
Abstract
The SARS-CoV-2 nucleocapsid protein (N) is responsible for RNA binding. Here we report the crystal structure of the C-terminal domain (NCTD) in open and closed conformations and in complex with guanine triphosphate, GTP. The crystal structure and biochemical studies reveal a specific interaction between the guanine, a nucleotide enriched in the packaging signals regions of coronaviruses, and a highly conserved tryptophan residue (W330). In addition, EMSA assays with SARS-CoV-2 derived RNA hairpin loops from a putative viral packaging sequence showed the preference interaction of the N-CTD to RNA oligonucleotides containing G and the loss of the specificity in the mutant W330A. Here we propose that this interaction may facilitate the viral assembly process. In summary, we have identified a specific guanine-binding pocket in the N protein that may be used to design viral assembly inhibitors.
Similar content being viewed by others

Introduction
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has emerged as a pandemic virus causing a global human health crisis. SARS-CoV-2 shares ≈80% genome identity with SARS-CoV1. At the 5′-terminus two-thirds of its genome, there are located the ORF1a and ORF1a/b encoding 16 non-structural proteins (NSPs 1-16), which are responsible for establishing the cellular conditions favorable for viral infection and viral mRNA synthesis2,3. The 3′-terminus one-third of the genome contains the ORFs for the four structural proteins: Spike (S), Envelope (E), Membrane (M), and Nucleocapsid (N) and other accessory proteins3. S, E and M are transmembrane proteins that are incorporated into the viral lipid envelope4. The protein N is a multifunctional protein that packs the genomic RNA and enhances the efficiency of virus transcription and assembly5,6,7,8,9,10,11,12,13. N contains an N-terminal domain, involved in RNA binding, and a C-terminal (NCTD) involved in RNA binding, protein dimerization, and interaction with the viral membrane protein (M)14,15. In the case of the Murine Hepatitis coronavirus, MHV (a betacoronavirus) the NCTD has been implicated in the interaction with a conserved packaging signal (PS) necessary for viral RNA packaging16.
To date, 11 crystallographic structures of the SARS-CoV-2 NCTD have been deposited at the protein databank presenting minor differences between them (PDB: 7N0I, 7F2B, 7F2E, 7C22, 7CE0, 6ZCO, 6YUN, 6WZO, 6WZQ, 6WJI, 7DE1, 7F2B, 7F2E). Some of them have been already published and others are pending publication17,18,19,20,21,22, however, none of these contain RNA or nucleotides that provide any insight into the RNA recognition process. The SARS-CoV-2 NCTD folds into a helical core with a protruding β-hairpin employed for protein dimerization. In this study, we report three SARS-CoV-2 NCTD structures and provide a co-crystal structure with GTP that might provide an insight into the specificity of the PS recognition.
Results
Structure of SARS-CoV-2 NCTD in open and closed conformations
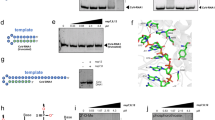
The crystal structure of NCTD was solved by molecular replacement to a resolution of 1.94 Å (Table 1). The crystal asymmetric unit contains two homodimers of NCTD with almost identical conformation (rmsd 0.14 Å). As described previously, it folds into five α-helices (α1–α5), two 310 helices (η1–η2), and two β-strands (β1–β2), presenting the following N- to C-sequence for the structural elements: η1–α1–α2–α3–α4–β1–β2–α5–η217,18,19,20,21 (Fig. 1a and Supplementary Fig. 1a). Two monomers are intertwined through the β-hairpin forming a four antiparallel β-sheet on one face of the dimer (Fig. 1). In striking contrast to the previously described structures, the superposition of the subunits in our structure shows a ≈5.5 Å movement of the β-hairpin (Fig. 1b). One subunit presents the β-hairpin in an extended and previously unseen conformation that we named “open”. Whereas the other subunit shows a β-hairpin in a flexed conformation that we named “closed”, similar to other structures of this protein (Fig. 1b and Supplementary Fig. 1b). The alternative conformation of the β-hairpin is associated with the structural displacement of the loop between α1 and α2 (residues 280–283) (Fig. 1b).

a Cartoon representation of the NCTD dimer. Each monomer is colored in blue and orange, respectively. The β-hairpin and the loop connecting the helices α1–α2 are highlighted in dark tones. Secondary structural elements and residues are numbered and labeled in order from N to C terminus: the symbol η corresponds to 310 helix; α to α-helix and β to β-strand. b Superimposition of the NCTD monomers. The common helical core is colored in gray. c, d Detailed view of the open (c) and the closed (d) conformations. The side chain of key residues is shown in sticks with carbon atoms colored according to the monomer to which they belong. Hydrophobic and polar interactions are represented as dashed black lines. The C-terminal of the symmetric molecule (sym) is colored in magenta. The symmetric Pro residue is indicated with an apostrophe. Nitrogen and oxygen atoms are colored in blue and red, respectively.
