
Abstract
Brucellosis is a zoonotic disease caused by bacteria of the Brucella genus. This study investigates the development of a multi-epitope mRNA vaccine aimed at combating Brucella infections. The approach involves selecting the architecture of the pathogenic type IV secretion system (T4SS) of Brucella using bioinformatics tools and reverse vaccinology methodologies. The candidate proteins chosen for this vaccine include VirB2, VirB3, and VirB6, along with the effector protein BtpA. Key findings include robust antigenicity scores for the candidate proteins (VirB2:0.5685; VirB3:0.5329;VirB6:0.5054;BtpA:0.5575).A variety of tools were essential for identifying potential epitopes suitable for incorporation into the mRNA vaccine, as well as for utilizing appropriate linkers for docking. Molecular dynamics simulations confirmed stable interactions between the vaccine and TLR4, with a docking score of − 350.15. Furthermore, computer cloning methods were employed alongside simulated gel electrophoresis to ensure a comprehensive evaluation of the vaccine’s properties. The results of this investigation indicate that the newly formulated mRNA vaccine effectively stimulates an immune response, presenting innovative strategies for prevention and management. Although comprehensive evaluations of vaccine design have been conducted using bioinformatics tools and molecular simulations, certain limitations still exist, primarily manifested in the reliance on computational predictions without validation from wet lab experiments. Future research must further verify the accuracy of computational results through laboratory experiments.
Similar content being viewed by others


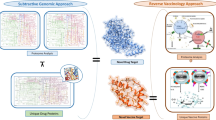
Introduction
Brucella, which poses significant health risks to both humans and animals1. Brucellosis represents a significant public health challenge globally, particularly for individuals residing in resource-limited environments2. In regions such as Central and South America, Central Asia, the Mediterranean, and the Middle East, the disease remains challenging to eradicate and control3. In Asian countries, the brucellosis epidemic is particularly severe in pastoral regions, notably in Xinjiang, China. Epidemiological research on Brucella in Xinjiang indicates that the prevention and control of brucellosis in this area should primarily focus on Brucella melitensis4,5. Brucella primarily infects the host by adhering to mucous membranes and penetrating the tissue6. In most patients with brucellosis, clinical symptoms commonly include fever, fatigue, hyperhidrosis, joint pain, and hepatosplenomegaly7. However, the effectiveness of clinical treatment is often limited, and issues with drug resistance are prevalent8. At present, there is no internationally recognized Brucella vaccine available for human use9. This presents a significant challenge to disease control and prevention, highlighting the urgent need to develop a safe and effective vaccine.
The pathogenesis of Brucella is dependent on the involvement of the Type IV secretion system (T4SS) encoded by the VirB gene, which expresses various virulence factors that facilitate the evasion of host immunity10,11. We selected VirB2, VirB3, and VirB6 from the structural proteins of the type IV secretion system (T4SS), in addition to the effector protein BtpA, as candidate proteins for the mRNA vaccine12. The effector protein BtpA, associated with T4SS, possesses a TIR domain that is structurally similar to Toll-like receptors (TLRs)13. This domain has the ability to inhibit nicotinamide adenine dinucleotide (NAD) in host cells, thereby regulating cellular metabolism and signal transduction12.
Candidate proteins for Brucella vaccines must demonstrate strong antigenicity, as well as non-toxicity and non-allergenicity14. Subsequently, utilize bioinformatics tools such as IEDB, NetMHCpan-4.1, NetMHCIIpan-4.0, ABCpred, and Elipro to perform a comprehensive analysis of the epitopes15. To ensure effective immune responses, we utilize suitable adapters to connect the multiple epitopes we have analyzed, integrating them into a single polypeptide16. The construction of mRNA vaccines necessitates a thorough analysis of the physical and chemical properties of the polypeptides utilized, alongside their capacity to elicit an immune response17. This study aims to design an mRNA vaccine and to provide new insights for future research directions in the prevention of brucellosis. Key obstacles encountered in clinical translation include the optimization of delivery systems, vaccine stability, and the durability of immune responses18. While mRNA vaccines offer advantages in design and production, challenges remain regarding their in vivo delivery efficiency, potential immunogenicity, and long-term stability, all of which necessitate further research19.
Materials and methods
Acquisition of candidate proteins
We accessed the UniProt database (https://www.uniprot.org/) to gather the protein sequences of several crucial components involved in the Type IV secretion system (T4SS). Specifically, we retrieved the sequences for VirB2, which is identified by the UniProt ID P0C528; VirB3, with the UniProt ID Q9RPY2; VirB6, denoted by the UniProt ID Q8YDZ2; and BtpA, recognized by its UniProt ID Q8YF53. This database provided us with the necessary information to facilitate our research on these important proteins.
Research analytical methods
Protein immunological properties
The protein of interest must exhibit strong antigenicity. To assess this, we utilize VaxiJen v2.0 (https://www.ddg-pharmfac.net/vaxijen/VaxiJen/VaxiJen.htmlthe first software capable of predicting whether a protein is a protective antigen without the need for alignment. In antigenicity prediction, the threshold for VaxiJen v2.0 is established at 0.4, which serves as the standard threshold recommended by the software for differentiating between antigenic and non-antigenic proteins21,22. Using the software ProtParam (https://web.expasy.org/protparam/, researchers can calculate various biological properties of proteins, including molecular weight, theoretical isoelectric point (pI), and total mean hydrophobicity (GRAVY)23. Additionally, as a vaccine candidate protein, it is essential for the protein to be non-allergenic to prevent allergic reactions. The software AlgPred 2.0 (https://webs.iiitd.edu.in/raghava/algpred2/) can effectively classify proteins into allergens and non-allergens24. Finally, ToxinPred2 (https://webs.iiitd.edu.in/raghava/toxinpred2/index.html) can be utilized to predict the toxicity of the protein of interest25.
Prediction of protein signal peptides
SignalP 6.0 (https://services.healthtech.dtu.dk/services/SignalP-6.0/) is capable of predicting five types of signal peptides, thereby facilitating the subsequent screening of dominant epitopes and more accurately identifying the sites where signal peptides are located26.
The physical and chemical properties of VirB2, VirB3, VirB6, and BtpA were analyzed using bioinformatics tools, including UniProt, VaxiJen v2.0, ProtParam, AlgPred 2.0, and ToxinPred227.
Prediction of T-cell epitopes
T-cell receptor and antigen-presenting cell recognition is a critical step in the T-cell response, and this process follows MHC restriction28. CD8 + T-cells, also known as cytotoxic T-cells, are limited by MHCI molecules, whereas CD4 + T-cells, or helper T-cells, are confined by MHCII molecules. Furthermore, the T-cell epitopes play a crucial role in promoting targeted T-cell immune responses29. In this study, we screened helper T-cell epitopes (CTLs) using the high-frequency alleles HLA-A*1101,HLA-A*0201,HLA-A*0301,HLA-DRB1*0701,HLA-DRB1*1501,and HLA-DRB1*0301 from Xinjiang30.
For cytotoxic T lymphocytes (CTLs), we utilize the Next-generation IEDB tools (https://nextgen-tools.iedb.org/) and NetMHCpan-4.1 (https://services.healthtech.dtu.dk/services/NetMHCpan-4.1/) with the peptide length set to 930. For MHC-II binding predictions, we employ the IEDB MHC-II Binding Predictions tool (http://tools.iedb.org/mhcii/) and NetMHCIIpan-4.0 (https://services.healthtech.dtu.dk/services/NetMHCIIpan-4.0/),with the peptide length set to 15; the remaining settings did not require modification14. Subsequently, the prediction results from the two software applications were compared, and the top ten dominant epitopes with the highest scores were selected for further analysis.
Prediction of linear epitopes in B-cells
ABCpred (https://webs.iiitd.edu.in/raghava/abcpred/index.html) was utilized to predict B-cell linear epitopes31. The software is an artificial neural network-based B-cell epitope prediction server. The selection window lengths are set at 10,12,14,16,18,and 20,with a default length of 16. When the threshold is set to 0.5,the server demonstrates an accuracy of 65.93%,exhibiting identical sensitivity and specificity when utilizing a window length of 1632. The top 10 dominant epitopes with the highest scores were employed for biological trait analysis.
