
Abstract
Climatic warming is expected to particularly alter greenhouse gas (GHG) emissions from soils in cold ecosystems such as tundra. We used 1 m2 open-top chambers (OTCs) during three growing seasons to examine how warming (+0.8–1.2 °C) affects the fluxes of carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O) from alpine tundra soils. Results showed that OTC warming increased soil CO2 efflux by 141% in the first growing season and by 45% in the second and third growing season. The mean CH4 flux of the three growing seasons was −27.6 and −16.7 μg CH4-C m−2h−1 in the warmed and control treatment, respectively. Fluxes of N2O switched between net uptake and emission. Warming didn’t significantly affect N2O emission during the first and the second growing season, but stimulated N2O uptake in the third growing season. The global warming potential of GHG was clearly dominated by soil CO2 effluxes (>99%) and was increased by the OTC warming. In conclusion, soil temperature is the main controlling factor for soil respiration in this tundra. Climate warming will lead to higher soil CO2 emissions but also to an enhanced CH4 uptake with an overall increase of the global warming potential for tundra.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
High-latitude and altitude tundra ecosystems are generally considered as one of the world’s most sensitive areas in response to global warming1. On the one hand, climatic warming of the earth’s surface is expected to be particularly strong at high northern latitudes and at high elevations2,3. On the other hand, soils of ‘cold’ ecosystems store large stocks of labile soil carbon4 and the temperature sensitivity of biogeochemical processes is the highest in cold climates5, suggesting that the greenhouse gas balance of cold systems responds particularly sensitive to the ongoing climate warming. Hydrological changes may lead to tradeoff between carbon dioxide (CO2) and methane (CH4) balance of ecosystems underlain by permafrost, with exchange rate of CO2 being usually several orders of magnitude greater than that of nitrous oxide (N2O) and CH4. However, CH4 and N2O have a greater warming potential than CO2. Hence, CH4 and N2O fluxes might be equally important to the greenhouse gas balance as soil CO2 effluxes6,7.
The net exchange rates of CH4, CO2 and N2O between soil and atmosphere are determined by the balance between sources and sinks. The fluxes of these greenhouse gases (GHG) are highly complex involving a number of soil biochemical processes, the composition of soil microbial and plant communities, as well as abiotic conditions8. Soil CO2 emission are mainly driven by temperature, moisture and the supply with substrate from plants5,9,10. Climate warming is generally increasing soil CO2 effluxes, but the overall net C balance of soils depends also on the C inputs by the vegetation11,12. For tundra and cold treeline ecosystems, it has been found that respiratory carbon losses responded more sensitive to increases in temperature than carbon gains by plants, which turned tundra ecosystems from a carbon sink into a carbon source13,14,15. Soil heating by 2.5 °C in a wet tundra increased soil CO2 emission by 39% at northeast coast of Greenland1. However, responses of soil respiration to warming are often short-lived and ecosystem-dependent16,17.
Fluxes of CH4 were found to be highly sensitive to climate warming in various cold ecosystems. Upland boreal soils in Siberia were found to consume atmospheric CH4 particularly during dry and warm summer months18. Warming is potentially increasing CH4 uptake due to soil drying and greater CH4 diffusion into soils7. In contrast, soil warming could decrease CH4 release by increasing soil NH4+ contents through a stimulated N mineralization which suppress the activities of methanotrophic bacteria19,20. In contrast, wetlands release large amounts of CH4 because methanogenesis is induced by anaerobic soil conditions. For instance, measurements in a high-arctic wet tundra ecosystem showed that the mean growing season CH4 fluxes ranged between 2.94 and 4.60 mg m−2 h−1 and were positively correlated with soil temperature21. However, in a boreal forest in interior Alaska, Wickland et al.22 did observe only weak relationships between CH4 flux and temperature. At an alpine treeline, Karbin et al.20 found that soil warming induced a shift in the depth distribution of methanotrophic bacteria, which, however, was not associated with significant responses of CH4 fluxes.
The N2O exchange between soil and atmosphere plays an important role in climate warming, global nitrogen cycling and stratospheric ozone depletion23. Climate warming may increase N2O fluxes by an accelerated N cycling but it may decrease the fluxes from partly anaerobic soils by a declining soil moisture23,24,25. Experimental warming (+3.5 °C) increased N2O emission in upland grassland where mean annual temperature was 8.7 °C23. However, the warming effects were only observed during the growing season but not in winter.
