
Abstract
The associations of SNPs in TOX3 gene with breast cancer risk were investigated by some Genome-wide association studies and epidemiological studies, but the study results were contradictory. To derive a more precise estimate of the associations, we conducted a meta-analysis. ORs with 95% CI were used to assess the strength of association between TOX3 polymorphisms and breast cancer risk in fixed or random effect model. A total of 37 publications with 97275 cases and 128686 controls were identified. We observed that the rs3803662 C > T, rs12443621 A > G and rs8051542 C > T were all correlated with increased risk of breast cancer. In the stratified analyses by ethnicity, significantly elevated risk was detected for all genetic models of the three SNPs in Caucasians. In Asian populations, there were significant associations of rs3803662 and rs8051542 with breast cancer risk. Whereas there was no evidence for statistical significant association between the three SNPs and breast cancer risk in Africans. Additionally, we observed different associations of rs3803662 with breast cancer risk based on different ER subtype and BRCA1/BRCA2 mutation carriers. In conclusion, the meta-analysis suggested that three SNPs in TOX3 were significantly associated with breast cancer risk in different populations.
Similar content being viewed by others


Introduction
Breast cancer is the most generally diagnosed cancer and the most common cause of cancer death for females all over the world, particularly in the economically developing countries1. It is well known that breast cancer is a heterogeneous disease, not only in the aspect of various pathogenesis, but also in diversified clinical manifestation and outcome. Meanwhile, breast carcinoma is multifactorial disease, from a certain perspective, along with the combination of polygenic inheritance factor and environmental factor. Accompany with technological advances, more studies related with the genomic variation were conducted, in order to improve diagnosis and treatment for breast cancer patients. Mutations in some high and moderate penetrate genes, such as BRCA1, BRCA2 and ATM, were verified to be connected with the increased risk of breast cancer2,3. Nonetheless, these mutations constitute a part of the disease risk and it remains unclarified for the majority of genetic variations related with breast cancer susceptibility, particularly for low penetrate genes. It is noteworthy that genome-wide association studies (GWASs) about hundreds of single nucleotide polymorphisms (SNPs) provide strong evidences in elaborating the associations between low penetrate genes and breast cancer risk.
The TOX3 gene, formerly known as trinucleotide repeat containing 9 (TNRC9), is located in the chromosome 16q 12 and has a tri-nucleotide repeat motive. The gene encoded a protein containing a putative high mobility group (HMG) box4, indicating that it might play a potential role in calcium dependent transcription as a transcription factor5. In the recent years, the associations between genetic variants in TOX3 region and breast cancer susceptibility have been validated by GWASs and epidemiological studies in European, Asian and African American populations6,7,8,9,10,11,12,13. The SNP rs3803662 is located in 8 kb upstream of TOX3 and the rs12443621 and rs8051542 are both lied in an linkage disequilibrium (LD) block containing the 5′ end of TOX36.
The TOX3 rs3803662 was identified to exhibit association with breast cancer by GWASs6,7,10, with ascertainment of the association in Hispanic and non-Hispanic white women by Slattery et al.11. However, no significant association was found between rs3803662 and breast cancer risk in Asian and African ancestry8,13. Analogously, there was no evidence for the association between rs12443621 or rs8051542 and increased risk of breast cancer in Chinese women14,15,16. Whereas, Shan et al. reported that rs8051542 was significantly correlated with breast cancer risk in Tunisians17. Additionally, some studies found different relationships of three SNPs and breast cancer risk among different populations, which might result from different sample size or diverse allele frequencies and LD pattern among populations.
Meanwhile, the most recent meta-analysis related to the associations between the above-mentioned 3SNPs with breast cancer risk omitted some important studies18 and thus had limited statistical power to demonstrate the associations. Therefore, we performed an updated meta-analysis to aim to come up with the highest level of evidence for the associations between three SNPs in TOX3 gene and breast cancer risk among diverse ancestry populations and distinct tumor subtypes stratified by estrogen receptor (ER) or BRCA1/BRCA2.