Prediction of B-cell conformational epitopes
Utilizing Elipro (http://tools.iedb.org/ellipro/) allows for the prediction and visualization of antibody epitopes within a given protein sequence or structure of interest33. It is important to note that, in addition to the score, the amino acid sequence between the two residues must be fully specified when selecting the conformational epitope.
Docking of T-cell epitopes and HLA alleles
Molecular docking was conducted using the HDOCK server (http://hdock.phys.hust.edu.cn/) to facilitate the interaction between HLA-A*02:01 (PDB ID: 4u6y) and cytotoxic T lymphocytes (CTLs), as well as to connect HLA-DRB1*01:01 (PDB ID: 5v4n) with helper T lymphocytes (HTLs)14. In the docking assessment, the parameters provided by the HDOCK server—such as scoring, confidence, and RMSD—serve as standard metrics for evaluating intermolecular interactions34,35. The reasonable ranges of these parameters are supported by extensive literature, effectively reflecting the binding stability between molecules.
Screening epitope Follow-up assessment
All selected dominant epitopes must be adequately antigenic, non-allergenic, and non-toxic15. Therefore, VaxiJen v2.0 is employed to predict antigenicity, AlgPred 2.0 is utilized to confirm that the selected epitope is non-sensitizing, and ToxinPred2 is used to evaluate the toxicity of all peptides.
mRNA vaccine construction design
Connection methods
In the first step, the final epitopes will be screened and linked using appropriate linkers. This study have selected widely used flexible and rigid linkers, as the choice of an appropriate linker significantly enhances protein function and intermolecular interactions36.We employ AAY to connect all CTLs, GPGPG for all HTLs, KK for all B-cell epitopes, and GGGS to link the His tag (HHHHHHH) at the C-terminus of the protein sequence27. EAAAK can further enhance the immunogenicity of vaccines37.The AAY linker can enhance the immunogenicity of an epitope vaccine38. The GPGPG linker facilitates the immune processing and presentation processes39. The KK linker maintains the independent immune activity of B-cell epitopes in vaccines14. In the second step, the online prediction software EMBOSS Backtranseq (https://emboss.bioinformatics.nl/cgi-bin/emboss/backtranseq) is utilized to convert the ligated protein sequence into a nucleotide sequence. Subsequently, NovoPro (https://www.novopro.cn/tools/codon-optimization.html) is used to perform codon optimization while excising the restriction enzyme sites BamHI and XhoI30,40.
To construct an mRNA vaccine, the 5’cap structure serves to prevent degradation and facilitates correct translation. Additionally, the inclusion of the 5’and 3’untranslated regions (UTRs) enhances the efficiency of translation. A Kozak sequence, which contains a start codon, along with a tPA leading sequence (UniProt ID: P00750),is also introduced. Finally, optimized mRNA sequences are incorporated, culminating in the addition of a poly(A) tail structure at the 3’ end to improve RNA stability41.
The constructed mRNA vaccine was evaluated for antigenicity using VaxiJen v2.0, sensitization with AlgPred 2.0,and toxicity assessment through ToxinPred2. The software ProtParam was employed to calculate the biological characteristics of the vaccine protein sequence, including molecular weight, theoretical isoelectric point (pI), and total mean hydrophobicity (GRAVY), among others. Solubility predictions were made using SOLpro(https://scratch.proteomics.ics.uci.edu/index.html)42.
Prediction of secondary structure of mRNA vaccines
The secondary structure of mRNA vaccines can be predicted using the Vienna RNA webserver(http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi)43.
In Silico cloning of mRNA vaccines
For the in silico cloning of mRNA vaccines, we employed the software SnapGene (https://www.snapgene.com/) to design primers for in silico PCR23. Subsequently, pVAX1 was selected as the vector for in silico cloning, and the designed gene of interest was inserted between the restriction enzyme sites BamHI and XhoI44. In this study, we present an experimental simulation using SnapGene to analyze the results of an agarose gel electrophoresis (1.0% agarose). The constructed mRNA vaccine is loaded in the first lane, followed by plasmid pVAX1 in the second lane, and the cloned plasmid in the third lane. This setup allows for a comparative assessment of the molecular weights and integrity of the different nucleic acid constructs.
Immune simulation prediction
We utilized C-ImmSim (http://150.146.2.1/C-IMMSIM/index.PHP) to evaluate the immune response elicited by the constructed vaccine. This program successfully replicates the immune reaction that occurs due to interactions between epitopes and T-cell receptors45. Our analysis focused on high-frequency alleles prevalent in Xinjiang, specifically HLA-A*1101,HLA-A*0201,HLA-B*5101,HLA-B*3501,HLA-DRB1*0701,and HLA-DRB1*150130. The simulation parameters were established as follows: a random seed of 12,345, a simulation count of 50, and a step size of 1050. Three doses of immunization were administered automatically, with intervals of 1,84,and 168 time steps between the immunizations, respectively46. The three vaccinations were administered 0,28, and 56 days apart, with each shot containing a dose of 1000 units.
Prediction of secondary and tertiary structure of peptides
The secondary structure of peptides, including α-helices,β-sheets, and random coils, was predicted using the SOPMA tool (https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl? page=/NPSA/npsa_sopma.html)47. For tertiary structure prediction, the Robetta server (https://robetta.bakerlab.org/login.php) was utilized, which employs both de novo modeling and comparative modeling techniques48. The accuracy of the predicted three-dimensional structure was assessed for potential errors based on the Z-score using the ProSA-web server (https://prosa.services.came.sbg.ac.at/prosa.php)49. Additionally, the stability of the tertiary structure was evaluated using the Verify3D server (https://www.doe-mbi.ucla.edu/verify3d/)50. We also utilized SWISS-MODEL to predict the Ramachandran plots of the tertiary structure, thereby enhancing the reliability of the prediction results.
Disulfide engineering
Disulfide engineering involves the identification of potential disulfide bonds using Disulfide by Design 2.0 (http://cptweb.cpt.wayne.edu/DbD2/),a software tool that analyzes B factors in protein regions associated with the predicted disulfide bonds50. This technique has the potential to enhance the stability of vaccines51.
Predicting discontinuous B-Cell epitopes in vaccine 3D structures
We utilized Elipro to predict B-cell conformational epitopes by inputting the PDB format of the vaccine following disulfide bond engineering52. We selected results with high scores for subsequent analysis. The server employed default parameters for epitope prediction, with a minimum score of 0.5 and a maximum score of 0.653.
Molecular Docking
Molecular docking involves Toll-like receptors (TLRs), specifically Toll-like receptor-4 (TLR-4) (PDB ID: 3FXI), which play a crucial role in antigen-presenting cell activity by recognizing Brucella abortus and initiating adaptive immunity54. We aim to construct an effective vaccine and TLR-4 molecule using the HDOCK server (http://hdock.phys.hust.edu.cn/) for simulating molecular docking. Finally, the tertiary structure of the docking complex will be visualized using PyMOL.
Molecular dynamics simulations
Molecular dynamics simulations were conducted using GROMACS 2024.2, with the protein modeled using the amber14sb_parmbsc1.ff55 force field and the TIP3P water model. The box boundary was set at a distance of 1.0 nm from the system’s atoms. The simulations were performed at 298.15 K, 1 atm, and a physiological salt concentration. Initially, energy minimization was carried out with an emtol of 100.0 kJ mol⁻¹ nm⁻², followed by constrained dynamics on the protein atoms for 300 ps, utilizing a time step of 1 fs. The formal simulation was subsequently set for 100 ns with a time step of 2 fs. During the simulation, the Coulomb interactions were managed using the PME method56, with a Coulomb cutoff radius of 1.0 nm. The van der Waals (vdW) interactions were treated with a cutoff method, establishing the van der Waals cutoff radius at 1.0 nm. The pressure coupling algorithm employed was the Parrinello-Rahman method57. GROMACS built-in tools were utilized to calculate the ligand’s RMSD58, root mean square fluctuation (RMSF), radius of gyration (RG), and MM-PBSA.
