
Abstract
Listeria monocytogenes is well known for having the ability to cross the placental barrier, leading to fetal infections and abortion. However, the mechanisms leading to infectious abortion are poorly understood. In this study, we demonstrate that interferon γ-induced GTPase (IGTP) contributes to the invasion of L. monocytogenes into trophoblast giant (TG) cells, which are placental immune cells. Knockdown of IGTP in TG cells decreased the relative efficiencies of L. monocytogenes invasion. Moreover, IGTP accumulated around infected L. monocytogenes in TG cells. Treatment of TG cells with phosphatidylinositol 3-kinase (PI3K)/Akt inhibitors also reduced bacterial invasion. PI3K/Akt inhibitor or IGTP knockdown reduced the amount of phosphorylated Akt. Monosialotetrahexosylganglioside (GM1) gangliosides, lipid raft markers, accumulated in the membrane of L. monocytogenes-containing vacuoles in TG cells. Furthermore, treatment with a lipid raft inhibitor reduced bacterial invasion. These results suggest that IGTP-induced activation of the PI3K/Akt signaling pathway promotes bacterial invasion into TG cells.
Similar content being viewed by others

Introduction
Listeria monocytogenes has the ability to cross the intestinal, blood-brain and placental barriers, leading to gastroenteritis, meningoencephalitis and maternofetal infections, respectively. Maternofetal infection results in abortion occasionally. A key feature of the virulence of L. monocytogenes is its ability to avoid the killing mechanisms of professional and non- professional phagocytic host cells1,2. L. monocytogenes infections in humans are caused mainly by the ingestion of contaminated food, such as dairy products, raw vegetables, fish, poultry, processed chicken and beef3.
Pregnancy leads to a generalized suppression of the adaptive immune system, typified by significantly decreased cell-mediated immunity and reduced T helper cell (Th) 1 responsiveness4,5. This immunosuppressed state prevents maternal rejection of the fetus but has the unfortunate consequence of increasing maternal susceptibility to certain infectious agents6,7. Immunity against L. monocytogenes is principally mediated by cellular immune responses because it is an intracellular pathogen8. For many other intracellular bacterial and protozoan pathogens, it has been shown that interferon-γ (IFN-γ) is an important component of Th1 immune responses and contributes to control through its ability to stimulate macrophages to kill more microbes. The infectious abortion model using a pregnant mouse is a powerful tool for investigating the mechanisms of bacterial pathogenesis. In our previous study, we demonstrated that abortion-inducing bacteria in human and animals, such as L. monocytogenes and Brucella abortus, cause abortion in pregnant mice model9,10. In Brucella infection, we found that there was a higher degree of bacterial colonization in the placenta than in other organs, that there were many bacteria in trophoblast giant (TG) cells in the placenta and that abortion was not induced in an intracellular replication-defective mutant11. In addition, we demonstrated that infection induced a transient increase in IFN-γ in pregnant mice. This transient IFN-γ production also contributes to infectious abortion and its neutralization serves to prevent abortion11. These studies of Brucella infection suggest that bacterial infection of TG cells plays a key role in causing abortion and that TG cells are closely linked to the avoidance of maternal immune rejection.
TG cells are polyploid cells differentiated from trophoblast stem (TS) cells by many morphological and functional developments; they form the fetal component of the placenta12. In particular, TG cells play crucial roles in implantation and the formation of a diffuse network of blood sinuses13 and promote maternal blood flow to the implantation site in mice14. TG cells are essential for the establishment of pregnancy. TG cells in the mouse placenta are parallel to extravillous cytotrophoblast cells in the human placenta14. Trophoblast cells also have a phagocytic ability. During implantation, trophoblast cells invade maternal tissue by phagocytosing uterine epithelial cells and stroma15. Several molecular mechanisms involved in phagocytosis by trophoblast cells have been reported16, however, the complete process remains unclear. It has also been reported that trophoblast cells can phagocytose pathogens and that this activity is enhanced by IFN-γ treatment17. Therefore, trophoblast cells may act in a manner similar to that of macrophages in phagocytosis. These studies suggested that trophoblast cells play a role not only in the development and maintenance of placenta but also in the placental defense system.
IFN-γ-induced GTPase (IGTP), also known as Irgm3, belongs to a family of 47 kDa IFN-γ-responsive GTPases (IRG). These family proteins are known to play critical roles in mediating specific resistance to intracellular pathogens including protozoa, bacteria and viruses18,19,20. Because IGTP localizes predominantly to the endoplasmic reticulum, it is assumed to be involved in the processing and trafficking of immunologically relevant proteins21,22. IGTP has been found to be essential for host resistance to acute infections by the protozoans Toxoplasma gondii23 and Leishmania major22, but its antimicrobial mechanism is still not clear.
In the present study, we investigated the role of IGTP in the invasion of L. monocytogenes into TG cells. Our results suggested that IGTP induces the activation of phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway and promotes bacterial invasion into TG cells.
