
Abstract
Single nucleotide polymorphisms (SNPs) of p53 rs1042522, MDM2 rs2279744 and p21 rs1801270, all in the p53 pathway, which plays a crucial role in DNA damage and genomic instability, were reported to be associated with cancer risk and pathologic characteristics. This case-control study was designed to analyse the association between these SNPs and retinoblastoma (RB) in a Chinese Han population. These SNPs in 168 RB patients and 185 adult controls were genotyped using genomic DNA from venous blood. No significant difference was observed in allele or genotypic frequencies of these SNPs between Chinese RB patients and controls (all P > 0.05). However, the rs1042522 GC genotype showed a protective effect against RB invasion, as demonstrated by event-free survival (HR = 0.53, P = 0.007 for GC versus GG/CC). This effect was significant for patients with a lag time >1 month and no pre-enucleation treatment (P = 0.007 and P = 0.010, respectively), indicating an interaction between p53 rs1042522 and clinical characteristics, including lag time and pre-enucleation treatment status. Thus, the rs1042522 SNP may be associated with RB invasion in the Han Chinese population; however, further large and functional studies are needed to assess the validity of this association.
Similar content being viewed by others


Introduction
Retinoblastoma (RB), a rare eye tumour observed in paediatric patients, is the most common intraocular malignancy among children worldwide1. Its global annual incidence is one case per 15,000–20,000 newborns—an estimated 9,000 new cases every year2. Both germinal/heritable (40%) and non-germinal/non-heritable RB (60%) originate in the primordial retinal cells and are predominantly initiated by biallelic inactivation of the RB1 gene1. The deletion of Rb family members leads to the compensatory up-regulation of the cyclin-dependent kinase inhibitor3 p21, which is the primary protein upregulated by p53 in response to DNA damage4. The p53 pathway, which is the master control system of the cell cycle, genome stability and cell apoptosis, is controlled by a negative feedback loop in which p53 transcriptionally activates MDM2, which in turn functions as a negative regulator by promoting the proteolytic degradation of p535. In addition to the loss of function of the RB1 gene, several other genes have been shown to be dysregulated in RB cells6,7,8 through single nucleotide polymorphisms (SNPs) in p53 pathway genes and may influence RB cancer risk among other ethnic populations9,10,11,12,13, indicating that SNPs in the p53 pathway genes and RB1 gene mutations may coincide in RB.
The Arg to Pro change in codon 72 (also known as p.Arg72Pro, rs1042522 G > C) is the most frequently studied functional SNP in p53. Compared with the G allele, the C allele of rs1042522 in p53 reduces proapoptotic activity and ultimately mediates the attenuation of the p53 pathway in concert with its induction of the MDM2 gene and other p53 targets14. An explanation of this diminished proapoptotic activity for the C allele is suggested to be a high affinity binding of the inhibitor of the apoptosis-stimulating protein of the p53 family15. Furthermore, the rs1042522 SNP modulates susceptibility to several human cancers, including hereditary RB in an Italian population10. In addition, the G allele has been associated with increased tumour stage16,17,18 and higher invasive or metastatic risk of cancer19,20. For the MDM2 gene, the G allele of a naturally occurring SNP at position 309 in the MDM2 promoter (also known as c.14 + 309T > G, rs2279744 T > G) has been shown to produce a higher-affinity DNA-binding site for Sp1, which leads to increased MDM2 mRNA and protein and thus increases the degradation of p53 and hinders p53-induced apoptosis21. Increased MDM2 expression is related to elevated cancer risk in sporadic and hereditary malignancies and to an increased likelihood of distant metastases22 and rs2279744 in the MDM2 gene is also associated with RB development9,10,11. Overall, these data indicate a risk-modifying effect of p53 and MDM2 SNPs on cancer23,24 and an association of these SNPs with pathological tumour characteristics25,26.
p21 is the primary protein that is upregulated by activated p53 in response to DNA damage4; conversely, p21 acts as a negative regulator of p53 stability27. Moreover, MDM2 negatively regulates p21 protein stability via proteasome-mediated degradation28. Most studies have focused on the p21 SNP in codon 31 (also known as p.Ser31Arg, rs1801270 C > A), which results in a non-synonymous serine-to-arginine substitution in the DNA-binding zinc finger motif of the protein29 and can lead to the failure of p21 to accumulate upon DNA damage, despite a normal p53 protein response30. Thus, this p21 SNP may affect the expression and activity of p21, thereby disturbing the p53 pathway activity and playing a role in the susceptibility to cancers31 including RB13.