In the closed conformation the β-hairpin interacts with the C-terminal proline residue (P364) of a symmetry-related protein (Fig. 1c, d and Supplementary Fig. 1b). This interdimeric interaction between W330 and P364 through a π–π stacking (face-to-face rings interaction), and making hydrogen bonds with T325 and S327 (Fig. 1d), is a common feature with seven NCTD structures determined previously. But strikingly, our structure shows that in the open conformation, the side chain of the residue W330 is rotated 180° towards the β-hairpin making hydrophobic contacts with the side chain of the residues T325 and S327 (Fig. 1c). Therefore, our structure shows that upon a side-chain arrangement the β-hairpin changes from an open conformation to a closed conformation that might favor the interdimeric interaction.
Previous structures of SARS-CoV-2 NCTD showed that the β-hairpin that does not interact with the symmetry-related molecule has a molecule of acetate (PDB 7C22)20 or sulfate (PDB 6WZQ)19 interacting with the side chain of W330 (Supplementary Fig. 2a, b).
Structure of SARS-CoV-2 NCTD bound to GTP
Based on the structural data we hypothesized that W330 could be a suitable residue for RNA recognition, as W330 could interact with the phosphate moiety, similarly to the sulfate, or via π–π stacking with the nitrogen base. To test our hypothesis, we attempted to individually co-crystallize NCTD with the oxynucleotides UTP, ATP, CTP, or GTP. Although crystals were obtained in all the mixtures, only the GTP was co-crystallized with NCTD. The structure of the binary complex NCTD-GTP was solved in two different space groups, P21 and P1, to a resolution of 1.8 and 2 Å, respectively (Table 1). Both crystal forms contain two protein dimers in the asymmetric unit. However, in the space group P21 there is one molecule of GTP bound to one of the dimers, whereas in the P1 the asymmetric unit contains two molecules of GTP, each bound to one dimer (Fig. 2). The electron density map is well defined for the three GTP molecules, which present temperature factors that increase from the guanine moiety to the phosphates (Fig. 2 and Supplementary Fig. 3a), suggesting that the guanine is well anchored to the protein and the phosphates are more flexible. In fact, the phosphates β and γ in the crystal P1 show two alternative dispositions, one identical to the crystal P21 (Fig. 2). In the two crystal forms the GTP binds to a cleft between the two subunits of the dimer, and adjacent to the β-hairpin in the closed conformation (Fig. 3a). The cleft corresponds to a cavity adjacent and perpendicular to W330, reducing the accessibility of the tryptophan to the solvent (Fig. 3a and Supplementary Fig. 3b).

Cartoon representation of the two dimers contained in the asymmetric unit of the crystal in the space group P21 (a) and P1 (b). Each monomer is colored in white and black for one dimer, and orange and blue for the second dimer. GTP is represented in sticks with carbon atoms colored green. The electron density map 2Fo−Fc (σ = 1) of the GTP is represented in blue. The moieties guanine, ribose, and phosphates are labeled. Nitrogen, oxygen, and phosphorus atoms are colored in blue, red, and orange, respectively. The temperature factors (B-factors) of nitrogen and oxygen atoms of the nucleoside and phosphorus atoms of phosphates are indicated below.

a Upper panel, the NCTD dimer in complex with GTP is represented in the cartoon. Each monomer is colored in blue and orange, respectively. The β-hairpins of the dimer are labeled and highlighted in dark tones. The GTP molecule is shown in sticks with its electron density map 2Fo−Fc (σ = 1) in green color. The close view of the GTP binding site is represented below. The side chain of key residues and the GTP molecule are shown in sticks with carbon atoms colored according to the monomer to which they belong. H-bond interactions of the ligand are represented as dashed black lines. The C-terminal of the symmetric molecule (sym) is colored in magenta with residues indicated with apostrophes. Secondary structural elements are numbered and labeled in order from N to C terminus. Nitrogen, oxygen, and phosphorus atoms are colored in blue, red, and orange, respectively. b Quenching of fluorescence (Fo/F) represented against increasing concentration of acrylamide for the NCTD and NCTD-W330A proteins in the presence or absence of GTP. Trajectories are fitted to linear regression. Statistical differences are indicated with asterisks (****p < 0.0001, n.s. p > 0.05). c Thermal unfolding curves of NCTD and NCTD-W330A in the presence or absence of GTP. Trajectories are fitted to a sigmoid. The corresponding melting temperature (Tm) is indicated. d GTP binding quantified thermophoretically. GTP is titrated to a constant amount of fluorescently labeled NCTD and NCTD-W330A. The binding affinity (Kd) is indicated. The error bar represents the standard error of the mean of at least four experiments.