Results
Physical and chemical properties of candidate proteins
Virulence proteins can effectively stimulate the body’s immune response and serve as promising immune candidate proteins59. The sequences of the virulence proteins VirB2,VirB3,VirB6,and BtpA were obtained from UniProt and saved in FASTA format for subsequent proteomic analysis. The antigenicity of the target protein, predicted using VaxiJen v2.0,exceeded the threshold of 0.4. Additionally, results from AlgPred 2.0 and ToxinPred2 indicated that the target protein is neither allergenic nor toxic. Finally, ProtParam was utilized to calculate the molecular weight, theoretical isoelectric point (pI), and overall average hydrophobicity (GRAVY) of the target protein. The results are presented in Table 1.
Prediction and excision of signal peptides
The results obtained from the online prediction software SignalP 6.0 (Fig. 1) indicate the cleavage site of VirB2 (Fig. 1A) is located between positions 36 and 37,with a probability of 0.866036. In contrast, the likelihood of VirB3 (Fig. 1B), VirB6 (Fig. 1C), and BtpA (Fig. 1D) possessing signal peptides is considerably low. These will be selected at a later stage. When identifying the dominant epitope, it is essential to remove the peptide containing the signal peptide.

(A-D) SignalP 6.0 provides predictions regarding the signal peptide of the target protein. A: Presents the prediction results for the VirB3 signal peptide,B:Illustrates the prediction results for the VirB6 signal peptide,C:Displays the prediction results for the VirB6 signal peptide,D:Provides the prediction results for the BtpA signal peptide.
T-cell epitope prediction results
The software tools IEDB and NetMHCpan-4.1 were utilized to jointly predict cytotoxic T lymphocyte (CTL) epitopes, resulting in the selection of seven dominant epitopes based on the outcomes from both programs. Concurrently, IEDB and NetMHCIIpan-4.0 were employed to predict helper T lymphocyte (HTL) epitopes, leading to the identification of six dominant epitopes. In total, seven CTL epitopes and six HTL epitopes were chosen for the subsequent construction of mRNA vaccines. Importantly, all selected dominant epitopes exhibit favorable antigenicity and are characterized as non-allergenic and non-toxic. Table 2 presents all final selected CTL dominant epitopes, while Table 3 displays the HTL dominant epitopes.
Docking results between T-Cell epitopes and HLA alleles
To evaluate the relationship between T-cell epitopes and HLA alleles, we conducted molecular docking studies. For the docking results involving HLA alleles and cytotoxic T lymphocyte (CTL) epitopes (Fig. 2A), the top-ranked model was selected, yielding a docking score of −244.52,a confidence score of 0.8688,and a ligand root mean square deviation (RMSD) of 85.58 Å. In the case of the docking results for HLA alleles and helper T lymphocyte (HTL) epitopes (Fig. 2B), the top-ranked model exhibited a docking score of −237.75,a confidence score of 0.8526,and a ligand RMSD of 170.71 Å.

The results between T-cell epitopes and HLA alleles. A: Illustrates the docking results for HLA alleles and CTL, B: Displays the docking results for HLA alleles and HTL. This image generation comes from the HDOCK SERVER.
B-cell linear epitope prediction results
The software ABCpred identified four B-cell linear epitopes of the target protein, selecting those with favorable antigenicity, non-allergenic properties, and non-toxicity. Ultimately, we selected seven B-cell linear epitopes, as presented in Table 4.
B-cell conformational epitope prediction results
Elipro predicted the B-cell conformational epitopes for four target proteins, ultimately selecting one dominant epitope each from VirB2,VirB3,and BtpA, as well as two dominant epitopes from VirB6 for subsequent mRNA construction. The results are presented in Table 5; Fig. 3(A–E).

B-cell conformational epitope prediction results. A: A: G70,A: Y71,A: M73,A: A74,A: F75,A: H77,A: A78,A: R79,A: F80,A: M81,A: D82,A: V84; B: A: E7,A: S8,A: N9,A: A10,A: R11,A: S12,A: A13,A: G14,A: Y15,A: G17; C: A: A326,A: G327,A: G328,A: N329,A: A330,A: M331,A: W332,A: S333,A: P334,A: A335,A: F336,A: R337,A: Q338; D: A: S307,A: S308,A: G309,A: G310,A: G311,A: G312,A: S313,A: G314,A: S315,A: A316,A: K317,A: A318,A: G319,A: G320,A: E321,A: S322,A: S323,A: Y324;E:A: M1,A: S2,A: K3,A: E4,A: K5,A: Q6,A: A7,A: Q8,A: S9,A: K10,A: A11,A: H12,A: K13,A: A14,A: Q15,A: Q16,A: A17,A: I18,A: S19,A: S20,A: A21,A: K22,A: S23,A: L24,A: S25,A: T26,A: Q27,A: K28,A: S29,A: K30,A: M31,A: S32,A: E33,A: L34,A: E35,A: R36,A: A37,A: T38,A: R39,A: D40,A: G41,A: A42,A: A43,A: I44,A: G45,A: K46,A: K47,A: A49,A: D50. This image is generated by ElliPro.
Construction of mRNA vaccine
The mRNA vaccine we designed is constructed sequentially from the N-terminus to the C-terminus show in Fig. 4,specifically comprising the following components: 5’ m7GCap,5’ UTR, Kozak sequence, tPA (signal peptide), The mRNA vaccine we designed is constructed sequentially from the N-terminus to the C-terminus, specifically comprising the following components: 5’ m7GCap-5’ UTR-Kozak sequence-tPA (signal peptide)-ALNTSLKSVAAYKVALNTSLKAAYSIAAIEPNLAAYILPLIVPIVAAYSISSAIFSRAAYAMLFGVPVIAAYVLSEHFFSKAAYKAKQLSSYQAKQFKAGPGPGEYDFFISHASEDKEAGPGPGLIVSIAAIEPNLAHAGPGPGLGGALVVGAAAEIASGPGPGRPAMLFGVPVIPLVIGPGPGPSSTIYQTLDNSLGKGPGPGFFSKQWPARELDGLTAKKIVSIAAIEPNLAHANGKKSVWISMFILPLIVPIVKKTIYQTLDNSLGKGWNIKKAPQESNARSAGYRGDPKKSGSSGGGGSGSAKAGGKKSELERATRDGAAIGKKKKSSGGGGSGSAKAGGESSYKKGYKMAFRHARFMDVVKKMSKEKQAQSKAHKAQQAISSAKSLSTQKSKMSELERATRDGAAIGKKRADKKAGGNAMWSPAFRQKKESNARSAGYRG-GGGSHHHHHH- stop codon-3’ UTR-Poly(A) tail.

The mRNA vaccine we designed is constructed sequentially from the N-terminus to the C-terminus, incorporating the use of linkers.
Vaccine physical and chemical properties
The molecular formula of the constructed vaccine is C2109H3369N601O610S8,with a molecular weight of 47161.09 g/mol and comprising 455 amino acids. The instability index (II) is computed to be 37.75,classifying the protein as stable. The grand average of hydropathicity (GRAVY) is −0.276. The results from the Scratch program indicate a probability of dissolution of 0.943206 when the protein is overexpressed (≥ 0.5),suggesting good dissolution ability. Furthermore, the vaccine exhibits favorable characteristics, including good antigenicity, non-allergenicity, and non-toxicity, as detailed in Table 6.
Codon optimization and computer cloning
The online software EMBOSS Backtranseq was utilized to reverse-translate the ligated protein sequence into its corresponding nucleotide sequence. Subsequently, NovoPro was employed to perform codon optimization, focusing on the excision restriction endonuclease sites BamHI and XhoI. Prior to optimization, the Codon Adaptation Index (CAI) was 0.55, and the GC content was 68.64%. After optimization, the CAI improved to 0.80, while the GC content decreased to 53.33%. BamHI and XhoI were added to the 5’ and 3’ ends of the optimized target gene, respectively. The designed primers are as follows: the forward primer, 5’GTCAGGATCCGCCACCATGGATGGACGCCATGAA, has a melting temperature (TM) of 64 °C, and the reverse primer, 5’GTCACTCGAGGTGGTGATGATGGTGATGGGAGCC, also has a TM of 64 °C.