Additionally, the p53 pathway targets pRB for degradation and controls the cell cycle and apoptosis in retinal cone precursor cells, from which the RB cell lineage originates32. Thus, the rs1042522, rs2279744 and rs1801270 SNPs in p53 pathway genes may affect the fate of RB. Previous studies have highlighted the contribution of the common functional SNPs rs1042522, rs2279744 and rs1801270 to RB cancer risk in non-Chinese populations9,10,11,12,13; however, to our knowledge, the association between these SNPs and RB invasion has not been reported. In the current study, for the first time, we genotyped these SNPs (rs1042522, rs2279744 and rs1801270) and evaluated their association with RB cancer risk and tumour invasion in a Chinese population.
Results
Study-population
This study was conducted on 168 RB patients and 185 cancer-free adult controls. The mean age of all patients at presentation was 20.0 ± 16.6 months, with a range from 1.0 to 118 months. No significant difference in gender was detected between the RB patients and the controls (P = 0.081). The clinical characteristics of the RB patients are summarised in Table 1. According to the International Classification of Retinoblastoma (ICRB), in the 168 patients, 131 eyes from 131 patients were enucleated, including 67 eyes (51.1%) in hazard group D and 64 eyes (48.9%) in hazard group E. Among enucleated patients, the percentage who received pre-enucleation treatment was higher in bilateral RB (78.9%) than in unilateral RB (5.4%) (P = 0.000) and a significantly negative correlation was observed between the age at presentation and lag time or pre-enucleation treatment (r = −0.21, P = 0.018; r = −0.25, P = 0.004, respectively). Histopathology reports were retrospectively staged for pTNM-staging characteristics and re-examination of the histopathological findings by a second reviewer (R.C.) showed 100% concurrence regarding the grade of RB invasion. Of the 131 enucleated eyes, 37 (28.2%), 49 (37.4%) and 45 (34.4%) enucleated eyes showed no, moderate and high invasion, respectively.
rs1042522, rs2279744 and rs1801270 SNPs and RB cancer risk
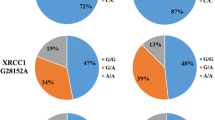
The p53 rs1042522, MDM2 rs2279744 and p21 rs1801270 SNPs were clearly distinguished by matrix-assisted laser desorption/ionisation time-of-flight (MALDI-TOF) mass spectrometry (MS) and the success rates of the genotype calls for these SNPs were 100%, 99.7% and 100%, respectively. Representative spectral peak diagrams are depicted in Fig. 1. In the control group, the genotype distributions of the evaluated SNPs were consistent with Hardy-Weinberg equilibrium (HWE) (all P > 0.05, Table 2). As shown in Table 2, neither the minor allele frequencies (MAFs) nor the genotype frequencies of the three selected SNPs were significantly different between RB and control groups (all P > 0.05).

Typical raw data obtained using the Sequenom MassARRAY system for single nucleotide polymorphisms of (a–c) p53 rs1042522 G/C, (d–f) MDM2 rs2279744 T/G and (g–i) p21 rs1801270 C/A. The blue arrows above the wave peak indicate the resulting genotypes.
rs1042522, rs2279744 and rs1801270 SNPs and RB invasion
Briefly, the distributions of the analysed genotypes were not significantly associated with any clinical characteristics of the patients (see Supplementary Table S1), except for the distribution of rs1042522 genotypes in laterality and pre-enucleation treatment status (P = 0.001 and P = 0.013, respectively) and of rs2279744 genotypes in lag time (P = 0.007); we adjusted for these factors in the Cox regression model analysis. For the patients with enucleated eyes, the analyses revealed almost identical associations of the selected SNPs with moderate and high invasion (see Supplementary Table S2). Therefore, the moderate and high invasion groups were combined into one group.
As shown in Fig. 2, a significant association was observed only for the rs1042522 SNP. Relative to the combined homozygous genotypes (GG and CC) in an over-dominant genetic model, the heterozygous GC genotype of p53 rs1042522 was significantly associated with event-free survival (EFS) for RB invasion (P = 0.000, Fig. 2a-b). For the genotypic association analysis of p21 rs1801270 and MDM2 rs2279744, no significant association with EFS was observed (Fig. 2c-d).

Kaplan-Meier analysis of the association of the selected SNPs of p53 pathway genes with event-free survival (EFS) for RB invasion among enucleated patients.
(a) Cumulative EFS curve of the enucleated patients for the three genotypes of p53 rs1042522 (P = 0.001 for GC versus GG, P = 0.914 for CC versus GG and P = 0.003 for GC versus CC); (b) Cumulative EFS curve of enucleated patients according to the over-dominant genetic model of p53 rs1042522 (P = 0.000 for GC versus GG/CC). (c) Cumulative EFS curve of enucleated patients according to the three genotypes of MDM2 rs2279744 (P = 0.204 for TG versus TT, P = 0.265 for GG versus TT and P = 0.940 for TG versus GG); (d) Cumulative EFS curve of enucleated patients according to the three genotypes of p21 rs1801270 (P = 0.524 for CA versus CC, P = 0.614 for AA versus CC and P = 0.938 for CA versus AA).