Results
IGTP expression is induced by IFN-γ in TG cells
L. monocytogenes infects placental cells and induces cell death in vitro and in vivo24,25,26, which results in infectious abortion occasionally. In a previous study, we demonstrated that IFN-γ promotes bacterial invasion into TG cells27,28. We speculated that factors induced by IFN-γ should play a key role in the bacterial infection of TG cells. In this study, we focused on IGTP. First, to confirm the expression of IGTP protein and RNA in TG cells, we performed reverse transcription PCR (RT-PCR) and immunoblotting with or without IFN-γ treatment and compared the results. IFN-γ treatment induced IGTP expression at both the RNA (Fig. 1a) and protein (Fig. 1b) level.

Increased IGTP expression in TG cells treated with IFN-γ.
TG cells were treated with or without IFN-γ (3,000 units/ml) for 24 h (a and b); IGTP expression was analyzed by RT-PCR (a) and immunoblotting (b). β-actin was used as a control. The blots were scanned and analyzed using ImageJ software. For each sample, IGTP or β-actin expression observed in the absence of IFN-γ was set at 100% and IGTP or β-actin expression in the presence of IFN-γ was expressed relative to this value (lower panels).
IGTP promotes bacterial invasion into TG cells
To examine the effect of IGTP on bacterial invasion into TG cells, we used siRNA to knockdown IGTP in TG cells. Immunoblotting confirmed that IGTP expression levels were reduced in TG cells with or without IFN-γ treatment (Fig. 2a). The relative L. monocytogenes invasion efficiencies were significantly reduced by the knockdown of IGTP by siRNA in TG cells, with or without IFN-γ treatment (Fig. 2b). L. monocytogenes invasion did not affect IGTP expression (Supplementary Figure S1). Next we examined the localization of IGTP in TG cells 30 min after bacterial infection. IGTP accumulated around infected L. monocytogenes in TG cells (Fig. 3). The number of co-localized bacteria with IGTP per 100 intracellular bacteria were counted in 5 times independent experiments and 32.2 ± 10.38% of bacteria co-localized with IGTP. These results suggested that IGTP contributes to bacterial invasion into TG cells.

Effect of IGTP depletion on bacterial invasion into TG cells.
(a) TG cells were treated for 48 h with siRNA-targeting IGTP, with or without IFN-γ (3,000 units/ml) for 24 h (IGTP) simultaneously during cell differentiation. A negative control sample was treated with AllStars Negative Control siRNA (Neg.). Expression of the indicated proteins was monitored by immunoblotting. β-actin was used as a control. The blots were scanned and analyzed using ImageJ software. For each sample, IGTP or β-actin expression of negative control was set at 100% and IGTP or β-actin expression treated with siRNA-targeting IGTP was expressed relative to this value (lower panels). (b) Bacterial invasion into IGTP-depleted TG cells with or without IFN-γ was studied using a bacterial invasion assay. The efficiency of L. monocytogenes invasion into TG cells was increased 2.3 times by IFN-γ treatment. Data are the averages of triplicate samples from three identical experiments and the error bars represent standard deviations. Statistically significant differences compared with the negative control are indicated by asterisks (*, P < 0.05).

Localization of L. monocytogenes and IGTP in TG cells by immunofluorescence microscopy.
TG cells were infected with L. monocytogenes for 30 min, then fixed and stained for IGTP (green) and intracellular bacteria (red). Arrows indicate bacteria colocalized with IGTP. Scale bar indicates 10 μm.
IGTP induces the activation of PI3K/Akt in TG cells
IGTP overexpression can activate the PI3K/Akt signaling pathway in HeLa cells29. To examine the effect of PI3K/Akt on bacterial invasion into TG cells, TG cells were treated with wortmannin, a PI3K inhibitor and with Akt 1/2 kinase inhibitor. Bacterial invasion was reduced by treatment of TG cells with PI3K/Akt inhibitors, with or without IFN-γ treatment (Fig. 4a). In addition, IFN-γ treatment induced the phosphorylation of Akt (p-Akt) in TG cells. Phosphorylation of Akt was reduced by treatment of PI3K/Akt inhibitors in TG cells with or without IFN-γ treatment (Fig. 4b). To examine the effect of IGTP on the activation of Akt, we used siRNA to knockdown IGTP in TG cells. The amount of p-Akt was reduced by the knockdown of IGTP (Fig. 5), suggesting that IGTP contributes to Akt activation.

Effect of PI3K/Akt inhibitor on bacterial invasion and phosphorylation of Akt in TG cells.