The majority of warming experiments in tundra quantifying fluxes of greenhouse gases have been carried out in arctic region26,27. The aim of our study was to investigate the responses of GHG fluxes to warming in a mountain tundra of China. In China, tundra exists only in Altai Mountain and Changbai Mountain. Changbai Mountain is a dormant volcano reaching up to 2734 m a.s.l. where the tundra zone is above 2000 m a.s.l.28. At the Songjiang weather station near Changbai Mountain, mean annual temperature in the region increased at a rate of 0.6 °C per decade from 2.2 °C in 1978 to 3.9 °C in 200629. In our study, here, we established open-top chambers (OTCs) according to the ITEX (International Tundra Experiment) standard to increase the growing season temperature (June to September). We have measured GHG fluxes in response to OTC-warming for three growing seasons from 2011 to 2013. Generally warming will lead to lower soil moisture. Thus, we expected that growing season warming will increase greenhouse gas exchange due to accelerated microbial activities and to a lower soil moisture promoting the gas diffusion into soils. Consequently, we hypothesized that the warming during the growing season will increase soil CO2 effluxes, but also the uptake of CH4 and N2O.
Result
Climatic variables
Mean air temperature during the growing seasons was 0.8 to 1.2 °C higher in the OTCs than in the control plots during the whole study period. The increase in mean soil temperature at 10 cm depths was about 0.2 °C. OTCs had negligible effects on relative air humidity, averaging +1.1%. Soil water contents were 0.02 to 0.03 m3m−3 higher in the OTCs as compared to the control plots during the three growing seasons, which corresponded to an increase of approximately 10%.
Soil CO2 efflux
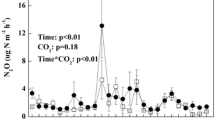
Warming increased soil CO2 efflux across three growing seasons (P = 0.017), but the magnitude of effect decreased from +141% in the first treatment year to approximately +45% in the following two years (Table 1). The mixed effect model for individual years indicated significant warming effects in 2011 and 2013, but not in 2012. The relatively high effect of OTC warming on soil CO2 efflux during the 2011 growing season was mainly attributed to the pulse in July (Fig. 1), but treatment effects were still significant without the initial peak (P = 0.035). Averaged across the three growing seasons in the Changbai Mountain tundra, the respiratory CO2 loss was 61% higher from the OTC warming than from the control plots.

Air relatively humidity, soil moisture, air temperature, soil temperature at 10 cm depth; CO2, CH4 and N2O fluxes for the warming OTCs (red) and the control plots (black) over three growing seasons’ period from 2011 to 2013.
Data show mean ± SE.
At a given soil temperature, soil CO2 effluxes were higher in the OTCs than in the controls and soil CO2 efflux increased exponentially with soil temperature at 10 cm depth for both OTCs and control plots (Fig. 2). The corresponding Q10 value in the control plots was 27% higher than that in the OTCs. No significant relationships between CO2 effluxes and air or soil moisture were found because of little variation of moisture throughout the experimental period (Fig. 1).

The relationship between soil temperature at 10 cm depth and soil CO2 efflux.
The Q10 values were 3.0 for the warming OTC (circles) and 3.8 for the control plot (diamonds), respectively.
CH4 Flux
Tundra soils in Changbai Mountain were a net sink for CH4 with OTC warming increasing the sink strength (Fig. 1; Table 1). Net production of CH4 was observed on 6 out of 27 measurements in the control treatment, but only at one date under OTC warming. Over the three-year long experimental period, OTC warming significantly increased CH4 flux (P = 0.014), but this effect differed among dates (PTreatment×Time < 0.001). In 2011 and 2013, there were significantly higher CH4 flux from the OTC warming than from the control plots (+122 and +96%, respectively; P < 0.05), but not in 2012 (+32%; P = 0.121). Averaged among all measurements, soils were a net sink for CH4 with −27.6 μg CH4-C m−2h−1 in the warmed and −16.7 μg CH4-C m−2h−1 in the control treatment. Over the entire measurement period and all plots, there were no significant relationships between CH4 fluxes and soil moisture as well as temperatures in air and soil.
