
Abstract
Recent studies have demonstrated that adiponectin (APN) attenuates cerebral ischemic/reperfusion via globular adiponectin (gAD). However, the therapeutic role of gAD in cerebral ischemic injury in type 1 diabetes mellitus (T1DM) remains unclear. Our results showed that gAD improved neurological scores and reduced the infarct volumes in the 8-week T1DM (T1DM-8W) mice, but not in the 2-week T1DM (T1DM-2W) mice. Moreover, the ischemic penumbra APN levels increased and peaked in T1DM-2W mice and reduced to normal in T1DM-8W mice, while the APN receptor 1 (AdipoR1) expression change was the opposite. Administration of rosiglitazone in T1DM-2W mice up-regulated the expression of AdipoR1 and restored the neuroprotection of gAD, while intracerebroventricular injection of AdipoR1 small interfering RNA (siRNA) in T1DM-8W mice reversed it. Furthermore, the expression of p-PERK, p-IRE1 and GRP78 were increased whereas the expressions of CHOP and cleaved caspase-12 as well as the number of apoptotic neurons were decreased after gAD treatment in T1DM-8W mice. These beneficial effects of gAD were reversed by pretreatment with AdipoR1 siRNA. These results demonstrated a dynamic dysfunction of APN/AdipoR1 accompanying T1DM progression. Interventions bolstering AdipoR1 expression during early stages and gAD supplementation during advanced stages may potentially reduce the cerebral ischemic injury in diabetic patients.
Similar content being viewed by others

Introduction
Diabetes mellitus (DM) is a complex metabolic disease and a major public health concern in China, with a prevalence rate of 9.7%1. Studies have found that diabetic patients have double the risk of suffering from stroke and 1-fold excess risk of stroke recurrence as compared to the normal population2. Moreover, the mortality and morbidity of stroke also increases in DM3. Therefore, the reduction of cerebral ischemic injury in DM is a major concern for scientists.
Adiponectin (APN) is regarded as an adipocyte-specific adipocytokine. Primary sequence analysis has revealed that full-length APN has four main domains, with the globular segment (gAD) at the carboxy terminus being much more potent than the full protein4. APN has multiple functions not only in the peripheral tissues, but also in the central nervous system. A previous study has confirmed that APN enhances tolerance against brain ischemia, with APN-KO mice suffering greater infarct volumes than wild-type mice5. Previous studies suggested the protective action of APN via an endothelial nitric oxide synthase-dependent mechanism, while Chen et al. confirmed the anti-inflammatory action of APN against cerebral ischemia/reperfusion (I/R) injury6. However, the contribution of other mechanisms to the neuroprotective action of APN requires further investigation.
Endoplasmic reticulum (ER) stress plays a critical role in various kinds of diseases including DM and neurodegeneration7. In the event of severe damage, apoptosis would be activated through ER stress-related unfolded protein response, leading to the elimination of damaged cells8. Accumulating evidences have suggested the involvement of ER stress in the regulation of post-cerebral I/R neuronal apoptosis9. Moreover, attenuation of ER stress could be a potential target for interfering brain I/R injury10. Recent data has suggested that the protective effect of gAD against myocardial I/R was modulated through suppressing of ER stress11. The involvement of ER stress in ischemia-induced vascular pathology in type II DM has also been proved12.
The current study aimed to investigate the neuroprotective effect of gAD on diabetic cerebral I/R model and further explore the underlying mechanisms. We assumed that with a proper therapeutic window, gAD could induce an anti-apoptotic effect against cerebral I/R injury through ER stress-related signaling pathway.
Results
gAD ameliorated cerebral I/R injury in T1DM-8W, but not in T1DM-2W mice
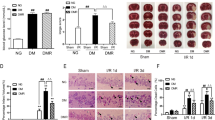
gAD failed to ameliorate I/R injury in T1DM-2W mice. There was no statistical difference in the neurological behavior among I/R, gAD + I/R and Veh + I/R groups (P > 0.05) and the infarct size 24 h after reperfusion in all these groups was comparable (P > 0.05). Furthermore, the neurological behavior scores in these groups were significantly deteriorated as compared with the Sham group (P < 0.05, Fig. 1A). On the other hand, the gAD + I/R group demonstrated significant improvements in neurological deficit scores 24 h after I/R compared with the I/R group in T1DM-8W mice (P = 0.028). Mice receiving gAD from 8-week DM group showed a significant decrease in infarct size 24 h after I/R compared with the I/R group (P = 0.009). No significant difference was observed between I/R and Veh + I/R groups (P = 0.79, Fig. 1B).

Varied effects of gAD against cerebral I/R.
Neurological scores and representative samples of TTC-stained brain sections from (A) T1DM-2W and (B) T1DM-8W mice. Cerebral infarct size was assessed by TTC staining. TTC-stained areas (red) indicate viable tissue while the white area shows the ischemic region. gAD significantly improved the neurological scores and decreased the infarct volumes in 8 W DM mice but not in the 2 W DM mice. Data are expressed as mean ± SD (n = 8 per group). #P < 0.05 vs. I/R group. Scale bars = 5 mm.
