
Abstract
Vitamin D deficiency is increasingly common in systemic lupus erythematosus (SLE) patients and is associated with the disease activity and proteinuria. Recently, alterations in metabolism have been recognized as key regulators of SLE pathogenesis. Our objective was to identify differential metabolites in the serum metabolome of SLE with vitamin D deficiency. In this study, serum samples from 31 SLE patients were collected. Levels of 25(OH)D3 were assayed by ELISA. Patients were divided into two groups according to their vitamin D level (20 ng/ml). Untargeted metabolomics were used to study the metabolite profiles in serum by high-performance liquid chromatography-tandem mass spectrometry. Subsequently, we performed metabolomics profiling analysis to identify 52 significantly altered metabolites in vitamin D deficient SLE patients. The area under the curve (AUC) from ROC analyses was calculated to assess the diagnostic potential of each candidate metabolite biomarker. Lipids accounted for 66.67% of the differential metabolites in the serum, highlighted the disruption of lipid metabolism. The 52 differential metabolites were mapped to 27 metabolic pathways, with fat digestion and absorption, as well as lipid metabolism, emerging as the most significant pathways. The AUC of (S)-Oleuropeic acid and 2-Hydroxylinolenic acid during ROC analysis were 0.867 and 0.833, respectively, indicating their promising diagnostic potential. In conclusion, our results revealed vitamin D deficiency alters SLE metabolome, impacting lipid metabolism, and thrown insights into the pathogenesis and diagnosis of SLE.
Similar content being viewed by others
Introduction
Systemic lupus erythematosus (SLE) is a chronic autoimmune disease characterized by production of pathogenic antibodies leading to tissue damage in multiple organs, including, but not limited to, the skin, kidney, blood and central nervous system1. The pathogenesis of SLE remains unclear, however, it has been reported that genetic susceptibility, epigenetics and environmental factors are involved in the pathogenesis of SLE2.
While previous research has primarily focused on genomic, transcriptomic, and proteomic changes in SLE, recent attention has turned to the role of metabolomics in autoimmune diseases. Abnormal metabolic pathways of immune cells, such as macrophages, dendritic cells, T cells and B cells, play pivotal roles in the pathogenesis of SLE3. Notably, CD4+ T cells from lupus-prone mice and SLE patients demonstrate enhanced glycolysis and mitochondrial oxidative metabolism, alongside an altered lipid raft glycosphingolipid profile4,5,6. Concurrently, dysregulated mammalian target of rapamycin (mTOR) activation has been observed in CD4+ T cells from SLE patients and lupus-prone mice, leading to metabolic disruptions, affecting glycolysis, fatty acid synthesis, mRNA translation, lipid synthesis, amino acids and growth factors7,8,9. Immunometabolomics provides a novel lens for elucidating SLE mechanisms and may inform targeted therapeutic strategies10. Metabolites are emerging as potential biomarkers for SLE diagnosis, disease activity assessment11,12,13, and organ involvement evaluation14,15. The gut microbiota’s interaction with the host immune system and its association with serum metabolites offer additional insights into SLE’s complex etiology16,17. These findings underscore the significance of metabolic profiling in advancing our understanding of SLE and guide the development of future diagnostic and therapeutic approaches.
Vitamin D is considered an immunomodulatory agent that regulates innate and adaptive immune systems. Vitamin D and vitamin D receptor (VDR) have been shown to modulate the metabolic activity of immune cells, which impact cell activation, differentiation, and cytokine production, ultimately might affecting the immune response in autoimmune diseases18,19. Vitamin D is also believed to play a role in modulating the pathogenesis of SLE. Various studies have shown that vitamin D deficiency is observed in SLE patients20, which may be associated with disease activity20, renal involvement21 and clinical characteristics22. Drug administration may affect vitamin D levels of SLE patients. The therapeutic regimens, including corticosteroids and hydroxychloroquine, may modulate the vitamin D status in SLE patients, albeit with some disagreement among researchers regarding the former’s impact23. Genetic studies suggested that VDR polymorphism may predispose to SLE development24. The active metabolite of vitamin D, 1,25(OH)2D3, has demonstrated immunomodulatory capabilities by dampening the activation of dendritic cells curbing the expression of CD40, MHC class II, and CD86 molecules, and attenuating dendritic cell activation and IFN-α-related gene expression25,26. Additionally, vitamin D is theorized to safeguard podocytes from autoimmune antibody-induced damage in lupus nephritis27. However, vitamin D supplementation in SLE treatment showed conflicting results. Several studies showed that vitamin D supplementation increased serum vitamin D concentration, ameliorate disease activity scores, reduce anti-dsDNA antibodies and proteinuria. It has also been suggested to bolster naive CD4+ T cell and Treg cell populations while diminishing Th1 cells, Th17 cells and memory B cells. In contrast, other studies have reported negligible changes in SLE disease activity or serological markers, including anti-dsDNA antibody and complement protein measurements28. Elucidating the complex interplay of Vitamin D in the immunopathogenesis of SLE presents a significant challenge. Prior research endeavors have predominantly focused on the modulation of immune cell function and cytokine profiles by vitamin D in SLE. Despite these efforts, the link between Vitamin D deficiency, metabolite levels in patients, and their pathogenic contribution to SLE remains to be fully elucidated.