One side of the guanine ring establishes a π–π T-shaped interaction (edge-to-face) with the indole ring of W330 and hydrophobic interactions with K338 (Fig. 3a). The other side of the guanine ring stacks over the guanidinium group of R259, which makes hydrogen bonds with the OH– groups of the ribose ring (Fig. 3a). R259 and R262 are responsible for multiple hydrogen-bonding contacts with the β- and γ-phosphates (Fig. 3a). At the bottom of the cleft the guanine moiety interacts with the M317 and with the main chain of the residues K338 and A336 (Fig. 3a). These interactions suggest high specificity for guanine, as other nitrogen bases have not as many favorable interactions as the guanine (Supplementary Fig. 4). For instance, adenine lacks the C6 oxygen that makes interactions with the M317, K338, and A336 in the case of guanine (Supplementary Fig. 4). All protein–ligand contacts are summarized in the Supplementary Table 1. Importantly, the C-terminal tail of the symmetry-related dimer also contributes to the GTP binding, covering the cleft as a lid, with Van der Waals and hydrophobic interactions between T362´ and the ribose and with the π–π stacking between P364´ and W330 (Fig. 3a). The structure suggests that the GTP binding favors swapping of the β-hairpin and interdimeric interaction.
Characterization of GTP binding
To confirm our structural results, we studied the binding of GTP in solution using the intrinsic fluorescence of tryptophan. The protein contains two tryptophan residues: W330, located in the β-hairpin and exposed to the solvent, and W301 partially buried in the protein core (Supplementary Fig. 5a). NCTD shows a fluorescence peak profile with a maximum fluorescence emission centered at 340 nm that decreases concomitantly with the increasing concentration of acrylamide, confirming the tryptophan fluorescence is quenchable (Fig. 3b and Supplementary Fig. 5b).
We also made the mutant NCTD-W330A, replacing W330 with alanine, and confirmed that the fluorescence of the remaining W301 is also quenchable (Fig. 3b). We determined the quenching of the fluorescence in the absence or presence of 0.5 mM GTP. Whereas GTP decreased significantly the quenching slope of NCTD (from 6.31 ± 0.08 to 5.17 ± 0.09; p-val ****, n = 7 and n = 4), the quenching of NCTD-W330A (1.99 ± 0.06; n = 4) is not affected by the presence of GTP (1.74 ± 0.06; n = 4), indicating that only the accessibility of W330 is affected by GTP (Fig. 3b and Supplementary Tables 2 and 3). This effect is not observed in the presence of ATP or UTP, but it is shown in the presence of CTP to a much lesser degree (p-val **) (Supplementary Fig. 6 and Supplementary Table 3). We hypothesized that the binding of GTP to the homodimerization interface could stabilize the protein dimer. To confirm this hypothesis, we perform differential scanning fluorimetry (DSF) experiments with NCTD and NCTD-W330A in the presence or absence of 5 mM GTP (Fig. 3c). NCTD presents a sigmoidal trajectory in response to temperature, with a melting temperature (Tm) of 48.96 °C (±0.03) that increased to 49.97 °C (±0.03) in presence of GTP (Fig. 3c). This difference is statistically significant (Aikake test, see the “Methods” section). In contrast, the mutant NCTD-W330A shows a Tm = 47.98 °C (±0.07) unaffected by the presence of GTP, Tm = 47.67 (±0.07) (Fig. 3c).
We also measure the binding affinity (Kd) for GTP of NCTD and NCTD-W330A using Microscale Thermophoresis (MST) (Fig. 3d and Supplementary Table 4). NCTD presents a Kd value of 196 μM for GTP, which increases to 858 μM in the mutant NCTD-W330A. This result confirms the participation of W330 for the GTP binding in solution (Fig. 3d). Although the affinity for GTP is in μM range, it should be taken into account that this Kd value corresponds to the affinity of one nucleotide of a protein that binds RNA, in which a synergistic effect of multiple nucleotide interactions would favor the avidity (see below for RNA hairpin binding affinity).
W330 confers specificity of the N-CTD to the guanine-containing RNA oligonucleotides
Phylogenetic analysis suggests the existence of conserved short structured repeats, termed repetitive structural motifs or RSM in the gRNAs from coronaviruses, that may facilitate the genome packaging by specific interaction with protein factors23. These RNA elements are called stem-loop (SL) 1–6 and SL5 is the most interesting as displays a conserved sequence 5′-UUYCGU-3′ in the tripartite apical substructures, SL5a–c. Interestingly these sequences contain a highly conserved G (in bold) in nearly all alpha- and beta-coronavirus23.