Adiponectin levels increased at 2-week and decreased at 8-week T1DM
We assessed APN in cerebral tissues by ELISA at various time points during T1DM progression. As shown in Fig. 2A, cerebral APN levels markedly increased after 1-week T1DM duration and peaked at 2-week T1DM; the levels then gradually decreased through the remainder of the study, up to 8 weeks.

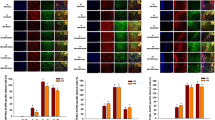
Dynamic alterations of APN and AdipoRs during T1DM progression.
(A) Total APN levels were determined by ELISA after various T1DM durations. Data are expressed as mean ± SD (n = 8 per group). $P < 0.05 vs. Control group; ^P < 0.05 vs. 2 W group. (B) Representative double immunofluorescence staining for AdipoR1, AdipoR2 (green) and neuron (NeuN, red) in brain sections. Scale bars = 20 μm. (C,D) Cerebral expression of AdipoR1 and AdipoR2 was determined by Western blot analysis. The upper part is the photograph of AdipoRs and their corresponding Tublin bands. Data are expressed as mean ± SD (n = 4 per group). ^P < 0.05 vs. 2 W group.
Cerebral AdipoR1 was up-regulated in T1DM-8W mice, but not in T1DM-2W mice
A small number of AdipoR1-positive cells were observed in the brain tissue of the T1DM-2W mice. However, the number of AdipoR1-positive neurons significantly increased in the T1DM-8W group. There was no difference in the number of AdipoR2-positive cells between the two groups (Fig. 2B). The same results were observed via Western blot analysis with higher AdipoR1 expression (P = 0.001; Fig. 2C) in the T1DM-8W group, but not in the AdipoR2 group (P > 0.05, Fig. 2D).
Up-regulation of AdipoR1 expression restored neuroprotection of gAD in T1DM-2W mice
According to the Western blot result, AdipoR1 was significantly increased in the RSG-treated group, while it showed no change in the Vehicle group (Fig. 3A).

Effect of rosiglitazone on neuroprotection of gAD in T1DM-2W mice.
(A) RSG significantly up-regulated the protein expression of AdipoR1 indicated by Western blot analysis (n = 4 per group). The upper part is the photograph of AdipoR1 and its corresponding Tublin bands. $P < 0.05 vs. Sham group. (B–D) Pretreatment with RSG significantly improved the protective effect of gAD in 2-week DM mice, with lower neurological scores and decreased infarct volumes (n = 8 per group). &P < 0.05 vs. gAD + I/R group. Scale bars = 5 mm.
The infarct volume in RSG + gAD + I/R group mice was significantly decreased as compared to that in gAD treated group 24 h after I/R in T1DM-2W mice (14.7% ± 4.3% vs. 31.2% ± 5.8%, P = 0.038, gAD + I/R vs. RSG + gAD + I/R). gAD failed to ameliorate the behavioral deficits while the neurological behavior outcome in mice pretreated with RSG was significantly improved 24 h after reperfusion injury (P = 0.028, gAD + I/R vs. RSG + gAD + I/R). There was no significant difference in the infarct volume or behavioral deficits score among I/R, gAD + I/R and Veh (R) + I/R groups in T1DM-2W mice (Fig. 3B–D).
Knockdown of AdipoR1 expression reversed neuroprotection of gAD in T1DM-8W mice
After intracerebroventricular administration of AdipoR1 siRNA or control siRNA, the AdipoR1 protein in DM mice brains was analyzed by Western blot. The results indicated that AdipoR1 expression was down-regulated by AdipoR1 siRNA compared to Sham or siRNA (n) (Fig. 4A).

Effect of AdipoR1 siRNA on neuroprotection of gAD in T1DM-8W mice.
(A) AdipoR1 siRNA significantly down-regulated the protein expression of AdipoR1 indicated by Western blot analysis (n = 4 per group). The upper part is the photograph of AdipoR1 and its corresponding Tublin bands. $P < 0.05 vs. Sham group. (B–D) Pretreatment with AdipoR1 siRNA attenuated the protective effect of gAD in 8-week DM mice with disruptive neurological behavior and increased infarct volumes, whereas that with AdipoR1 negative siRNA showed no such effect. Values are presented as mean ± SD (n = 8 per group). &P < 0.05 vs. gAD + I/R group. Scale bars = 5 mm.
Compared with the gAD + I/R group, the infarct volume in mice pretreated with AdipoR1 siRNA was significantly increased 24 h after I/R in T1DM-8W mice (14.7% ± 4.3% vs. 31.2% ± 5.8%, P = 0.038, gAD + I/R vs. siRNA + gAD + I/R). gAD could ameliorate the behavioral deficits while the neurological behavior outcome in siRNA + gAD + I/R group had significantly worsened 24 h after cerebral I/R (P = 0.028, gAD + I/R vs. siRNA + gAD + I/R). No significant difference in infarct volume or neurological scores was observed between the gAD + I/R and siRNA(n) + gAD + I/R groups in T1DM-8W mice (Fig. 4B–D).
gAD alleviated ER stress in T1DM-8W mice with cerebral I/R injury
GRP78, CHOP and caspase-12 are markers of ER stress while PERK and IRE1 are 2 major ER stress sensor protein. In the present study, the expressions of these proteins from the penumbra were investigated in T1DM-8Wmice. At 24 h after reperfusion, the expression of p-PERK, p-IRE1 and ER stress chaperone protein GRP78 significantly increased in gAD + I/R group (P < 0.05, gAD + I/R vs. I/R). However, AdipoR1-specific siRNA reversed this effect of gAD as indicated by the p-PERK, p-IRE1 and GRP78 expression significantly reduced in siRNA + gAD + I/R group (Fig. 5A,E).