Materials and Methods
Literature search strategy
We carried out a comprehensive literature search from PubMed and EMBASE databases up to March 2015, using the following search terms “TOX3” or “TNRC9” and “polymorphism” or “genetic variant” or “rs3803662” or “rs12443621” or “rs8051542” and “breast cancer” or “breast carcinoma” or “breast tumor” . First, we retrieved all potentially relevant articles, whose abstracts contained information related to our research purpose. Second, the references from eligible studies were carefully checked for additional relevant literature. Finally, only the comprehensive or the most recent study was brought into this meta-analysis, in the case that the same study population was included in several different articles.
Selection criteria
Eligible studies had to fulfill the following criteria: (1) case-control studies or cohort studies evaluating the association between TOX3 polymorphism (rs3803662, rs12443621 or rs8051542) and breast cancer risk; (2) odds ratio (OR) and 95% confidence interval (CI) or genotype data of rs3803662, rs12443621 or rs8051542 in breast cancer patients and cancer-free female to calculate OR and 95% CI; (3) studies were confined to human female groups; (4) articles in English.
Data extraction
A standard protocol was applied to extract data. For every eligible study, the following data were extracted: First author’s surname, year of publication, country of origin, population ethnicity, genotyping method, the genotype counts in cases and control (TT, CT and CC genotypes for TOX3 rs3803662; GG, AG and AA genotypes for rs12443621; TT, CT and CC genotypes for rs8051542) and P-value for the HWE in control groups. Two investigators independently extracted the above relative data with any disagreement resolved by discussion. If no consensus wasn’t reached, another investigator joined in the discussion. And the final decision was made by the majority of the votes.
Statistical methods
The strength of associations between TOX3 polymorphisms and breast carcinoma risk were estimated by OR with corresponding 95% CI. For all studies, we assessed the association under five different genetic models for calculating OR. Those were homozygote codominant model, heterozygote codominant model, dominant model, recessive model and allele model. Hardy-Weinberg equilibrium (HWE) was assessed by using χ2 test to compare expected and actual genotype frequencies among controls of each study. Q-statistic was applied to investigate heterogeneity among studies. P-value greater than 0.1 for Q test suggested a lack of statistically significant heterogeneity and the fixed-effect model (Mantel-Haenszel method)19 was used to calculate pooled ORs. Otherwise, heterogeneity was present and the random-effect model (DerSimonian-Laird method)20 was more appropriate. In addition, the I2-test was employed to accurately measure the degree of heterogeneity. Furthermore, the I2-value less than 25% was equivalent to mild heterogeneity and values between 25% and 50% was equivalent to moderate heterogeneity, whereas values greater than 50% was equivalent to large heterogeneity among studies. Potential publication bias was estimated by symmetry of funnel plot of OR versus the standard error of log (OR) and the visual symmetrical plot indicated that there was no publication bias among studies. Sensitivity analyses were conducted to assess the robustness of the results by eliminating each study in turn to show whether the individual data set influenced the pooled OR. Stratified analyses were conducted in terms of ethnicity, estrogen receptor (ER) status, BRCA1 and BRCA2 mutation. All statistical tests in this meta-analysis were two-tailed and P-value ≤ 0.05 was considered statistically significant unless otherwise noted. All statistical analyses were performed with Review Manager 5.2 software recommended by Cochrane Collaboration and Comprehensive Meta Analysis V2 software.