We analyzed the binding of the N-CTD to an RNA oligonucleotide derived from the apical region of the stem-loop 5a (SL5a) of SARS-CoV-2 gRNA (Oligo-G, Fig. 4). Incubation of increasing concentrations of NCTD to the Oligo-G showed the binding and the protein–RNA complex formation, Kd = 32 ± 5 nM, (Fig. 4). Interestingly mutation of the only G to U, Oligo-U, induces a decrease in the binding affinity, Kd = 140 ± 29 nM (Fig. 4a, b). To study the role of the W330, we generated and purified the mutant NCTD-W330A. EMSA assays showed that the mutant W330A binds less efficiently to the oligonucleotide Oligo-G (Kd = 130 ± 1 nM, n = 3) than the NCTD-wt to Oligo-G (Kd = 32 ± 5 nM, n = 3), and with a similar affinity to Oligo-U (W330A:Oligo-U Kd = 90 ± 19 nM, n = 3 and NCTD-wt:Oligo-U Kd = 140 ± 34 nM, n = 3), (Fig. 4a and b). This supports the specific recognition of the guanine in the RNA hairpin by the N-CTD and that the mutation W330A losses this specific recognition.

a Ag-EMSA gels of titration experiments. Binding of NCTD and NCTD-W330A to an ssRNA oligonucleotide from the apical part of the SL5a region of SARS-CoV-2 gRNA with conserved guanine (arrowhead, Oligo-G) or with a G to U mutation (Oligo-U, lower panels). b ssRNA oligonucleotide shift with increasing concentration of protein. Protein–ssRNA combinations are represented in different colors. Trajectories are fitted to a sigmoidal one-site binding model. The error bar represents the standard error of the mean of at least four experiments.
Discussion
In coronavirus assembly of the genomic RNA, gRNA, is a fundamental problem as infection produces several copies of subgenomics RNAs, sgRNA. The specific interaction of N to PS sequences will assure the packaging of gRNA versus other sgRNAs, mRNAs from the host and other RNAs from the infected cells24. In the alpha- and beta-coronavirus, like SARS-CoV, SARS-CoV-2 and MERS-CoV phylogenetic data suggest the conservation of several of the RNA stem-loops (SL) such as SL5a-c that might constitute regions that participate as authentic PS23. Although we did not succeed to co-crystallize N-CTD with the SL5a hairpin, we found that the NCTD is able to specifically recognize the SL5a apical sequence from SARS-CoV-2, supporting previous suggestions indicating that this protein domain might be the region of the N protein able to recognize specific features of coronavirus gRNA24. If the C-terminal domain of the M membrane protein of SARS-CoV-2 is implicated in this recognition as others have found in murine coronavirus16,24, will need further studies.
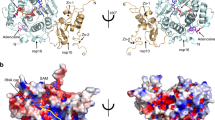
Comparison to other human CoVs NCTD reveals that there are fifteen invariant residues among human coronaviruses, three of which belong to the identified GTP binding pocket (R259, R262, F274; numbers corresponding to SARS-CoV-2) (Fig. 5). The pocket is conserved in SARS-CoV-1, and presents only one conservative change in MERS that intriguingly correspond to the swinging W330, which changes to F (Fig. 5). The structural comparison with SARS-CoV-1 (PDB 2GIB)25, MERS (PDB 6G13)26, and NL63 (PDB 5EPW)27 further suggests that the cleft is also conserved in other human coronaviruses (Fig. 5a–d). Intriguingly, the equivalent swinging tryptophan in SARS-CoV-1 also presents two rotamers, suggesting that the conformational mechanism of the β-hairpin described here could be common for both viruses (Fig. 5a, b). Moreover, the structure of MERS NCTD shows a molecule of trimethylamine oxide (TMO) that mimics the interactions shown here with the phosphates of GTP, strongly suggesting that the cleft of MERS NCTD could also be a suitable pocket for ligand binding (Fig. 5c).

Cartoon representation of the GTP binding pocket in SARS-CoV-2 (a), SARS-CoV-1 (PDB 2GIB) (b), MERS (PDB 6G13) (c) and NL63 (PDB 5EPW) (d). Each monomer of the dimer is represented in different colors. The residues that mediate the GTP binding in SARS-CoV-2 and equivalent residues in other coronaviruses are shown in sticks with carbons in the same color as the monomer to which they belong. The GTP molecule is represented in semi-transparent sticks with carbon atoms in green. The trimethylamine oxide (TMO) bound to MERS is shown in sticks with carbons in cyan. e Sequence alignment of NCTD of human coronaviruses. Invariant residues are in bold and indicated with an asterisk above the sequence. Residues for the GTP binding are in bold and highlighted in blue, and conservative changes are in light green. Secondary structural elements of the SARS-CoV-2 NCTD are represented below the sequence and labeled and numbered from N to C. η corresponds to 310 helices; α to α-helices and β to β-strands.