The results of the multivariate analysis of EFS using the Cox proportional hazards model are presented in Table 3. Only the carriers of the p53 rs1042522 GC genotype exhibited a significantly decreased risk of RB invasion (HR = 0.53; 95% CI = 0.33–0.84; P = 0.007) after adjustment for age (≤12 or >12 months), gender (male or female), laterality (unilateral or bilateral), clinical predictors (whether presented), ICRB (group D or E), lag time (≤1 or >1 month) and pre-enucleation treatment (administered or not). Furthermore, a lag time greater than 1 month and no pre-enucleation treatment were also associated with a reduced risk of RB invasion based on multivariate analysis (HR = 0.10, 95% CI = 0.05–0.24, P = 0.000 and HR = 0.02, 95% CI = 0.05–0.24, P = 0.000, respectively); a significant association was observed between age older than 12 months or bilateral eyes and the risk of RB invasion in the univariate analyses, whereas no association was observed in the multivariate analyses (HR = 1.13, 95% CI = 0.68–1.88, P = 0.635 and HR = 1.38, 95% CI = 0.623–3.11, P = 0.431, respectively). However, no significant association was detected between the genotypes of the other two SNPs and the risk of RB invasion.
Moreover, a subgroup analysis of the association of p53 rs1042522 with RB invasion risk was performed according to lag time and pre-enucleation treatment status. As shown in Table 4, in only the separate analyses of subgroups of lag time greater than 1 month and no prior treatment, the patients exhibited a significantly increased EFS duration (P = 0.008 and P = 0.004, respectively) and reduced invasion risk (HR = 0.31, P = 0.007 and HR = 0.50, P = 0.010, respectively) for the GC carriers relative to the homozygous carriers. However, no significant association between the MDM2 or p21 genotypes and RB invasion was observed according to the subgroup analyses of lag time or pre-enucleation treatment (see Supplementary Table S3).
Discussion
This study reports for the first time that the SNPs rs1042522, rs2279744 and rs1801270 in p53 pathway genes are not correlated with RB cancer risk, but rs1042522 is associated with RB invasion in a Chinese Han population. RB was classified according to the extent of tumour invasion to determine the association of RB invasion with these SNPs. The heterozygous GC genotype of p53 rs1042522 was negatively associated with RB invasion among patients with a lag time greater than 1 month and among those who had not undergone pre-enucleation treatments; however, no associations were observed between MDM2 rs2279744 or p21 rs1801270 and invasion.
Several studies have proposed a role for the p53 rs1042522 SNP as a risk factor for cancer due to its influence on cellular processes via p53-induced apoptosis33. Although the C allele frequency in Chinese RB (0.443) was relatively higher than that found in an Italian population (0.32) described in a previous report10, there was no association between the MAF of rs1042522 and RB cancer risk. This finding may be the result of a relatively high frequency of the C allele in the controls (0.438), which was similar to the C allele frequency of 0.489 in the Chinese population (HapMap-CHB database, http://hapmap.ncbi.nlm.nih.gov). Additionally, no association between RB cancer risk and the rs1042522 genotypes was found in this study. This finding was not in agreement with a previous study, which found an association between the p53 CC genotype and hereditary RB among 111 patients in an Italian population10. At present, there is no mechanism to support any proposed role for the rs1042522 SNP as a risk factor for RB and this lack of evidence may be due to conflicting results from different studies, which may also be attributed to the diverse distributions of SNPs in different races.
In addition, the mean age at presentation to our centre was 20.0 months, which was approximately the same as that in other Asian populations34,35,36. For the enucleated patients, those older than 12 months showed an increased risk of RB invasion based on univariate analysis (P = 0.001); this result was in accord with a previous study showing that older age is strongly associated with a high risk of histopathological findings37, whereas age was correlated with pre-enucleation treatment status and lag time in our study. Meanwhile, our data are consistent with a previous finding that a relatively higher percentage of RB patients initially presented with unilateral RB34,36. The enucleated patients with bilateral RB exhibited a reduced risk of RB invasion in the univariate analysis (P = 0.000), whereas the patients with bilateral RB tended to require pre-enucleation treatment in our study. An explanation of these findings is that chemotherapy before the enucleation of eyes containing RB diminishes the pathologic characteristics of extraocular extension38 and the distribution of p53 rs1042522 genotypes was correlated with RB laterality, pre-enucleation treatment status and lag time. For these reasons, the RB invasion risk was evaluated using multivariate analysis. After adjustment for clinical characteristics, the heterozygous genotype of p53 rs1042522 remained significantly associated with decreased RB invasion (P = 0.007).