In our study, we performed metabolomics studies in patients with SLE, comparing those with vitamin D deficiency to those without vitamin D deficiency. Our findings shed light on the metabolic differences associated with vitamin D deficiency and emphasize the crucial role of vitamin D in regulating lipid metabolism. These insights contribute to our understanding of the pathological mechanisms underlying SLE, potentially paving the way for novel diagnostic strategies.
Materials and methods
Study population
Between May and June 2022, a total of 31 SLE patients aged from 14 to 69 years (mean 38.35 ± 2.32 years) from our hospital admissions were enrolled in the study. All fulfilled the 1997 American College of Rheumatology classification criteria29. Blood was taken for the assay of 25(OH)D3. Current SLE disease activity was measured using the SLE Disease Activity Index 2000 (SLEDAI-2 K)30. The patients’ characteristics, including age, gender, routine blood test results, kidney and liver biochemistry test results, lipid levels, serum ferritin levels, serum anti-dsDNA antibody, immunoglobin, serum complement, lymphocyte subsets and urinary protein results were recorded. Medical history were registered. All participants were given written consent to the study approved by the Ethics Committee of the Drum Tower Clinical Medical School of Nanjing Medical University.
Assay of 25(OH)D3 and definition of deficiency
Levels of 25(OH)D3 were measured in serum samples, in duplicate, using a commercial ELISA kit according to the manufacturer’s instructions (MULTISCIENCES, China). All blood samples were collected at 6 a.m. Vitamin D deficiency was defined as a 25(OH)D3 level of < 20 ng/ml18.
Sample preparation for metabolomics study
The serum of SLE patients were collected and transferred to − 80 °C for long term storage. 100 µL serum was thoroughly mixed with 400 µL of cold methanol acetonitrile (v/v, 1:1) via vortexing. And then the mixture was processed with sonication for 1 h in ice baths. The mixture was then incubated at − 20 °C for 1 h, and centrifuged at 4 °C for 20 min with a speed of 14,000 g. The supernatants were then harvested and dried under vacuum liquid chromatography–mass spectrometry (LC–MS) analysis31.
LC–MS metabolomics data acquisition
Metabolomics profiling was analyzed using a UPLC-ESI-Q-Orbitrap-MS system (UHPLC, Shimadzu Nexera X2 LC-30AD, Shimadzu, Japan) coupled with Q-Exactive Plus (Thermo Scientific, San Jose, USA). For liquid chromatography (LC) separation, samples were analyzed using a ACQUITY UPLC® HSS T3 column (2.1 × 100 mm, 1.8 μm) (Waters, Milford, MA, USA). The flow rate was 0.3 mL/min and the mobile phase contained: A: 0.1% FA in water and B: 100% acetonitrile (ACN). The gradient was 0% buffer B for 2 min and was linearly increase to 48% in 4 min, and then up to 100% in 4 min and maintained for 2 min, and then decreased to 0% buffer B in 0.1 min, with 3 min re-equilibration period employed. The electrospray ionization (ESI) with positive-mode and negative mode were applied for MS data acquisition separately. The HESI source conditions were set as follows: Spray Voltage: 3.8 kv (positive) and 3.2 kv (negative); Capillary Temperature: 320 °C; Sheath Gas (nitrogen) flow: 30 arb (arbitrary units); Aux Gas flow: 5 arb; Probe Heater Temp: 350 °C; S-Lens RF Level: 50. The instrument was set to acquire over the m/z range 70–1050 Da for full MS. The full MS scans were acquired at a resolution of 70,000 at m/z 200, and 17,500 at m/z 200 for MS/MS scan. The maximum injection time was set to for 100 ms for MS and 50 ms for MS/MS. The isolation window for MS2 was set to 2 m/z and the normalized collision energy (stepped) was set as 20, 30 and 40 for fragmentation31.
LC–MS metabolomics data analysis
The raw MS data were processed using MS-DIAL for peak alignment, retention time correction and peak area extraction. The metabolites were identified by accuracy mass (mass tolerance < 10 ppm) and MS/MS data (mass tolerance < 0.02 Da) which were matched with HMDB, massbank and other public databases and our self-built metabolite standard library. In the extracted-ion features, only the variables having more than 50% of the nonzero measurement values in at least one group were kept.
R (version:4.0.3) and R packages were used for all data analyses and modeling. Data were mean-centered using Pareto scaling. To understand the difference of metabolomics profile between SLE patients with low serum vitamin D level and normal serum vitamin D level, principal component analysis (PCA) was carried out. At the same time, the discriminating metabolites were obtained using Fold change analysis and two-tailed Student’s t test on the normalized raw data at univariate analysis level. Further volcano plot analysis with fold-change (FC) > = 1.5 or FC < = 1.5 and FDR-adjusted P < 0.05 were considered to be statistically significant metabolites. On the other side, the identified differential metabolites were used to perform cluster analyses with R package. The Spearman’s correlation was used to describe the relationships between different metabolites. The ROC analysis was carried out to evaluate each candidate metabolite biomarker. The AUC from ROC analyses was computed using the SPSS software package (version 24, IBM)31.