(a) TG cells were treated with wortmannin (Wort.) or Akt 1/2 kinase inhibitor (Akt) for 1 h. Treated TG cells were infected with L. monocytogenes in the presence or absence of IFN-γ. Bacterial invasion into TG cells was measured using a bacterial invasion assay. Data represent the averages and standard deviations of triplicate samples from three identical experiments. Statistically significant differences between control and treated groups are indicated by asterisks (*, P < 0.05). (b) TG cells were treated for 1 h with wortmannin or Akt 1/2 kinase inhibitor with or without IFN-γ (3,000 units/ml), for 24 h. Expression of the indicated proteins was detected by immunoblotting. β-actin was used as a control. Cropped gels/blots are used in immunoblotting data. Full length images are presented in supplementary information (Supplementary Figure S2).

Correlation between IGTP expression and phosphorylation of Akt.
TG cells were treated for 48 h with either siRNA-targeting IGTP, with or without IFN-γ (3,000 units/ml), for 24 h (IGTP) simultaneously during cell differentiation. A negative control sample was treated with AllStars Negative Control siRNA (Neg.). Expression of the indicated proteins was detected by immunoblotting. β-actin was used as a control. Cropped gels/blots are used in immunoblotting data. Full length images are presented in supplementary information (Supplementary Figure S3).
Lipid rafts contribute to bacterial internalization into TG cells
Because lipid rafts mediate PI3K/Akt signaling30, we examined the localization of GM1 gangliosides, which are associated with lipid rafts, in L. monocytogenes-infected TG cells. GM1 gangliosides accumulated in the membrane of L. monocytogenes-containing vacuoles in TG cells (Fig. 6a). Furthermore, bacterial invasion was reduced by treatment of TG cells with lipid raft inhibitors, such as β-cyclodextrin, nystatin and chlorpromazine, with or without IFN-γ treatment (Fig. 6b). Because several intracellular pathogens infect into host cells using lipid raft-mediated macropinocytosis31,32, we assessed the effect of the macropinocytosis inhibitor, amiloride, on the invasion of L. monocytogenes into TG cells. Amiloride also inhibited bacterial invasion into TG cells (Fig. 7). These results suggested that lipid raft-mediated macropinocytosis contributes to the invasion of L. monocytogenes into TG cells.

Localization of L. monocytogenes and GM1 ganglioside in TG cells using immunofluorescence microscopy.
(a) TG cells were infected with L. monocytogenes for 30 min, then fixed and stained for GM1 ganglioside (red) and intracellular bacteria (green). Arrows indicate colocalized bacteria with GM1 ganglioside. Scale bar indicates 10 μm. (b) TG cells were treated with β-cyclodextrin (Cd, 5 μg/ml), Nystatin (Nys, 20 μg/ml), or chlorpromazine (Cp, 15 μg/ml) for 1 h. Treated TG cells were infected with L. monocytogenesin the presence or absence of IFN-γ (3,000 units/ml) for 24 h. Bacterial invasion into TG cells was measured using a bacterial invasion assay. Data represent the averages and standard deviations of triplicate samples from three identical experiments. Statistically significant differences between control and treated groups are indicated by asterisks (*, P < 0.05).

Macropinocytosis is associated with L. monocytogenes invasion into TG cells.
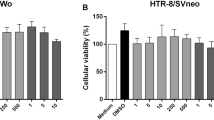
TG cells were treated with amiloride at the indicated concentrations for 1 h. Treated TG cells were then infected with L. monocytogenes. Bacterial invasion into TG cells was measured using a bacterial invasion assay. Data represent the averages and standard deviations of triplicate samples from three identical experiments. Statistically significant differences between control and treated groups are indicated by asterisks (*, P < 0.05).
Discussion
It is well known that L. monocytogenes infection induces IFN-γ production and that IFN-γ is the most important cytokine for host defense against L. monocytogenes because administration of recombinant IFN-γ enhances antilisterial resistance33,34 and administration of an anti-IFN-γ antibody eliminates this resistance35. Mice lacking IFN-γ or the IFN-γ receptor are highly susceptible to L. monocytogenes infection36,37. On the other hand, IFN-γ is known to be harmful for pregnancy because recent studies have shown that factors induced by bacterial infection, such as IFN-γ and “regulated on activation, normal T cell expressed and secreted” (RANTES), induce infectious abortion in Brucella infection38. However, the detailed mechanisms of infectious abortion remain unclear. It has also been reported that TG cells have a phagocytic activity and that this activity is enhanced by IFN-γ treatment27,28. Therefore, we thought that IFN-γ-inducible factors may be involved in the invasion of L. monocytogenes into TG cells.