Furthermore, subgroup analyses supported the association between the p53 rs1042522 GC genotype and RB invasion within the subgroup of patients with a lag time greater than 1 month (P = 0.007) or without prior treatment (P = 0.010). Therefore, an interaction between p53 rs1042522 genotypes and clinical characteristics (lag time and pre-enucleation treatment) contributed to RB invasion. This finding may be related to a decreased risk of future RB invasion because this result was observed in patients with a less invasive phenotype. These results may represent a “heterozygote advantage” situation in which the heterozygotes exhibit favourable survival characteristics39. Similar results have been observed in sickle cell anaemia40, HPV status in women from South Brazil,41 and breast cancer in an Iranian Azeri population42. Therefore, our findings may be useful in the near future for the promotion of prevention strategies guided by the patient’s genotype because the p53 rs1042522 GC genotype was associated with reduced RB invasion (over-dominant model). However, contrary to our finding, the GC genotype of rs1042522 showed a “heterozygote disadvantage” in which the heterozygote displayed an increased risk of metastasis in breast-invasive ductal carcinoma, based on a study conducted in northeast Brazil43. Although two wild-type alleles of rs1042522 have different potentials in binding components of the transcriptional machinery, activating transcription, inducing apoptosis and repressing cancer cells, recent studies demonstrate that rs1042522 could also be associated with cancer through an epigenetic mechanism, including RNAi suppression44 and DNA methylation45. Thus, the exact biological and genetic mechanisms underlying such associations are uncertain and further studies are necessary to confirm the association of the rs1042522 GC genotype with RB invasion.
Although the p53 gene is not mutated in primary RB46, the T-to-G variant of MDM2 has been shown to result in increased transcription, which plays a crucial role in the p53 pathway and in accelerating tumour formation21,47,48,49,50. The lack of an association of the MDM2 rs2279744 SNP contrasts with reports showing that the G allele appears to protect against RB development9,11. Moreover, our experiment did not show any association between the MDM2 rs2279744 SNP and RB invasion or the patients’ clinical characteristics. Thus, our findings appear to be plausible, considering a previous report showing that this association is only observed in p53-mutated cancers51.
Contrary to several molecular epidemiological studies that have reported the effects of the p21 rs1801270 SNP on cancer risk, no increased risk of developing RB was found in the studied Chinese population (P = 0.712). This finding contradicted the increased RB risk shown in Brazilian carriers of the CA genotype13. Although the p21 rs1801270 A allele results in a 38% reduction of p21 mRNA expression52, we found no association between the rs1801270 SNP and RB invasion based on both univariate and multivariate analyses (adjusted or not adjusted for the clinical characteristics of the patients) of the overall data. These results may partly account for the variation in the data regarding the MAF of rs1801270 in the population, ranging from 4% in a Swedish population to 50% in a Chinese population53. Compared to the frequency of the A allele (21%) in RB patients, which was reported by Carvalho et al., a relatively high A allele frequency (45.5%) was found in our study.
In conclusion, we have demonstrated that the p53 rs1042522 GC genotype, but not the MDM2 rs2279744 or p21 rs1801270 genotypes, is associated with a reduced risk of RB invasion in patients with a lag time greater than 1 month or in patients who received no prior treatment before enucleation, suggesting an interaction of rs1042522 with lag time and pre-enucleation treatment status in RB invasion. Considering how these SNPs interact with germline mutations in the RB1 gene, the present findings—if validated in other populations—could provide novel therapeutic implications.
Methods
Ethics statement
This study was approved by the medical ethics committee of Zhongshan Ophthalmic Center in Guangzhou (No. 2013PRLL019). Additionally, informed consent was obtained from all individuals after a thorough explanation of the procedure and its risk in accordance with the principles of the Declaration of Helsinki.
Study population
A total of 168 RB patients and 185 cancer-free controls were included in the present study. In brief, the patients were consecutively recruited from Zhongshan Ophthalmic Center of Sun Yat-sen University, Guangzhou, China, between February 2013 and October 2014. Unrelated controls of similar gender and ethnicity were recruited from healthy adults who visited the clinic for a general health check-up and had no individual history of cancer. Additionally, all subjects included in this study were Han Chinese. Subsequently, a peripheral blood sample was obtained from each recruited individual for genomic DNA extraction and the clinical information about the RB patients (age at presentation, gender, family history of RB, lag time between first symptom and treatment onset, laterality, clinical predictors at presentation, ICRB classification54, therapeutic regimen (including chemotherapy, focal treatment and enucleation), histopathology reports and duration between first symptom and enucleation onset) was collected from their medical records. The included clinical predictors were glaucoma, pseudohypopyon, hyphaema, staphyloma and neovascularisation of the iris37,55.