KEGG enrichment analysis
To identify the perturbed biological pathways, the differential metabolite data were performed KEGG pathway analysis using KEGG database (http://www.kegg.jp). KEGG enrichment analyses were carried out with the Fisher’s exact test, and FDR correction for multiple testing was performed. Enriched KEGG pathways were nominally statistically significant at the P < 0.05 level31.
Statistics
The clinical data of SLE patients were summarized as the mean ± standard error (mean ± SEM). To compare clinical data of two groups, the Student’s t-test was performed. The correlations were determined by Pearson Correlation Coefficients. All statistical analyses were performed using GraphPad Prism 9 software (Graph-Pad, San Diego, CA, USA).
Ethical approval
All participants gave their written Informed consent approved by the Ethics Committee of the Drum Tower Clinical Medical School of Nanjing Medical University in accordance with the Declaration of Helsinki.
Results
Characteristics of SLE patients
In this study, a total of 31 SLE patients were included, and patients were divided into with vitamin D deficiency (VD1, serum vitamin D < 20 ng/ml, n = 21) and without vitamin D deficiency (VD2, serum vitamin D > 20 ng/ml, n = 10) groups, based on their serum vitamin D levels18,32. The demographics and clinical characteristics of SLE patients were shown in Table 1. There were no significant differences in routine blood test results, lymphocytes, kidney and liver biochemistry, lipid levels, serum ferritin levels, anti-dsDNA antibody, immunoglobin, or serum complement between the two groups. However, VD1 SLE patients tended to have higher SLE disease activity index (SLEDAI) scores and proteinuria than VD2 SLE patients. The average dosage of glucocorticoids (calculated according to the equivalent dose of methylprednisolone) of the two groups was 23.45 ± 5.48 mg versus 17.50 ± 4.55 mg. The vast majorities were also treated with HCQ and other immunosuppressants. A total of 57.14% patients with vitamin D deficiency were taking Vitamin D supplements, while 50.00% patients without vitamin D deficiency were taking Vitamin D supplements.
Serum metabolites profiling in SLE patients with or without serum vitamin D deficiency
To uncover the metabolome differences between SLE patients with vitamin D deficiency and without vitamin D deficiency, we performed liquid chromatography–mass spectrometry (LC–MS) analysis and identified a total of 1599 metabolites (Fig. 1A). PCA with QC samples was performed to assess the experiment quality, which showed that the QC samples were closely clustered together in the positive and negative ion mixed mode (Fig. 1B). This indicates that our experiment repeated well and met the required quality for subsequent difference analysis. Using volcano plot with fold change analysis and t-test, we selected 52 significantly altered metabolites based on fold change (FC) > = 1.5 or FC < = − 1.5 and FDR-adjusted p-values < 0.05 (Fig. 1C). Among these, etofylline was up-regulated in VD1 SLE patients, while the remaining 51 metabolites were all downregulated in VD1 SLE patients, predominantly comprising lipids (Fig. 1D). The differential metabolites screened in metabolomics mainly included lipid and lipid-like molecules, benzenoids, organic acids and derivatives, organic oxygen compounds and organoheterocyclic compounds. Lipid and lipid-like molecules accounted for a major proportion (66.67%) of metabolites identified (Fig. 1E). Specifically, the differential lipid and lipid-like molecules mainly included fatty acyls (41.18%), prenol lipids (29.41%), steroids and steroid derivatives (14.71%), glycerophospholipids (8.82%) and sphingolipids (5.88%) (Fig. 1F). These findings suggest that lipid metabolism is significantly altered in SLE patients with vitamin D deficiency.

Serum metabolites profiling in SLE patients with or without serum vitamin D deficiency. (A) Classified bubble plot. The horizontal coordinate represents the number interval of differential metabolites, and the vertical coordinate represents the logarithmic transformation of FC. Each circle represents a differential metabolite, and the larger the circle, the higher the difference significance. Different colors indicate a primary class, where the differentiated metabolites of the same class are arranged together. (B) PCA score plots and heatmap of metabolomics. Overview of PCA score plots from all VD1 (red), all VD2 (green), and QC (blue) in positive and negative mixed mode. (C) Volcano Plot. Metabolites with FC > = 1.5 or FC < = − 1.5 and FDR-adjusted P < 0.05 were considered as statistically significant metabolites. The vertical dashed line analysis indicated log2(1/1.5) and log2(1.5). The red dots represent up-regulated metabolites, and the blue dots represent down-regulated metabolites. (D) Complex heatmap for significantly differential metabolites. The abundance of each metabolite is normalized by Z score normalization. The color is positively correlated with the intensity of change in metabolites, with red indicating up-regulation and green indicating down-regulation. Metabolite ontology, P value and FC value are shown in the left of the heatmap. (E). Classified pie chart of HMDB super class. The numbers and proportions of differential metabolites are shown. (F) Classified pie chart of HMDB class. The numbers and proportions of differential metabolites are shown.
