc supplementation clinical trial at the onset of calf diarrhea, calf diarrheal cure, and the last day of 14 cumulative days of zinc or placebo treatment were analyzed. Antimicrobial susceptibility testing was conducted for Enterococcus spp. (n = 167) and E. coli (n = 44), with one representative isolate of each commensal bacteria tested per sample. Parametric survival interval regression models were constructed to evaluate the association between zinc treatment and phenotypic AMR, with exponentiated accelerated failure time (AFT) coefficients adapted for MIC instead of time representing the degree of change in AMR (MIC Ratio, MR). Findings from our study indicated that zinc supplementation did not significantly alter the MIC in Enterococcus spp. for 13 drugs: gentamicin, vancomycin, ciprofloxacin, erythromycin, penicillin, nitrofurantoin, linezolid, quinupristin/dalfopristin, tylosin tartrate, streptomycin, daptomycin, chloramphenicol, and tigecycline (MR = 0.96–2.94, p > 0.05). In E. coli, zinc supplementation was not associated with resistance to azithromycin (MR = 0.80, p > 0.05) and ceftriaxone (MR = 0.95, p > 0.05). However, a significant reduction in E. coli MIC values was observed for ciprofloxacin (MR = 0.17, 95% CI 0.03–0.97) and nalidixic acid (MR = 0.28, 95% CI 0.15–0.53) for zinc-treated compared to placebo-treated calves. Alongside predictions of MIC values generated from these 17 AFT models, findings from this study corroborate the influence of age and antimicrobial exposure on phenotypic AMR.
Similar content being viewed by others


Introduction
Antimicrobial resistance (AMR) is one of the most significant public health threats faced in this century1. The global challenge to address AMR has prompted studies to better ascertain its distribution across diverse host populations and environments, to infer the pathways of its spread, and to identify the risk factors associated with its emergence and persistence. Comparably, understanding of specific factors that modulate AMR and the extent to which they promote or decrease resistance, particularly in food-producing animals, remains limited.
The occurrence of AMR in food-producing animals not only limits therapeutic options that compromises animal health, but also raises concerns of the potential for its dissemination through animal populations, environmental matrices, and the food chain2. In dairy cattle, it has been well documented that a higher frequency of AMR is typically observed in calves compared to older cattle3,4,5,6. This observed difference in resistance between younger and older cattle may partially be attributed to factors such as increased susceptibility to disease during early life, dietary changes, and initial exposure to the environment and antimicrobial treatments7,8. Improving our understanding of AMR dynamics during early calf life is thus of considerable importance as this period presents an opportunity to reduce AMR acquisition in bovine hosts and their environments.
Diarrhea is the leading cause of morbidity and mortality in pre-weaned dairy calves, with causative agents including viral (e.g. bovine rotavirus), parasitic (e.g. Cryptosporidium parvum), and bacterial pathogens (e.g. Salmonella enterica)9. In diarrheic calves, the frequency of co-infection with more than one pathogen and increased susceptibility to bacteremia from compromised small intestinal function makes antimicrobial therapy a critical and frequent component of treatment protocols10,11; the USDA reported in 2014 that 21.1% of pre-weaned heifers were affected with diarrhea or other digestive disorders, of which 75.9% were treated with antimicrobials12. To ensure animal health and mitigate AMR, alternatives such as zinc supplementation has been explored13,14. Zinc is a trace mineral with a crucial role in many aspects of cellular metabolism and a key constituent of over 300 structural or catalytic enzymatic functions15. It also has an essential role in cellular signal transduction, cell proliferation and differentiation, and regulation of innate and adaptive immunity15,16. Deficiency compromises mechanisms involved in pathogen neutralization and alters cytokine production to impact inflammatory responses, resulting in increased risk for disease and infection in hosts15. The immunity and gut integrity benefits of zinc makes its application in dairy calves of particular interest to simultaneously reduce incidence of diarrheal diseases, improve calf health and growth, and decrease antimicrobial use13,17,18,19.
In a previously conducted double-blind, block-randomized, placebo-controlled clinical trial, zinc supplementation was shown to be effective in delaying diarrheal onset and expediting diarrhea recovery14. However, the utility of zinc supplementation may have unintended consequences as heavy metal exposure has been suggested to enhance the spread of AMR due to the genetic and physiological linkages between metal resistance and AMR. Mechanistically, co-selection for metal resistance and AMR could putatively arise through cross-resistance (one gene/mechanism confers both metal resistance and AMR), co-regulation (the expression of transcriptionally linked metal resistance and AMR systems are modulated by a common gene/regulator), and/or co-resistance (metal resistance and AMR genes occur on the same genetic element)20. Studies evaluating the relationship between dietary zinc and AMR have primarily focused on swine, with several studies providing supporting evidence for the selection of multidrug resistance and increase in occurrence and abundance of AMR genetic determinants from zinc supplementation21,22,23,24. In bovine hosts, studies on dietary zinc and AMR have focused on beef cattle with mixed findings of no AMR selection25,26 or increased AMR to certain antimicrobials in enteric bacteria27. Currently, little is known on the effect of dietary zinc supplementation on AMR in calves. Additionally, the influence on AMR from other contributing factors in conjunction with dietary zinc remains unclear.
To address these knowledge gaps, the aim of this study was to evaluate the impact of dietary zinc supplementation in pre-weaned dairy calves on the phenotypic AMR of fecal commensal bacteria. The objectives were to determine the association between zinc supplementation and AMR in E. coli and Enterococcus spp. and quantify changes in resistance with respect to dietary zinc exposure and calf-level factors.
Materials and methods
The original trial procedures that generated the repository of fecal samples for the current study were approved by the University of California Davis Institutional Animal Care and Use Committee (protocol number 18067 Approved: March 6, 2014) and performed in accordance with relevant guidelines and regulations. The trial that generated the sample repository was a double-blind, block randomized, placebo-controlled clinical trial evaluating different dietary zinc supplementation treatments and their effect on diarrhea prevention and health in pre-weaned calves. The trial was conducted on a single San Joaquin Valley dairy (Kings County, CA, USA). The detailed study and sampling design were previously described14. Briefly, healthy Holstein heifer or bull calves from the one dairy were enrolled at birth (24–48 h of age) and block-randomized by time of birth to zinc or placebo treatment groups. Calves were examined by a veterinarian or trained personnel and excluded if exhibiting obvious morbidities or congenital defects14. A random sample of enrolled calves (approximately 8–10% of the study population given budgetary constraints for sample testing) were selected for fecal sampling. Calves analyzed for this study included all sampled calves (n = 67) in the zinc sulfate (n = 28) and placebo (n = 39) treatment groups that had onset of diarrhea at some point over the duration of the clinical trial.