Assessment of RB invasion
The extent of tumour invasion into the anterior chamber, the iris, the ciliary body, the choroid, the sclera, the extra-scleral tissue, the optic nerve preceding or beyond the lamina and the optic nerve section margin were recorded based on written pathology reports and were confirmed via a retrospective re-review of haematoxylin and eosin-stained slides. For some analyses, the enucleation group was further subdivided into the following three groups based on the AJCC pTNM stage of RB invasion: no (pT1), moderate (pT2), or high (pT3, pT4) tumour invasion56 (Table 5).
Laboratory methods
Total DNA extraction
Total genomic DNA was extracted from 100 μL of EDTA-Na2 anticoagulated peripheral blood samples from each individual according to a previously described protocol57. The extracted DNA was eluted in 40 μL of TE buffer and stored at −20 °C in a dedicated area that was only used for PCR. The absorbance ratio of 260 nm to 280 nm was used to assess the purity of the isolated DNA using a Multiskan GO Microplate Spectrophotometer (Thermo Fisher Scientific, Inc., Waltham, MA, USA) and an absorbance ratio of ~1.8 was generally accepted as “pure”.
Genotyping. Based on the Sequenom Mass-ARRAY iPLEX platform, the SNP alleles were detected by allele-specific primer extension products using MALDI-TOF MS (Sequenom Inc., San Diego, California, USA). A multiplexed SNP assay was designed to test up to 3 SNPs using the “MySequenom” online design tools (https://www.mysequenom.com/Tools). According to the designated specifications, polymerase chain reaction (PCR) amplification primers and single-base extension (SBE) oligonucleotides for each SNP assay were synthesised and purified by Sangon Biotech Co., Ltd (Shanghai, China). PCR primers within the same multiplex were mixed at a final concentration of 1 μM each and the final concentrations of the SBE oligonucleotides were optimised according to the recommendations.
The genotyping reactions were performed on batches of 96-well plates according to the manufacturer’s instructions (http://www.sequenom.com). For PCR amplification, 10 ng of genomic DNA, dNTPs, primer mix and HotStart Taq polymerase (Qiagen Inc., Valencia, CA) were added to each well and the DNA was amplified via multiplex PCR. Unincorporated dNTPs present in the PCR product were neutralised using shrimp alkaline phosphatase (SAP), which was supplied with the iPLEX Gold reagent kit (Sequenom Inc.). After the SAP reaction, the SBE reactions were performed by adding extension primers, DNA polymerase and di-deoxynucleotide triphosphates (Sequenom Inc.) to each well. Subsequently, clean resin (Sequenom Inc.) was added to the mixture to remove extraneous salts that could interfere with MALDI-TOF analysis. The primer extension products were cleaned and aliquots of each sample were dispensed onto a 96-pad SpectroCHIP (Sequenom Inc.). Ultimately, genotype calling was performed in real time using MassARRAY RT software (version 3.0.0.4, Sequenom Inc.) and the results were analysed using MassARRAY Typer software (version 4.0, Sequenom Inc.). Duplicate samples and negative controls were included to verify the genotyping quality.
Statistical analysis
HWE of the genotype frequencies was tested using a goodness-of-fit Chi-square test with one degree of freedom among the controls. Comparisons of the SNPs between groups were analysed using Pearson’s Chi-square tests. Considering the major allele as the reference, logistic regression analysis was used to calculate the odds ratios (OR) and their corresponding 95% confidence intervals (CIs), which assessed the relative RB cancer risk conferred by each of the genotypes and alleles. In the univariate analysis, each genotype was independently analysed for a correlation with the duration of EFS for RB invasion. The Kaplan-Meier method was performed to estimate survival curves and the log-rank test was used to compare the EFS duration between the subgroups of RB patients. In the multivariate analysis, the association between the genetic models of the selected SNPs and RB invasion risk was estimated by computing the hazard ratio (HR) and the respective 95% CI using a Cox proportional hazards model, which was adjusted for potential risk factors that were significantly or marginally associated with invasion risk in the univariate analysis. If a P value was less than 0.05, the corresponding factor was included as a covariate in the Cox regression analysis. In the association analysis, all factors displaying P < 0.05 were subjected to subgroup analysis. All statistical tests were two-sided and were performed using SPSS version 20.0 for Mac OS (IBM Corporation, Armonk, New York, USA). The P value was set at a significance level of 0.05. For all multivariate analyses, the Bonferroni correction was applied.
Additional Information
How to cite this article: Chen, R. et al. Association of p53 rs1042522, MDM2 rs2279744 and p21 rs1801270 polymorphisms with retinoblastoma risk and invasion in a Chinese population. Sci. Rep. 5, 13300; doi: 10.1038/srep13300 (2015).