Correlations between differential metabolites
We analyzed the linear correlations between these significant differential metabolites and screened out significant correlations with absolute correlation coefficient > 0.9(|r|> 0.9). There were 19 pairs of strong correlations between these differential metabolites in total. Interestingly, 12 of these pairs involved correlations between lipids and lipids, especially fatty acyls, while 5 pairs of them were between lipids and 2-Hydroxy-4,5′,8a′-trimethyl-1′-oxo-4-vinyloctahydro-1′H-spiro[cyclopentane-1,2′-naphthalene]-5′-carboxylic acid (Fig. 2A and B). Moreover, they were all positive associated. Their strong positive association suggested that these metabolites may participate in the same metabolic pathway, or biological process.

Correlations between differential metabolites. (A) Correlation coefficient matrix heatmap. Positive correlation is shown in red, while negative correlation is shown in blue. The larger the proportion of coloring intervals, the stronger the correlation. (B) Circos plot. From outside to inside, there are metabolite names, HMDB classification of metabolites, log2(fold change), P-value, and correlation lines.
KEGG pathways of SLE patients with or without serum vitamin D deficiency
A total of 52 differential metabolites were submitted to the KEGG, and they were matched to 27 metabolic pathways. Among these pathways, lipid metabolism and digestive system pathway were significant secondary classific pathways (Fig. 3A). In terms of the primary classific pathways, fat digestion and absorption (P = 4.75E − 05), pathways in cancer(P = 2.82E − 04) and vitamin digestion and absorption (P = 4.46E − 04) were significant (Fig. 3B). Further pathway impact analyses revealed that sphingolipid metabolism, glycerolipid metabolism, glycerophospholipid metabolism and primary bile acid biosynthesis were all significant pathways and they all belong to lipid metabolism pathway (Fig. 3C, D), indicating the important role of lipid metabolism in SLE patients with vitamin D deficiency.

Metabolic pathway analysis of differential metabolites using MetaboAnalyst 4.0 based on the KEGG. (A) Secondary classific significant pathway bubble plot. (B) Significant pathway bubble plot. The significance of pathways in each class decreases from top to bottom. Size of circle represents the number of differential metabolites annotated into the pathway. Color of circle represents the corrected P value. (M, Metabolism; G, Genetic Information Processing; E, Environmental Information Processing; C, Cellular Processes; O, Organismal Systems; H, Human Diseases; D, Drug Development.) (C) Pathway impact plot calculated by Betweenness centrality method. (D) Pathway impact plot calculated by Out degree centrality method.
Regulatory network analysis of differential metabolites and pathways
We searched the screened differential metabolites in the KEGG database for their corresponding pathways and performed regulatory network analysis. The most important differential metabolites were cholesterol (C00187), Lysophosphatidic acid (18:2) (C00681) and spectral match to Lyso-Sphingomyelin from NIST14 (C00550). The mainly associated pathways were vitamin digestion and absorption (hsa04977), neuroactive ligand-receptor interaction (hsa04080), basal cell carcinoma (hsa05217), fat digestion and absorption (hsa04975), regulation of lipolysis in adipocytes (hsa04923), sphingolipid metabolism (hsa00600) and glycerophospholipid metabolism (hsa00564) (Fig. 4). These results suggest that vitamin D-altered metabolites may involve in lipid digestion, absorption and metabolism, as well as cellular interaction in SLE patients with different vitamin D levels.

Regulatory network analysis of differential metabolites. Red circle represents metabolic pathway. Purple circle represents modular information. Yellow circle represents enzyme. Green circle represents background compound of a metabolic pathway. Blue circle represents chemical reaction. Green square represents differential metabolite.
Diagnostic potential of differential metabolites
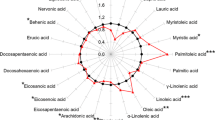
To explore the diagnostic potential of the differential metabolites in predicting vitamin D levels and diagnosing SLE, we further compared relative abundance of these differential metabolites. The result showed that totally 9 metabolites were significantly decreased in VD1 SLE patients, while Etofylline was significantly increased in VD1 SLE patients (Fig. 5A). Then we performed classical univariate receiver operating characteristic (ROC) curve analyses to generate ROC curve, to calculate area under the curve (AUC) and their 95% confidence intervals. The ROC analysis showed that the AUC of (S)-Oleuropeic acid was 0.867 (0.710, 0.993) and 2-Hydroxylinolenic acid was 0.833 (0.669, 0.948) (Fig. 5B). The correlations between (S)-Oleuropeic acid and 2-Hydroxylinolenic acid, and clinical parameters were analyzed. It was found that the levels of (S)-Oleuropeic acid were positively correlated with vitamin D, IgA and IgG levels in SLE patients (Table 2). The levels of 2-Hydroxylinolenic acid were positively correlated with vitamin D, ESR, IgG and IgE levels in SLE patients (Table 3). These results indicated that (S)-Oleuropeic acid and 2-Hydroxylinolenic acid might have promising diagnostic potential as biomarkers for predicting vitamin D levels and the disease activity of SLE patients.