Expression of ER stress protein and ER morphology in T1DM-8W mice
. Western blot analyses were performed for (A) GRP78, (B) CHOP and (C) cleaved caspase-12 (E) PERP, p-PERP, IRE-1, p-IRE-1 in the penumbra of different groups. The upper part is the photograph of ER stress proteins and their corresponding β-Actin bands. After MCAO, all the ER stress-related proteins were up-regulated. After gAD treatment, the expression of p-PERP, p-IRE-1, GRP78 was increased while that of CHOP and cleaved caspase-12 was decreased. The effects of gAD were reversed by pretreatment with AdipoR1 siRNA. Values are presented as mean ± SD (n = 4 per group). *P < 0.05 vs. Sham group. #P < 0.05 vs. I/R group. &P < 0.05 vs. gAD + I/R group. (D) Representative electron microscopy images of ER morphology. After I/R injury, the ER was disaggregated and difficult to identify. With gAD treatment, some ER could be identified as having regular shape. Scale bars = 200 nm.
The results revealed that I/R markedly increased expressions of CHOP (P = 0.001; I/R vs. Sham) and activated caspase-12 (P = 0.001, I/R vs. Sham), while gAD lowered them (P < 0.05; I/R vs. gAD + I/R, Fig. 5B [CHOP] and P = 0.028; I/R vs. gAD + I/R, Fig. 5C [activated caspase-12]). However, when pretreated with AdipoR1 siRNA, the expressions of CHOP (P < 0.05; gAD + I/R vs. siRNA + gAD + I/R) and activated caspase-12 protein were significantly lower than those in gAD group (P < 0.05; gAD + I/R vs. siRNA + gAD + I/R).
The ultrastructure of neurons was examined by TEM. The ER morphology changes such as clumping, margination and condensation were considered the most important evidence of injury. In I/R group, the damaged ER was condensed, verifying degeneration, while gAD treatment could ameliorate the injury of ER (Fig. 5D).
gAD inhibited neuronal apoptosis in T1DM-8W mice with cerebral I/R injury
Figure 6 showed the representative photomicrographs of TUNEL staining in the ischemic penumbra from different groups. In the Sham group, few TUNEL-positive cells were observed in the brain tissue 24 h after surgery. The number of TUNEL-positive neurons significantly increased in I/R group (P < 0.001, I/R vs. Sham) and markedly diminished in the gAD group (P = 0.003, gAD + I/R vs. I/R). However, when pretreated with AdipoR1 siRNA rather than the control siRNA, more apoptotic neurons were detected compared with gAD group (P = 0.038, gAD + I/R vs. siRNA + gAD + I/R).

Neuronal apoptosis in T1DM-8W mice with cerebral I/R.
Representative photomicrographs of TUNEL in the ischemic penumbra by immunohistology and quantitative analysis. gAD significantly decreased the number of TUNEL-positive cells. AdipoR1 siRNA reversed the anti-apoptotic effect of ER, whereas control siRNA showed no such effect. Scale Bar = 20 μm. Values are presented as mean ± SD (n = 5 per group). *P < 0.05 vs. Sham group. #P < 0.05 vs. I/R group. &P < 0.05 vs. gAD + I/R group.
Discussion
DM is a severe metabolic disorder with a worldwide prevalence of 6.4%. The total diabetic population is expected to reach 438 million13. APN is a cytokine that is primarily synthesized and secreted by fat cells. It has been proven to play a critical role in DM14,15. Stroke is a severe and common complication of DM. Diabetic patients are at an increased risk for cerebral ischemic injury16. Although there still remain some controversies and arguments17,18, some of the previous studies proved that low APN level is correlated with the severity of stroke and negatively related with brain infarct volume, as well as with the NIHSS score19,20. It is also associated with atherosclerosis21. A study by Sasaki et al. showed that circulating APN level is temporarily decreased in acute ischemic stroke patients and then recovers to baseline levels 14 days later22. It has also been proved that APN is effective against cerebral I/R through anti-inflammatory and anti-oxidative mechanisms23,24. Moreover, APN supplementation reduced the myocardial ischemic injury in T1DM mice25. Increased plasma APN was associated with decreased risk of Type 2 DM and thus, reduced risk of cardiovascular disease26. As globular APN is the main functional fragment of APN, our present study proved that gAD has a neuroprotecitve effect against cerebral I/R injury in 8-week DM mice. However, the protection was not observed in the 2-week DM mice.
To evaluate the dynamic therapeutic effects of gAD against cerebral ischemic injury in diabetic models, we focused on the change in APN level during the course of DM. We observed that APN levels peaked in the second week of T1DM and decreased during DM progression. Some studies indicated that APN concentration is positively related to cardiovascular risk in T1DM patients27,28, while others showed the opposite29,30. The dynamic change in the APN level might at least partially explain the previous contradictory results of APN in T1DM.