IFN-γ induces resistance to bacterial infection through broad transcriptional programs involving a variety of genes, many of which remain uncharacterized39,40. IGTP is a member of the immunity-related GTPase family M (IRGM) subfamily of the family of IFN-γ-responsive GTPases (IRG). The IRG play a critical role in IFN-γ-induced resistance to intracellular pathogens. LRG-47 knockout mice are susceptible to a range of organism, including intracellular protozoa (L. major, Trypanosoma cruzi and T. gondii) and intracellular bacteria (L. monocytogenes, Salmonella typhimurium, Mycobacterium avium, M. tuberculosis and Chlamydia trachomatis)39,41. In contrast, IGTP knockout mice are susceptible to infection with T. gondii, L. major and C. trachomatis but are resistant to T. cruzi, L. monocytogenes, S. typhimurium, M. avium, M. tuberculosis, murine cytomegalovirus and Ebola virus39,41. In this study, we observed that the efficienciy of L. monocytogenes invasion was significantly reduced by siRNA mediated knockdown of IGTP in TG cells. We do not know if there is a direct relationship between the inhibition of bacterial invasion caused by knockdown of IGTP in TG cells and the resistance to bacterial infection exhibited by IGTP knockout mice. IGTP may contribute to the invasion of L. monocytogenes into other types of cells, such as macrophages and epithelial cells and that may be involved in resistance to bacterial infection.
IGTP localizes predominantly to the endoplasmic reticulum and has been found to be essential for host resistance to acute infection by T. gondii42. Elimination of T. gondii is dependent on IGTP and requires PI3K activity in macriphages43. IGTP can be seen clustered adjacent to T. gondii44. We also observed IGTP accumulated around infected L. monocytogenes in TG cells. Therefore, we suspected that bacterial invasion via IGTP may be mediated by the PI3K/Akt pathway. Indeed, we demonstrated that invasion of L. monocytogenes requires PI3K/Akt activity, suggesting that there are mediator molecules upstream of the PI3K/Akt pathway on the surface of TG cells. In our previous studies, several receptors expressed on TG cells contributed to the invasion of L. monocytogenes, such as Toll-like receptor (TLR) 2, class B scavenger receptor type 1 (SR-B1) and Mannose receptor, C type 1 (MRC1)27,28. Because the expression of these receptors was not induced by IFN-γ27,28, IFN-γ-mediated enhancement of bacterial invasion into TG cells may result from contributions by other cell surface receptors. On the other hand, TLR2 and SR-B1 are known to activate of PI3K45. Several receptors, including TLR2, SR-B1 and MRC1, may mediate the signals of bacterial invasion and activate the PI3K/Akt pathway in TG cells.
Signal transduction through cell surface receptors, including TLR2 and SR-B1, is involved in lipid rafts46,47. Lipid rafts are specialized areas of the plasma membrane which are enriched in cholesterol, sphingolipids, glycolipids and glycosylphosphatidylinositol-linked proteins48,49. Several intracellular pathogens invade into host cells through lipid rafts50,51,52. Invasion of L. monocytogenes into epithelial cells also involves lipid rafts53. We observed that lipid rafts and macropinocytosis contributed to bacterial invasion into TG cells in this study. These results suggested that putative cell surface receptors in lipid rafts mediate the signaling necessary for bacterial invasion into TG cells and activate the PI3K/Akt pathway and that this signaling is enhanced by IFN-γ.
The PI3K/Akt pathway is also known to be involved in the expression of heme oxygenease-1 (HO-1)54. HO-1 plays key roles in cytoprotection, anti-oxidation and anti-inflammation55,56. HO-1 expression in TG cells is decreased by L. monocytogenes infection and by treatment with IFN-γ. The reduction of HO-1 expression induces apoptosis in TG cells9. TG cell death inhibits the maintenance of pregnancy and results in infectious abortion. The modulation of the production of IFN-γ and IRG family members is an important host response for fetal survival and pathogen control. However, L. monocytogenes exploits this host defense system to achieve the survival strategy of the bacterium in the pregnant host. Recently, it has been reported that L. monocytogenes EGD strain has a point mutation in transcriptional regulator PrfA (PrfA*), leading to constitutive expression of several major virulence genes57. Since the mutation may affect invasion of L. monocytogenes into TG cells in vitro and in vivo, further analysis of bacterial virulence factors is also needed to clarify mechanism of bacterial invasion into TG cells.
Methods
Bacterial strains
L. monocytogenes EGD was maintained as a frozen glycerol stock and cultured in brain heart infusion (BHI) broth (Becton Dickinson, Franklin Lakes, NJ) or on BHI broth containing 1.5% agar.
Cell culture
The mouse TS cell line was a gift from Dr. Tanaka58,59. TS cells were cultured in a mixed medium comprising a 3:7 ratio of TS medium to mouse embryonic fibroblast-conditioned medium, supplemented with 25 ng/ml fibroblast growth factor 4 (TOYOBO, Osaka, Japan) and 1 μg/ml heparin (Sigma, St. Louis, MO), as described in our previous study59. TS medium was prepared by adding 20% fetal bovine serum (FBS), 1 mM sodium pyruvate, 100 μM β-mercaptoethanol and 2 mM l-glutamine to RPMI 1640 medium. To induce differentiation of TS cells to TG cells, TS cells were cultured in the TS medium alone for 3 days at 37°C in an atmosphere containing 5% CO2. TG cells were seeded (1–2 × 105 per well) in 48-well tissue culture plates. Size of TG cells is clearly larger than TS cells and the differentiation was usually confirmed by microscopy.