The computer-simulated cloning process is illustrated using BioRender, as depicted in Fig. 5A. The software SnapGene was then used to perform simulated PCR on the target gene using the designed primers, resulting in an amplified product of 1473 base pairs (Fig. 5D). This amplified product was subsequently cloned into plasmid pVAX1, yielding a final cloned product of 4402 base pairs (Fig. 5B). The results of the simulated agarose gel experiment (1.0% agarose) are presented in Fig. 5C. The first lane contains the constructed mRNA vaccine, the second lane features the plasmid pVAX1, and the third lane displays the cloned plasmid.

The results of the computer simulation of the cloning process are presented as follows: A:Computer-simulated cloning process;B:Map depicting the simulated cloning products; C:Results from the simulated agarose gel experiment (1.0% agarose), where the first lane represents the mRNA vaccine, the second lane corresponds to the plasmid pVAX1, and the third lane displays the cloned product. D:Map illustrating the amplification of the simulated mRNA vaccine;.
RNA vaccine structure and protein secondary structure
The structure of RNA vaccines and their protein configurations can be analyzed using RNAfold, which predicts the secondary structure of mRNA vaccines. The optimal secondary structure exhibits a minimum free energy of −463.90 kcal/mol, while the free energy of the thermodynamic ensemble is −484.19 kcal/mol. The centroid secondary structure has a minimum free energy of −334.37 kcal/mol. These negative free energy values indicate that the formation of these structures is exothermic. A greater absolute value of free energy suggests increased stability; thus, the mRNA structure is considered highly stable. Simultaneous predictions of the minimum free energy (MFE) secondary structure (Fig. 6A) and centroid secondary structure (Fig. 6B) are presented, along with the thermodynamic integration of the MFE structure, RNA structure, and the mountain map of the centroid structure (Fig. 6C).
The software SOPMA predicted the secondary structure of the peptide, as illustrated in Fig. 6D. The analysis indicated that the alpha helix constituted 43.22% of the structure, while the extended strand represented 14.71%, and the random coil accounted for 42.07%.

The results of predicting the secondary structure of mRNA vaccines and peptides. A: The minimum free energy (MFE) secondary structure, B: The centroid secondary structure, C: The thermodynamic integration of the MFE structure, RNA structure, and the mountain map of the centroid structure, D: The secondary structure of the peptide. The images A-C are generated from RNAfold(2.6.3) and diagram D is generated from SOPMA.
Protein tertiary structure and evaluation
From the five structural models predicted by the Robetta server, we selected the model with the highest score and saved it in PDB format. Subsequently, we utilized the ProSA-web server to evaluate the tertiary structure, obtaining a z-score of −3.24 (Fig. 7 7A), local model quality (Fig. 7B), and a residue energy map (Fig. 7 C), which ranges from blue to red in order of increasing residue energy. We then employed SWISS-MODEL to generate the pull chart (Fig. 7D), which shows that 94.04% of residues are in the Ramachandran favored region, while Ramachandran outliers account for 1.55% and rotamer outliers for 0.30%. These results indicate that the majority of amino acid residues fall within acceptable limits, suggesting a high level of confidence in the prediction of this tertiary structure model.

The evaluation results of the protein tertiary structure. A: The ProSA-web server predicts the Z-score for the tertiary structure.B: Assessment of the quality of the local model.C: Residue energy map analysis.D: The Ramachandran diagram, as predicted by SWISS-MODEL.
Disulfide engineering results
Upon uploading the protein’s tertiary structure in PDB format to Disulfide by Design 2.0, the amino acid residues involved in disulfide engineering are detailed in Table 7. The yellow regions in Fig. 8 indicate the presence of the disulfide bond.

The image on the left depicts the structure prior to the introduction of a disulfide bond, while the image on the right illustrates the structure following the addition of the disulfide bond, with the bond highlighted in yellow. This image generation comes from Disulfide by Design 2.0.
Discontinuous B-Cell epitopes in the tertiary structure
In the tertiary structure, the results of the discontinuous B-cell epitopes are as follows: a total of three discontinuous B-cell epitopes were predicted, all of which exhibited very high scores. Notably, the antigenicity predictions for all three epitopes exceeded the threshold of 0.4, as shown in Table 8. The structural diagram presented in Fig. 9 indicates that after vaccine construction, the discontinuous B-cell epitopes can be effectively exposed, thereby meeting the requirements for vaccines.

he visualization results of discontinuous B-cell epitopes in the tertiary structure of the vaccine are presented.A: A: K275, A: K276, A: A277, A: P278, A: Q279, A: E280, A: S281, A: N282, A: A283, A: R284, A: S285, A: A286, A: G287, A: Y288, A: R289, A: G290, A: D291, A: P292, A: K293, A: K294, A: S295, A: G296, A: S297, A: S298, A: G299, A: G300, A: G301, A: G302, A: S303, A: G304, A: S305, A: A306, A: K307, A: A308, A: G309, A: G310, A: K311, A: K312, A: S313, A: E314, A: L315, A: E316, A: R317, A: A318, A: T319, A: R320, A: D321, A: G322, A: A323, A: A324, A: I325, A: G326, A: K327, A: K328, A: K329, A: K330, A: S331, A: S332, A: G333, A: G334, A: G335, A: G336, A: S337, A: G338, A: S339, A: A340, A: K341, A: A342, A: G343, A: G344, A: E345, A: S346, A: S347, A: Y348, A: K349;B:A: A1, A: L2, A: N3, A: T4, A: S5, A: L6, A: K7, A: S8, A: V9, A: A10, A: A11, A: C12, A: K13, A: V14, A: S52, A: C53, A: I54, A: F55, A: S56, A: R57, A: A58, A: A59, A: A61, A: R403, A: A404, A: C405, A: R406, A: D407, A: G408, A: A409, A: A410, A: I411, A: G412, A: K413, A: K414, A: R415, A: A416, A: D417, A: K418, A: K419, A: A420, A: G421, A: G422, A: N423, A: A424, A: M425, A: S427, A: P428, A: A429, A: F430, A: R431, A: Q432, A: K433, A: K434, A: E435, A: S436, A: N437, A: A438, A: R439, A: S440, A: A441, A: G442, A: Y443, A: R444, A: G445, A: G448, A: C449, A: C450, A: H451, A: H452, A: H453, A: H454, A: H455C: A: A94, A: K95, A: F97, A: K98, A: A99, A: G100, A: P101, A: G102, A: P103, A: G104, A: E105, A: C106, A: S128, A: I129, A: A131, A: I132, A: E133, A: P134, A: N135, A: L136, A: A137, A: H138, A: A139, A: G140, A: P141, A: G142, A: P143, A: V178, A: I179, A: G180, A: P181, A: G182, A: P183, A: G184, A: P185, A: S186, A: S187, A: T188, A: I189, A: Y190, A: Q191, A: T192, A: L193, A: D194, A: N195, A: S196, A: L197, A: G198, A: K199, A: G200, A: P201, A: G202, A: P203, A: G204, A: F205, A: F206, A: S207, A: K208, A: Q209, A: P211, A: A212, A: D216, A: G217, A: L218, A: T219, A: A220, A: K221, A: V224, A: C225, A: C226, A: A227, A: A228, A: I229, A: E230, A: P231, A: N232, A: L233, A: A234, A: H235, A: A236, A: N237, A: G238, A: K239, A: K240, A: S241, A: W243, A: I244. This image is generated by ElliPro.
Molecular Docking
The online prediction software HDOCK generates the top ten models. We select the first model, which has the highest score, as illustrated in Fig. 10. This model has a docking score of −350.15, a ligand RMSD of 99.20 Å, and a confidence score of 0.9821. In this study, red represents the receptor and blue denotes the ligand. We also employ PyMOL for docking visualization, with the orange regions indicating the presence of hydrogen bonds.

The molecular docking results are presented as follows: Figure A displays the surface diagram of the molecular docking complex, with the red portion representing the vaccine as the ligand and the blue portion representing the TLR4 molecule as the receptor. Figure B illustrates the molecular docking visualization performed using PyMOL 2.6.