APN exerts its biological effects via its two receptors, AdipoR1 and AdipoR2 and previous studies offered some evidences for the expression of AdipoRs in the central nervous system31. However, the alteration of cerebral AdipoRs expression in a diabetic model, which would be essential for understanding the APN-AdipoR signaling pathway in the metabolic progression, has not been studied earlier. From our result, AdipoR1 expression decreased 2 weeks after the onset of DM but increased in the later stage, i.e. 8 weeks. It is interesting to note that no significant difference in AdipoR2 was observed in either phase. These results might help deduce the different roles of the 2 receptors in the course of DM.
On one hand, we tried to explain the possible reason for the failure of gAD in 2-week DM mice. Increased up-regulation of endogenous APN was observed at the early stage; however, it failed to protect against cerebral I/R. The administration of gAD in 2-week DM mice also showed no significant differences with respect to the infarct size and neurological behavior. This might be related to the lower expression of AdipoR1. To further prove our hypothesis, we used RSG as a peroxisome proliferator activated receptor gamma activator. It could increase AdipoR1 expression, thus activating the downstream AMPK-related signaling pathway32. After RSG injection, a neuroprotective effect of gAD was restored in 2-week T1DM mice, evidenced by a decreased infarct volume and improved neurological behavior. Therefore, we proved that failure of gAD in the early stage of DM was at least partially associated with decreased AdipoR1 expression.
On the other hand, to explain the mechanism of gAD in 8-week DM mice, we proved that AdipoR1 siRNA reversed the protective effect of gAD. This indicated the important role of AdipoR1 in the neuroprotective effect of gAD. Furthermore, the downstream signaling pathway was also explored. Cerebral ischemic reperfusion injury, together with DM, triggers multiple overlapping signaling pathways in the brain. It has been suggested that ER stress is involved in cerebral ischemic pathophysiology and neuronal apoptosis33. It also contributes to autoimmunity and insulin resistance in diabetic patients34.
ER chaperones are highly sensitive to I/R injury and are representative of ER stress35. We found that damaged ER morphology was ameliorated with gAD. ER stress markers such as GRP78, CHOP and caspase-12 were enhanced after I/R injury and reversed by gAD treatment in T1DM-8W mice (except for GRP78, which was further increased). GRP78 regulates protein folding and facilitates protein translocation in ER36. A previous study indicated that up-regulation of GRP78 attenuated the induction of CHOP and reduced ER stress-related apoptosis37. Therefore, the increased expression of GRP78 by gAD might contribute to the neuroprotective effect against I/R injury. Moreover, CHOP as a proapoptotic transcription factor plays a key role in ER stress-related apoptosis, while caspase-12 as a murine protein dissociates from the ER membrane and gets activated after ER stress38,39. Our results showed that the activation of both CHOP and caspase-12 was significantly suppressed by gAD. In addition, with the knockdown of AdipoR1, the regulation of gAD on GRP78, CHOP and caspase-12 expression were reversed, which showed the anti-ER stress was in close relationship with the gAD-AdipoR1 signaling pathway. For further exploring the possible molecular mechanisms of gAD, ER stress sensor proteins R-like endoplasmic reticulum kinase (PERK) and inositol-requiring enzyme 1 (IRE1) in neurons were examined. PERK and IRE1 are major ER stress sensor proteins. ER stress leads to isolation of GRP78 from PERK and IRE1, which in turn activates downstream ER stress response. Previous study suggested that PERK and IRE1 phosphorylation are crucial for maintaining the balance between cell survival and death40,41. Our results indicated that gAD could lead to the further activation of PERK and IRE1 pathways, which might lead subsequently to down-regulation of the pro-apoptotic protein CHOP and caspase12, while blocking AdipoR1 could reverse the effect of gAD. The alterations in p-IRE1 and p-PERK levels were consistent with those in GRP78 level. Dong et al., reported that ER stress proteins were unaffected by adiponectin in db/db diabetic obese mice42. The differences between results might due to the different type of mice as well as different injury model. Moreover, neuronal apoptosis is in close relationship with ER stress. Dysfunction of this organelle occurring at an early stage after ischemia might be the initial step of apoptotic cascades in neurons43. This apoptosis could reflect the function of the ER to some extent, while prolonged ER stress would ultimately lead to cell apoptosis. Therefore, the anti-apoptotic effect of gAD and the role of AdipoR1 in our study further support our assumption.
This study also had some limitations. Firstly, although we offered an evidence for the dynamic dysfunction of APN/AdipoR1 during T1DM progression as well as the protective role of gAD, the mechanisms might not be applicable for explaining type 2 diabetic models with highly variable metabolic properties. Further studies are required to determine the role of gAD and AdipoRs during type 2 DM and in different models. Secondly, our previous study demonstrated the protective effect of gAD against ischemic stroke in normal adult mice24. In the current study we focused only on T1DM and did not compare the differences between normal and DM mice. Thirdly, the study is not mechanistic enough. Our previous study showed that the neuroprotective action of gAD against ischemic stroke in normal adult mice resulted from the promotion of antioxidant capacity by inhibiting the NOX2 signaling system. Moreover, we also proved that electroacupuncture alleviated cerebral ischemic injury in diabetic mice stroke model through adiponectin/AdipoR1-mediated signaling pathway, in which glycogen synthase kinase 3-β might be one crucial molecule in the downstream pathway of AdipoR144. Nevertheless, the possible molecular mechanism of gAD for treating diabetic stroke should be addressed in future researches.