N2O flux
In all years and in both treatments, N2O fluxes in Changbai mountain tundra switched between positive and negative values. Overall, the soils were a net N2O source during the growing seasons in 2011 and 2012, but a net N2O sink in 2013 (Pseason = 0.042). The experimental warming using OTCs tended to affect N2O fluxes (P = 0.094, across all three years). There was a significant difference between the two treatments for a number of measurements (12 out of 27 dates). However, as the direction of the treatment effect was not consistent, the warming effect during whole growing seasons were not significant in 2011 and 2012, but significant in 2013 when soils were mostly a N2O sink (P = 0.042; Table 1). In the third treatment year, OTC warming increased the mean N2O uptake during the growing season from −0.08 μg NO2-N m−2h−1 in the control plots to −1.95 μg NO2-N m−2h−1 in the OTC plots (P = 0.003). We did not find obvious relationship between N2O fluxes and microclimatic parameters either on an annual basis or for individual measurements.
Global warming potential
The GWP considering the fluxes of all three gases was dominated by soil CO2 effluxes (Table 1). Including CH4 and N2O fluxes decreased the GWP of CO2 emissions by less than 1%. Over the entire three growing seasons, OTC warming increased the GWP significantly by 70% (P = 0.018).
Discussion
Greenhouse gas fluxes from cold ecosystems such as tundra are assumed to respond particularly sensitive to climatic warming18,30,31,32. Our experimental warming in the Chinese Changbai Mountain tundra using OTCs supports this assumption. The passive warming increased soil CO2 effluxes and CH4 uptake for three growing seasons which was consistent with our hypothesis. The likely reasons for the altered greenhouse gases fluxes from the mountain tundra are an increased microbial activity and a change in the soil microclimate.
The stimulated belowground activity under warming is indicated by an accelerated soil respiration, which is in agreement with a number of warming studies in alpine and arctic environments17,33,34. Concurrent measurements of soil microbial biomass in the Changbai Mountain tundra did not reveal a significant warming effect in 201235, suggesting an enhanced metabolic activity of the soil microbial community with a less efficient use of C and smaller allocation of C into microbial growth36. The declining C use efficiency is in line with other studies in mountain ecosystems37,38 which can be explained by differential effects of temperature on growth and respiration36,39. In our OTC study, we found that the stimulation of soil respiration decreased strongly with treatment time. This decline is in agreement with other warming experiment and it is usually attributed to an acclimation of soil CO2 effluxes to warming either by a substrate depletion, an adaptation of soil microbial communities or a decreasing C allocation of plant assimilates to the rhizosphere16,38,40,41,42. Due to the moderate warming in our OTC study with an increase in air and soil temperature by less than 1.5 °C and relatively high soil organic C contents of 7% in the uppermost 10 cm, it seems unlikely that the depletion of labile soil carbon was primarily responsible for the strong decline after first growing season. Here, however, we can only speculate about the exact mechanism, but we rather relate the rapid and short-lived initial warming effect to an altered C allocation to the rhizosphere, which responds highly sensitive to microclimatic changes even at a daily time scale43.
The increased soil CO2 effluxes do not necessarily imply increased net ecosystem C losses because a frequently observed stimulation in plant growth in tundra ecosystems by warming12,17 potentially increases C inputs and hence, counterbalances C losses. In our study, we have indeed found a mean growth stimulation of approx. 7% in aboveground vegetation height (unpublished data), suggesting at least a partial offset of the enhanced soil CO2 effluxes by an increased plant growth. However, here, we cannot estimate a complete C balance, but a two-decade long summer warming in the Alaskan tundra by Sistla et al.12 indicated largely unchanged total soil C pools despite increased C cycling rates.
Our results also showed that soil respiration exponentially increased with soil temperature at 10 cm depth for both OTCs and control plots. This is in agreement with a number of studies observing a close positive relationship between soil respiration and temperature44,45,46,47. Soil moisture is often reported to control soil respiration48,49. However, in our study, soil moisture did not significantly affect soil respiration for both warming OTCs and control plots, probably because soil moisture ranged always between 0.2 and 0.3 mm−3 and hence in an range assumed to be optimal for respiratory activity. Moreover, it did not vary strongly during the whole measurement period and between treatments. We therefore conclude that soil temperature was the main controlling factor for soil respiration in this tundra. The relationship of soil CO2 efflux with temperature indicated that at a given temperature, the effluxes were higher in the warmed OTCs than the control plots (Fig. 2). This pattern suggests that the warming effect in our study goes beyond a simple temperature response, which we attribute to an indirect warming effect through an enhanced plant growth and to a higher allocation of assimilates to the belowground by the warming of soil under ‘cold’ conditions43.