Molecular dynamic simulation
During the simulation of the interaction between the docking complex using GROMACS2024.2 software (Fig. 11), several metrics were analyzed to assess the stability and characteristics of the interaction. In Fig. 11A, different colors represent the levels of free energy, with red indicating regions of high free energy and blue indicating regions of low free energy. Molecules are more stable in low free energy regions, and the figure illustrates the variation of free energy concerning two principal components. In the simulation of protein-ligand binding, after 10 ns of simulation, the RMSD (Fig. 11B) value of the protein backbone stabilized, indicating that the protein maintained a stable conformation without significant deformation. By superimposing the protein backbone and calculating the RMSD of the ligand, a smaller RMSD value for the ligand indicates less positional change relative to the protein. In this simulation, after 60 ns, the RMSD value of the ligand backbone also stabilized, indicating that the ligand was stably positioned at the active site. Figure 11C illustrates the variation in the radius of gyration of the complex over time. The Rg curve remains relatively stable throughout the simulation, indicating that the compactness of the molecule does not undergo significant changes and that the overall structure remains stable. The Root Mean Square Fluctuation (RMSF) was utilized to assess the flexibility of various protein regions. As illustrated in Fig. 11D to H, the fluctuation range for the receptor chains remained below 0.5 nm. This observation indicates a high level of overall structural stability, characterized by minimal conformational changes. The analysis of hydrogen bonds (Fig. 11I) indicated that the number of hydrogen bonds between the complex fluctuated between 5 and 27 throughout the simulation. This stability underscores the significant role of hydrogen bonds in maintaining protein conformation during the interaction. The results of MM-PBSA can be found in the supplementary files Figure S1-S3.

Molecular dynamics simulation results. A: A discrete free energy surface plot. B: The blue line represents the root mean square deviation (RMSD) of chain ABCD, while the purple line indicates the RMSD of both chain ABCD and chain E. C: This panel illustrates the change in the radius of gyration of chain ABCD over time. D-H: These panels display the root mean square fluctuation (RMSF) of each residue in the respective chains. I: This panel shows the variation in the number of hydrogen bonds over time during the simulation.
Immune simulation reaction
The immune simulation conducted after three injections, utilizing the C-ImmSim server for online prediction, demonstrates that the mRNA vaccine effectively activates both humoral and cellular immunity. It can be observed that after three injections, the antigen concentration initially rises rapidly and then gradually declines. In Fig. 12 A, various antibodies exhibit differing degrees of increase in response to antigen stimulation, with IgG1 and IgG2 persisting in the later stages. This persistence indicates that the body has generated an immune response and formed immune memory. As illustrated in Fig. 12B, during the initial phase of immunization, the total number of B cells experiences an initial increase followed by a subsequent decrease. In contrast, B memory cells gradually form and are maintained at a stable level in the later phase. Additionally, various subtypes of B cells, such as IgG1 and IgG2, exhibit distinct dynamic changes, which reflect the processes of differentiation and subtype switching of B cells during the immune response. Figure 12 C indicates that the organism developed long-term immune memory of T helper cells following three immunization injections. Figure 12D illustrates the activation, effector functions, and memory formation processes of cytotoxic T cells during the immune response. Figure 12E illustrates the activation and cytotoxic functions of natural killer (NK) cells during the initial stages of immune defense following immune stimulation, with their numbers gradually stabilizing thereafter. Figure 12 F illustrates that immature dendritic cells (DCs) are initially present in greater numbers, subsequently decreasing over time. Concurrently, DCs in various presenting states emerge and assume crucial roles throughout the infection process. This observation underscores the dynamic changes of dendritic cells during antigen capture, maturation, and presentation. Figure 12G illustrates that the number of active macrophages remained constant at 100 cells per cubic millimeter throughout the three-dose vaccination process. However, by 100 days post-vaccination, the quantity of active macrophages exhibited a rapid decline, stabilizing at 20 cells per cubic millimeter. Figure 12H illustrates that the quantity of epithelial cells tends to stabilize throughout the entire immune cycle. As illustrated in Fig. 12I, cytokines such as IFN-γ, IL-12, and TGF-β are concurrently involved in the immune response during the three injections.

The immune simulation conducted after three injections. A: Antibody production increased with each injection, with IgM + IgG levels. B: B cell counts following each vaccine injection exhibited an upward trend. C: The counts of helper T cells. D: The counts of cytotoxic T cells. E: The number of NK cells in three injections; F: The number of DC cells in three injections; G: The number of macrophages in three injections; H: Changes in epithelial cells across three injections; I: Changes in cytokines across three injections.
Discussion
Brucellosis, caused by the endoparasitic bacterium Brucella, is a zoonotic disease that poses a significant threat to the health and lives of both humans and animals60. Several scholars have discussed the application of the One Health approach in combating brucellosis61. This method emphasizes that the prevention and control of Brucella-related harm require collaboration among multiple agencies and disciplines, highlighting the need for increased attention from policymakers. Currently, disease control efforts primarily emphasize managing infections in animals, notably through the use of veterinary Brucella vaccines; however, these vaccines are not appropriate for human use. Although there have been instances of Brucella abortus S19 being administered to humans, it poses a risk of causing Brucella infection in this population6. Furthermore, live attenuated vaccines have demonstrated safety concerns. Therefore, it is crucial to focus on the development of effective and safe vaccines.
In this study, we employed bioinformatics and reverse vaccinology technologies to develop an mRNA vaccine that targets multiple epitopes of Brucella melitensis. Compared to traditional vaccines, mRNA vaccines are cost-effective, exhibit strong immune responses, and have fewer side effects. Previous research has demonstrated that mRNA vaccines represent a promising strategy for immunological treatment62. Because conventional peptide or subunit vaccines often fail to effectively activate the body’s immune response, mRNA vaccines, which possess self-adjuvant properties, can significantly enhance the immune response generated by subunit vaccines63. The stability of mRNA translation depends on the functional 5’ cap structure, while both the 5’ cap and the poly(A) tail are important determinants of messenger RNA translation efficiency64. To construct an mRNA vaccine, the elements such as the 3’ and 5’ UTR regions, Kozak sequences, and the nucleotide sequence of the multi-epitope protein are introduced65. Currently, mRNA can be efficiently synthesized using in vitro transcription technology, and various modified nucleotides are employed to enhance both the stability and translation efficiency of mRNA66. For example, the incorporation of modified nucleotides such as pseudouridine and N1-methylpseudouridine improves mRNA stability, reduces its degradation rate by nucleases in vivo, thereby extending its half-life and increasing the duration for protein expression67.Additionally, a His tag (HHHHHH) was incorporated at the C-terminus of the protein to facilitate subsequent protein purification68.In the design of multi-epitope proteins, we referenced the Type IV secretion system (T4SS), which is a significant pathogenic factor in Brucella69.
The delivery system is pivotal for the effective functioning and safety assurance of mRNA vaccines70. It ensures the stability of mRNA within the body, preventing enzymatic degradation, and efficiently delivers mRNA to specific immune cells, such as B cells in the spleen, to elicit immune responses71. Furthermore, it mitigates adverse effects, such as hepatotoxicity, thereby enhancing the overall performance and clinical applicability of mRNA vaccines72. However, in vitro and in vivo validation studies are critical components of research on mRNA vaccine delivery systems. In vitro cell experiments provide preliminary insights into cellular uptake efficiency, transfection efficiency, and cytotoxicity of the delivery system, offering valuable references for subsequent in vivo studies. In vivo validation assesses distribution, targeting ability, and immunogenicity in animals, thereby further optimizing the delivery system and enhancing the safety and efficacy of mRNA vaccines. In the context of protein expression and purification, we will subsequently provide optimized codons for plasmid construction, which will then be transfected into Escherichia coli (e.g., BL21)73. Single colonies will be selected on a medium containing the appropriate antibiotic for further expansion74. Following IPTG induction under suitable conditions, the cells will be sonicated and centrifuged to collect soluble proteins, which will then undergo further purification and validation75.