Above all, our study indicated the therapeutic window of globular APN against cerebral ischemic injury in diabetic mice. Accordingly, the dynamic dysfunction of APN and AdipoR1 could partially explain the deficient neuroprotective effect of gAD against I/R in 2-week T1DM mice but efficiently for 8-week T1DM mice. Moreover, ER stress-apoptosis signal pathway might play an important role for I/R injury in the late stage of T1DM mice and gAD-AdipoR1 could mitigate the injury. Interventions bolstering AdipoR1 expression during early stages and gAD supplementation during advanced stages may potentially reduce the cerebral ischemic injury in diabetic patients. We hope that our current results provide a reliable theoretical basis for future clinical treatment of ischemic stroke patients with DM.
Materials and Methods
Experimental protocol
All experiments were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23) revised in 1996 and associated guidelines. The experimental protocol used in this study was approved by the Ethics Committee for Animal Experimentation of the Fourth Military Medical University, Xi’an, China and the study was conducted according to the Guidelines for Animal Experimentation of the same university. Adult male C57BL/6 mice weighing 20–25 g were purchased from the Experimental Animal Center of the University and housed under controlled conditions with a 12-h light/dark cycle, at a temperature of 21 ± 2 °C and a humidity of 60–70% for at least one week prior to experimentation. The animals were allowed free access to standard rodent diet and water.
T1DM mice were induced via injection of low dose of streptozotocin (50 mg/kg, ip) for five consecutive days. The blood sugar was detected 10 days after the first day of injection and the mice with blood glucose level >13.3 mmol/L were considered as successful models for the experiment. This time point was defined as T1DM Day 0. The blood glucose and weight of the mice were measured weekly thereafter.
Experiment I
In order to test the neuroprotective effect of gAD against cerebral I/R injury in T1DM mice, 64 male mice were randomly assigned to Sham, I/R, gAD + I/R (gAD treatment, i.c.v. 30 min before I/R), or Veh + I/R (Vehicle as phosphate buffered saline [PBS] i.c.v. 30 min before I/R) treatment groups in 2-week T1DM (T1DM-2W) mice and 8-week T1DM (T1DM-8W) mice. I/R was performed to induce focal cerebral ischemic stroke.
Experiment II
In order to assess the dynamic change in APN level and alteration of AdipoR expression in diabetic mice, 88 mice were involved and the group was divided as Control, T1DM-2W mice and T1DM-8W mice.
Experiment III
In order to evaluate the effect of AdipoR1 on gAD-induced neuroprotection in T1DM-2W mice, 40 animals were randomly divided into I/R, gAD + I/R, RGS + I/R ([rosiglitazone, RGS], intraperitoneal injection, 2 mg/kg, 4 h before I/R), RSG + gAD + I/R and Veh (R) + I/R groups. RGS is an anti-diabetic drug and acts as a peroxisome proliferator-activated receptor gamma activator. In the current study, it was used to up-regulate AdipoR1 expression. Its doses were based on the previously published results and those of our preliminary study4,6,45,46.
Experiment IV
In order to investigate the role of AdipoR1 in T1DM-8W mice and to further explore the possible mechanisms of gAD, 84 T1DM-8W mice were assigned to I/R, gAD + I/R, siRNA + gAD + I/R (AdipoR1-specific siRNA, 0.3 nmol/μL, 1.2 nmol, i.c.v, 3 days before I/R) and siRNA (n) + gAD + I/R (negative siRNA, 3 nmol/μL, 1.2 nmol, i.c.v, 3 days before I/R) groups.
Drug injection
For gAD pretreatment, the mice were deeply anesthetized with 2.5% isoflurane and placed on a stereotaxic frame (Stoelting, Wood Dale, IL, USA). Recombinant mouse globular APN (gAD, produced in Escherichia coli [Peprotech EC, London, UK]) was injected into cerebroventricularl (i.c.v, bregma: 1.0 mm lateral, 0.5 mm posterior, 2.0 mm deep) at 30 min before ischemia and in a dose of 0.1μg/g body weight (diluted in PBS) as previously published45. PBS (pyrogen-free, 0.01 M, pH 6.8-7.4, 2 μL) was also injected as a vehicle control.
Transient focal cerebral ischemia
Focal cerebral ischemia was induced by middle cerebral artery occlusion (MCAO, under anesthesia by inhaling 2.5% isoflurane) in mice using the intraluminal filament technique, as previously described5. The suture was withdrawn 90 min after MCAO. Regional cerebral blood flow was measured by laser Doppler flowmetry (Periflux 5000, Perimed, Stockholm, Sweden) with a flexible probe fixed on the skull, 1 mm posteriorly and 6 mm laterally from the bregma and monitored throughout surgery.