Efficiency of bacterial invasion into cultured cells
Overnight culture of L. monocytogenes strains were deposited onto TG cells at a multiplicity of infection (MOI) of 10 by centrifugation at 150 × g for 10 min at room temperature. To measure the efficiency of bacterial uptake, the infected cells were incubated at 37°C for 30 min, washed once with TS medium and then incubated in TS medium containing gentamicin (50 µg/ml) for 30 min. The cells were washed thrice with phosphate-buffered saline (PBS) and lysed with cold distilled water. Colony forming units (CFUs) were determined by serial dilution on BHI agar plates. Wortmannin (5 μM, Sigma), Akt 1/2 kinase inhibitor (20 μM, Sigma), nystatin (20 μg/ml, Wako, Osaka, Japan), β-cyclodextrin (5 μg/ml, Wako), chlorpromazine (15 μg/ml, Wako), or amiloride (indicated concentrations, Sigma) was added to the RPMI 1640 medium 1 h before infection. Recombinant IFN-γ (3,000 units/ml, Cedarlane Laboratories, Ontario, Canada) was added to the TS medium 24 h before infection.
Immunoblotting
TG cells were washed twice with PBS and lysed in lysis buffer [ice-cold Tris-buffered saline (TBS) containing 1% Triton X-100, 0.1% sodium dodecyl sulfate, 1% sodium deoxycholate, 10 mM EDTA and 1× Halt Protease Inhibitor Cocktail Kit (Thermo Fisher Science, Rockford, IL)] at 4°C for 60 min. The cell lysates were centrifuged (13,200 rpm, 4°C, 30 min) and supernatants were collected. Protein concentrations were determined using the Bio-Rad Protein Assay (Bio-Rad, Richmond, CA). Protein samples were separated on 10% polyacrylamide gels and transferred to a polyvinylidene difluoride membranes, which were incubated for overnight at 4°C with anti-mouse IGTP goat antibody (1:1000, Santa Cruz Biotechnology, Dallas, TX), anti-mouse Akt rabbit antibody (1:2000, Cell Signaling, Beverly, MA), or anti-mouse phospho-Akt rabbit antibody (1:2000, Cell Signaling) in 1% skim milk. The membranes were then washed thrice in TBS with 0.02% Tween 20, incubated for 30 min with a horseradish peroxidase (HRP)-conjugated secondary antibody (0.01 μg/ml) at room temperature and washed again. Immunoreactions were visualized using the enhanced chemiluminescence detection system (GE Healthcare Life Science, Little Chalfont, UK). The anti-mouse β-actin antibody was purchased from Sigma. Immunoblots were quantified using the ImageJ 1.32 software (National Institutes of Health, Bethesda, MD) after densitometric scanning of the films by LAS-3000 Imaging System (Fujifilm Life Science, Tokyo, Japan).
siRNA experiment
The siRNA duplexes used to silence mouse IGTP (target sequence:5′-TCCCATGGATTTAGTCACAAA-3′) and AllStars Negative Control siRNA which was used as a scrambled control were purchased from QIAGEN (Hilden, Germany). TG cells were transiently transfected using the Lipofectamine® RNAiMAX Transfection Reagent (Invitrogen, Carlsbad, CA) with or without siRNAs at a final concentration of 1 nM.
RNA isolation and reverse transcription (RT)-PCR
Total RNA was isolated from TG cells using the RNAeasy Plus Mini Kit (QIAGEN). Purified RNA samples were stored at −80°C prior to use. The RNA was quantified by absorption at 260 nm using a SmartSpec3000 spectrophotometer (Bio-Rad). RT-PCR was carried out using the SuperScript One-Step RT-PCR kit with Platinum Taq polymerase (Invitrogen). The primer sequences were as follows. IGTP: 5′-GCTGCTCCTGCCTCTTCTAA-3′ and 5′-ATTTAGACCACGGGCTGATG-3′ (in this study) and β-actin: 5′-TGGAATCCTGTGGCATCCATGAAAC-3′ and 5′-TAAAACGCAGCTCAGTAACAGTCCG-3′58.