Result
Study Characteristics
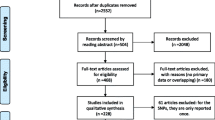
Based on the above selection criteria, a total of 37 eligible studies were included in the pooled analyses, involving 97275 cases and 128686 controls for rs3803662 polymorphism7,8,9,10,11,12,13,14,15,16,17,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46. For rs12443621, 14 studies8,12,14,15,16,17,24,28,29,31,41,42,45,46 involved a total of 17750 cases and 19488 controls. Moreover, there were 13 studies8,12,15,16,17,28,29,31,39,41,42,45,46 with 20965 cases and 21580 controls for rs8051542. Of particular note was that it’s smaller than 0.05 for the P-value of Hardy-Weinberg equilibrium in the controls of two studies, Campa et al. and Garcia-Closas et al.37,38, but we still included the two studies after sensitivity analyses were done. Additionally, in three included studies, genotype frequencies were shown separately according to different ethnic groups7,31,39. Therefore, the corresponding genotype counts in the study were separately considered for analyses. For rs3803662, five studies11,15,16,26,38 concerned with ER subtype of breast cancers and three studies26,30,35 related with BRCA1/2 mutation carriers were analysed as subgroups. The Fig. 1 expounded the study selection process. The Table 1 and 2 described the main features of these studies, especially for the genotype counts.

The flowchart of the study selection process.
Meta-analysis results
The mixtures of adjusted and crude estimates were used to calculate pooled ORs. The available adjusted variables of included studies were listed in supplementary table 1. Owing to large heterogeneity among studies, we used random-effect model to calculate pooled ORs for the associations of rs3803662 and rs12443621 with breast cancer risk. In contrast, fix-effect model was applied to calculate pooled ORs for rs8051542. In aggregate, T-rs3803662 and T-rs8051542 were all statistically associated with increased risk of breast cancer in all genetic models. However, the association between G-rs12443621 and breast cancer risk was only observed in Caucasians under all genetic models. The pooled ORs and 95%CI for these associations in all genetic models were shown in detail in Table 3, 4, 5, respectively. Forest plots related to the association of rs3803662, rs12443621 and rs8051542 with breast cancer susceptibility in homozygote model were shown in Fig. 2, Fig. 3 and Fig. 4, respectively.

Forest plot of TOX3 rs3803662 polymorphism and breast cancer risk.
Random-effect model was used for the analysis (homozygote codominant model TT vs. CC). The squares and horizontal lines correspond to the specific OR and 95% CI for every study. The area of the squares reflects the study specific weight. The diamond stands for the pooled OR and 95% CI.

Forest plot of TOX3 rs12443621 polymorphism and breast cancer risk.
Random-effect model was used for the analysis (homozygote codominant model GG vs. AA).

Forest plot of TOX3 rs8051542 polymorphism and breast cancer risk.
Fixed-effect model was used for the analysis (homozygote codominant model TT vs. CC).
In the subgroup analysis by ethnicity, our results indicated statistically significant associations between the three SNPs and breast cancer susceptibility in Caucasians under all genetic models. Nevertheless, as for Asian populations, T-rs3803662 and T-rs8051542 were shown to be statistically significant correlated with increased risk of breast cancer in all genetic models. In addition, there was no evidence for the statistical significant associations between the three SNPs and increased risk of breast cancer in African population which were almost African-American in our study. The pool ORs and 95%CI for these stratified analyses were detailedly shown in Table 3, 4, 5 for all genetic modes.
When stratified by ER status for rs3803662, statistically significant increased risk was found in ER+ and ER− tumor (Fig. 5 and Fig. 6). Moreover, a stronger association was identified in ER+ than ER− subtype for breast cancer risk (Fig. 7). Additionally, our analysis demonstrated that there were significant relationships between elevated risk of breast cancer and BRCA1/2 mutation carriers for rs3803662 (Fig. 8 and Fig. 9). And the details about ORs and 95% CI under all genetic models were shown in Table 3.

Forest plot of TOX3 rs3803662 polymorphism and breast cancer risk stratified by ER (+).
Fixed-effect model was used for the analysis (allele model T vs. C).

Forest plot of TOX3 rs3803662 polymorphism and breast cancer risk stratified by ER (−).
Fixed-effect model was used for the analysis (allele model T vs. C).