Neurological score evaluation and infarct assessment
At 24 h after reperfusion, neurological deficit scores were assessed in a blinded manner according to the the 5-point scale method47. Briefly, normal motor function was scored as 0, flexion of the contralateral torso and forelimb on lifting the animal by the tail as 1, circling to the contralateral side but normal posture at rest as 2, leaning to the contralateral side at rest as 3 and no spontaneous motor activity as 4. After neurological score evaluation, the animals were decapitated under deep anesthesia (pentobarbital, 150 mg/kg body weight) and their brains were removed, frozen and sectioned (1 mm thick). The brain sections were stained with 2% (w/v) 2,3,5-Triphenyltetrazolium chloride (TTC; Sigma, St. Louis, MO, USA) to evaluate infarct volume, as previously described48.
ELISA
Samples of the ischemic penumbra were homogenized in cold RIPA lysis buffer with 1× Roche complete protease inhibitor cocktail and 1 mM phenylmethylsulfonyl fluoride on ice. The tissue extract was centrifuged at 12,000 × g at 4 °C for 30 min and the supernatant was removed and stored at −80 °C until use. The total APN concentration was measured by ELISA according to the instructions (Westang Biotechnology, Shanghai, China).
Immunofluorescence staining
For analysis of AdipoR1 and AdipoR2 expression, the mice from Experiment II were anesthetized and perfused with PBS and 4% paraformaldehyde 24 h after reperfusion. The brains were cut, prepared and stained as described in our previous study49. Different brain areas in each section from the same part of the brain in each group were observed and photographed by a microscope (20×; Olympus, BX51, Japan).
Western Blot Analysis
Immunoblot analysis was performed according to a previously described method48. The following primary antibodies were used: anti-phospho-IRE, anti-IRE1 (1:1000; Cell Signaling Technology, Danvers, MA, USA), anti-phospho-PERK, anti-PERK (1:1000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-GRP 78 antibody (1:200; Abcam, Cambridge, MA, USA), anti-CHOP antibody (1:1000; Abcam, Cambridge, MA, USA), anti-cleaved caspase-12 (1:200; Abcam, Cambridge, MA, USA), anti-AdipoR1 antibody (1:1000; BD Bioscience, Franklin Lakes, NJ, USA), anti-AdipoR2 antibody (1:1000; BD Bioscience, Franklin Lakes, NJ, USA), anti-β-actin antibody (1:10000; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and anti-β-tubulin antibody (1:10000; Santa Cruz Biotechnology, Santa Cruz, CA, USA). The membranes were then washed and incubated with suitable secondary antibodies for 1 h at room temperature. Antigens were detected using the chemiluminescence technique (Amersham Pharmacia Biotech Piscataway, NJ, USA). Image analysis was completed using the Bio-Rad Laboratories (Hercules) analysis software.
Transmission electron microscopy
Vibratome sections from the penumbra were selected and transferred onto the phosphate buffer. The tissues were rinsed in buffer and postfixed with 1% osmium tetroxide for 1 h, subjected to a graded ethanol dehydration series and infiltrated using a mixture of one-half propylene oxide and resin overnight. After 24 h, the tissues were embedded in resin. The 120-nm sections were cut and stained with 4% uranyl acetate for 20 min and 0.5% lead citrate for 5 min. The ultrastructure of the hippocampus was observed under transmission electron microscopy (TEM [Philips Tecnai 10, Philips, Holland]).
TUNEL staining
Neuronal cell apoptosis in the ischemic penumbra was quantified using a commercially available terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining kit (Roche Diagnostics, Mannheim, Germany) as previously described50. The total number of TUNEL-positive cells in the right hemisphere were counted in three different fields for each section in a naive manner using light microscopy (BX51; Olympus, Tokyo, Japan) and data from four animals in each group were averaged.
Statistical analysis
All values, excluding neurological scores, were expressed as mean ± SD. Statistical significance was tested by one-way analysis of variance and differences between the groups were detected using the Dunnett’s test. The neurological scores, presented as a median, were analyzed by a nonparametric method (Kruskal-Wallis test) followed by the Mann-Whitney U test with Bonferroni correction. P < 0.05 was considered statistically significant.
Additional Information
How to cite this article: Song, W. et al. Therapeutic window of globular adiponectin against cerebral ischemia in diabetic mice: the role of dynamic alteration of adiponectin/adiponectin receptor expression. Sci. Rep. 5, 17310; doi: 10.1038/srep17310 (2015).
References
Yang, W. et al. Prevalence of diabetes among men and women in China. N Engl J Med 362, 1090–1101 (2010).
Sarwar, N. et al. Diabetes mellitus, fasting blood glucose concentration and risk of vascular disease: a collaborative meta-analysis of 102 prospective studies. Lancet 375, 2215–2222 (2010).
Bejot, Y. & Giroud, M. Stroke in diabetic patients. Diabetes Metab 36 Suppl 3, S84–87 (2010).
Fruebis, J. et al. Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc Natl Acad Sci USA 98, 2005–2010 (2001).
Nishimura, M. et al. Adiponectin prevents cerebral ischemic injury through endothelial nitric oxide synthase dependent mechanisms. Circulation 117, 216–223 (2008).
Chen, B. et al. Adiponectin protects against cerebral ischemia-reperfusion injury through anti-inflammatory action. Brain Res 1273, 129–137 (2009).
Lin, J. H., Walter, P. & Yen, T. S. Endoplasmic reticulum stress in disease pathogenesis. Annu Rev Pathol 3, 399–425 (2008).