References
Dimaras, H. et al. Retinoblastoma. Lancet 379, 1436–1446 (2012). 10.1016/S0140-6736(11)61137-9.
Kivelä, T. The epidemiological challenge of the most frequent eye cancer: retinoblastoma, an issue of birth and death. Br. J. Ophthalmol. 93, 1129–1131 (2009). 10.1136/bjo.2008.150292.
Conkrite, K. et al. miR-17~92 cooperates with RB pathway mutations to promote retinoblastoma. Genes Dev. 25, 1734–1745 (2011). 10.1101/gad.17027411.
Gartel, A. L. & Radhakrishnan, S. K. Lost in transcription: p21 repression, mechanisms and consequences. Cancer Res. 65, 3980–3985 (2005). 10.1158/0008-5472.CAN-04-3995.
Perry, M. E. The regulation of the p53-mediated stress response by MDM2 and MDM4. Cold Spring Harb Perspect. Biol. 2, a000968 (2010). 10.1101/cshperspect.a000968.
Corson, T. W. & Gallie, B. L. One hit, two hits, three hits, more? Genomic changes in the development of retinoblastoma. Genes Chromosomes Cancer 46, 617–634 (2007). 10.1002/gcc.20457.
Chakraborty, S. et al. Identification of genes associated with tumorigenesis of retinoblastoma by microarray analysis. Genomics 90, 344–353 (2007). 10.1016/j.ygeno.2007.05.002.
Reis, A. H., Vargas, F. R. & Lemos, B. More epigenetic hits than meets the eye: microRNAs and genes associated with the tumorigenesis of retinoblastoma. Front. Genet. 3, 284 (2012). 10.3389/fgene.2012.00284.
Castéra, L. et al. MDM2 as a modifier gene in retinoblastoma. J. Natl Cancer Inst. 102, 1805–1808 (2010). 10.1093/jnci/djq416.
Epistolato, M. C. et al. p53 Arg72Pro and MDM2 309 SNPs in hereditary retinoblastoma. J. Hum. Genet. 56, 685–686 (2011). 10.1038/jhg.2011.82.
de Oliveira Reis, A. H. et al. Influence of MDM2 and MDM4 on development and survival in hereditary retinoblastoma. Pediatr. Blood Cancer 59, 39–43 (2012). 10.1002/pbc.24014.
McEvoy, J. et al. Analysis of MDM2 and MDM4 single nucleotide polymorphisms, mRNA splicing and protein expression in retinoblastoma. PLOS ONE 7, e42739 (2012). 10.1371/journal.pone.0042739.
Carvalho, I. N., Reis, A. H., Cabello, P. H. & Vargas, F. R. Polymorphisms of CDKN1A gene and risk of retinoblastoma. Carcinog. 34, 2774–2777 (2013). 10.1093/carcin/bgt308.
Dumont, P., Leu, J. I., Della Pietra, A. C., George, D. L. & Murphy, M. The codon 72 polymorphic variants of p53 have markedly different apoptotic potential. Nat. Genet. 33, 357–365 (2003). 10.1038/ng1093.
Sullivan, A. & Lu, X. ASPP: a new family of oncogenes and tumour suppressor genes. Br. J. Cancer 96, 196–200 (2007). 10.1038/sj.bjc.6603525.
Furihata, M. et al. p53 mutation arising in Arg72 allele in the tumorigenesis and development of carcinoma of the urinary tract. Clin. Cancer Res. 8, 1192–1195 (2002).
Schneider-Stock, R. et al. Retention of the arginine allele in codon 72 of the p53 gene correlates with poor apoptosis in head and neck cancer. Am. J. Pathol. 164, 1233–1241 (2004). 10.1016/S0002-9440(10)63211-7.
Pérez, L. O., Abba, M. C., Dulout, F. N. & Golijow, C. D. Evaluation of p53 codon 72 polymorphism in adenocarcinomas of the colon and rectum in La Plata, Argentina. World J. Gastroenterol. 12, 1426–1429 (2006).
Zhu, Z. Z. et al. Association of p53 codon 72 polymorphism with liver metastases of colorectal cancers positive for p53 overexpression. J. Zhejiang Univ. Sci. B 9, 847–852 (2008). 10.1631/jzus.B0820100.
Chen, W. C. et al. Distributions of p53 codon 72 polymorphism in bladder cancer--proline form is prominent in invasive tumor. Urol. Res. 28, 293–296 (2000). 10.1007/s002400000117.
Bond, G. L. et al. A single nucleotide polymorphism in the MDM2 promoter attenuates the p53 tumor suppressor pathway and accelerates tumor formation in humans. Cell 119, 591–602 (2004). 10.1016/j.cell.2004.11.022.