Forest plot of TOX3 rs3803662 polymorphism and breast cancer risk in ER+ subtype compared with ER− tumors.
Fixed-effect model was used for the analysis (allele model T vs. C).

Forest plot of TOX3 rs3803662 polymorphism and breast cancer risk stratified by BRCA1 mutation.
Fixed-effect model was used for the analysis (allele contrast model T vs. C).

Forest plot of TOX3 rs3803662 polymorphism and breast cancer risk stratified by BRCA2 mutation.
Fix -effect model was used for the analysis (allele contrast model T vs. C).
Sensitivity analyses and publication bias
Sensitivity analyses were conducted to assess the robustness of the results by eliminating each study in turn and all the results were not essentially altered, suggesting that the results of our meta-analysis were statistically stable. Publication bias of the eligible literature was evaluated by funnel plots and the shapes of funnel plots for literature about association between three SNPs and breast cancer risk were mostly symmetrical, indicating that no publication bias was detected.
Discussion
The TOX3 gene encoded a protein with an HMG box that is considered to be implicated in modification of DNA and chromatin structure47. Moreover, increased expression of TOX3 was relevant to bone metastasis in breast cancer patients48. Whereas, precise biological function of TOX3 is undetermined. Some GWASs and epidemiological studies have identified the associations of TOX3 polymorphisms with breast cancer susceptibility. However, study results were not consistent. Hence, in order to resolve the conflict, we performed this meta-analysis of the associations between the TOX3 rs3803662, rs12443621 and rs8051542 polymorphism and breast cancer risk.
The three SNPs locate in the 5’ end of TOX3 gene and a hypothetical gene LOC643714 on 16q12 and the region is contained in a 133kb linkage disequilibrium (LD) block12. Based on the International HapMap database, different LD patterns were observed between Asian and European ancestry. SNP rs3803662 was in moderate LD with rs12443621, with a Pearson’s correlation coefficient (r2) of 0.29 in the HapMap CEU population for European ancestry, but very weak LD was found between these two SNPs (r2 = 0.06) in Chinese8. Similarly, there was very weak association (r2 = 0.08) between rs3803662 and rs8051542 located 52 kb apart from each other in Chinese women45. However, the two SNPs showed moderate association (r2 = 0.15) with each other in European populations8. The substantial differences in genetic architecture among races, such as allele frequencies and LD structures, may partly account for our results which confirmed different association of the three SNPs with breast cancer risk in Caucasians, Asians and African-Americans. Rs3803662-T allele, rs12443621-G allele and rs8051542-T allele were statistically significantly associated with increased risk of breast cancer in Caucasians. Meanwhile, T-rs3803662 and T-rs8051542 were identified as risk factors of breast cancer in Asian populations. However, there was no evidence to prove that the three SNPs in African-Americans and G-rs12443621 in Asians were implicated in the breast tumor susceptibility, which was in line with the previous studies41,45. It’s worth mentioning that our study showed that T-rs3803662 and G-rs12443621 were protective factors in African-Americans in spite of no statistical significance.
In general, our study proved that the T-rs3803662 and T-rs8051542 in TOX3 were correlated with elevated breast cancer risk in all genetic models. It is notable that a previous meta-analysis directed by Chen et al.18 has showed that rs3803662 polymorphism was significantly associated with breast cancer risk, but no significant associations were observed for the rs12443621 and rs8051542. In addition, it only included eight case-control studies without stratified analyses. Compared with the previous meta-analysis, our study had more powerful and detailed analyses to prove our results. First and most obviously, more eligible literature and larger sample size were included. Second, the associations between breast cancer risk and rs3803662 polymorphism were considered with respect to ER status and BRCA1/2 mutation carriers. Third, stratified analyses were performed based on Caucasians, Asians and Africans, which was in favor of a more comprehensive understanding the associations in diverse populations. Finally and most importantly, we used mixture of adjusted and crude ORs rather than unadjusted estimates to calculate the pooled ORs. Meanwhile, the original genotype counts of eligible studies were also used to calculate the crude ORs. Supplementary table 2 showed the pooled ORs of the associations between the 3SNPs and breast cancer risk by using crude estimates. And there was no significant difference among the two results of pooled ORs based on different estimates, except for rs12443621. The crude ORs were incorporated to result in marginally association of rs12443621 with breast cancer risk under homozygote, dominant and allele genetic mode, but no association was found by using mixture ORs. That was probably because that adjusted estimates could yield more accurate results. Nevertheless, the two ways both demonstrated the relationship between rs12443621 and elevated risk of breast cancer in Caucasians.