However, the long-term protective efficacy of mRNA vaccines warrants greater attention76. For instance, studies have shown that six months after vaccination with the COVID-19 mRNA vaccine, the neutralizing antibody titers against the original strain decrease77. Due to the negative charge and large molecular characteristics of mRNA, it is challenging for it to cross the cell membrane and enter the cell interior. Therefore, delivery systems such as lipid nanoparticles (LNPs) are required to enhance cellular uptake efficiency76,78. Most mRNA vaccines need to be stored at ultra-low or low temperatures, which imposes extremely high requirements on cold chain facilities for storage and transportation, thereby increasing costs and difficulties, especially in resource-limited regions where achieving this is challenging79. Thus, it is crucial to establish a comprehensive vaccine safety monitoring system to conduct real-time monitoring and assessment of adverse reactions to mRNA vaccines, promptly identifying and addressing potential safety issues80.
Conclusion
In this study, specifically selected the structural proteins VirB2, VirB3, and VirB6, along with the effector protein BtpA, from the T4SS for our study. We utilized bioinformatics software to assess the antigenicity, allergenicity, toxicity, and water solubility of the proteins VirB2, VirB3, VirB6, and BtpA. Our findings indicate that these proteins fulfill the criteria to be considered as vaccine candidate proteins. In this study, high-frequency alleles from Xinjiang (HLA-A*1101, HLA-A*0201, HLA-A*0301, HLA-DRB1*0701, HLA-DRB1*1501, and HLA-DRB1*0301) were utilized to identify suitable cytotoxic T cells and helper T cells for subsequent vaccine development27. Additionally, the vaccine must incorporate both linear and conformational epitopes for B cells14. To enhance the immunogenicity of the vaccine, we linked the selected seven cytotoxic T cell epitopes, six helper T cell epitopes, seven B cell linear epitopes, and five B cell conformational epitopes according to their respective subunits to create multi-epitope proteins. Subsequently, we predicted the secondary and tertiary structures of these proteins and evaluated the most suitable tertiary structures for subsequent docking with TLR4 molecules.The most critical aspect of vaccine development is its capacity to activate both humoral and cellular immunity. We utilized C-ImmSim software to simulate the immune response elicited by our vaccine61. We analyzed the high-frequency alleles in Xinjiang, specifically HLA-A*1101, HLA-A*0201, HLA-B*5101, HLA-B*3501, HLA-DRB1*0701, and HLA-DRB1*1501. Following three injections, we observed that the vaccine mRNA we designed not only increases the number of B cells and T cells but also enhances the levels of antibody production.
Data availability
The data derived from public domain information: Uniprot database (https://www.uniprot.org/).We retrieved the sequences for VirB2,which is identified by the UniProt ID P0C528; VirB3,with the UniProt ID Q9RPY2; VirB6,denoted by the UniProt ID Q8YDZ2; and BtpA, recognized by its UniProt ID Q8YF53. The data that supports the findings of this study are available in the methods.For any questions, please contact the corresponding author.
References
Ali, A. et al. Recent advancement, immune responses, and mechanism of action of various vaccines against intracellular bacterial infections. Life Sci. 314, 121332. https://doi.org/10.1016/j.lfs.2022.121332 (Feb. 2023).
Laine, C. G., Johnson, V. E., Scott, H. M. & Arenas-Gamboa, A. M. Global Estimate of Human Brucellosis Incidence, Emerg Infect Dis, vol. 29, no. 9, pp. 1789–1797, Sep. (2023). https://doi.org/10.3201/eid2909.230052
Suárez-Esquivel, M. et al. Persistence of Brucella abortus lineages revealed by genomic characterization and phylodynamic analysis. PLoS Negl. Trop. Dis. 14 (4), e0008235. https://doi.org/10.1371/journal.pntd.0008235 (Apr. 2020).
Yang, X. et al. Analysis of the Brucella melitensis epidemic in xinjiang: genotyping, polymorphism, antibiotic resistance and tracing. Ann. Clin. Microbiol. Antimicrob. 23 (1), 71. https://doi.org/10.1186/s12941-024-00724-0 (Aug. 2024).
Lin, Z. Q. et al. IL-6 and INF-γ levels in patients with brucellosis in severe epidemic region, xinjiang, China. Infect. Dis. Poverty. 9 (1), 47. https://doi.org/10.1186/s40249-020-00666-7 (May 2020).
Tian, T. et al. The development of a human Brucella mucosal vaccine: what should be considered? Life Sci. 355, 122986. https://doi.org/10.1016/j.lfs.2024.122986 (Oct. 2024).
Jia, B. et al. The clinical features of 590 patients with brucellosis in xinjiang, China with the emphasis on the treatment of complications. PLoS Negl. Trop. Dis. 11 (5), e0005577. https://doi.org/10.1371/journal.pntd.0005577 (May 2017).
Rezaei Shahrabi, A., Moradkasani, S., Goodarzi, F., Beig, M. & Sholeh, M. Prevalence of Brucella melitensis and Brucella abortus tetracyclines resistance: A systematic review and meta-analysis. Microb. Pathog. 183, 106321. https://doi.org/10.1016/j.micpath.2023.106321 (Oct. 2023).
Yang, J. et al. Evasion of host defense by Brucella. Cell. Insight. 3 (1), 100143. https://doi.org/10.1016/j.cellin.2023.100143 (Feb. 2024).
Xiong, X. et al. The virb system plays a crucial role in Brucella intracellular infection. Int. J. Mol. Sci. 22, no., https://doi.org/10.3390/ijms222413637 (Jan. 2021). 24, Art. 24.
Amjadi, O., Rafiei, A., Mardani, M., Zafari, P. & Zarifian, A. A review of the Immunopathogenesis of brucellosis. Infect. Dis. 51 (5), 321–333. https://doi.org/10.1080/23744235.2019.1568545 (May 2019).
Ke, Y., Wang, Y., Li, W. & Chen, Z. Type IV secretion system of Brucella spp. and its effectors, Front. Cell. Infect. Microbiol., vol. 5, Oct. (2015). https://doi.org/10.3389/fcimb.2015.00072
Zheng, M. et al. Effector proteins of type IV secretion system: weapons of Brucella used to fight against host immunity. Current Stem Cell. Res. & Therapy, 19, 2, pp. 145–153, https://doi.org/10.2174/1574888X18666230222124529
Yin, Z. et al. Aug., Design of multi-epitope vaccine candidate against Brucella type IV secretion system (T4SS), PLOS ONE, vol. 18, no. 8, p. e0286358, (2023). https://doi.org/10.1371/journal.pone.0286358
Shang, K. et al. Development of a novel multi-epitope vaccine for brucellosis prevention. Heliyon 10 (14), e34721. https://doi.org/10.1016/j.heliyon.2024.e34721 (Jul. 2024).
Shah, M., Rafiq, S. & Woo, H. G. Challenges and considerations in multi-epitope vaccine design surrounding toll-like receptors. Trends Pharmacol. Sci. 45 (12), 1104–1118. https://doi.org/10.1016/j.tips.2024.10.013 (Dec. 2024).
mRNA-Based. Vaccine Designing against Epstein-Barr Virus to Induce an Immune Response Using Immunoinformatic and Molecular Modelling Approaches. Accessed: Dec. 22, 2024. [Online]. Available: https://www.mdpi.com/1660-4601/19/20/13054
Frontiers |. mRNA-based platform for preventing and treating Staphylococcus aureus by targeted staphylococcal enterotoxin B. Accessed: Apr. 27, 2025. [Online]. (2024). Available: https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.1490044/full.
Bergstrom, C., Fischer, N. O., Kubicek-Sutherland, J. Z. & Stromberg, Z. R. mRNA vaccine platforms to prevent bacterial infections, Trends Mol Med, vol. 30, no. 6, pp. 524–526, Jun. (2024). https://doi.org/10.1016/j.molmed.2024.02.013
The UniProt Consortium. UniProt: the universal protein knowledgebase in 2023. Nucleic Acids Res. 51, D523–D531. https://doi.org/10.1093/nar/gkac1052 (Jan. 2023). no. D1.