Kozutsumi, Y., Segal, M., Normington, K., Gething, M. J. & Sambrook, J. The presence of malfolded proteins in the endoplasmic reticulum signals the induction of glucose-regulated proteins. Nature 332, 462–464 (1988).
Nakka, V. P., Gusain, A. & Raghubir, R. Endoplasmic reticulum stress plays critical role in brain damage after cerebral ischemia/reperfusion in rats. Neurotox Res 17, 189–202 (2010).
Kishi, S. et al. Nerve growth factor attenuates 2-deoxy-d-glucose-triggered endoplasmic reticulum stress-mediated apoptosis via enhanced expression of GRP78. Neurosci Res 66, 14–21 (2010).
Guo, J. et al. Globular adiponectin attenuates myocardial ischemia/reperfusion injury by upregulating endoplasmic reticulum Ca2+−ATPase activity and inhibiting endoplasmic reticulum stress. J Cardiovasc Pharmacol 62, 143–153 (2013).
Amin, A. et al. Chronic inhibition of endoplasmic reticulum stress and inflammation prevents ischaemia-induced vascular pathology in type II diabetic mice. J Pathol 227, 165–174 (2012).
Wild, S., Roglic, G., Green, A., Sicree, R. & King, H. Global prevalence of diabetes: estimates for the year 2000 and projections for 2030. Diabetes Care 27, 1047–1053 (2004).
Spranger, J. et al. Adiponectin and protection against type 2 diabetes mellitus. Lancet 361, 226–228 (2003).
Maahs, D. M. et al. Low plasma adiponectin levels predict progression of coronary artery calcification. Circulation 111, 747–753 (2005).
Khoury, J. C. et al. Diabetes mellitus: a risk factor for ischemic stroke in a large biracial population. Stroke 44, 1500–1504 (2013).
Kuwashiro, T. et al. Significance of plasma adiponectin for diagnosis, neurological severity and functional outcome in ischemic stroke–Research for Biomarkers in Ischemic Stroke (REBIOS). Metabolism, doi: 10.1016/j.metabol.2014.04.012 (2014).
Arregui, M. et al. Adiponectin and risk of stroke: prospective study and meta-analysis. Stroke 45, 10–17 (2014).
Efstathiou, S. P. et al. Plasma adiponectin levels and five-year survival after first-ever ischemic stroke. Stroke 36, 1915–1919 (2005).
Chen, M. P. et al. Hypoadiponectinemia is associated with ischemic cerebrovascular disease. Arterioscler Thromb Vasc Biol 25, 821–826 (2005).
Gardener, H. et al. Adiponectin and carotid intima-media thickness in the northern Manhattan study. Stroke 43, 1123–1125 (2012).
Sasaki, M. et al. Hypoadiponectinemia in patients with cerebral infarction: comparison with other atherosclerotic disorders. Am J Med Sci 333, 140–144 (2007).
Jung, Y. S. et al. The role of adiponectin in secondary inflammatory reaction in cerebral ischemia. J Cerebrovasc Endovasc Neurosurg 15, 171–176 (2013).
Song, W. et al. Globular adiponectin elicits neuroprotection by inhibiting NADPH oxidase-mediated oxidative damage in ischemic stroke. Neuroscience 248C, 136–144 (2013).
Ma, Y. et al. Dynamic alteration of adiponectin/adiponectin receptor expression and its impact on myocardial ischemia/reperfusion in type 1 diabetic mice. Am J Physiol Endocrinol Metab 301, E447–455 (2011).
Lindberg, S. et al. Low adiponectin levels and increased risk of type 2 diabetes in patients with myocardial infarction. Diabetes Care 37, 3003–3008 (2014).
Shah, A. S. et al. Adiponectin and arterial stiffness in youth with type 1 diabetes: the SEARCH for diabetes in youth study. J Pediatr Endocrinol Metab 25, 717–721 (2012).
Fan, Q. et al. Nitrate tolerance aggravates postischemic myocardial apoptosis and impairs cardiac functional recovery after ischemia. Apoptosis 10, 1235–1242 (2005).
Messaaoui, A., Willems, D., Melot, C. & Dorchy, H. Risk markers for cardiovascular disease in young type 1 diabetic patients: lipoproteins, high-sensitivity C-reactive protein and adiponectin. Acta Clin Belg 67, 79–82 (2012).
Forsblom, C. et al. Serum adiponectin concentration is a positive predictor of all-cause and cardiovascular mortality in type 1 diabetes. J Intern Med 270, 346–355 (2011).
Thundyil, J. et al. Evidence that adiponectin receptor 1 activation exacerbates ischemic neuronal death. Exp Transl Stroke Med 2, 15 (2010).
Ding, G. et al. Adiponectin and its receptors are expressed in adult ventricular cardiomyocytes and upregulated by activation of peroxisome proliferator-activated receptor gamma. J Mol Cell Cardiol 43, 73–84 (2007).
Tajiri, S. et al. Ischemia-induced neuronal cell death is mediated by the endoplasmic reticulum stress pathway involving CHOP. Cell Death Differ 11, 403–415 (2004).