To date, more attention has been paid to the heterogeneity of associations between common genetic variants and breast cancer subtypes. The two large-scale studies37,38 and our result identified that rs3803662 polymorphism was associated with both ER+ and ER− subtype of breast cancer, in spite of the slightly weaker association for ER− breast cancer. Additionally, our study demonstrated that T-rs3803662 was statistically significant associated with increased risk in ER+ breast cancer compared with ER− subtype, which was accordance with the researches done by Stacey et al. and Broeks et al.7,49. Intriguingly, T-rs8051542 allele and rs12443621 AG/GG genotypes, rather than rs3803662, were significantly associated with elevated risk of ER+ breast cancer in Chinese women8,16. By contrast, the significant associations of rs8051542 and rs12443621 were observed with luminal A (ER/PR+, Her2−) and Her2+/ER− breast cancer only among whites, respectively50. Furthermore, the association was strongly confirmed between rs3803662 and triple-negative tumors25,49. Taken together, these studies indicated that there were somehow connections between the three SNPs in TOX3 gene and pathological subtype of breast tumor among different populations. And in another aspect, these studies provided further support for the hypothesis that different subtypes stem from diverse etiological pathways. Additionally, rs3803662 SNP in BRCA1 and BRCA2 mutation carriers was significantly associated with the increased risk of breast cancer in our analysis, which was in consistent with previous studies26,35. While Latif et al. confirmed that the genetic variant was only associated with breast tumor in BRCA2 mutation carriers30. Therefore, it’s necessary to further elucidate the relevance of rs3803662 to breast cancer risk with BRAC1 and BRCA2 mutation.
Despite the advantage of large sample size and stratified analyses, the meta-analysis had several limitations that should be taken into account. First, there was extreme heterogeneity for the outcomes of the association between rs3803662 polymorphism and breast cancer risk. Although we reduced the degree of heterogeneity by stratified analyses based on ethnicity, other sources of heterogeneity were not verified, such as different genotyping methods or tumor types. Second, the sample size of African populations (5462 cases and 7155 controls) was relatively small. Therefore there was insufficient statistical power to demonstrate the associations between the 3SNPs and breast cancer risk in Africans. Third, the criterion of control groups was not uniformly defined. The design of eligible studies was based on population or/and hospital patients, thus there were potential risks of breast cancer in control groups. Fourth, the mixtures of crude and adjusted publish estimates, rather than incorporation of adjusted ORs, were used in the meta-analysis. Because of the lack of some individual data, we were unable to adjust effect size with possible confounders related with lifestyle risk factors, such as age, obesity, smoking, alcohol consumption and menopausal status. Furthermore, we were unable to examine the interaction between genetic variables and environment. In recent years, some studies for gene-environment interactions showed that relative risks of breast cancer correlated with low-penetrance susceptibility variants (including rs3803662) didn’t vary significantly with established environmental risk factors, such as reproductive history, menopausal status and body mass index51,52,53. Nevertheless, more and more researches have elaborated combined effect of low-penetrance susceptibility loci with breast cancer risk. And the obviously elevated risk stemming from combining many low-penetrant risk alleles supports the polygenic inheritance model of breast cancer44. Finally, owing to merely include English articles, there might be language bias on some level. Additionally, positive reports are tended to be published, which might make certain bias.