Diagnostic potential of differential metabolites. (A) Relative presentations of differential metabolites in VD1 SLE patients and VD2 SLE patients. (B) ROC curves of the important altered metabolites.
Discussion
In recent years, metabolomics has emerged as a valuable tool for studying the underlying pathological mechanisms of SLE. Lipid metabolism, including anabolism and catabolism, is essential for almost every aspect of cellular functioning33. Vitamin D, on the other hand, has been shown to be involved in autoimmune diseases18. It is well-established that SLE patients often have insufficiency of vitamin D levels and lower serum levels of vitamin D were associated with proteinuria24,25,34,35. Our study investigated the serum metabolome differences between SLE patients with vitamin D deficiency and those without vitamin D deficiency using LC–MS analysis. We identified a total of 52 differentially distributed metabolites, with 34 of them being lipid and lipid-like molecules. Interestingly, all these lipids were downregulated in SLE patients with vitamin D deficiency. These results indicated a strong association between vitamin D and lipid metabolism of SLE patients.
Among the altered metabolites, 14 differential metabolites were fatty acyls, which can promote angiogenesis and reduce inflammation. A total of 10 differential metabolites were prenol lipids, which can regulate the progression of aging-related diseases, diabetes and inflammation and as regulator of bone health and cardiovascular homeostasis. A total of 5 differential metabolites were sterol lipids, which is the component of membrane lipids and can regulate T cell function as hormones and signaling molecules. A total of 3 differential metabolites were glycerophospholipids, which is the structural component of membrane and can affect membrane fluidity. The other 2 differential metabolites were sphingolipids, which is an important component of the cell membrane and can induce lipotoxicity and inflammation and regulate cell death36. Current evidence suggested that lipids and lipid metabolites played an important role in the pathogenesis of SLE. Lipid metabolism was considered as an important facilitator of T cell differentiation. Lipid biosynthesis, including cholesterol and fatty acids, was critical for the proliferation and differentiation of T cells, especially Th17 cells37. Inhibiting FA synthesis in memory CD4+T cells of SLE patients decreased interferon (IFN)-γ production and increased Foxp3 expression in T-bet+Foxp3+ cells. Fatty acid synthesis inhibitors may improve the pathological status by correcting Th1 subset imbalance and overproduction of IFN-γ in SLE38. Fatty acids could modulate the process of macrophage differentiation39. Targeting receptors associated lipid metabolism, such as peroxisome proliferator-activated receptor γ (PPARγ), 12/15-Lipoxygenase (12/15-LO), sphingosine-1-phosphate (S1P) and liver X receptors (LXRs), was able to affect differentiation of monocytes and production of proinflammatory cytokines, which were related to TLR7 and TLR9 expression40,41,42,43. Furthermore, we observed strong associations between the differential metabolites, particularly the lipids, suggesting that these metabolites may participate in the same metabolic pathway, or biological process. We speculated that the downregulated lipids in vitamin D deficient SLE patients may affect the proliferation, differentiation and production of proinflammatory cytokines of lymphocytes and innate immune cells and participated in the pathogenesis of SLE.
The fatty acid (FA) and cholesterol metabolic pathways are the most important pathways in lipid metabolism. Lipid synthesis is critical for the cell and determine cell survival44. Lipid degradation is also essential to sustain life. Lipolytic products and intermediates play a vital role in cellular signaling45. Our results showed that fat digestion and absorption, sphingolipid metabolism, glycerolipid metabolism, glycerophospholipid metabolism and primary bile acid biosynthesis pathways played a crucial part in regulating differential metabolites of vitamin D deficient SLE patients. These data shed light on the probable signal pathways by which these altered metabolites participated in the development of SLE.
Recent researches have shed light on the potential mechanisms by which vitamin D participates in the pathogenesis of SLE. VDR generally skews immune cells towards anti-inflammatory states46. Vitamin D has been shown to decrease neutrophil extracellular traps (NETs) and prevent endothelial damage in cultured neutrophiles derived from SLE patients, compared to controls, revealing the possibility of Vitamin D as supplementary therapy for SLE patients with hypovitamin D to prevent endothelial damage47. Liu et al.48 revealed that 1,25-(OH)2D3/VDR facilitated the recovery of SLE by downregulating Skp2 and upregulating p27 expression. New scientific approaches targeting VitD/VDR signaling at the cellular metabolic level may provide a better comprehension of its role in SLE progression. However, there is no related report till now. Our study indicated the metabolic interactions between VitD and SLE, providing new insights into the mechanism of SLE.