Zhu, M., Guo, M., Fei, L., Pan, X. Q. & Liu, Q. Q. 4-Phenylbutyric acid attenuates endoplasmic reticulum stress-mediated pancreatic beta-cell apoptosis in rats with streptozotocin-induced diabetes. Endocrine, doi: 10.1007/s12020-013-0132-7 (2013).
Truettner, J. S., Hu, K., Liu, C. L., Dietrich, W. D. & Hu, B. Subcellular stress response and induction of molecular chaperones and folding proteins after transient global ischemia in rats. Brain Res 1249, 9–18 (2009).
Wu, C. X. et al. Pinocembrin protects brain against ischemia/reperfusion injury by attenuating endoplasmic reticulum stress induced apoptosis. Neurosci Lett 546, 57–62 (2013).
Wang, X. Z. et al. Signals from the stressed endoplasmic reticulum induce C/EBP-homologous protein (CHOP/GADD153). Mol Cell Biol 16, 4273–4280 (1996).
Ma, Y., Brewer, J. W., Diehl, J. A. & Hendershot, L. M. Two distinct stress signaling pathways converge upon the CHOP promoter during the mammalian unfolded protein response. J Mol Biol 318, 1351–1365 (2002).
Nakagawa, T. & Yuan, J. Cross-talk between two cysteine protease families. Activation of caspase-12 by calpain in apoptosis. J Cell Biol 150, 887–894 (2000).
Duan, Z. et al. The PERK-eIF2alpha signaling pathway is involved in TCDD-induced ER stress in PC12 cells. Neurotoxicology 44, 149–159 (2014).
Han, Y. et al. Carbon monoxide offers neuroprotection from hippocampal cell damage induced by recurrent febrile seizures through the PERK-activated ER stress pathway. Neurosci Lett 585, 126–131 (2015).
Dong, F. & Ren, J. Adiponectin improves cardiomyocyte contractile function in db/db diabetic obese mice. Obesity 17, 262–268 (2009).
Hayashi, T. & Abe, K. Ischemic neuronal cell death and organellae damage. Neurol Res 26, 827–834 (2004).
Guo, F. et al. Electroacupuncture attenuates cerebral ischemia-reperfusion injury in diabetic mice through adiponectin receptor 1-mediated phosphorylation of GSK-3beta. Molecular Neurobiology, doi: 10.1007/s12035-014-8765-y (2014).
Qi, Y. et al. Adiponectin acts in the brain to decrease body weight. Nat Med 10, 524–529 (2004).
Masaki, T. et al. Peripheral, but not central, administration of adiponectin reduces visceral adiposity and upregulates the expression of uncoupling protein in agouti yellow (Ay/a) obese mice. Diabetes 52, 2266–2273 (2003).
Hase, Y. et al. Cilostazol, a phosphodiesterase inhibitor, prevents no-reflow and hemorrhage in mice with focal cerebral ischemia. Experimental Neurology 233, 523–533 (2012).
Wang, Q. et al. Limb remote postconditioning alleviates cerebral reperfusion injury through reactive oxygen species-mediated inhibition of delta protein kinase C in rats. Anesth Analg 113, 1180–1187 (2011).
Wang, Q. et al. Trans-activator of transcription-mediated delivery of NEP1-40 protein into brain has a neuroprotective effect against focal cerebral ischemic injury via inhibition of neuronal apoptosis. Anesthesiology 108, 1071–1080 (2008).
Wang, Q. et al. Activation of epsilon protein kinase C-mediated anti-apoptosis is involved in rapid tolerance induced by electroacupuncture pretreatment through cannabinoid receptor type 1. Stroke 42, 389–396 (2011).
Acknowledgements
This work was supported by the Overseas, Hong Kong & Macao Scholars Collaborative Researching Fund (Grant 81228022), the Program for Changjiang Scholars and Innovative Research Team in University (Grant IRT 1053) and the National Natural Science Foundation of China (Grants 81072888, 81171278, 81400952).
Author information
Authors and Affiliations
Contributions
W.S. acquired, analyzed and interpreted the data and drafted the manuscript. She is one of the guarantors of this work and had full access to all the data in the study and assumes responsibility for integrity of the data and accuracy of data analysis. F.G. analyzed and interpreted the data, performed statistical analysis and drafted the manuscript. H.Z., L.L. and R.Y. acquired the data. Q.W. handled the funding and supervision, analyzed and interpreted the data, performed statistical analysis and drafted the manuscript. L.X. conceived and designed the research, handled funding and supervision and made critical revisions to the manuscript with regard to important intellectual content.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Song, W., Guo, F., Zhong, H. et al. Therapeutic window of globular adiponectin against cerebral ischemia in diabetic mice: the role of dynamic alteration of adiponectin/adiponectin receptor expression. Sci Rep 5, 17310 (2015). https://doi.org/10.1038/srep17310
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep17310
- Springer Nature Limited
This article is cited by
-
Ischemic stroke and select adipose-derived and sex hormones: a review
Hormones (2018)
-
2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside protects murine hearts against ischemia/reperfusion injury by activating Notch1/Hes1 signaling and attenuating endoplasmic reticulum stress
Acta Pharmacologica Sinica (2017)
-
Cardioprotective effects of Notoginsenoside R1 against ischemia/reperfusion injuries by regulating oxidative stress- and endoplasmic reticulum stress- related signaling pathways
Scientific Reports (2016)