Pre-weaned calf management and treatment protocols
All calves were under the same management practices including housing and diet14. Throughout the study, calves were housed in individual metal hutches. The respective treatments were administered in the morning milk bottles for 14 days starting from enrollment; calves in the zinc sulfate treatment group received 0.22 g zinc sulfate monohydrate (80 mg of elemental zinc) (Sigma-Aldrich Company, St. Louis, MO, United States) with 0.44 g milk replacer powder, while those in the placebo group received only milk replacer powder (0.44 g). Antimicrobials included in dietary milk for calves 0–25 days of age included chlortetracycline hydrochloride (Pennchlor 64, Pharmgate Animal Health, Omaha, NE, United States) and neomycin sulfate (NeoMed 325, Bimeda, Inc., Le Sueur, MN, United States). Calves 25 days of age to weaning received dietary milk that included oxytetracycline hydrochloride (NT-10G, Agri-Best™, Strauss Feeds LLC, Watertown, WI, United States). Calves from which samples were employed for the current study were all treated for symptoms of diarrhea at some point over the duration of the trial and received the same treatment consisting of an oral mixture of 118.5 mL bismuth subsalicylate (Bismusal Suspension, Durvet, Inc., Blue Springs, MO, United States) and 31.5 mL of spectinomycin (SpectoGard, Bimeda, Inc., Le Sueur, MN, United States)14.
Fecal sample collection
Fecal samples were collected on the first day of calf diarrhea onset (D1), exit or cure from diarrhea (Dex), and on the last day of the 14-day treatment period (D14). Sampling was conducted only at two time points for calves where onset of diarrhea occurred on the same day as the last day of the treatment period (D1,D14). Sample collection was conducted as previously described; new gloves and sterile lubricant were used to collect fresh feces (~ 5 g) into 20 mL polypropylene jars (The Cary Company, QAddison, IL) by digital rectal simulation14. Fecal samples were stored at − 20 °C until analysis.
Bacterial isolation
Fecal samples were processed for bacterial isolation by enriching a saturated fecal cotton swab in 10 mL of Tryptic Soy Broth (TSB, Becton Dickinson, Franklin Lakes, NJ, United States) for 24 h. Two 1 mL aliquots from each TSB enrichment were transferred to two tubes, one containing 9 mL of MacConkey and another containing 9 mL of Enterococcosel broth and were incubated for 24 h at 35 °C and 45 °C, respectively. A loopful (10 µL) of MacConkey and Enterococcosel broth was streaked to MacConkey and Enterococcosel agar for isolation of E. coli and Enterococcus spp., respectively. One putative colony of E. coli and Enterococcus spp. based on typical morphology (pink colony for E. coli and beige colony with strong black halo for Enterococcus spp.) was randomly selected from each selective agar plate, and streaked to purity by subculturing on respective selective agars then to blood agar plates (Tryptic Soy Agar with 5% sheep blood, Thermo Scientific, Waltham, MA, United States). All incubation steps for agar plates were performed at 35 ± 2 °C for 18–24 h.
Bacterial confirmation
DNA was extracted from pure cultures of E. coli and Enterococcus spp. on blood agar plates using a boiling method. Briefly, colonies were suspended in 100 µL of molecular grade water in a sterile 1.5 mL microcentrifuge tube, incubated on a heating block at 100 °C for 20 min, and then centrifuged for 10 min at 5000 rpm28. Confirmation of isolates was then conducted as previously described using conventional PCR to screen for the presence of universal stress protein uspA for E. coli29,30, and 23S rRNA sequence using forward (ECST784F) and reverse (ENC854R) primers for Enterococcus spp31,32. Primer sequences used are provided in Supplementary Table 1. During the PCR step, a positive control (E. coli ATCC 25922 or Enterococcus faecalis ATCC 29212) and two negative controls comprised of a molecular grade water and PCR mastermix blank were used. Confirmed E. coli and Enterococcus spp. isolates were stored in cryovials containing TSB with 15% glycerol at − 80 °C for further testing.
Antimicrobial susceptibility testing (AST)
Antimicrobial susceptibility testing (AST) was conducted on E. coli isolates from D14 fecal samples and 167 enterococci isolates from fecal samples collected at all sample collection time points using the NARMS Gram negative (YCMV3AGNF) and Gram positive (YCMV3AGPF) AST panels, respectively5. Briefly, 3–5 colonies of fresh, pure culture were inoculated into 5 mL of dH2O. Turbidity of the suspension was adjusted to 0.5 McFarland standard, or the equivalent of a 0.08–0.1 OD at 625 nm. From the suspension, 10 µL was then transferred to 11 mL of Sensititre Mueller–Hinton Cation adjusted broth to yield 1 × 105 CFU/mL of bacterial suspension. Fifty microliters of the Mueller–Hinton inoculum were then transferred into each well of the AST plate and incubated at 35 °C for 24 h. E. coli ATCC 25922 and Enterococcus faecalis ATCC 29212 were used as quality controls. Minimum inhibitory concentration (MIC) values were recorded as the lowest concentration of each antimicrobial drug (AMD) which inhibited visible growth of bacteria. The antimicrobial drugs in each panel and their corresponding dilution ranges (µg/mL) are provided in Supplementary Table 2–3.
Statistical analysis
-
a.
All statistical analyses were conducted using Stata 17.0. Data visualization was conducted in RStudio (ggplot2) and BioRender
-
b.
Calf-level factors
Descriptive statistics of enrolled calves for this study and the characteristics of E. coli and Enterococcus spp.—with each isolate representing the status of a calf from which the fecal isolate was collected from—were determined based on daily assessment records of individual calves. This included treatment group (whether an isolate originated from a fecal sample collected from a placebo or zinc sulfate treated calf), and calf-level factors of age, diarrhea status, and therapeutic spectinomycin exposure at the time of fecal collection. Different specifications of calf-level covariates were evaluated. For the age of the calf, this included age in days and age by week. Diarrhea status was specified as days on or from diarrhea (positive values indicating days of ongoing diarrhea and negative values indicating days post diarrhea recovery), categorical diarrhea status (isolate collected from a calf that was pre-diarrheic, diarrheic, or recovered from diarrhea). Exposure to spectinomycin was specified as the number of days from the most recent treatment, the number of doses received, whether treatment was received or not, and a categorical variable of days from the most recent treatment (E. coli: 0 days, 3–5 days, 6–8 days, and 9–10 days; Enterococcus spp.: 0 days, 1–3 days, 4–7 days, 8–23 days).
-
c.