Immunofluorescence microscopy
Bacteria were deposited onto TG cells grown on coverslips by centrifugation at 150 × g for 5 min at room temperature. The coverslips were then incubated at 37°C for 30 min. The samples were washed twice with PBS, fixed with 4% paraformaldehyde in PBS for 30 min at room temperature, washed thrice with PBS and then successively incubated thrice for 5 min in blocking buffer (5% bovine serum albumin in PBS) at room temperature. The samples were permeabilized with 0.2% Triton X-100 and washed thrice with PBS and then treated with 5 µg/ml anti-L. monocytogenes rabbit antibody (Viro Stat, Portland, ME) diluted in blocking buffer to identify intracellular bacteria or anti-mouse IGTP goat antibody to identify IGTP. After incubation for 1 h at 37°C, the samples were washed thrice for 5 min with blocking buffer, stained with TRITC-labeled goat anti-rabbit IgG (Santa Cruz Biotechnology) or FITC-labeled donkey anti-goat IgG (Santa Cruz Biotechnology) in blocking buffer and incubated for 1 h at 37°C. Fluorescent images were obtained using a FluoView FV100 confocal laser scanning microscope (Olympus, Tokyo, Japan). To detect the localization of GM1 gangliosides, TG cell monolayers were incubated for 5 min with biotin-cholera toxin B subunit (CTB) (10 μg/ml, List Biological Laboratories, Campbell, CA), rinsed thrice in RPMI 1640 medium and then incubated with L. monocytogenes for 30 min at 37°C. The cells were washed once, fixed in periodate-lysine-paraformaldehyde (PLP)-sucrose and probed for extracellular bacteria, as above, before permeablization in 0.2% Triton X-100 for 20 min at room temperature. After three washes with PBS and incubation in blocking buffer, the biotin-CTB was detected using Alexa Fluor 594-streptavidin (1:500, Molecular Probes, Eugene, OR)32.
Statistical analysis
Statistical analyses were performed using Student's t-test. Statistically significant differences, compared with control, are indicated by asterisks (*, P < 0.05). Data are the averages of triplicate samples from three identical experiments and the error bars represent standard deviations.
References
Cossart, P. Illuminating the landscape of host-pathogen interactions with the bacterium Listeria monocytogenes. Proc Natl Acad Sci USA 108, 19484–19491 (2011).
Portnoy, D. A., Chakraborty, T., Goebel, W. & Cossart, P. Molecular determinants of Listeria monocytogenes pathogenesis. Infect Immun 60, 1263–1267 (1992).
Lorber, B. Listeriosis. Clin Infect Dis 24, 1–9 (1997).
Weinberg, E. D. Pregnancy-associated immune suppression risks and mechanisms. Microb Pathog 3, 393–397 (1987).
Raghupathy, R. Th1-type immunity is incompatible with successful pregnancy. Immunol Today 18, 478–482 (1997).
Sano, M., Mitsuyama, M., Watanabe, Y. & Nomoto, K. Impairment of T cell-mediated immunity to Listeria monocytogenes in pregnant mice. Microbiol Immunol 30, 165–176 (1986).
Krishnan, L. et al. Pregnancy impairs resistance of C57BL/6 mice to Leishmania major infection and causes decreased antigen-specific IFN-gamma response and increased production of T helper 2 cytokines. J Immunol 156, 644–652 (1996).
Parmer, E. G. Immune responses to Listeria monocytogenes. Nat Rev Immunol 4, 812–823 (2004).
Tachibana, M., Watanabe, K., Yamasaki, Y., Suzuki, H. & Watarai, M. Expression of heme oxygenase-1 is associated with abortion caused by Brucella abortus infection in pregnant mice. Microb Pathog 45, 105–109 (2008).
Tachibana, M., Hashino, M., Nishida, T., Shimizu, T. & Watarai, M. Protective role of heme oxygenase-1 in Listeria monocytogenes-induced abortion. PLoS One 6, e25046 (2011).
Kim, S. et al. Interferon-γ promotes abortion due to Brucella infection in pregnant mice. BMC Microbiol 5, 22 (2005).
Parast, M. M., Aeder, S. & Sutherland, A. E. Trophoblast giant-cell differentiation involves changes in cytoskeleton and cell motility. Dev Biol 230, 43–60 (2001).
Bevilacqua, E. M. & Abrahamsohn, P. A. Ultrastructure of trophoblast giant cell transformation during the invasive stage of implantation of the mouse embryo. J Morphol 198, 341–351 (1988).
Cross, J. C. Genetic insights into trophoblast differentiation and placental morphogenesis. Semin Cell Dev Biol 11, 105–113 (2000).
Welsh, A. O. & Enders, A. C. Trophoblast-decidual cell interactions and establishment of maternal blood circulation in the parietal yolk sac placenta of the rat. Anat Rec 217, 203–219 (1987).
Drake, B. L. & Rodger, J. C. Phagocytic properties of cultured murine trophoblast. Placenta 8, 129–139 (1987).
Amarante-Paffaro, A., Queiroz, G. S., Correa, S. T., Spira, B. & Bevilacqua, E. Phagocytosis as a potential mechanism for microbial defense of mouse placental trophoblast cells. Reproduction 128, 207–218 (2004).
Taylor, G. A. et al. Pathogen-specific loss of host resistance in mice lacking the IFN-gamma-inducible gene IGTP. Proc Natl Acad Sci USA 97, 751–755 (2000).