The clinical manifestations of SLE patients are highly heterogeneous, involving various symptoms in the gastrointestinal tract, skin, kidney, blood, nerves system, and other organs. Given the multidimensional nature of organ involvement and fluctuating disease activity, accurate assessment tools are essential for clinical decision-making. The SLEDAI30, BILAG index49, SSLICC/ACR DI50 are established metrics for this purpose. Emerging research leverages soluble mediators, such as cytokines, chemokines, and soluble receptors, to gauge clinical activity and forecast disease flares in SLE51. However, current tools are still not available to fully measure disease activity of SLE patients with different subtypes. Given the different clinical manifestations of SLE and the pathological differences between active and inactive disease, metabolomics may be an important platform for discovering different clinical phenotypic biomarkers for SLE15. Notably, SLE patients with vitamin D deficiency could represent an important subgroup. The metabolomic profiling in our study have the potential to unravel unique metabolic signatures associated with different clinical phenotypes in SLE. These metabolites could serve as specific biomarkers to better stratify patients, identify disease activity, and predict outcomes. Furthermore, the discovery of differential metabolites in SLE patients with vitamin D deficiency may shed light on the underlying mechanisms linking vitamin D status and disease pathogenesis in SLE.
The current study has several limitations. Firstly, the validation of metabolic biomarkers in SLE with vitamin D deficiency requires a larger patient cohort. Secondly, a more nuanced comparison beyond deficiency is needed, considering gradations of deficiency, insufficiency, and sufficiency. Lastly, the specific roles and mechanisms of action of individual metabolites in the pathophysiology of SLE merit deeper exploration. Addressing these limitations will enhance our understanding and management of SLE.
In conclusion, our study revealed the global metabolic differences in SLE patients with vitamin D deficiency, illustrating the extensive association between vitamin D and lipid metabolism in these patients. Further investigations are needed to further explore the underlying mechanism by which vitamin D regulates the lipid metabolism and affects immune cells. This study provides a rational basis for a better understanding of the pathogenetic mechanism underlying SLE and may pave the way for the development of targeted therapeutic approaches.
Data availability
The datasets generated during and/or analyzed during the current study are not publicly available out of concern for patient privacy, as data from medical records is considered sensitive, but de-identified datasets are available from the corresponding author on reasonable request.
References
Bernatsky, S. et al. Mortality in systemic lupus erythematosus. Arthritis Rheum. 54(8), 2550–2557. https://doi.org/10.1002/art.21955 (2006).
Ameer, M. A. et al. An overview of systemic lupus erythematosus (SLE) pathogenesis, classification, and management. Cureus 14(10), e30330. https://doi.org/10.7759/cureus.30330 (2022).
Wu, Y. et al. Immunometabolomics provides a new perspective for studying systemic lupus erythematosus. Int. Immunopharmacol. 118, 109946. https://doi.org/10.1016/j.intimp.2023.109946 (2023).
Yin, Y. et al. Normalization of CD4+ T cell metabolism reverses lupus. Sci. Transl. Med. 7(274), 274ra18. https://doi.org/10.1126/scitranslmed.aaa0835 (2015).
McDonald, G. et al. Normalizing glycosphingolipids restores function in CD4+ T cells from lupus patients. J. Clin. Invest. 124(2), 712–724. https://doi.org/10.1172/JCI69571 (2014).
Zhu, Y. et al. Lowering glycosphingolipid levels in CD4+ T cells attenuates T cell receptor signaling, cytokine production, and differentiation to the Th17 lineage. J. Biol. Chem. 286(17), 14787–14794. https://doi.org/10.1074/jbc.M111.218610 (2011).
Murray, P. J., Rathmell, J. & Pearce, E. SnapShot: Immunometabolism. Cell Metab. 22(1), 190-190 el. https://doi.org/10.1016/j.cmet.2015.06.014 (2015).
Zhang, C. X. et al. Immunometabolism in the pathogenesis of systemic lupus erythematosus. J. Transl. Autoimmun. 3, 100046. https://doi.org/10.1016/j.jtauto.2020.100046 (2020).
Robinson, G. A., Wilkinson, M. G. L. & Wincup, C. The role of immunometabolism in the pathogenesis of systemic lupus erythematosus. Front. Immunol. 12, 806560. https://doi.org/10.3389/fimmu.2021.806560 (2022).
Hu, J. Q. et al. Targeting abnormal lipid metabolism of T cells for systemic lupus erythematosus treatment. Biomed. Pharmacother. 165, 115198. https://doi.org/10.1016/j.biopha.2023.115198 (2023).
Zhang, Q. et al. Metabolomic profiling reveals serum L-pyroglutamic acid as a potential diagnostic biomarker for systemic lupus erythematosus. Rheumatology (Oxford) 60(2), 598–606. https://doi.org/10.1093/rheumatology/keaa126 (2021).
Li, Y. et al. Lipidomic and metabolomic profiling reveals novel candidate biomarkers in active systemic lupus erythematosus. Int. J. Clin. Exp. Pathol. 12(3), 857–866 (2019).