Accelerated failure time (AFT) models
To evaluate the association between isolates from zinc/placebo treatment groups and the degree of change in MIC values, an accelerated failure time (AFT) model was constructed for each antimicrobial drug. The AFT model is a parametric survival model that can be represented by:
For this study, the AFT model was adapted such that the dependent variable Ti, typically representing time-to-event, is specified as left (≤ AMDmin μg/mL), right (> AMDmax μg/mL), or interval censored (between two twofold tested antimicrobial drug concentrations) MIC data, where AMDmin and AMDmax represent the lowest and highest antimicrobial drug concentrations tested, respectively. As negative MIC values are not biologically possible, all left-censored MIC values were specified with a lower bound of 0 instead of − ∞. The relationship between Ti—the progression in expected concentration to inhibition for an antimicrobial for the ith isolate—and a random error term (εi) are assumed to follow a specified distribution. Fixed effects are denoted by the intercept (β0) and Xiβi representing covariates (Xi) and their corresponding regression coefficients (βi). To account for confounding from the biological influence of age and therapeutic antimicrobial treatment for diarrhea, calf age and exposure to spectinomycin were controlled for by forced inclusion as independent variables in all Enterococcus spp. models. Additionally, repeated measures from Enterococcus spp. collected at multiple time points were accounted for by calculating robust standard error estimates using a clustered sandwich variance estimator, which relaxed the assumption of independence among observations and allowed for intragroup correlation of isolates from the same calf33,34. As all E. coli isolates evaluated were recovered from fecal samples collected at the last day of the 14-day zinc or placebo treatments since calf enrollment (1–2 days of age), all E. coli isolates corresponded to calves of similar age (14–16 days), and only exposure to spectinomycin was controlled for in all E. coli models.
-
d.
AFT model specification
A manual forward model building approach was used, in which a base model with treatment group as the only independent variable was first evaluated based on BIC estimates to specify a best-fitting (lowest BIC) distribution (Weibull, exponential, or generalized gamma) for model building. The final model with addition of confounders (spectinomycin exposure for E. coli models and spectinomycin exposure and calf age for Enterococcus spp. models) was identified by evaluating the various specifications for calf-level covariates and using the method of change in estimates to assess confounding. Other calf-level covariates (e.g. diarrhea status) were evaluated and retained in the model if improving model fit. The significance of biologically relevant interactions, specifically calf age—which was collinear with the days of zinc/placebo treatment exposure—and treatment group was tested for all Enterococcus spp. models to assess if a dose-dependent treatment effect was present. The specified parametric distribution for each model was also re-evaluated after tentative multivariable models were constructed, with Wald tests used to confirm that the appropriate parametric model was selected. Lastly, the final model for each antimicrobial drug was selected on the criteria that it generated estimates within the hypothetical maximum concentration limit for the MIC (1 × 106 µg/mL)35,36. In favor of more parsimonious model selection, the Bayesian Information Criteria (BIC) was used to assess competing models, with lower values indicating better model fit. A 5% significance level was used for all models.
Results
Study population and characteristics of E. coli and Enterococcus spp.
Fecal samples from randomly selected pre-weaned dairy calves (28 calves from the zinc sulfate treatment group and 39 calves from the placebo treatment group) from a previously conducted zinc supplementation trial were assessed in this study14. Due to the observational nature of sample collection, fecal collection occurred across four possible time sequences as detailed in the study schematic (Fig. 1). Characteristics of pre-weaned dairy calves whose samples were identified for the current study are summarized in Table 1; no significant differences in sex, attitude score, fecal score, and age were observed between calves in the zinc sulfate and placebo treatment groups from which fecal samples were utilized in the present study.

Schematic of fecal collection and analysis of samples collected from pre-weaned dairy calves enrolled in a zinc supplementation clinical trial.
A total of 44 E. coli isolates from D14 fecal samples and 167 Enterococcus spp. from fecal samples collected at all of the aforementioned time points were included for antimicrobial susceptibility testing. Due to both budgetary constraints for laboratory testing and lower recovery of E. coli isolates from fecal samples collected at other time points, analysis of E. coli was restricted to isolates from D14 samples. As one representative isolate was tested per fecal sample, each commensal isolate represents the AMR status of a calf at the corresponding time point of fecal collection. Of the 44 E. coli isolates, 18 (40.91%) and 26 (59.09%) were from zinc and placebo treated calves, respectively. The average age of calves that E. coli isolates were collected from was 15.7 days (SD 0.51). Of the E. coli isolates, 15.91% corresponded to fecal samples from calves which received no spectinomycin treatment, 15.91% received 1 dose, and 68.18% received 2 doses. Further, 25%, 11.36%, and 63.64% of E. coli isolates were from healthy, diarrheic, and diarrhea-recovered calves, respectively. For the 167 enterococci isolates collected through repeated sampling of calves, 67 (40.12%) and 100 (59.88%) were from zinc and placebo treated calves, respectively. The average age of calves which enterococci isolates were collected from was 14.34 days (SD 5.47). Further, 28.74%, 17.37%, 51.50%, and 2.4% of enterococci isolates corresponded to calves having received 0, 1, 2, and 3 doses of spectinomycin treatment, respectively; while 6.59%, 31.74%, 39.52%, and 22.16% were from healthy, day of diarrhea diagnosis, diarrheic, and diarrhea-recovered calves, respectively. Characteristics of commensal isolates by treatment group are summarized in Fig. 2.

Characteristics of E. coli (n = 44) and Enterococcus spp. (n = 167) from pre-weaned dairy calves at the time of fecal sample collection by placebo and zinc treatment groups (A) Fecal sample type (B) Age of calf (C) Number of days from recent therapeutic spectinomycin treatment (D) Number of therapeutic spectinomycin treatments received (E) Days from diarrhea onset (positive numbers) and from diarrheal cure (negative numbers) and (F) Diarrhea disease status.
Accelerated failure time (AFT) models
Due to the high frequency of right-censored antimicrobial susceptibility testing (AST) data, certain antimicrobial drugs were determined to be unsuitable for modelling given data sparseness and omitted from analysis. This included tetracycline, sulfisoxazole, trimethoprim-sulfamethoxazole, ampicillin, streptomycin, cefoxitin, chloramphenicol, amoxicillin-clavulanic acid, gentamicin, and ceftiofur for E. coli; and tetracycline, kanamycin, and lincomycin for enterococci isolates. The distribution of minimum inhibitory concentration (MIC) values and descriptive statistics (MIC50 and MIC90) for all tested drugs from AST for E. coli and Enterococcus spp. isolates are presented in Supplementary Figs. 1–6 and Supplementary Tables 4 and 5.