The measurement of CH4 fluxes indicates that Changbai Mountain tundra is a CH4 sink which is in agreement with other observations in cold ecosystems with ‘upland’ soils in northern Siberia and the Alps, as well as in the high arctic tundra in Canada18,20,46. In our study, the OTC warming increased the CH4 sink strength by 66% (Table1), supporting experimental warming studies in the Canadian high Arctic32 and in the subarctic Scandes50. There are two potential reasons for the increased CH4 uptake under warming, higher soil temperature and a change in soil moisture51. Higher soil temperatures could enhance CH4 fluxes by accelerating microbial processes and enzyme activities. However, increases in soil temperature can stimulate both methanotrophic activity and methanogen activity at the same time52, which may balance out CH4 production and consumption in warmer soils53. In our study, we did not observe a significant relationship between CH4 uptake and temperature (air and soil temperatures) when data were aggregated over three growing seasons, but we cannot rule out that for individual measurements, warmer soil temperatures have induced a higher activity of soil methanotrophs. Similar poor correlations have also been found in other ecosystems, including tundra7,20,53. Alternatively, the increased CH4 uptake could be induced by an associated decrease in soil moisture under warming, because CH4 fluxes are closely related to the moisture status of soils by its effects on anaerobicity and gas diffusivity54,55. Higher temperatures generally induce a decline in soil moisture and hence, soil warming could decrease methanogenesis and facilitate CH4 diffusion into soils56. However, in our study, OTC warming increased soil moisture by about 10% due to the blocking of winds. We therefore conclude that it is unlikely that soil moisture was responsible for the greater CH4 uptake in the Changbai Mountain tundra. In contrast, the increased soil moisture might have partly balanced out the presumably positive temperature effect on soil methanotrophs. Thus, we expect that the increase in the CH4 sink strength we have observed here will even be larger during the ‘natural’ ongoing climate warming with an associated decline in soil moisture.
Observed responses of N2O flux to warming are highly variable, ranging from enhanced uptakes to increased emissions in various ecosystems7, even in the same ecosystem under different vegetation types26. In the present study in the Changbai Mountain tundra, N2O fluxes also varied between net emissions and net uptakes on an inter- and intra-annual timescale. Warming treatment alleviated the N2O emission in 2011 and 2012 though not significant but strongly enhanced N2O uptake during the growing season 2013. Similar results were found by Hu et al.45 in an alpine meadow where the effects of a less than 2 °C warming during the growing season on N2O fluxes varied from positive to negative responses with year and season. Also, in a temperate heathland ecosystem, a plateau peatland and a treeline ecotone, elevated temperatures did not significantly affect N2O fluxes20,51,57. The likely reason for the small and inconsistent responses in our and other studies are the number of processes involved with warming potentially accelerating N-cycling and hence denitrification58.
Conclusion
Our results indicated that the three-year OTC warming during growing season increased soil CO2 effluxes in the Changbai mountain tundra, but at the same time stimulated CH4 uptake and decreased N2O emission. Soil CO2 emissions dominated the global warming potential and OTC warming increased the overall GHG fluxes from soil. However, the GHG fluxes from soils might have partly been balanced out by an enhanced C uptake by plants and we thus expect the effect of warming to be smaller for the overall ecosystems GHG balance. We also have to recognize that our measurements were confined to growing seasons and for an annual ‘complete’ budget, estimates of GHG fluxes during the long winter would be needed. In addition, warming responses are frequently short-lived and only long-term manipulation experiments may represent the ongoing climatic warming. We therefore assume that our experiment is indicative for the intra-annual variability with greater GHG emissions to be expected during warmer summers.