Yan, R. et al. Fecal metabolites were altered, identified as biomarkers and correlated with disease activity in patients with systemic lupus erythematosus in a GC-MS-based metabolomics study. Front. Immunol. 11, 2138. https://doi.org/10.3389/fimmu.2020.02138 (2020).
Zhang, W. et al. Integration of metabolomics and lipidomics reveals serum biomarkers for systemic lupus erythematosus with different organs involvement. Clin. Immunol. 241, 109057. https://doi.org/10.1016/j.clim.2022.109057 (2022).
Kalantari, S. et al. Metabolomics approach reveals urine biomarkers and pathways associated with the pathogenesis of lupus nephritis. Iran. J. Basic Med. Sci. 22(11), 1288–1295. https://doi.org/10.22038/ijbms.2019.38713.9178 (2019).
Luo, X. M. et al. Gut microbiota in human systemic lupus erythematosus and a mouse model of lupus. Appl. Environ. Microbiol. 84(4), e02288-e2317. https://doi.org/10.1128/AEM.02288-17 (2018).
Yang, W. & Cong, Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol. Immunol. 18(4), 866–877. https://doi.org/10.1038/s41423-021-00661-4 (2021).
Harrison, S. R. et al. Vitamin D, autoimmune disease and rheumatoid arthritis. Calcif. Tissue Int. 106(1), 58–75. https://doi.org/10.1007/s00223-019-00577-2 (2020).
Bishop, E. L. et al. Vitamin D and immune regulation: Antibacterial, antiviral, anti-inflammatory. JBMR Plus 5(1), e10405. https://doi.org/10.1002/jbm4.10405 (2021).
Sahebari, M., Nabavi, N. & Salehi, M. Correlation between serum 25(OH)D values and lupus disease activity: An original article and a systematic review with meta-analysis focusing on serum VitD confounders. Lupus 23(11), 1164–1177. https://doi.org/10.1177/0961203314540966 (2014).
Wang, X. R. et al. Decreased serum/plasma vitamin D levels in SLE patients: A meta-analysis. Curr. Pharm. Des. 24(37), 4466–4473. https://doi.org/10.2174/1381612825666190111145848 (2018).
Mok, C. C. et al. Vitamin D levels in Chinese patients with systemic lupus erythematosus: relationship with disease activity, vascular risk factors and atherosclerosis. Rheumatology (Oxford) 51(4), 644–652. https://doi.org/10.1093/rheumatology/ker212 (2012).
Athanassiou, L., Kostoglou-Athanassiou, I., Koutsilieris, M. & Shoenfeld, Y. Vitamin D and autoimmune rheumatic diseases. Biomolecules 13(4), 709. https://doi.org/10.3390/biom13040709 (2023).
Islam, M. A. et al. Vitamin D status in patients with systemic lupus erythematosus (SLE): A systematic review and meta-analysis. Autoimmun. Rev. 18(11), 102392. https://doi.org/10.1016/j.autrev.2019.102392 (2019).
Ben-Zvi, I. et al. The impact of vitamin D on dendritic cell function in patients with systemic lupus erythematosus. PLoS One 5(2), e9193. https://doi.org/10.1371/journal.pone.0009193 (2010).
Lerman, M., Burnham, J. & Behrens, E. 1,25 dihydroxyvitamin D3 limits monocyte maturation in lupus sera. Lupus 20(7), 749–753. https://doi.org/10.1177/0961203310394542 (2011).
Yu, Q. et al. Vitamin D protects podocytes from autoantibodies induced injury in lupus nephritis by reducing aberrant autophagy. Arthritis Res. Ther. 21(1), 19. https://doi.org/10.1186/s13075-018-1803-9 (2019).
Cutolo, M., Smith, V., Paolino, S. & Gotelli, E. Involvement of the secosteroid vitamin D in autoimmune rheumatic diseases and COVID-19. Nat. Rev. Rheumatol. 19(5), 265–287. https://doi.org/10.1038/s41584-023-00944-2 (2023).
Hochberg, M. C. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheum. 40(9), 1725. https://doi.org/10.1002/art.1780400928 (1997).
Gladman, D. D., Ibanez, D. & Urowitz, M. B. Systemic lupus erythematosus disease activity index 2000. J. Rheumatol. 29(2), 288–291 (2002).
Menezes, E. B. et al. Uncovering sperm metabolome to discover biomarkers for bull fertility. BMC Genom. 20(1), 714. https://doi.org/10.1186/s12864-019-6074-6 (2019).
Kowalówka, M. et al. Clinical significance of analysis of vitamin D status in various diseases. Nutrients 12(9), 2788. https://doi.org/10.3390/nu12092788 (2020).
Sun, W. et al. Lipid metabolism: Immune regulation and therapeutic prospectives in systemic lupus erythematosus. Front. Immunol. 13, 860586. https://doi.org/10.3389/fimmu.2022.860586 (2022).
Kowalowka, M. et al. Clinical significance of analysis of vitamin D status in various diseases. Nutrients 12(9), 2788. https://doi.org/10.3390/nu12092788 (2020).