A total of 17 accelerated failure time (AFT) models, consisting of models for 4 antimicrobial drugs for E. coli (Supplementary Table 6–9) and 13 antimicrobial drugs for Enterococcus spp. (Supplementary Tables 10–22), were constructed. For E. coli, the ciprofloxacin model with spectinomycin exposure specified as the number of doses (BIC 189.5612) was selected as the final model over the competing model with a binary spectinomycin treated or not variable (BIC 188.6194) despite having a lower BIC in the latter given the significant coefficient for the number of doses variable (Supplementary Table 7). For Enterococcus spp., the generalized gamma model for streptomycin (BIC 260.1047, Supplementary Table 13) was selected over its Weibull model counterpart (BIC 249.5898) due to estimates from the latter model exceeding the hypothetical maximum concentration for MIC; for the same reason, the exponential model for gentamicin (BIC 524.5724, Supplementary Table 22) was selected as the final model over its Weibull model counterpart (BIC 268.565). Lastly, the Enterococcus spp. linezolid model with categorical number of days from last spectinomycin treatment (BIC 303.3108) was selected as the final model over the one with a binary spectinomycin (treated or not) variable (BIC 299.7655) given the significant coefficient for the categorical variable (Supplementary Table 16).
Impact of zinc supplementation on AMR of fecal E. coli and Enterococcus spp.
For each AFT model, the exponentiated coefficient for the primary effect of treatment group (MIC Ratio, or MR) represents the change in MIC values associated with treatment status, in which a MR > 1 indicates that zinc supplementation was associated with an increase in phenotypic AMR whereas a MR < 1 indicates that zinc supplementation was associated with a decrease in phenotypic AMR35,36. Overall findings from AFT models indicated that zinc supplementation was not significantly associated with the MIC of Enterococcus spp. to 13 drugs: gentamicin, vancomycin, ciprofloxacin, erythromycin, penicillin, nitrofurantoin, linezolid, quinupristin/dalfopristin, tylosin tartrate, streptomycin, daptomycin, chloramphenicol, and tigecycline (MR = 0.96–2.94, p > 0.05) (Fig. 3, Table 2, and Supplementary Tables 10–22). In E. coli, no significant association was observed between zinc supplementation and resistance of isolates as measured using MIC values for ceftriaxone (MR = 0.95, p > 0.05) and azithromycin (MR = 0.80, p > 0.05) with the exception of ciprofloxacin (MR = 0.17, 95% CI 0.03–0.97) and nalidixic acid (MR = 0.28, 95% CI 0.15–0.53) where a protective effect against resistance was observed (Fig. 3, Table 3, and Supplementary Tables 6–9). No significant dose-dependent effect of zinc supplementation was observed across all antimicrobial drugs evaluated for E. coli and Enterococcus spp..

Treatment group minimum inhibitory concentration ratio (MR) estimates and corresponding 95% confidence intervals from accelerated failure time models, after controlling for confounders by (A) Enterococcus spp. and (B) E. coli.
Therapeutic antimicrobial treatment and calf age were significantly associated with AMR
In this study, therapeutic spectinomycin treatment of pre-weaned dairy calves and calf age were significantly associated with AMR for certain antimicrobial drugs in fecal commensal isolates. The effect measures of these confounders represented the corresponding direct effects on AMR when other covariates were held constant; as indirect effects were not accounted for, it should be noted that the magnitude and direction of these associations were not necessarily representative of total effect estimations. Spectinomycin exposure was significantly associated with an increase in MIC for azithromycin, ciprofloxacin, and nalidixic acid in E. coli (Supplementary Tables 6, 7, and 8) and tigecycline and linezolid in enterococci isolates (Supplementary Tables 10 and 16). For Enterococcus spp., it was also significantly associated with a decrease in MIC for tylosin tartrate, nitrofurantoin, erythromycin, and gentamicin (Supplementary Tables 14, 17, 19, and 22). Calf age at time of sampling was found to be significantly associated with AMR in Enterococcus spp. models only; as all E. coli isolates were from calves of similar age (14–16 days), age was not evaluated in E. coli models. Age was a significant predictor in 5 of the 13 Enterococcus spp. models, in which it was significantly associated with a decrease in MIC for tigecycline, nitrofurantoin, penicillin, ciprofloxacin, and gentamicin (Supplementary Tables 10, 17, 18, 20, and 22).
Predictions of phenotypic antimicrobial resistance
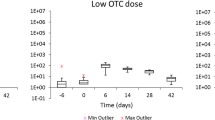
To further understand the dynamics between zinc supplementation and AMR, predictions of MIC from AFT models for Enterococcus spp. (Fig. 4) and E. coli isolates (Fig. 5) were assessed. The predicted MIC for enterococci isolates after controlling for calf age was higher in zinc-treated compared to placebo-treated calves across all levels of spectinomycin exposure (Fig. 4, Supplementary Tables 27–39). Predicted MIC decreased after spectinomycin exposure for tylosin tartrate, gentamicin, chloramphenicol, erythromycin, daptomycin, nitrofurantoin, and penicillin and increased for ciprofloxacin, tigecycline, quinupristin/dalfopristin, streptomycin, and vancomycin (Fig. 4). In contrast to predictions from Enterococcus spp. models, predictions from E. coli models indicated that across all levels of spectinomycin exposure, isolates from placebo-treated calves had higher MIC than those from zinc-treated calves for all antimicrobial drugs (Fig. 5, Supplementary Tables 23–26). Differences in MIC were statistically significant between treatment groups for predicted E. coli resistance to nalidixic acid for calves that received 2 doses of spectinomycin treatment; E. coli from calves in the placebo group that received 2 doses of spectinomycin treatment on average had 4.76 µg/mL higher MIC for nalidixic acid compared to those from calves in the zinc treatment group that received the same number of spectinomycin doses (Fig. 5, Supplementary Table 25). Following spectinomycin exposure, the predicted E. coli MIC increased for ceftriaxone (Fig. 5, Supplementary Table 26). For azithromycin, the predicted MIC for E. coli exhibited a cyclical trend in days post spectinomycin treatment in which the MIC increased (3–5 days), decreased (6–8 days) and then increased (9–10 days) (Fig. 5, Supplementary Table 23). Across all models, zinc supplementation and therapeutic spectinomycin were not predicted to increase or decrease the MIC to above or below the CLSI resistance breakpoints of tested antimicrobials.

Predicted minimum inhibitory concentrations (MIC, µg/mL) and corresponding 95% confidence intervals for Enterococcus spp. accelerated failure time (AFT) models for each antimicrobial drug model and by therapeutic spectinomycin treatment status, controlling for calf age. Dashed lines mark resistant breakpoints based on CLSI criteria for each antimicrobial drug.