Moreover, our results suggest that guanine binding would promote the oligomerization of the N protein through interdimeric interactions to form the ribonucleoparticle. This sequence and structural conservation expand the possibility of using this guanine-binding pocket as a therapeutic target against SARS-CoV-2, SARS-CoV-1, highly probable against MERS, and possibly against previous human coronaviruses as NL63. In addition, the comparison of the guanine-binding pocket of NCTD with structurally similar human proteins that also bind guanine nucleotides, such as Rho6, RaI-A, and CD38 (Supplementary Fig. 7) shows low conservation in the set of residues and their structural organization for the nucleotide binding, suggesting that the design of specific compounds against SARS-CoV-2 NCTD may be feasible.
Methods
DNA methods
The synthetic gene for the SARS-CoV-2 NCTD (residues 256–364) cloned into the vector pET28a(+) was provided by Biomatik. The insert is in frame with the N-terminal His6-tag, and a TEV protease cleavage site. This construction was used as a template to introduce the mutation W330A using the Q5-Site Direct Mutagenesis Kit (NEB) with the oligonucleotides W330A-fw 5′-GAGCGGTACCGCCCTGACCTATACCG- and W330A-rv 5′-GGGGTAACTTCCATGCCA-. The final construct was sequenced in the IBV-Core Sequencing service.
Protein production and purification
E. coli BL21 (DE3) (Novagen) cells transformed with NCTD and NCTD-W330A constructs were grown at 37 °C in LB medium supplemented with 33 g/ml kanamycin to exponential phase (optical density of O.D. = 0.6, measured at λ = 600 nm), and then, protein overexpression was induced by addition of 1 mM isopropyl-β-d-1-thiogalactopyranoside (IPTG) for 16 h at 20 °C. After induction, cells were harvested by centrifugation at 4 °C for 30 min at 4000 × g, resuspended in buffer A (150 mM Tris–HCl pH 7.5, 250 mM NaCl) supplemented with 1 mM phenylmethanesulfonyl fluoride (PMSF) and lysed by sonication. The clarified lysate was loaded onto a 5 mL His Trap HP column (GE Healthcare) pre-equilibrated in buffer A. Following through wash with 20 column volumes of buffer A supplemented with 10 mM imidazole, the protein was eluted with buffer A supplemented with 500 mM imidazole. The sample was concentrated using an Amicon Ultracentrifugation device with 10 kDa cutoff, and dialyzed against buffer B (25 mM Tris–HCl pH 7.5 and 250 mM NaCl). The dialyzed protein was flash-frozen in liquid nitrogen and stored at −80 °C.
Fluorescence spectroscopy measurements
Protein fluorescence quenching was determined in a TECAN Spark instrument using acrylamide as a quencher. Samples were prepared at 5 μM protein and increasing concentration of acrylamide (0–1 M) in the absence or presence of 0.5 mM GTP. Samples of protein with or without GTP, and the acrylamide were prepared by separate at 2× in buffer B. Equal volumes of sample and acrylamide were mixed to a final volume of 50 μl (1×). Sample fluorescence was measured in 96-well black plates exciting at λex = 295 nm (bandwidth of 5 nm; step size of 1 nm) and collecting the emission between λem = 312 and 380 nm (bandwidth of 5 nm; step size of 1 nm).
The emission at 340 nm was used for calculation. Quenching was calculated by dividing the fluorescence of the sample in absence of acrylamide (Fo) by the fluorescence of the sample in the presence of acrylamide (F), (ratio Fo/F) at each acrylamide concentration.
Differential scanning fluorimetry (DSF)
The structural integrity of the protein at increasing temperature was performed using SYPRO Orange as an external fluorophore in a 7500 Real-Time PCR System (Applied Biosystems). 20 μl samples were prepared at 5x SYPRO Orange fluorophore (Sigma), 80 μM of protein and 5 mM GTP (when indicated) in buffer B. Samples were loaded in 96-well PCR plates and heated from 20 to 85 °C with a heating rate of 1 °C/min. Fluorescence intensity was normalized and analyzed using GraphPad Prism software.