Mu, Q., Zhang, H. & Luo, X. M. SLE: Another autoimmune disorder influenced by microbes and diet?. Front. Immunol. 6, 608. https://doi.org/10.3389/fimmu.2015.00608 (2015).
Zhang, C. et al. Lipid metabolism in inflammation-related diseases. Analyst 143(19), 4526–4536. https://doi.org/10.1039/c8an01046c (2018).
Hu, X. et al. Sterol metabolism controls T(H)17 differentiation by generating endogenous RORgamma agonists. Nat. Chem. Biol. 11(2), 141–147. https://doi.org/10.1038/nchembio.1714 (2015).
Iwata, S. et al. Enhanced fatty acid synthesis leads to subset imbalance and IFN-gamma overproduction in T helper 1 cells. Front. Immunol. 11, 593103. https://doi.org/10.3389/fimmu.2020.593103 (2020).
Verescakova, H. et al. Nitro-oleic acid regulates growth factor-induced differentiation of bone marrow-derived macrophages. Free Radic. Biol. Med. 104, 10–19. https://doi.org/10.1016/j.freeradbiomed.2017.01.003 (2017).
Mohammadi, S., Saghaeian-Jazi, M., Sedighi, S. & Memarian, A. Immunomodulation in systemic lupus erythematosus: induction of M2 population in monocyte-derived macrophages by pioglitazone. Lupus 26(12), 1318–1327. https://doi.org/10.1177/0961203317701842 (2017).
Uderhardt, S. et al. 12/15-lipoxygenase orchestrates the clearance of apoptotic cells and maintains immunologic tolerance. Immunity 36(5), 834–846. https://doi.org/10.1016/j.immuni.2012.03.010 (2012).
Cartier, A. & Hla, T. Sphingosine 1-phosphate: Lipid signaling in pathology and therapy. Science 366(6463), eaar5551. https://doi.org/10.1126/science.aar5551 (2019).
Kim, H. A. et al. The liver X receptor is upregulated in monocyte-derived macrophages and modulates inflammatory cytokines based on LXRalpha polymorphism. Mediat. Inflamm. 2019, 6217548. https://doi.org/10.1155/2019/6217548 (2019).
Howie, D. et al. The role of lipid metabolism in T lymphocyte differentiation and survival. Front. Immunol. 8, 1949. https://doi.org/10.3389/fimmu.2017.01949 (2017).
Zechner, R. et al. FAT SIGNALS–Lipases and lipolysis in lipid metabolism and signaling. Cell Metab. 15(3), 279–291. https://doi.org/10.1016/j.cmet.2011.12.018 (2012).
Yang, C. Y. et al. The implication of vitamin D and autoimmunity: A comprehensive review. Clin. Rev. Allergy Immunol. 45(2), 217–226. https://doi.org/10.1007/s12016-013-8361-3 (2013).
Handono, K. et al. Vitamin D prevents endothelial damage induced by increased neutrophil extracellular traps formation in patients with systemic lupus erythematosus. Acta Med. Indones. 46(3), 189–198 (2014).
Liu, D. et al. 1,25-(OH)(2)D(3)/Vitamin D receptor alleviates systemic lupus erythematosus by downregulating Skp2 and upregulating p27. Cell Commun. Signal 17(1), 163. https://doi.org/10.1186/s12964-019-0488-2 (2019).
Isenberg, D. A. et al. BILAG 2004. Development and initial validation of an updated version of the British Isles Lupus Assessment Group’s disease activity index for patients with systemic lupus erythematosus. Rheumatology (Oxford) 44(7), 902–906. https://doi.org/10.1093/rheumatology/keh624 (2005).
Petri, M. et al. Derivation and validation of the Systemic Lupus International Collaborating Clinics classification criteria for systemic lupus erythematosus. Arthritis Rheum. 64(8), 2677–2686. https://doi.org/10.1002/art.34473 (2012).
Thanou, A. et al. Clinical disease activity and flare in SLE: Current concepts and novel biomarkers. J. Autoimmun. 119, 102615. https://doi.org/10.1016/j.jaut.2021.102615 (2021).
Acknowledgements
The authors acknowledge Hong Yu for assistance in analysing the data.
Funding
The work was supported by Nanjing Medical Science and technique Development Foundation (JQX20004), and funding for Clinical Trials from the Affiliated Drum Tower Hospital, Medical School of Nanjing University (2021-LCYJ-PY-16).
Author information
Authors and Affiliations
Contributions
L.G., L.S. designed the study. Y.Y. collected the data, performed the experiments and performed the statistical analysis. Y.Y., F.Y. and Q.L. analysed the data. Y.Y., L.G. and X.F. wrote the manuscript. All authors approved the final manuscript as submitted and agree to be accountable for all aspects of the work.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yan, Y., Yu, F., Li, Q. et al. Metabolic alterations in vitamin D deficient systemic lupus erythematosus patients. Sci Rep 14, 18879 (2024). https://doi.org/10.1038/s41598-024-67588-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-67588-4
- Springer Nature Limited