Predicted minimum inhibitory concentration (MIC, µg/mL) and corresponding 95% confidence intervals for E. coli accelerated failure time (AFT) models for each antimicrobial drug model, by therapeutic spectinomycin treatment status. Dashed lines mark resistant breakpoints based on CLSI criteria for each antimicrobial drug.
Discussion
The development of AMR is frequently attributed to use of antimicrobials, but there is increasing recognition that non-antimicrobial agents (e.g. biocides, antiseptics, and heavy metals) may also play a role in the modulation of AMR37,38. The large-scale application of these agents in agricultural and livestock production in particular, may pose considerable impact from veterinary medicine, environmental, and public health standpoints; hence, better understanding of non-antimicrobial influences on AMR dynamics in these systems is of great importance. The present study investigated the impact of dietary zinc supplementation in pre-weaned calves on phenotypic AMR using fecal E. coli and Enterococcus spp. as Gram-negative and Gram-positive indicator organisms, respectively.
In this study, zinc supplementation did not significantly change the MIC to all antimicrobials tested for E. coli and Enterococcus spp. with the exception of nalidixic acid and ciprofloxacin, in which zinc supplementation was associated with a protective effect against quinolone resistance in E. coli. Although the temporality of exposure to zinc preceding resistance cannot be definitively established from our study data due to the cross-sectional nature of evaluating E. coli isolates collected at one time point, a previous study evaluating the in vitro selection dynamics of zinc on ciprofloxacin provides evidence in agreement with findings in this study39. In laboratory conditions of 0.00625 µg/mL ciprofloxacin and no zinc, ciprofloxacin-resistant E. coli was shown to have a significant growth rate advantage relative to a ciprofloxacin-susceptible strain, with this advantage disappearing in the presence of zinc sulfate (0.5 mM and 1.0 mM) at the same ciprofloxacin concentration39. Vos et al. further showed through a competition assay that ciprofloxacin-susceptible E. coli outcompetes its resistant counterpart under conditions of 0.5 mM zinc sulfate and 0.025 µg/mL ciprofloxacin39. While additional work is needed to fully elucidate the role of zinc on quinolone resistance, these congruent findings collectively support the potential antagonistic effects of zinc sulfate on E. coli resistance to certain antimicrobial classes.
Limited studies have evaluated the impact of dietary zinc on AMR in calves, with existing zinc supplementation studies conducted in other livestock and evaluating different zinc concentrations. The concentration of zinc fed once per day through milk to pre-weaned dairy calves in this study was 0.22 g zinc sulfate monohydrate, or 80 mg of elemental zinc. Such a non-toxic zinc dose for calves was previously established through a clinical trial13, which alongside the field trial that contributed samples for the present analysis and other independent studies, collectively found beneficial impacts of zinc supplementation on the growth performance and diarrheal recovery in pre-weaned calves14,19,40. The lack of evidence for the promotion of AMR by dietary zinc in this study is consistent with a factorial trial on feedlot cattle that found copper and/or zinc supplementation (30 or 300 mg/kg) over a 32-day period resulted in minimal changes to MICs of copper, zinc, and antibiotics of fecal E. coli and Enterococcus spp25. These results are further corroborated through another feedlot cattle study, where quantified resistance to ceftriaxone and tetracycline among enteric E. coli were unaffected by dietary zinc concentrations (0–90 mg/kg)26. In contrast, one feedlot cattle study found that dietary zinc was associated with a significant increase in macrolide resistance of fecal enterococci isolates27. The relationship between zinc supplementation and AMR has been more extensively studied in swine, where concentrations of > 2000 mg/kg are typically used in feed additives to prevent post-weaning diarrhea and to enhance growth performance41. In contrast to findings from dietary zinc in cattle, multiple studies on swine have found that high dietary zinc may promote AMR, with one study finding that dietary zinc oxide (2,103 mg/kg) increased multi-resistant E. coli in feces, digesta, and mucosa21; and another finding an increased proportion of multi-resistant E. coli in ileum and colon digesta of between zinc oxide (18.6%; 2500 mg/kg) and control (0%; 50 mg/kg) groups22. The conflicting findings across these studies suggests that the modulation of AMR by zinc may be in part, or in combination with other unidentified factors, affected by: (1) the host site of bacterial isolation, (2) the duration and dose of supplementation, and (3) the form of zinc (organic/inorganic) administered. In cattle and swine, the absorption of trace minerals such as zinc occurs primarily in the small intestine, with endogenous zinc loss mainly occurring through excretion of feces42,43. Differences in the bioavailability and dose of administered zinc may result in variable quantities and duration of zinc presence in the intestinal lumen, which could subsequently exert disparate effects on the gut resistome. Zinc absorption and excretion are homeostatically regulated, with a previous study finding that pigs fed 2000 ppm zinc did not become zinc loaded and excrete increased quantities of zinc in feces until after 13 to 14 days of supplementation44,45. Although the digestive tracts of swine (non-ruminant) and cattle (ruminant) at maturity are structurally different, the gastrointestinal (GI) tract of pre-weaned calves is functionally monogastric like that of swine. These GI physiological similarities and the same duration of zinc supplementation evaluated in this study (14 days) suggest that improved health outcomes of pre-weaned calves from zinc supplementation (80 mg elemental zinc) in this study likely occurred with minimal excess zinc excretion.
The age-dependent acquisition and loss of AMR in pre-weaned dairy calves has been well documented to show transient increases in AMR during early calf life3,5,7,46,47,48,49, which corroborate findings from this study. Higher levels of multiple resistance in fecal E. coli isolates were observed in calves 2 weeks of age compared to day-old calves (OR = 53.6)3. Additionally, a case–control study evaluating the occurrence of AMR in fecal E. coli from diarrheic and healthy dairy calves found higher prevalence of resistance to ampicillin, tetracycline, or sulfonamide (p ≤ 0.05) in diarrheic calves50. Similarly, this study found high phenotypic AMR (right-censoring of MIC data) in diarrhea-associated E. coli isolates collected from calves around 2 weeks of age, which limited our ability to model AMR dynamics of several antimicrobial drugs for E. coli. In enterococci isolates, age was significantly associated with a decrease in MIC for 5 of the 13 antimicrobials evaluated after accounting for zinc and spectinomycin exposure. The age-dependent trends in calf AMR has been thought to be driven by host immune status of increased susceptibility during early life that allows ease of colonization by resistant bacteria, exposure through the environment and dam (e.g. colostrum), and rapid establishment of the early gut microbiome7,8. A previous study demonstrated that the acquisition of the resistome throughout calf life occurs even in the absence of antimicrobial exposure8. Liu et al. also demonstrated that facultative anaerobes such as Escherichia and Enterococcus, which are frequent carriers of AMR, constitute greater portions of the fecal microbiota during early life but decrease in relative abundance over time8. Calf diarrhea has been shown to result in further perturbation of the gut microbiota, with multiple studies observing shifts or differences in gut microbial composition between healthy and diarrheic calves51,52,53,54. These findings collectively support the hypothesis that stabilization and maturation of the calf gut microbiota with age may impart differential fitness costs for resistance, providing susceptible bacteria a competitive advantage and ultimately leading to a decrease in the resistant enteric microbial population as observed in Enterococcus spp. in our study.