MicroScale termophoresis (MST)
His-tagged NCTD was fluorescently labeled with NT-647 amine-reactive dye from the Monolith NT Protein Labeling kit RED-NHS (NanoTemper Technologies) according to the manufacturer’s instructions. The protein was prepared at 20 μM in 250 mM NaCl and mixed with 3x dye solution (1:1 volume), the mixture was incubated 30 min in dark at room temperature after soon the labeled protein was purified to remove free dye by gravity column with buffer C (25 mM Tris–HCl pH 7.5, 250 mM NaCl and 0.1% pluronic acid F127). Labeled NCTD protein was used at a final concentration of 20 nM. A 16-point twofold dilution series (ranged from 50 mM) of GTP in buffer C was mixed with NCTD-labeled solution (1:1), and incubated 15 min at room temperature. Samples were filled into premium-coated capillaries (K005, NanoTemper Technologies) and MST measurement was performed on a Monolith NT.115 (NanoTemper Technologies) at 25 °C in RED channel using a 77% LED excitation power and MST power of 40%. Analyses were performed with M.O. Affinity Analysis software (NanoTermper Technologies)28 using the signal from an MST on-time of 1.5 s.
Crystallization
Crystals were grown in sitting drops at 21 °C with a vapor-diffusion approach. Crystallization trials were set up in the Crystallogenesis service of the IBV-CSIC using commercial screenings JBS I and II (JENA Biosciences) and JCSG+ (Molecular Dimensions) in 96-well plates. Crystallization drops were generated by mixing equal volumes (0.3 µl) of each protein solution and the corresponding reservoir solution and were equilibrated against 100 µl reservoir solution. His tagged NCTD was crystallized at a protein concentration of 8 mg/ml in a reservoir solution consisting of 0.1 M tri-sodium citrate pH 4.5; 0.1 M bisTris pH 5.5 and 25% PEG 3350. Crystals were cryoprotected with a solution consisting on reservoir solution supplemented with 10% glycerol.
For crystallization in complex with GTP, his tagged NCTD at 15 mg/ml was incubated overnight at 4 °C with 5 mM GTP. Crystals were grown in a reservoir solution consisting on 30% PEG 3350 and 0.1 M sodium acetate pH 4.6. Crystals were cryoprotected with a solution consisting of 30% PEG 3350, 10% glycerol, and 0.1 M sodium acetate pH 4.6. Both crystal forms (P21, P1) were obtained within the same crystallization drops.
Data collection and model building
X-ray diffraction was performed at 100 K in an XALOC beamline at ALBA synchrotron (Barcelona, Spain). Processing of collected data collection was performed with XDS and iMosflm programs29,30, and the data were reduced with Aimless31 from the CCP4 suite32. The data-collection statistics for the best data sets used in structure determination are shown in Extended Data Table 1. In crystal form I, the spacegroup P21 was selected during automatic data processing, and although orthorhombic space groups were also explored for this data set, no correct solutions were found in that crystal system, likely due to the fact that the two dimers contained in the asymmetric unit are not entirely identical and no higher symmetry can be applied. Phases were obtained by molecular replacement using Phaser program33 and PDB 6WZO19 as search model. Model building and refinement were done by iterative cycles using Refmac34 and Coot35. Data refinement statistics are given in Extended Data Table 1.
Electrophoretic mobility shift assay (EMSA)
Single strand oligonucleotides 5′-CGGUUUCGCCG-3 and 5′-CGGUUUCUCCG-3 labeled on 5′ with IRDye-800CW fluorophore (IDT) were used at a final concentration of 300 nM and serial dilution of protein ½ from 95 to 0 μM protein. Mixtures were prepared at a final volume of 10 μl and incubated at room temperature for 20 min. EMSA was performed on agarose 0.5% gels in acidic conditions on a buffer consisting of 0.355% (m/v) β-alanine adjusted to pH 4 with glacial acetic acid. 3 μl of methyl green loading buffer (50% glycerol, traces of methyl-green, 0.15% (v/v) glacial acetic acid adjusted to pH 5 with KOH) were added to the mixtures and gels were run at 4 °C for 20 min at 100 V. Gels were revealed in an Odyssey Imaging System (LI-COR) and images analyzed using ImageStudio. Quantification was done by normalizing the signal from the free oligo to 1.0 at 0 μM protein and plotting the free oligo versus the concentration of the N-CTD protein (in Log scale). Data were adjusted to a nonlinear curve of the form One site equation (Prism 6 software). Data of at least three independent experiments were plotted. Error bars represent the standard error of the mean.