In conclusion, this meta-analysis indicated that there were different associations between the 3SNPs in TOX3 gene and breast cancer risk in different ethnic groups or subtype tumor. The 3SNPs were associated with the increased risk of breast cancer in Caucasians, while weren’t correlated in Africans. Additionally, rs3803662 and rs8051542 were risk factors for breast cancer in Asians. Furthermore, there were stronger associations between rs3803662 polymorphism and breast cancer risk in ER+ subtype than ER− tumors. Increased risk of breast cancer associated with rs3803662 was confirmed in BRCA1/BRCA2 mutation carriers. However, studies with larger sample size, which use uniform genotyping methods and criterion of control groups, have sufficiently corresponding individual data and consider the interactions of gene-gene and gene-environment will be needed to verify our results for TOX3 rs3803662, rs12443621 and rs8051542 as predisposition markers to breast cancer in clinical application.
Additional Information
How to cite this article: Zhang, L. and Long, X. Association of three SNPs in TOX3 and breast cancer risk: Evidence from 97275 cases and 128686 controls. Sci. Rep. 5, 12773; doi: 10.1038/srep12773 (2015).
References
Jemal, A. et al. Global cancer statistics. CA Cancer J Clin 61, 69–90 (2011).
Gabai-Kapara, E. et al. Population-based screening for breast and ovarian cancer risk due to BRCA1 and BRCA2. Proc Natl Acad Sci USA 111, 14205–14210 (2014).
Tavtigian, S. V. et al. Rare, evolutionarily unlikely missense substitutions in ATM confer increased risk of breast cancer. Am J Hum Genet 85, 427–446 (2009).
O’Flaherty, E. & Kaye, J. TOX defines a conserved subfamily of HMG-box proteins. BMC Genomics 4, 13–22 (2003).
Yuan, S. H., Qiu, Z. & Ghosh,. A. TOX3 regulates calcium-dependent transcription in neurons. Proc Natl Acad Sci USA 106, 2909–2914 (2009).
Easton, D. F. et al. Genome-wide association study identifies novel breast cancer susceptibility loci. Nature 447, 1087–1093 (2007).
Stacey, S. N. et al. Common variants on chromosomes 2q35 and 16q12 confer susceptibility to estrogen receptor-positive breast cancer. Nat Genet 39, 865–869 (2007).
He, X., Yao, G., Li, F., Li, M. & Yang, X. Risk-association of five SNPs in TOX3/LOC643714 with breast cancer in southern China. Int J Mol Sci 15, 2130–2141 (2014).
Elematore, I. et al. Association of genetic variants at TOX3, 2q35 and 8q24 with the risk of familial and early-onset breast cancer in a South-American population. Mol Biol Rep 41, 3715–3722 (2014).
Low, S. K. et al. Genome-wide association study of breast cancer in the Japanese population. PLoS One 8, e76463 (2013).
Slattery, M. L. et al. Replication of five GWAS-identified loci and breast cancer risk among Hispanic and non-Hispanic white women living in the Southwestern United States. Breast Cancer Res Treat 129, 531–539 (2011).
Udler, M. S. et al. Fine scale mapping of the breast cancer 16q12 locus. Hum Mol Genet 19, 2507–2515 (2010).
Ruiz-Narváez, E. A. et al. Polymorphisms in the TOX3/LOC643714 locus and risk of breast cancer in African-American women. Cancer Epidemiol Biomarkers Prev 19, 1320–1327 (2010).
Jiang, Y. et al. Risk of genome-wide association study newly identified genetic variants for breast cancer in Chinese women of Heilongjiang Province. Breast Cancer Res Treat 128, 251–257 (2011).
Li, L. et al. TNRC9/LOC643714 polymorphisms are not associated with breast cancer risk in Chinese women. Eur J Cancer Prev 18, 285–290 (2009).
Liang, J. et al. Genetic variants in trinucleotide repeat containing 9 (TNRC9) are associated with risk of estrogen receptor positive breast cancer in a Chinese population. Breast Cancer Res Treat 124, 237–241 (2010).