Dalsass, M., Brozzi, A., Medini, D. & Rappuoli, R. Comparison of Open-Source reverse vaccinology programs for bacterial vaccine antigen discovery. Front. Immunol. 10, 113. https://doi.org/10.3389/fimmu.2019.00113 (Feb. 2019).
Doytchinova, I. A. & Flower, D. R. VaxiJen: a server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 8, p. (4). https://doi.org/10.1186/1471-2105-8-4 (Jan. 2007).
S. K et al., Development of a novel multi-epitope vaccine for brucellosis prevention. Heliyon, 10, 14, Jul. (2024). https://doi.org/10.1016/j.heliyon.2024.e34721
Sharma, N. et al. AlgPred 2.0: an improved method for predicting allergenic proteins and mapping of IgE epitopes. Brief. Bioinform. 22 (4), bbaa294. https://doi.org/10.1093/bib/bbaa294 (Jul. 2021).
Sharma, N., Naorem, L. D., Jain, S. & Raghava, G. P. S. ToxinPred2: an improved method for predicting toxicity of proteins, Briefings in Bioinformatics, vol. 23, no. 5, p. bbac174, Sep. (2022). https://doi.org/10.1093/bib/bbac174
Teufel, F. et al. SignalP 6.0 predicts all five types of signal peptides using protein Language models. Nat. Biotechnol. 40 (7), 1023–1025. https://doi.org/10.1038/s41587-021-01156-3 (2022).
Shi, H. et al. Development of innovative multi-epitope mRNA vaccine against central nervous system tuberculosis using in Silico approaches. PLoS One. 19 (9), e0307877. https://doi.org/10.1371/journal.pone.0307877 (Sep. 2024).
Tan, D. et al. Apr., Construction and efficacy testing of DNA vaccines containing HLA-A*02:01-restricted SARS-CoV-2 T-cell epitopes predicted by immunoinformatics, Acta Biochim Biophys Sin (Shanghai), vol. 56, no. 7, pp. 986–996, (2024). https://doi.org/10.3724/abbs.2024039
Zhang, F. et al. The immunogenicity of OMP31 peptides and its protection against Brucella melitensis infection in mice. Sci. Rep. 9, 3512. https://doi.org/10.1038/s41598-019-40084-w (Mar. 2019).
S. J et al., In Silico designed novel multi-epitope mRNA vaccines against Brucella by targeting extracellular protein BtuB and LptD. Scientific Reports, 14, 1, Mar. (2024). https://doi.org/10.1038/s41598-024-57793-6
Wang, Y. et al. A multi-epitope subunit vaccine based on CU/ZN-SOD, OMP31 and BP26 against Brucella melitensis infection in BALB/C mice. Int. Immunopharmacol. 127, 111351. https://doi.org/10.1016/j.intimp.2023.111351 (Jan. 2024).
Galanis, K. A. et al. Linear B-Cell epitope prediction for in Silico vaccine design: A performance review of methods available via Command-Line interface. Int. J. Mol. Sci. 22 (6), 3210. https://doi.org/10.3390/ijms22063210 (Mar. 2021).
Ponomarenko, J. et al. ElliPro: a new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 9 (1), 514. https://doi.org/10.1186/1471-2105-9-514 (Dec. 2008).
Yan, Y., Tao, H., He, J. & Huang, S. Y. The HDOCK server for integrated protein-protein Docking. Nat. Protoc. 15 (5), 1829–1852. https://doi.org/10.1038/s41596-020-0312-x (May 2020).
Yan, Y., Zhang, D., Zhou, P., Li, B. & Huang, S. Y. HDOCK: a web server for protein-protein and protein-DNA/RNA Docking based on a hybrid strategy. Nucleic Acids Res. 45, W365–W373. https://doi.org/10.1093/nar/gkx407 (Jul. 2017). no. W1.
Wei, Y. et al. Advances of computational methods enhance the development of multi-epitope vaccines. Brief. Bioinform. 26 (1), bbaf055. https://doi.org/10.1093/bib/bbaf055 (Feb. 2025).
Azami, Z., Farahmand, M. & Kavousi, M. A new multi-epitope DNA vaccine against Helicobacter Pylori infection in a BALB/c mouse model, Heliyon, vol. 10, no. 21, p. e39433, Oct. (2024). https://doi.org/10.1016/j.heliyon.2024.e39433
Felice, A. G. et al. In Silico construction of a multi-epitope vaccine (RGME-VAC/ATS-1) against the Rickettsia genus using immunoinformatics. Mem. Inst. Oswaldo Cruz. 120, e240201. https://doi.org/10.1590/0074-02760240201 (2025).
Ismail, S. & Barakat, K. Designing a multi-neoantigen vaccine for melanoma: integrating immunoinformatics and biophysics methods. Comput. Biol. Med. 190, 110081. https://doi.org/10.1016/j.compbiomed.2025.110081 (May 2025).
Zaib, S. et al. Designing multi-epitope Monkeypox virus-specific vaccine using immunoinformatics approach. J. Infect. Public Health. 16 (1, p. 107, ). https://doi.org/10.1016/j.jiph.2022.11.033 (Dec. 2022).
Jaan, S., Zaman, A., Ahmed, S., Shah, M. & Ojha, S. C. mRNA vaccine designing using Chikungunya virus E glycoprotein through Immunoinformatics-Guided approaches. Vaccines (Basel). 10 (9). https://doi.org/10.3390/vaccines10091476 (Sep. 2022).
Cn, M., P, B. & R. A, and SOLpro: accurate sequence-based prediction of protein solubility. Bioinf. (Oxford England). 25 (17). https://doi.org/10.1093/bioinformatics/btp386 (Jan. 2009).
Kishore Kumar, G. et al. Polymorphisms of extrinsic death receptor apoptotic genes (FAS – 670 G > A, FASL – 844 T > C) in coronary artery disease. Apoptosis 21 (5), 558–565. https://doi.org/10.1007/s10495-016-1232-7 (May 2016).
Y. Z et al., Nano DNA vaccine encoding Toxoplasma gondii histone deacetylase SIR2 enhanced protective immunity in mice. Pharmaceutics, 13, 10, Sep. (2021). https://doi.org/10.3390/pharmaceutics13101582
Stolfi, P. et al. In-silico evaluation of adenoviral COVID-19 vaccination protocols: assessment of immunological memory up to 6 months after the third dose. Front. Immunol. 13 https://doi.org/10.3389/fimmu.2022.998262 (Oct. 2022).
Negahdari, B., Sarkoohi, P., Nezhad, F. G., Shahbazi, B. & Ahmadi, K. Design of multi-epitope vaccine candidate based on ompa, CarO and ZnuD proteins against multi-drug resistant Acinetobacter baumannii. Heliyon 10 (14), e34690. https://doi.org/10.1016/j.heliyon.2024.e34690 (Jul. 2024).
G, D. & G. C and SOPMA: significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Comput. Appl. Biosciences: CABIOS. 11 (6). https://doi.org/10.1093/bioinformatics/11.6.681 (Dec. 1995).
Baek, M. et al. Accurate prediction of protein structures and interactions using a 3-track neural network. Science 373 (6557), 871–876. https://doi.org/10.1126/science.abj8754 (Aug. 2021).
Mj, S. W. M and ProSA-web: interactive web service for the recognition of errors in three-dimensional structures of proteins, Nucleic acids research, vol. 35, no. Web Server issue, Jul. (2007). https://doi.org/10.1093/nar/gkm290
Tn, A. Design of a novel multi-epitopes vaccine against Escherichia fergusonii: a pan-proteome based in- Silico approach. Front. Immunol. 14 https://doi.org/10.3389/fimmu.2023.1332378 (Aug. 2023).
H. M et al., Vaccinomics strategy for developing a unique multi-epitope monovalent vaccine against Marburg marburgvirus. Infection Genet. Evolution: J. Mol. Epidemiol. Evolutionary Genet. Infect. Diseases, 70, Jun. (2019). https://doi.org/10.1016/j.meegid.2019.03.003
Farzan, M. et al. Immunoinformatics-based multi-epitope vaccine design for the re-emerging Monkeypox virus. Int. Immunopharmacol. 123, 110725. https://doi.org/10.1016/j.intimp.2023.110725 (Oct. 2023).