Statistics and reproducibility
Statistical analyses were used in all graphs represented in Figs. 3 and 4. Figure 3b was analyzed using a one-way ANOVA with Tukey’s multiple comparison test. Figure 3c was analyzed by comparing the nonlinear fitting curve using the Aikake test (AIC) with a >99.9% probability that the Tm of N and Tm of N + GTP is different, with an AIC of 221.1 (using GraphPad prism 6.0). In Fig. 4 statistics was analyzed by comparing the nonlinear fitting curve (one site equation in GraphPad Prism 6.0) using the Aikake test (AIC) with a >99.9% probability that the Kd of the binding of N-CTD to Oligo-G is statistically different from the Kd of the binding of N-CTD to Oligo-U is with an AIC of 24.1.
Data availability
The coordinates and structure factors of the crystallographic structures have been deposited in the Protein Data Bank with accession codes PDB 7O05, 7O35 and 7O36. Graph raw data and gel images are deposited at Mendeley Data at the address https://data.mendeley.com/datasets/8cfsrmd8by/draft?a = a0d0ad46-dfb5-474e-b9c5-37c59410e6a3.
References
Zhou, P. et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 270–273 (2020).
Prentice, E., McAuliffe, J., Lu, X., Subbarao, K. & Denison, M. R. Identification and characterization of severe acute respiratory syndrome coronavirus replicase proteins. J. Virol. 78, 9977–9986 (2004).
Wu, F. et al. A new coronavirus associated with human respiratory disease in China. Nature 579, 265–269 (2020).
Neuman, B. W. & Buchmeier, M. J. Supramolecular architecture of the coronavirus particle. Adv. Virus Res. 96, 1–27 (2016).
Cong, Y. et al. Nucleocapsid protein recruitment to replication-transcription complexes plays a crucial role in coronaviral life cycle. J. Virol. 94, e01925-19 (2020).
Surjit, M., Liu, B., Chow, V. T. K. & Lal, S. K. The nucleocapsid protein of severe acute respiratory syndrome-coronavirus inhibits the activity of cyclin-cyclin-dependent kinase complex and blocks S phase progression in mammalian cells. J. Biol. Chem. 281, 10669–10681 (2006).
Surjit, M., Liu, B., Jameel, S., Chow, V. T. K. & Lal, S. K. The SARS coronavirus nucleocapsid protein induces actin reorganization and apoptosis in COS-1 cells in the absence of growth factors. Biochem. J. 383, 13–18 (2004).
Zeng, Y. et al. The nucleocapsid protein of SARS-associated coronavirus inhibits B23 phosphorylation. Biochem. Biophys. Res. Commun. 369, 287–291 (2008).
Zhang, L. et al. SARS-CoV nucleocapsid protein induced apoptosis of COS-1 mediated by the mitochondrial pathway. Artif. Cells Blood Substit. Immobil. Biotechnol. 35, 237–253 (2007).
Lu, X., Pan, J., Tao, J. & Guo, D. SARS-CoV nucleocapsid protein antagonizes IFN-β response by targeting initial step of IFN-β induction pathway, and its C-terminal region is critical for the antagonism. Virus Genes 42, 37–45 (2011).
Yasui, F. et al. Prior immunization with severe acute respiratory syndrome (SARS)-associated coronavirus (SARS-CoV) nucleocapsid protein causes severe pneumonia in mice infected with SARS-CoV. J. Immunol. 181, 6337–6348 (2008).
Zúñiga, S. et al. Coronavirus nucleocapsid protein facilitates template switching and is required for efficient transcription. J. Virol. 84, 2169–2175 (2010).
Mu, J. et al. SARS-CoV-2-encoded nucleocapsid protein acts as a viral suppressor of RNA interference in cells. Sci. China Life Sci. 63, 1413–1416 (2020).
Luo, H. et al. Severe acute respiratory syndrome coronavirus membrane protein interacts with nucleocapsid protein mostly through their carboxyl termini by electrostatic attraction. Int. J. Biochem. Cell Biol. 38, 589–599 (2006).
Lu, S. et al. The SARS-CoV-2 nucleocapsid phosphoprotein forms mutually exclusive condensates with RNA and the membrane-associated M protein. Nat. Commun. 12, 502 (2021).
Kuo, L., Koetzner, C. A. & Masters, P. S. A key role for the carboxy-terminal tail of the murine coronavirus nucleocapsid protein in coordination of genome packaging. Virology 494, 100–107 (2016).
Peng, Y. et al. Structures of the SARS-CoV-2 nucleocapsid and their perspectives for drug design. EMBO J. 39, e105938 (2020).
Zinzula, L. et al. High-resolution structure and biophysical characterization of the nucleocapsid phosphoprotein dimerization domain from the Covid-19 severe acute respiratory syndrome coronavirus 2. Biochem. Biophys. Res. Commun. 538, 54–62 (2021).