Methods
Site description
The study was conducted in a tundra ecosystem at an elevation of 2028 m a.s.l., Changbai Mountain in northeastern China (41°58′–42°42′N; 127°67′–128°27′E). The climate is characterized by long and cold winters and short and cool summers. The mean annual temperature is −1.6 °C, with the highest mean diel temperature of 28 °C in August and the lowest one with −35 °C in January59. The distribution of precipitation over the year is uneven with the largest amount of rainfall in July and August. The study site is dominated by dwarf shrubs of Dryas octopetala var. asiatica, Vaccinium uliginosum and Rhododendron aureum, with an average coverage of 56%, 29% and 7%, respectively. Additional species are Carex atrata, Polygonum ochotense etc. The mean height of the vegetation canopy is about 8 cm during growing season (June to September). In the uppermost 10 cm of soil, the total contents of organic carbon, nitrogen and phosphorus were 6.95%, 0.37% and 0.54%, respectively.
Experimental design
Ten hexagon open-top chambers (OTCs) were installed on tundra according to the criteria of International Tundra Experiment in June 2010. The distance between two adjacent chambers was about 4 to 5 meters. All chambers were placed on relatively flat ground surface with similar vegetation cover. The experimental area almost covered 300 m2. The chambers were made of translucent plexiglas which had a high solar transmittance. They were 45 cm high and had inwardly inclined sides (60 cm at bottom, total area approx. 1 m2) which helped to trap heat and decrease wind speed. The control plot was set adjacent to each OTC within two meters having the same area and a similar vegetation. We used two sets of HOBO (Bourne, MA, USA) weather station locating in the OTC and the adjacent outside control, at a height of 15 cm above the ground surface recording air temperature, air relative humidity and photosynthetically active radiation (PAR) every half hour. Soil temperature and moisture was measured at 10 cm depth. Due to the thick snow cover, strong winds and low temperatures during winter in Changbai tundra, the experiment could only be conducted during the growing season, which lasted on average from June until September.
Flux measurements
Fluxes of CH4, CO2 and N2O fluxes were measured once a week during the growing season (June to September) from 2011 to 2013 except during rainy weather. The fluxes of the three greenhouse gas were measured using the static chamber method60 with six permanently installed cubic stainless-steel collars (length × width × height = 15.6 × 15.6 × 25 cm each) which were inserted into the soil to a depth of 6 cm in three OTCs and three control plots. Vegetation in the collars was removed during the whole measurements. For each measurement, a stainless-steel chamber was placed for 30 minutes on a water-filled rim of each collar providing a gas tight seal. Air temperature was measured inside each chamber with thermocouples. Gas samples were taken with a 30 cm3 plastic syringe four times at equal intervals of 10 min. The gas sampling was usually carried at about 9:00 a.m. at local time to represent daily mean flux, since previous studies in tropic forests and wetlands revealed that the flux at 9:00 a.m. almost equals the daily mean flux48,60. Sample gases were transported to the laboratory and were analyzed within four hours using gas chromatography (Hewlett 5890, USA). The CH4, CO2 and N2O flux rates were calculated from the slope of the temporal change in gas concentrations within the closed chamber. Further details of the method can be found in Song et al.60. Positive values refer to the flux from the soil to the atmosphere (emission) and negative values refer to the flux from the atmosphere to the soil (uptake or consumption).
Data analysis and statistics
The relationship between soil respiration (SR) and soil temperature at 10 cm depth (T) was estimated by fitting the following exponential function to the data of the three growing seasons:

where a and b were coefficients. The Q10 values were then calculated with Q10 = e10b.
The net global warming potential (GWP) from soils was calculated as sum of net emissions of CO2, N2O and CH4 by converting each gas unit to CO2 equivalent at a 100-yr time scale with climate change feedbacks using a conversion factor of 1 for CO2, 298 for N2O and 34 for CH4. The warming effect on GWP was estimated by the difference of the GWP in the warmed and control treatment.