Shan, J. et al. Genome-Wide Association Studies (GWAS) breast cancer susceptibility loci in Arabs: susceptibility and prognostic in Tunisians. Breast Cancer Res Treat 135, 715–724 (2012).
Chen, M. B. et al. Association between polymorphisms of trinucleotide repeat containing 9 gene and breast cancer risk: evidence from 62,005 subjects. Breast Cancer Res Treat 126, 177–183 (2011).
Mantel, N. & Haenszel, W. Statistical aspects of the analysis of data from retrospective studies of disease. J Natl Cancer Inst 22, 719–748 (1959).
DerSimonian, R. & Laird, N. Meta-analysis in clinical trials. Control Clin Trials 7, 177–188 (1986).
Long, J. et al. Evaluating genome-wide association study-identified breast cancer risk variants in African-American women. PLoS One 8, e58350 (2013).
Kim, H. C. et al. A genome-wide association study identifies a breast cancer risk variant in ERBB4 at 2q34: results from the Seoul Breast Cancer Study. Breast Cancer Res 14, R56 (2012).
Huo, D. et al. Evaluation of 19 susceptibility loci of breast cancer in women of African ancestry. Carcinogenesis 33, 835–840 (2012).
Chan, M. et al. Association of common genetic variants with breast cancer risk and clinicopathological characteristics in a Chinese population. Breast Cancer Res Treat 136, 209–220 (2012).
Stevens, K. N. et al. Common breast cancer susceptibility loci are associated with triple-negative breast cancer. Cancer Res 71, 6240–6249 (2011).
Mulligan, A. M. et al. Common breast cancer susceptibility alleles are associated with tumour subtypes in BRCA1 and BRCA2 mutation carriers: results from the Consortium of Investigators of Modifiers of BRCA1/2. Breast Cancer Res 13, R110 (2011).
Han, W. et al. Common genetic variants associated with breast cancer in Korean women and differential susceptibility according to intrinsic subtype. Cancer Epidemiol Biomarkers Prev 20, 793–798 (2011).
Long, J. et al. Evaluation of breast cancer susceptibility loci in Chinese women. Cancer Epidemiol Biomarkers Prev 19, 2357–2365 (2010).
Long, J. et al. Identification of a functional genetic variant at 16q12.1 for breast cancer risk: results from the Asia Breast Cancer Consortium. PLoS Genet 6, e1001002 (2010).
Latif, A. et al. Breast cancer susceptibility variants alter risks in familial disease. J Med Genet 47, 126–131 (2010).
Barnholtz-Sloan, J. S. et al. FGFR2 and other loci identified in genome-wide association studies are associated with breast cancer in African-American and younger women. Carcinogenesis 31, 1417–1423 (2010).
Gorodnova, T. V. et al. Distribution of FGFR2, TNRC9, MAP3K1, LSP1 and 8q24 alleles in genetically enriched breast cancer patients versus elderly tumor-free women. Cancer Genet Cytoqenet 199, 69–72 (2010).
Hemminki, K. et al. Low-risk variants FGFR2, TNRC9 and LSP1 in German familial breast cancer patients. In t J Cancer 126, 2858–2862 (2010).
Mcinerney, N. et al. Low penetrance breast cancer predisposition SNPs are site specific. Breast Cancer Res Treat 117, 151–159 (2009).
Antoniou, A. C. et al. Common breast cancer-predisposition alleles are associated with breast cancer risk in BRCA1 and BRCA2 mutation carriers. Am J Hum Genet 82, 937–948 (2008).
Tapper, W. et al. The influence of genetic variation in 30 selected genes on the clinical characteristics of early onset breast cancer. Breast cancer Res 10, R108 (2008).
Campa, D. et al. Interactions between genetic variants and breast cancer risk factors in the breast and prostate cancer cohort consortium. J Natl Cancer Inst 103, 1252–1263 (2011).