Ye, Q., West, A. M. V., Silletti, S. & Corbett, K. D. Architecture and self-assembly of the SARS-CoV-2 nucleocapsid protein. Protein Sci. 29, 1890–1901 (2020).
Zhou, R., Zeng, R., von Brunn, A. & Lei, J. Structural characterization of the C-terminal domain of SARS-CoV-2 nucleocapsid protein. Mol. Biomed. 1, 2 (2020).
Yang, M. et al. Structural insight into the SARS-CoV-2 nucleocapsid protein C-terminal domain reveals a novel recognition mechanism for viral transcriptional regulatory sequences. Front. Chem. 8, 624765 (2020).
Ye, Q., Lu, S. & Corbett, K. D. Structural basis for SARS-CoV-2 nucleocapsid protein recognition by single-domain antibodies. Front. Immunol. 12, 719037 (2021).
Chen, S.-C., Olsthoorn, R. C. L. & Yu, C.-H. Structural phylogenetic analysis reveals lineage-specific RNA repetitive structural motifs in all coronaviruses and associated variations in SARS-CoV-2. Virus Evol. 7, veab021 (2021).
Masters, P. S. Coronavirus genomic RNA packaging. Virology 537, 198–207 (2019).
Yu, I.-M., Oldham, M. L., Zhang, J. & Chen, J. Crystal structure of the severe acute respiratory syndrome (SARS) coronavirus nucleocapsid protein dimerization domain reveals evolutionary linkage between corona- and arteriviridae. J. Biol. Chem. 281, 17134–17139 (2006).
Nguyen, T. H. V. et al. Structure and oligomerization state of the C-terminal region of the Middle East respiratory syndrome coronavirus nucleoprotein. Acta Crystallogr. D. Struct. Biol. 75, 8–15 (2019).
Szelazek, B. et al. Structural characterization of human coronavirus NL63 N protein. J. Virol. 91, e02503-16 (2017).
Wienken, C. J., Baaske, P., Rothbauer, U., Braun, D. & Duhr, S. Protein-binding assays in biological liquids using microscale thermophoresis. Nat. Commun. 1, 100 (2010).
Powell, H. R., Johnson, O. & Leslie, A. G. W. Autoindexing diffraction images with iMosflm. Acta Crystallogr. D. Biol. Crystallogr. 69, 1195–1203 (2013).
Battye, T. G. G., Kontogiannis, L., Johnson, O., Powell, H. R. & Leslie, A. G. W. iMOSFLM: a new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallogr. D. Biol. Crystallogr. 67, 271–281 (2011).
Evans, P. R. & Murshudov, G. N. How good are my data and what is the resolution? Acta Crystallogr. D. Biol. Crystallogr. 69, 1204–1214 (2013).
Winn, M. D. et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D. Biol. Crystallogr. 67, 235–242 (2011).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Murshudov, G. N. et al. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D. Biol. Crystallogr. 67, 355–367 (2011).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D. Biol. Crystallogr. 66, 486–501 (2010).
Acknowledgements
We would like to thank to Dr. Encarnación Martínez-Salas from CBM-CSIC for her suggestions on the EMSA assays, and Dr. Santiago Ramón-Maiques and Dr. Francisco Del Caño-Ochoa from IBV-CSIC, for their technical support and critical reading of the manuscript and the staff of the synchrotron for technical assistance. The X-ray diffraction experiments were performed in XALOC beamline at ALBA Synchrotron (Barcelona; Spain). This work was supported by the COVID research grant COV20/01265 awarded to M.V. from the Instituto de Salud Carlos III (ISCIII; Spain) and by the European Commission–NextGenerationEU (Regulation EU 2020/2094), through CSIC’s Global Health Platform (PTI Salud Global). X-ray diffraction data collection was supported by the Spanish Synchrotron Radiation Facility ALBA through the COVID Proposal 2020074407 awarded to J.R.C.-T. and M.V.
Author information
Authors and Affiliations
Contributions
Conceptualization: J.R.C.-T. and M.V.; Methodology: J.R.C.-T., M.L.F., and M.V.; Investigation: J.R.C.-T. and M.V.; writing-original draft, J.R.C.-T. and M.V.; funding acquisition: M.V.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Stephanie Gras, Mukesh Kumar, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Isabelle Lucet and Gene Chong. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rafael Ciges-Tomas, J., Franco, M.L. & Vilar, M. Identification of a guanine-specific pocket in the protein N of SARS-CoV-2. Commun Biol 5, 711 (2022). https://doi.org/10.1038/s42003-022-03647-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-022-03647-8
- Springer Nature Limited
This article is cited by
-
Crykey: Rapid identification of SARS-CoV-2 cryptic mutations in wastewater
Nature Communications (2024)