Although this study was unable to evaluate the effects of zinc supplementation in absence of antimicrobial exposures, these data provide practical insights on the combined effects of antimicrobial treatment, dietary heavy metal exposure, and host-level factors on phenotypic AMR, interactions that remain undercharacterized. The AMR selective pressures in our study were from dietary milk (same concentration and duration of tetracycline and neomycin for all calves) and therapeutic spectinomycin (same dosage but varied in time and duration of treatment for each calf) which were reflected in the high right-censoring to tested drugs from the same antimicrobial class in our E. coli and Enterococcus spp. study isolates. These broad-spectrum antibiotics correspond to tetracycline and aminoglycoside/aminocyclitol classes and are commonly used for the treatment of diarrhea in calves; USDA reports from 2014 indicate that of pre-weaned heifers treated with antimicrobials for diarrhea and digestive disorders, aminoglycoside and tetracycline were the third and fourth most commonly administered primary antimicrobials, with 14.7% and 11.2% of calves with these diseases treated with drugs corresponding to these respective classes12. The other reported antimicrobials frequently used—third-generation cephalosporins (27.6%) and trimethoprim/sulfamethoxazole (18.7%)12—were not administered to calves in our study. Despite this, we also observed right-censored or elevated MICs to these drugs (trimethoprim/sulfamethoxazole and ceftriaxone) in our data, indicating the occurrence of AMR selection in absence of direct selective pressures. Regarding the differential exposure of therapeutic spectinomycin for each calf that was controlled for in all models, neither zinc supplementation nor spectinomycin modulated the predicted MIC of all tested antimicrobials to above or below CLSI resistant breakpoints. These predictions from AFT models allowed for the quantification of absolute changes in resistance to better understand AMR selection dynamics. For example, model predictions indicated that after receiving two doses of therapeutic spectinomycin, E. coli from placebo-treated calves had a significantly higher nalidixic acid MIC relative to those from zinc-treated calves. These predictions support aforementioned in vivo findings that the putative inhibitory effects of zinc on quinolone resistance may occur in combination with antimicrobial selective pressures39. Antimicrobial therapy has been shown to transiently modify the calf gut microbiota, which may favorably enrich resistant bacteria55,56. Indeed, model predictions from this study indicated that following spectinomycin treatment, MICs to certain antimicrobials increase in both E. coli (ceftriaxone) and Enterococcus spp. (ciprofloxacin, tigecycline, quinupristin/dalfopristin, streptomycin, and vancomycin). However, the decreasing trend in MIC to other antimicrobials—primarily in Enterococcus spp. to tylosin tartrate, gentamicin, chloramphenicol, erythromycin, daptomycin, nitrofurantoin, and penicillin—and the cyclical pattern in azithromycin MIC of E. coli following spectinomycin treatment underscores the complexity of AMR selection. With respect to the latter finding, previous studies have observed similar cyclical resistance trends in fecal Enterobacteriaceae from dairy cattle following antimicrobial treatment. This included one study that observed cyclical peaks in neomycin resistant Enterobacteriaceae counts following treatment with the drug57, and another which observed cyclical re-emergence of ceftiofur-resistant Enterobacteriaceae counts following systemic antimicrobial drug treatment58. In some bacteria, antimicrobial exposure has been shown to induce mutagenesis and conjugation59,60,61, which can be followed by emergence of additional compensatory mutations that alleviate fitness costs associated with resistance (e.g. positive epistasis)62,63. Such variation in fitness costs would support how modifications of the calf enteric microbiota from collective influences of antimicrobial therapy, disease (e.g. diarrhea), and other changes in host-factors culminate a microbial system that is conducive to the divergent and bidirectional selection of bacterial resistance.
In conclusion, the current study found a lack of evidence for the selection of phenotypic AMR to the majority of tested antimicrobial drugs in fecal commensal bacteria from dietary zinc supplementation in pre-weaned dairy calves. The primary limitations of this study include the cross-sectional assessment of E. coli at one time point which limited our ability to determine the temporality of association between dietary zinc treatment and AMR, and the small sample size of E. coli which may impact the reliability of study findings. Additionally, our study may have limited external validity due to samples originating from just one dairy. Future studies conducted across different dairies with varied environmental factors and management practices, with larger sample sizes, and evaluating multiple isolates per sample are needed to validate the effect sizes observed in our study, particularly for the significant findings of the antagonistic association between zinc supplementation and quinolone resistance in E. coli. As our main objective was to assess the impact of dietary zinc on AMR, AFT models specified in this study were not designed to obtain unbiased point estimates on the relationship of calf age and antimicrobial treatment with AMR. Additionally, the extreme model estimates observed in our study for two models (gentamicin and streptomycin for Enterococcus spp.) indicated that AFT models may not be optimal for data with a high frequency of right- and left- censoring; hence, alternative approaches such as truncated interval censored models should be explored for these types of data in future studies. Further research that evaluates a longer follow-up period (e.g. through post-weaning), the impact of other forms of dietary zinc (e.g. chelated zinc), and different resistance outcomes are needed. Additionally, genomic analysis of isolates in this study would provide additional insight on the AMR dynamics of dietary zinc supplementation, antimicrobial treatment, and disease status in pre-weaned dairy calves. Collectively, this work corroborates previous reports of host-level drivers (e.g. age and antimicrobial exposure) on AMR, and provides important insights into the largely unexplored dynamics of dietary zinc and AMR in pre-weaned dairy calves.
Data availability
Data from this study are presented in the Supplementary File. Additional source data are available from the corresponding author upon reasonable request.
References
Murray, C. J. et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 399, 629–655 (2022).
Economou, V. & Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 8, 49–61 (2015).
Berge, A. C. B., Atwill, E. R. & Sischo, W. M. Animal and farm influences on the dynamics of antibiotic resistance in faecal Escherichia coli in young dairy calves. Prev. Vet. Med. 69, 25–38 (2005).
Berge, A. C. B., Moore, D. A. & Sischo, W. M. Field trial evaluating the influence of prophylactic and therapeutic antimicrobial administration on antimicrobial resistance of fecal Escherichia coli in dairy calves. Appl. Environ. Microbiol. 72, 3872–3878 (2006).