Ponomarenko, J. et al. ElliPro: a new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 9, 514. https://doi.org/10.1186/1471-2105-9-514 (Dec. 2008).
Jy, L., Y, L., Xx, G., H, C. & G. X, and TLR2 and TLR4 signaling pathways are required for Recombinant Brucella abortus BCSP31-induced cytokine production, functional upregulation of mouse macrophages, and the Th1 immune response in vivo and in vitro. Cell Mol. Immunol. 11 (5). https://doi.org/10.1038/cmi.2014.28 (Sep. 2014).
Sorin, E. J. & Pande, V. S. Exploring the helix-coil transition via all-atom equilibrium ensemble simulations, Biophys J, vol. 88, no. 4, pp. 2472–2493, Apr. (2005). https://doi.org/10.1529/biophysj.104.051938
Essmann, U. et al. A smooth particle mesh Ewald method, The Journal of Chemical Physics, vol. 103, no. 19, pp. 8577–8593, Nov. (1995). https://doi.org/10.1063/1.470117
Parrinello, M. & Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method, Journal of Applied Physics, vol. 52, no. 12, pp. 7182–7190, Dec. (1981). https://doi.org/10.1063/1.328693
Khan, N. et al. A novel therapeutic approach to prevent Helicobacter pylori induced gastric cancer using networking biology, molecular docking, and simulation approaches. J. Biomol. Struct. Dyn. 42 (24), 13876–13889. https://doi.org/10.1080/07391102.2023.2279276 (2024).
M. M et al., Proteome-Wide screening of potential vaccine targets against Brucella melitensis. Vaccines, 11, 2, Jan. (2023). https://doi.org/10.3390/vaccines11020263
Jiao, H. et al. The mechanism of facultative intracellular parasitism of Brucella. Int. J. Mol. Sci. 22 (7), 3673. https://doi.org/10.3390/ijms22073673 (Apr. 2021).
Ghanbari, M. K. et al. One health approach to tackle brucellosis: a systematic review. Trop. Med. Health. 48 (1), 86. https://doi.org/10.1186/s41182-020-00272-1 (Oct. 2020).
mRNA, V. & Era—Mechanisms Drug Platform and Clinical Prospection. Accessed: Dec. 24, 2024. [Online]. Available: https://www.mdpi.com/1422-0067/21/18/6582#B23-ijms-21-06582
Sahin, U., Karikó, K. & Türeci, Ö. mRNA-based therapeutics–developing a new class of drugs, Nat Rev Drug Discov, vol. 13, no. 10, pp. 759–780, Oct. (2014). https://doi.org/10.1038/nrd4278
Gallie, D. R. The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 5 (11), 2108–2116. https://doi.org/10.1101/gad.5.11.2108 (Nov. 1991).
Jaan, S., Zaman, A., Ahmed, S., Shah, M. & Ojha, S. C. mRNA vaccine designing using Chikungunya virus E glycoprotein through Immunoinformatics-Guided approaches. Vaccines (Basel). 10 (9), 1476. https://doi.org/10.3390/vaccines10091476 (Sep. 2022).
Isaac, I. et al. Tetrahydropyrimidine Ionizable Lipids for Efficient mRNA Delivery, ACS Nano, vol. 18, no. 42, pp. 29045–29058, Oct. (2024). https://doi.org/10.1021/acsnano.4c10154
Zeng, J. Y. et al. Cholesterol-Derived mannosylated Polypeptide-Formed lipid nanoparticles for efficient in vivo mRNA delivery. Small Methods. 2401712, e. https://doi.org/10.1002/smtd.202401712 (Apr. 2025).
Zhao, D. & Huang, Z. Effect of His-Tag on Expression, Purification, and Structure of Zinc Finger Protein, ZNF191(243–368), Bioinorg Chem Appl, vol. p. 8206854, 2016, (2016). https://doi.org/10.1155/2016/8206854
Ke, Y., Wang, Y., Li, W. & Chen, Z. Type IV secretion system of Brucella spp. And its effectors. Front. Cell. Infect. Microbiol. 5, 72. https://doi.org/10.3389/fcimb.2015.00072 (2015).
Liu, J. et al. Ionizable lipids with branched linkers enhance the delivery of mRNA vaccines. ACS Appl. Mater. Interfaces. 17 (9), 13552–13564. https://doi.org/10.1021/acsami.4c21289 (Mar. 2025).
Wang, F. et al. A Spleen-Targeted tolerogenic mRNA-LNPs vaccine for the treatment of experimental asthma. Adv. Sci. (Weinh). 12, e2412543. https://doi.org/10.1002/advs.202412543 (Apr. 2025).
Suzuki, Y. et al. Splenic B cell-targeting lipid nanoparticles for safe and effective mRNA vaccine delivery. J. Control Release. 382, 113687. https://doi.org/10.1016/j.jconrel.2025.113687 (Apr. 2025).
Yuan, L. et al. A broad-spectrum multiepitope vaccine against seasonal influenza A and B viruses in mice. eBioMedicine 106 https://doi.org/10.1016/j.ebiom.2024.105269 (Aug. 2024).
Gomes, L., Monteiro, G. & Mergulhão, F. The impact of IPTG induction on plasmid stability and heterologous protein expression by Escherichia coli biofilms. Int. J. Mol. Sci. 21 (2), 576. https://doi.org/10.3390/ijms21020576 (Jan. 2020).
Murakami, M., Murakami, A. M. & Itagaki, S. A dual prokaryotic (E. coli) expression system (pdMAX). PLoS One. 16 (10), e0258553. https://doi.org/10.1371/journal.pone.0258553 (2021).
Heiser, B. J., Veyssi, A. & Ghosh, D. Recent strategies for enhanced delivery of mRNA to the lungs. Nanomed. (Lond). 1–27. https://doi.org/10.1080/17435889.2025.2485669 (Apr. 2025).
Azeem, M. et al. Efficacy and limitations of SARS-CoV-2 vaccines - A systematic review. Life Sci. 371, 123610. https://doi.org/10.1016/j.lfs.2025.123610 (Jun. 2025).
Wu, K. et al. Dec., Characterization of mRNA-LNP structural features and mechanisms for enhanced mRNA vaccine immunogenicity, J Control Release, vol. 376, pp. 1288–1299, (2024). https://doi.org/10.1016/j.jconrel.2024.11.007
Leong, K. Y., Tham, S. K. & Poh, C. L. Revolutionizing immunization: a comprehensive review of mRNA vaccine technology and applications. Virol. J. 22 (1), 71. https://doi.org/10.1186/s12985-025-02645-6 (Mar. 2025).
Jeon, J. & Kim, E. Exploring future pandemic preparedness through the development of preventive vaccine platforms and the key roles of international organizations in a global health crisis. Vaccines (Basel). 13 (1), 56. https://doi.org/10.3390/vaccines13010056 (Jan. 2025).
Acknowledgements
Not applicable.
Funding
This work was supported by Youth Science and Technology Top Talent Program (2022TSYCCX0112); Science and Technology Department of Xinjiang Uygur Autonomous Region, Outstanding Youth Project(2022D01E69); National Natural Science Foundation of China and regional science foundation projects(82360394,82460399).
Author information
Authors and Affiliations
Contributions
LJR is responsible for writing the original manuscript and editing. QXX is in charge of conducting full-text grammar checks and writing the abstracts. TTT oversees the methods and materials section, while SKY is responsible for simulating secondary and tertiary structures. SHD handles computer cloning, and LC is tasked with visual analysis. CZL conducts immune simulations, DJB performs a full-text review. ZFB and ZYJ are responsible for critical review, providing comments on the full text, and revising the title. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Luo, JR., Qi, XX., Tian, TT. et al. Design of a multi-Epitope mRNA vaccine against Brucella type IV secretion system using reverse vaccinology and immunogenicity approaches. Sci Rep 15, 30698 (2025). https://doi.org/10.1038/s41598-025-09509-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-09509-7
- Springer Nature Limited