Collazo, C. M. et al. The function of gamma interferon-inducible GTP-binding protein IGTP in host resistance to Toxoplasma gondii is Stat1 dependent and requires expression in both hematopoietic and nonhematopoietic cellular compartments. Infect Immun 70, 6933–6939 (2002).
Singh, S. B., Davis, A. S., Taylor, G. A. & Deretic, V. Human IRGM induces autophagy to eliminate intracellular mycobacteria. Science 313, 1438–1441 (2006).
Taylor, G. A. et al. The inducibly expressed GTPase localizes to the endoplasmic reticulum, independently of GTP binding. J Biol Chem 272, 10639–10645 (1997).
Taylor, G. A., Feng, C. G. & Sher, A. p47 GTPases: regulators of immunity to intracellular pathogens. Nat Rev Immunol 4, 100–109 (2004).
Halonen, S. K., Taylor, G. A. & Weiss, L. M. Gamma interferon-induced inhibition of Toxoplasma gondii in astrocytes is mediated by IGTP. Infect Immun 69, 5573–5576 (2001).
Abram, M. et al. Murine model of pregnancy-associated Listeria monocytogenes infection. FEMS Immunol Med Microbiol 35, 177–182 (2003).
Irvin, E. A., Williams, D., Voss, K. A. & Smith, M. A. Listeria monocytogenes infection in pregnant guinea pigs is associated with maternal liver necrosis, a decrease in maternal serum TNF-alpha concentrations and an increase in placental apoptosis. Reprod Toxicol 26, 123–129 (2008).
Le Monnier, A., Join-Lambert, O. F., Jaubert, F., Berche, P. & Kayal, S. Invasion of the placenta during murine listeriosis. Infect Immun 74, 663–672 (2006).
Watanabe, K., Shin, E. K., Hashino, M., Tachibana, M. & Watarai, M. Toll-like receptor 2 and class B scavenger receptor type I are required for bacterial uptake by trophoblast giant cells. Mol Immunol 47, 1989–1996 (2010).
Hashino, M., Tachibana, M., Shimizu, T. & Watarai, M. Mannose receptor, C type 1 contributes to bacterial uptake by placental trophoblast giant cells. FEMS Immunol Med Microbiol 66, 427–435 (2012).
Liu, Z. et al. Focal adhesion kinase mediates the interferon γ-inducible GTPase-induced phosphatidylinositol 3-kinase/Akt survival pathway and further initiates a positive feedback loop of NF-κB activation. Cell Microbiol 10, 1787–1800 (2008).
Arcaro, A. et al. Critical role for lipid raft-associated Src kinases in activation of PI3K-Akt signaling. Cell Signal 19, 1081–1092 (2007).
Hartlova, A., Cerveny, L., Hubalek, M., Krocova, Z. & Stulik, J. Membrane rafts: a potential gateway for bacterial entry into host cells. Microbial Immunol 54, 237–245 (2010).
Watarai, M., Makino, S., Fujii, Y., Okamoto, K. & Shirahata, T. Modulation of Brucella-induced macropinocytosis by lipid rafts mediates intracellular replication. Cell Microbiol 4, 341–355 (2002).
Chen, Y., Nakane, A. & Minagawa, T. Recombinant murine gamma interferon induces enhanced resistance to Listeria monocytogenes infection in neonatal mice. Infect Immun 57, 2345–2349 (1989).
Portnoy, D. A., Schreiber, R. D., Connelly, P. & Tilney, L. G. Gamma interferon limits access of Listeria monocytogenes to the macrophage cytoplasm. J Exp Med 170, 2141–2146 (1989).
Nakane, A., Numata, A. & Minagawa, T. Endogenous tumor necrosis factor, interleukin-6 and gamma interferon levels during Listeria monocytogenes infection in mice. Infect Immun 60, 523–528 (1992).
Huang, S. et al. Immune response in mice that lack the interferon-gamma receptor. Science 259, 1742–1745 (1993).
Dai, W. J. et al. Impaired macrophage listericidal and cytokine activities are responsible for the rapid death of Listeria monocytogenes-infected IFN gamma receptor-deficient mice. J Immunol 158, 5297–5304 (1997).
Watanabe, K. et al. Regulated upon activation normal T-cell expressed and secreted (RANTES) contributes to abortion caused by Brucella abortus infection in pregnant mice. J Vet Med Sci 70, 681–686 (2006).
Martens, S. & Howard, J. The interferon-inducible GTPases. Annu Rev Cell Dev Biol 22, 559–89 (2006).
Zhang, S. Y. et al. Inborn errors of interferon (IFN)-mediated immunity in humans: insights into the respective roles of IFN-alpha/beta, IFN-gamma and IFN-lambda in host defense. Immunol Rev 226, 29–40 (2008).
Hunn, J. P., Feng, C. G., Sher, A. & Howard, J. C. The immunity-related GTPases in mammals: a fast-evolving cell-autonomous resistance system against intracellular pathogens. Mamm Genome 22, 43–54 (2011).