Li, X. et al. Phenotypic antimicrobial resistance profiles of E. coli and Enterococcus from dairy cattle in different management units on a Central California dairy. Clin. Microbiol. 07, 2 (2018).
Oh, S.-I., Ha, S., Roh, J.-H., Hur, T.-Y. & Yoo, J. G. Dynamic changes in antimicrobial resistance in fecal Escherichia coli from neonatal dairy calves: An individual follow-up study. Animals 10, 1776 (2020).
Khachatryan, A. R., Hancock, D. D., Besser, T. E. & Call, D. R. Role of calf-adapted Escherichia coli in maintenance of antimicrobial drug resistance in dairy calves. Appl. Environ. Microbiol. 70, 752–757 (2004).
Liu, J. et al. The fecal resistome of dairy cattle is associated with diet during nursing. Nat. Commun. 10, 4406 (2019).
Cho, Y. & Yoon, K.-J. An overview of calf diarrhea—infectious etiology, diagnosis, and intervention. J. Vet. Sci. 15, 1–17 (2014).
Constable, P. D. Antimicrobial use in the treatment of calf diarrhea. J. Vet. Intern. Med. 18, 8–17 (2004).
Constable, P. D. Treatment of calf diarrhea: Antimicrobial and ancillary treatments. Vet. Clin. N. Am. Food Anim. Pract. 25, 101–120 (2009).
USDA. Health and Management Practices on U.S. Dairy Operations, 2014; https://www.aphis.usda.gov/animal_health/nahms/dairy/downloads/dairy14/Dairy14_dr_PartIII.pdf. Accessed 20 October 2023.
Glover, A. D. et al. A double-blind block randomized clinical trial on the effect of zinc as a treatment for diarrhea in neonatal Holstein calves under natural challenge conditions. Prev. Vet. Med. 112, 338–347 (2013).
Feldmann, H. R., Williams, D. R., Champagne, J. D., Lehenbauer, T. W. & Aly, S. S. Effectiveness of zinc supplementation on diarrhea and average daily gain in pre-weaned dairy calves: A double-blind, block-randomized, placebo-controlled clinical trial. PLOS One 14, e0219321 (2019).
Bonaventura, P., Benedetti, G., Albarède, F. & Miossec, P. Zinc and its role in immunity and inflammation. Autoimmun. Rev. 14, 277–285 (2015).
Maywald, M., Wessels, I. & Rink, L. Zinc signals and immunity. Int. J. Mol. Sci. 18, 2222 (2017).
Ma, F. T. et al. Zinc-methionine acts as an anti-diarrheal agent by protecting the intestinal epithelial barrier in postnatal Holstein dairy calves. Anim. Feed Sci. Technol. 270, 114686 (2020).
Liu, J., Ma, F., Degen, A. & Sun, P. The effects of zinc supplementation on growth, diarrhea, antioxidant capacity, and immune function in Holstein dairy calves. Animals 13, 2493 (2023).
Chang, M. N. et al. Effects of different types of zinc supplement on the growth, incidence of diarrhea, immune function, and rectal microbiota of newborn dairy calves. J. Dairy Sci. 103, 6100–6113 (2020).
McNeilly, O., Mann, R., Hamidian, M. & Gunawan, C. Emerging concern for silver nanoparticle resistance in Acinetobacter baumannii and other bacteria. Front. Microbiol. 12, 652863 (2021).
Ciesinski, L. et al. High dietary zinc feeding promotes persistence of multi-resistant E. coli in the swine gut. PLoS One 13, e0191660 (2018).
Bednorz, C. et al. The broader context of antibiotic resistance: Zinc feed supplementation of piglets increases the proportion of multi-resistant Escherichia coli in vivo. Int. J. Med. Microbiol. 303, 396–403 (2013).
Johanns, V. C. et al. Effects of a four-week high-dosage zinc oxide supplemented diet on commensal Escherichia coli of weaned pigs. Front. Microbiol. 10, 2734 (2019).
Vahjen, W., Pietruszyńska, D., Starke, I. C. & Zentek, J. High dietary zinc supplementation increases the occurrence of tetracycline and sulfonamide resistance genes in the intestine of weaned pigs. Gut Pathog. 7, 23 (2015).
Jacob, M. E. et al. Effects of feeding elevated concentrations of copper and zinc on the antimicrobial susceptibilities of fecal bacteria in feedlot cattle. Foodborne Pathog. Dis. 7, 643–648 (2010).
Van Bibber-Krueger, C. L. et al. Effects of supplemental zinc sulfate on growth performance, carcass characteristics, and antimicrobial resistance in feedlot heifers1. J. Anim. Sci. 97, 424–436 (2019).
Murray, S. A. et al. Effects of zinc and menthol-based diets on co-selection of antibiotic resistance among E. coli and Enterococcus spp. in beef cattle. Animals 11, 259 (2021).
Atwill, E. R. et al. Transfer of Escherichia coli O157:H7 from simulated wildlife scat onto romaine lettuce during foliar irrigation. J. Food Prot. 78, 240–247 (2015).
Anastasi, E. M. et al. Prevalence and persistence of Escherichia coli strains with uropathogenic virulence characteristics in sewage treatment plants. Appl. Environ. Microbiol. 76, 5882–5886 (2010).
Chen, J. & Griffiths, M. W. PCR differentiation of Escherichia coli from other Gram-negative bacteria using primers derived from the nucleotide sequences flanking the gene encoding the universal stress protein. Lett. Appl. Microbiol. 27, 369–371 (1998).
Ludwig, W. & Schleifer, K.-H. How quantitative is quantitative PCR with respect to cell counts?. Syst. Appl. Microbiol. 23, 556–562 (2000).
Frahm, E. & Obst, U. Application of the fluorogenic probe technique (TaqMan PCR) to the detection of Enterococcus spp. and Escherichia coli in water samples. J. Microbiol. Methods 52, 123–131 (2003).
STATA. https://www.stata.com/manuals/rvce_option.pdf#rvce_option. Accessed 20 October 2023.
STATA. https://www.stata.com/manuals/u20.pdf#u20.22Obtainingrobustvarianceestimates. Accessed 20 October 2023.
Breen, M. J. et al. Effect of group housing of preweaned dairy calves: Health and fecal commensal antimicrobial resistance outcomes. Antibiotics 12, 1019 (2023).
Okello, E., ElAshmawy, W. R., Williams, D. R., Lehenbauer, T. W. & Aly, S. S. Effect of dry cow therapy on antimicrobial resistance of mastitis pathogens post-calving. Front. Vet. Sci. 10, 1132810 (2023).