Rayburn, E., Zhang, R., He, J. & Wang, H. MDM2 and human malignancies: expression, clinical pathology, prognostic markers and implications for chemotherapy. Curr. Cancer Drug Targets. 5, 27–41 (2005). 10.2174/1568009053332636.
Sun, Y. F., Leu, J. D., Chen, S. M., Lin, I. F. & Lee, Y. J. Results based on 124 cases of breast cancer and 97 controls from Taiwan suggest that the single nucleotide polymorphism (SNP309) in the MDM2 gene promoter is associated with earlier onset and increased risk of breast cancer. BMC Cancer 9, 13 (2009). 10.1186/1471-2407-9-13.
Li, G. et al. MDM2 gene promoter polymorphisms and risk of lung cancer: a case-control analysis. Carcinogenesis 27, 2028–2033 (2006). 10.1093/carcin/bgl047.
Heist, R. S. et al. MDM2 polymorphism, survival and histology in early-stage non-small-cell lung cancer. J. Clin. Oncol. 25, 2243–2247 (2007). 10.1200/JCO.2006.08.8914.
Paulin, F. E. et al. MDM2 SNP309 is associated with high grade node positive breast tumours and is in linkage disequilibrium with a novel MDM2 intron 1 polymorphism. BMC Cancer 8, 281 (2008). 10.1186/1471-2407-8-281.
Broude, E. V. et al. p21 (CDKN1A) is a negative regulator of p53 stability. Cell Cycle 6, 1468–1471 (2007). 10.4161/cc.6.12.4313.
Zhang, Z. et al. MDM2 is a negative regulator of p21WAF1/CIP1, independent of p53. J. Biol. Chem. 279, 16000–16006 (2004). 10.1074/jbc.M312264200.
Mousses, S. et al. Two variants of the CIP1/WAF1 gene occur together and are associated with human cancer. Hum. Mol. Genet. 4, 1089–1092 (1995). 10.1093/hmg/4.6.1089.
Johnson, G. G. et al. A novel type of p53 pathway dysfunction in chronic lymphocytic leukemia resulting from two interacting single nucleotide polymorphisms within the p21 gene. Cancer Res. 69, 5210–5217 (2009). 10.1158/0008-5472.CAN-09-0627.
Liu, F. et al. P21 codon 31 polymorphism associated with cancer among white people: evidence from a meta-analysis involving 78,074 subjects. Mutagenesis 26, 513–521 (2011). 10.1093/mutage/ger010.
Xu, X. L. et al. Retinoblastoma has properties of a cone precursor tumor and depends upon cone-specific MDM2 signaling. Cell 137, 1018–1031 (2009). 10.1016/j.cell.2009.03.051.
Whibley, C., Pharoah, P. D. & Hollstein, M. p53 polymorphisms: cancer implications. Nat. Rev. Cancer 9, 95–107 (2009). 10.1038/nrc2584.
Chung, S. E. et al. Clinical manifestations and treatment of retinoblastoma in Korea. Br. J. Ophthalmol. 92, 1180–1184 (2008). 10.1136/bjo.2008.140046.
Chang, C. Y. et al. Retinoblastoma in Taiwan: survival rate and prognostic factors. Jpn. J. Ophthalmol. 50, 242–249 (2006). 10.1007/s10384-005-0320-y.
Zhao, J., Li, S., Shi, J. & Wang, N. Clinical presentation and group classification of newly diagnosed intraocular retinoblastoma in China. Br. J. Ophthalmol. 95, 1372–1375 (2011). 10.1136/bjo.2010.191130.
Kashyap, S. et al. Clinical predictors of high risk histopathology in retinoblastoma. Pediatr. Blood Cancer 58, 356–361 (2012). 10.1002/pbc.23239.
Zhao, J. et al. Pre-enucleation chemotherapy for eyes severely affected by retinoblastoma masks risk of tumor extension and increases death from metastasis. J. Clin. Oncol. 29, 845–851 (2011). 10.1200/JCO.2010.32.5332.
Savas, S., Liu, G. & Xu, W. Special considerations in prognostic research in cancer involving genetic polymorphisms. BMC Med. 11, 149 (2013). 10.1186/1741-7015-11-149.
Dean, M., Carrington, M. & O’Brien, S. J. Balanced polymorphism selected by genetic versus infectious human disease. Annu. Rev. Genomics Hum. Genet. 3, 263–292 (2002). 10.1146/annurev.genom.3.022502.103149.
Hartwig, F. P. et al. Evidence for an epistatic effect between TP53 R72P and MDM2 T309G SNPs in HIV infection: a cross-sectional study in women from South Brazil. PLOS ONE 9, e89489 (2014). 10.1371/journal.pone.0089489.