Collazo, C. M. et al. The Function of gamma interferon-inducible GTP-binding protein IGTP in host resistance to Toxoplasma gondii is Stat1 dependent and requires expression in both hematopoietic and nonhematopoietic cellular compartments. Infect Immun 70, 6933–6939 (2002).
Ling, Y. M. et al. Vacuolar and plasma membrane stripping and autophagic elimination of Toxoplasma gondii in primed effector macrophages. J Exp Med 203, 2063–2071 (2006).
Melzer, T., Duffy, A., Weiss, L. M. & Halonen, S. K. The gamma interferon (IFN-gamma)-inducible GTP-binding protein IGTP is necessary for Toxoplasma vacuolar disruption and induces parasite egression in IFN-stimulated astrocytes. Infect Immun 76, 4883–4894 (2008).
Huang, H. L., Chiang, M. F., Lin, C. W. & Pu, H. F. Lipopolysaccharide directly stimulates aldosterone production via toll-like receptor 2 and toll-like receptor 4 related PI3K/Akt pathway in rat adrenal zona glomerulosa cells. J Cell Biochem 111, 872–880 (2010).
Soong, G., Reddy, B., Sokol, S., Adamo, R. & Prince, A. TLR2 is mobilized into an apical lipid raft receptor complex to signal infection in airway epithelial cells. J Clin Invest 113, 1482–1489 (2004).
Triantafilou, M. et al. Lipoteichoic acid and toll-like receptor 2 internalization and targeting to the Golgi are lipid raft-dependent. J Biol Chem 279, 40882–40889 (2004).
Simons, K. & Ikonen, E. Functional rafts in cell membranes. Nature 387, 569–572 (1997).
Lenne, P. F. et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J 25, 3245–3256 (2006).
Gatfield, J. & Pieters, J. Essential role for cholesterol in entry of mycobacteria into macrophages. Science 288, 1647–1650 (2000).
Shin, J. S., Gao, Z. & Abraham, S. N. Involvement of cellular caveolae in bacterial entry into mast cells. Science 289, 785–788 (2000).
Lafont, F., Tran Van Nhieu, G., Hanada, K., Sansonetti, P. & van der Goot, F. G. Initial steps of Shigella infection depend on the cholesterol/sphingolipid raft-mediated CD44-IpaB interaction. EMBO J 21, 4449–4457 (2002).
Seveau, S., Bierne, H., Giroux, S., Prévost, M. C. & Cossart, P. Role of lipid rafts in E-cadherin– and HGF-R/Met–mediated entry of Listeria monocytogenes into host cells. J Cell Biol 166, 743–753 (2004).
Mo, L. et al. PI3K/Akt signaling pathway-induced heme oxygenase-1 upregulation mediates the adaptive cytoprotection of hydrogen peroxide preconditioning against oxidative injury in PC12 cells. Int J Mol Med 30, 314–320 (2012).
Hegazi, R. et al. Carbon monoxide ameliorates chronic murine colitis through a heme oxygenase 1-dependent pathway. J Exp Med 202, 1703–1713 (2005).
Nakahira, K. et al. Carbon monoxide differentially inhibits TLR signaling pathways by regulating ROS-induced trafficking of TLRs to lipid rafts. J Exp Med 203, 2377–2389 (2006).
Becavin, C. et al. Comparison of widely used Listeria monocytogenes strains EGD, 10403S and EGD-e highlights genomic differences underlying variations in pathogenicity. mBio 5, e00969–14 (2014).
Tanaka, S., Kunath, T., Hadjantonakis, A. K., Nagy, A. & Rossant, J. Promotion of trophoblast stem cell proliferation by FGF4. Science 282, 2072–2075 (1998).
Watanabe, K. et al. Heat shock cognate protein 70 contributes to Brucella invasion into trophoblast giant cells that cause infectious abortion. BMC Microbiol 8, 212 (2008).
Acknowledgements
This work was partially supported by a Grant-in-Aid for Scientific Research (C) 22580333, MEXT TOKUBETSUKEIHI to M.W. from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) and by a KAKENHI Grant (Number 26850181) from the Japan Society for the Promotion of Science (JSPS). M.H. acknowledges the support from a JSPS Research Fellowship for Young Scientists (DC1).
Author information
Authors and Affiliations
Contributions
M.T. and M.W. planned the experiments and wrote the main manuscript. M.H., K.W. and T.S. discussed the experimental findings and interpretation of the results. M.T., M.H. and K.W. participated in the immunological and genetic experiments. M.T., T.S. and M.W. participated in the cell biology experiments.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Tachibana, M., Hashino, M., Watanabe, K. et al. Interferon γ-induced GTPase promotes invasion of Listeria monocytogenes into trophoblast giant cells. Sci Rep 5, 8195 (2015). https://doi.org/10.1038/srep08195
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep08195
- Springer Nature Limited