Poole, K. At the nexus of antibiotics and metals: The impact of Cu and Zn on antibiotic activity and resistance. Trends Microbiol. 25, 820–832 (2017).
Wales, A. D. & Davies, R. H. Co-selection of resistance to antibiotics, biocides and heavy metals, and its relevance to foodborne pathogens. Antibiotics 4, 567–604 (2015).
Vos, M. et al. Zinc can counteract selection for ciprofloxacin resistance. FEMS Microbiol. Lett. 367, fnaa038 (2020).
Wo, Y. et al. Supplementation with zinc proteinate increases the growth performance by reducing the incidence of diarrhea and improving the immune function of dairy calves during the first month of life. Front. Vet. Sci. 9, 911330 (2022).
Burrough, E. R., De Mille, C. & Gabler, N. K. Zinc overload in weaned pigs: Tissue accumulation, pathology, and growth impacts. J. Vet. Diagn. Invest. 31, 537–545 (2019).
Broom, L. J., Monteiro, A. & Piñon, A. Recent advances in understanding the influence of zinc, copper, and manganese on the gastrointestinal environment of pigs and poultry. Animals 11, 1276 (2021).
Miller, W. J. Zinc nutrition of cattle: A review. J. Dairy Sci. 53, 1123–1135 (1970).
Rincker, M. J., Hill, G. M., Link, J. E., Meyer, A. M. & Rowntree, J. E. Effects of dietary zinc and iron supplementation on mineral excretion, body composition, and mineral status of nursery pigs1,2. J. Anim. Sci. 83, 2762–2774 (2005).
Hill, G. M. & Shannon, M. C. Copper and zinc nutritional issues for agricultural animal production. Biol. Trace Elem. Res. 188, 148–159 (2019).
Cao, H. et al. Age-associated distribution of antimicrobial-resistant Salmonella enterica and Escherichia coli Isolated from dairy herds in Pennsylvania, 2013–2015. Foodborne Pathog. Dis. 16, 60–67 (2019).
Carey, A. M. et al. Prevalence and profiles of antibiotic resistance genes mph(A) and qnrB in extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from dairy calf feces. Microorganisms 10, 411 (2022).
Gaire, T. N., Scott, H. M., Sellers, L., Nagaraja, T. G. & Volkova, V. V. Age dependence of antimicrobial resistance among fecal bacteria in animals: A scoping review. Front. Vet. Sci. 7, 622495 (2021).
Merle, R. et al. The therapy frequency of antibiotics and phenotypical resistance of Escherichia coli in calf rearing sites in Germany. Front. Vet. Sci. 10, 1152246 (2023).
de Verdier, K., Nyman, A., Greko, C. & Bengtsson, B. Antimicrobial resistance and virulence factors in Escherichia coli from Swedish dairy calves. Acta Vet. Scand. 54, 2 (2012).
Gomez, D. E., Arroyo, L. G., Costa, M. C., Viel, L. & Weese, J. S. Characterization of the fecal bacterial microbiota of healthy and diarrheic dairy calves. J. Vet. Intern. Med. 31, 928–939 (2017).
Fan, P. et al. The gut microbiota of newborn calves and influence of potential probiotics on reducing diarrheic disease by inhibition of pathogen colonization. Front. Microbiol. 12, 772863 (2021).
Kim, E.-T. et al. Dynamic changes in fecal microbial communities of neonatal dairy calves by aging and diarrhea. Animals 11, 1113 (2021).
Malmuthuge, N. & Guan, L. L. Understanding the gut microbiome of dairy calves: Opportunities to improve early-life gut health. J. Dairy Sci. 100, 5996–6005 (2017).
Beyi, A. F. et al. Enrofloxacin alters fecal microbiota and resistome irrespective of its dose in calves. Microorganisms 9, 2162 (2021).
Ma, T. et al. Linking perturbations to temporal changes in diversity, stability, and compositions of neonatal calf gut microbiota: Prediction of diarrhea. ISME J. 14, 2223–2235 (2020).
Cella, E. et al. Estimating the rates of acquisition and loss of resistance of enterobacteriaceae to antimicrobial drugs in pre-weaned dairy calves. Microorganisms 9, 2103 (2021).
Sheedy, D. B. et al. Effect of antimicrobial treatment on the dynamics of ceftiofur resistance in enterobacteriaceae from adult california dairy cows. Microorganisms 9, 828 (2021).
Lopatkin, A. J. et al. Antibiotics as a selective driver for conjugation dynamics. Nat. Microbiol. 1, 16044 (2016).
Revitt-Mills, S. A. & Robinson, A. Antibiotic-Induced Mutagenesis: Under the microscope. Front. Microbiol. 11, 585175 (2020).
Ding, M. et al. Subinhibitory antibiotic concentrations promote the horizontal transfer of plasmid-borne resistance genes from Klebsiellae pneumoniae to Escherichia coli. Front. Microbiol. 13, 1017092 (2022).
Marcusson, L. L., Frimodt-Møller, N. & Hughes, D. Interplay in the selection of fluoroquinolone resistance and bacterial fitness. PLOS Pathog. 5, e1000541 (2009).
Melnyk, A. H., Wong, A. & Kassen, R. The fitness costs of antibiotic resistance mutations. Evolut. Appl. 8, 273–283 (2015).
Acknowledgements
K.Y.L. was supported by the Austin Eugene Lyons fellowship from the University of California, Davis. This study was additionally made possible in part by the Western Center for Agricultural Health and Safety (WCAHS) Small Grant Program (Project PI: K.Y.L.) through a larger CDC/NIOSH grant #U54OH007550. We gratefully thank Jennifer Chase and Xiaohong Wei for their technical support in the laboratory.
Author information
Authors and Affiliations
Contributions
Conceptualization: K.Y.L., E.R.A., X.L., B.C.W., and S.S.A. Data collection and curation: K.Y.L., H.R.F., D.R.W., and S.S.A. Laboratory analysis: K.Y.L. Formal analysis: K.Y.L. and S.S.A. Investigation: K.Y.L. and S.S.A. Resources: X.L., E.R.A., and S.S.A. Writing—original draft preparation: K.Y.L. Writing—Review and editing: D.R.W., B.C.W., S.S.A. Visualization: K.Y.L. Project administration: K.Y.L. and S.S.A. Funding acquisition: K.Y.L. All authors have read the article and approved the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lee, K.Y., Atwill, E.R., Li, X. et al. Impact of zinc supplementation on phenotypic antimicrobial resistance of fecal commensal bacteria from pre-weaned dairy calves. Sci Rep 14, 4448 (2024). https://doi.org/10.1038/s41598-024-54738-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-54738-x
- Springer Nature Limited