Saadatian, Z. et al. Association of rs1219648 in FGFR2 and rs1042522 in TP53 with premenopausal breast cancer in an Iranian Azeri population. Asian Pac. J. Cancer Prev. 15, 7955–7958 (2014). 10.7314/APJCP.2014.15.18.7955.
Ramalho, E. A. et al. Assessment of changes in the BRCA2 and P53 genes in breast invasive ductal carcinoma in northeast Brazil. Biol. Res. 47, 3 (2014). 10.1186/0717-6287-47-3.
Weige, C. C. et al. Transcriptomes and shRNA suppressors in a TP53 allele-specific model of early-onset colon cancer in African Americans. Mol. Cancer Res. 12, 1029–1041 (2014). 10.1158/1541-7786.MCR-13-0286-T.
Rebbani, K. et al. TP53 R72P polymorphism modulates DNA methylation in hepatocellular carcinoma. Mol. Cancer 14, 74 (2015). 10.1186/s12943-015-0340-2.
Kato, M. V. et al. Loss of heterozygosity on chromosome 17 and mutation of the p53 gene in retinoblastoma. Cancer Lett. 106, 75–82 (1996). 10.1016/0304-3835(96)04305-4.
Polsky, D. et al. HDM2 protein overexpression and prognosis in primary malignant melanoma. J. Natl Cancer Inst. 94, 1803–1806 (2002). 10.1093/jnci/94.23.1803.
Giordana, M. T. et al. MDM2 overexpression is associated with short survival in adults with medulloblastoma. Neuro-oncology. 4, 115–122 (2002).
Haitel, A., Wiener, H. G., Baethge, U., Marberger, M. & Susani, M. mdm2 expression as a prognostic indicator in clear cell renal cell carcinoma: comparison with p53 overexpression and clinicopathological parameters. Clin. Cancer Res. 6, 1840–1844 (2000).
Ranuncolo, S. M. et al. Prognostic value of Mdm2, p53 and p16 in patients with astrocytomas. J. Neurooncol. 68, 113–121 (2004). 10.1023/B:NEON.0000027741.19213.99.
Hu, Z. et al. MDM2 promoter polymorphism SNP309 contributes to tumor susceptibility: evidence from 21 case-control studies. Cancer Epidemiol. Biomarkers Prev. 16, 2717–2723 (2007). 10.1158/1055-9965.EPI-07-0634.
Su, L. et al. P53 (codon 72) and P21 (codon 31) polymorphisms alter in vivo mRNA expression of p21. Lung Cancer 40, 259–266 (2003). 10.1016/S0169-5002(03)00081-3.
Birgander, R. et al. The codon 31 polymorphism of the p53-inducible gene p21 shows distinct differences between major ethnic groups. Hum. Hered. 46, 148–154 (1996). 10.1159/000154344.
Kiss, S., Leiderman, Y. I. & Mukai, S. Diagnosis, classification and treatment of retinoblastoma. Int. Ophthalmol. Clin. 48, 135–147 (2008). 10.1097/IIO.0b013e3181693670.
Chawla, B. et al. Correlation between clinical features, magnetic resonance imaging and histopathologic findings in retinoblastoma: a prospective study. Ophthalmology 119, 850–856 (2012). 10.1016/j.ophtha.2011.09.037.
Finger, P. T. et al. Retinoblastoma in AJCC Cancer Staging Manual (ed. Edge, S. et al. ) 561–568 (Springer Science and Business Media, 2010).
Loparev, V., Cartas, M., Monken, C., Velpandi, A. & Srinivasan, A. An efficient and simple method of DNA extraction from whole blood and cell lines to identify infectious agents. J. Virol. Methods 34(1), 105–112 (1991). 10.1016/0166-0934(91)90126-K.
Acknowledgements
This study was partially supported by the National Natural Science Foundation of China (Grant No. 81470664). The funding organisation played no role in the design or performance of this study.
Author information
Authors and Affiliations
Contributions
H.S.Y. conceived and designed the experiments. R.C., S.L., H.J.Y., J.L., S.A. and Y.M. designed the experiments. R.C., S.L., L.C., X.L., Y.D. and Y.G.D. performed the experiments. W.M., X.L. and J.L. contributed reagents/materials/analysis tools. P.Z. reported and confirmed the RB histopathology. R.C., S.L. and Q.L. analysed the data. R.C. and S.L. wrote the manuscript. H.S.Y. supervised the research.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Chen, R., Liu, S., Ye, H. et al. Association of p53 rs1042522, MDM2 rs2279744 and p21 rs1801270 polymorphisms with retinoblastoma risk and invasion in a Chinese population. Sci Rep 5, 13300 (2015). https://doi.org/10.1038/srep13300
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep13300
- Springer Nature Limited