Garcia-Closas, M. et al. Heterogeneity of breast cancer associations with five susceptibility loci by clinical and pathological characteristics. PLoS Genet 4, e1000054 (2008).
Barzan, D. et al. Comparison of genetic variation of breast cancer susceptibility genes in Chinese and German populations. Eur J Hum Genet 21, 1286–1292 (2013).
Rinella, E. S. et al. Genetic variants associated with breast cancer risk for Ashkenazi Jewish women with strong family histories but no identifiable BRCA1/2 mutation. Hum Genet 132, 523–536 (2013).
Zheng, W. et al. Evaluation of 11 breast cancer susceptibility loci in African-American women. Cancer Epidemiol Biomarkers Prev 18, 2761–2764 (2009).
Butt, S. et al. Genetic predisposition, parity, age at first childbirth and risk for breast cancer. BMC Res Notes 5, 414 (2012).
Mizoo, T. et al. Effects of lifestyle and single nucleotide polymorphisms on breast cancer risk: a case-control study in Japanese women. BMC Cancer 13, 565 (2013).
Harlid, S. et al. Combined effect of low-penetrant SNPs on breast cancer risk. Br J Cancer 106, 389–396 (2012).
Zheng, W. et al. Genetic and clinical predictors for breast cancer risk assessment and stratification among Chinese women. J Natl Cancer Inst 102, 972–981 (2010).
Tamimi, R. M. et al. Birth weight, breast cancer susceptibility loci and breast cancer risk. Cancer Causes Control 21, 689–696 (2010).
O’Flaherty, E. & Kaye, J. TOX defines a conserved subfamily of HMG-box proteins. BMC Genomics 4, 13 (2003).
Smid, M. et al. Genes associated with breast cancer metastatic to bone. J Clin Oncol 24. 2261–2267 (2006).
Broeks, A. et al. Low penetrance breast cancer susceptibility loci are associated with specific breast tumor subtypes: findings from the Breast Cancer Association Consortium. Hum Mol Genet 20, 3289–3303 (2011).
O’Brien, K. M. et al. Breast cancer subtypes and previously established genetic risk factors: a bayesian approach. Cancer Epidemiol Biomarkers Prev 23, 84–97 (2014).
Milne, R. L. et al. Assessing interactions between the associations of common genetic susceptibility variants, reproductive history and body mass index with breast cancer risk in the breast cancer association consortium: a combined case-control study. Breast Cancer Res 12, R110 (2010).
Travis, R. C. et al. Gene-environment interactions in 7610 women with breast cancer: prospective evidence from the Million Women Study. Lancet 375, 2143–2151 (2010).
Warren Andersen, S. et al. The associations between a polygenic score, reproductive and menstrual risk factors and breast cancer risk. Breast Cancer Res Treat 140, 427–434 (2013).
Acknowledgements
The Project-sponsored by SRF for ROCS, SEM. This work was supported by grants from National Natural Science Foundation of China (30873044 and 81272372).
Author information
Authors and Affiliations
Contributions
L.Z. and X.L. conceived and designed the experiments; L.Z. analyzed the data, L.Z. and X.L. wrote the manuscript. All authors reviewed and approved the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhang, L., Long, X. Association of three SNPs in TOX3 and breast cancer risk: Evidence from 97275 cases and 128686 controls. Sci Rep 5, 12773 (2015). https://doi.org/10.1038/srep12773
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep12773
- Springer Nature Limited
This article is cited by
-
Low-penetrance susceptibility variants and postmenopausal oestrogen receptor positive breast cancer
Journal of Genetics (2020)
-
An in silico approach to characterize nonsynonymous SNPs and regulatory SNPs in human TOX3 gene
Journal of Genetics (2019)
-
Significant association of TOX3/LOC643714 locus-rs3803662 and breast cancer risk in a cohort of Iranian population
Molecular Biology Reports (2019)