The effects of warming and measurement date on CH4, CO2 and N2O fluxes was analyzed by fitting mixed-effects models by maximum likelihood (http://www.R-project.org/). The models included the sequential fixed effects Block, Treatment (Control vs. OTC chamber), as well as Time and Plot as the random effect accounting for the split-plot design and repeated measurement structure. Sampling times were used as categorical variables. Residuals of repeated measures showed a first-order autoregressive covariate structure, which was included in the model using the corAR1 function. For each measurement, the significant difference in three gas fluxes between the warming OTCs and the control plots was also assessed by One-way ANOVA followed by a Least Significance Difference (LSD) test using SPSS 16.0 system (SPSS Inc., Chicago, IL, USA). The dependent variables were all log or square-root transformed to meet the assumptions of normality and homoscedasticity. The relationship between temperature and moisture and gas fluxes was tested for all data of the three growing seasons using Pearson’s correlation analyses. Differences at the P < 0.05 level were considered significant.
Additional Information
How to cite this article: Zhou, Y. et al. Experimental warming of a mountain tundra increases soil CO2 effluxes and enhances CH4 and N2O uptake at Changbai Mountain, China. Sci. Rep. 6, 21108; doi: 10.1038/srep21108 (2016).
References
Mertens, S. et al. Influence of high temperature on end-of-season tundra CO2 exchange. Ecosystems 4, 226–236 (2001).
Pepin, N. et al. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Change 5, 424–430 (2015).
Jiang, Y., Zhuang, Q. & O’Donnell, J.A. Modeling thermal dynamics of active layer soils and near-surface permafrost using a fully coupled water and heat transport model. J. Geophys. Res. 117, D11110 (2012).
Sjögersten, S., Turner, B.L., Mahieu, N., Condron, L.M. & Wookey, P.A. Soil organic matter biochemistry and potential susceptibility to climatic change across the forest-tundra ecotone in the Fennoscandian mountains. Global Change Biol. 9, 759–772 (2003).
Davidson, E.A. & Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165–173 (2006).
Frolking, S. & Roulet, N.T. Holocene radiative forcing impact of northern peatland carbon accumulation and methane emissions. Global Change Biol. 13, 1079–1088 (2007).
Dijkstra, F.A. et al. Effects of elevated carbon dioxide and increased temperature on methane and nitrous oxide fluxes: evidence from field experiments. Front. Ecol. Environ. 10(10), 520–527 (2012).
Groffman, P.G., Hardy, J.P., Driscoll, C.T. & Fahey, T.J. Snow depth, soil freezing and fluxes of carbon dioxide, nitrous oxide and methane in a northern hardwood forest. Global Change Biol. 13, 1748–1760 (2006).
Lellei-Kovács, E. et al. Thresholds and interactive effects of soil moisture on the temperature response of soil respiration. Eur. J. Soil Biol. 47, 247–255 (2011).
Luan, J., Liu, S., Zhu, X., Wang, J. & Liu, K. Roles of biotic and abiotic variables in determining spatial variation of soil respiration in secondary oak and planted pine forests. Soil Biol. Biochem. 44, 143–150 (2012).
Smith, P. & Fang, C. Carbon cycle: A warm response by soils. Nature 464, 499–500 (2010).
Sistla, S.A. et al. Long-term warming restructures Arctic tundra without changing net soil carbon storage. Nature 497, 615–618 (2013).
Biasi, C. et al. Initial effects of experimental warming on carbon exchange rates, plant growth and microbial dynamics of a lichen-rich dwarf shrub tundra in Siberia. Plant Soil 307, 191–205 (2008).
Hagedorn, F. et al. Short-term responses of ecosystem carbon fluxes to experimental warming at the Swiss alpine treeline. Biogeochemistry 97, 7–19 (2010).
Natali, S.M. et al. Effects of experimental warming of air, soil and permafrost on carbon balance in Alaskan tundra. Global Change Biol. 17, 1394–1407 (2011).
Melillo, J.M. et al. Soil warming and carbon-cycle feedbacks to the climate system. Science 298, 2173–2176 (2002).
Dieleman, W.I.J. et al. Simple additive effects are rare: a quantitative review of plant biomass and soil process responses to combined manipulations of CO2 and temperature. Global Change Biol. 18, 2681–2693 (2012).
Flessa, H. et al. Landscape controls of CH4 fluxes in a catchment of the forest tundra ecotone in northern Siberia. Global Change Biol. 14, 2040–2056 (2008).
Stiehl-Braun, P.A., Hartmann, A.A., Kandeler, E., Buchmann, N. & Niklaus, P.A. Interactive effects of drought and N fertilization on the spatial distribution of methane assimilation in grassland soils. Global Change Biol. 17, 2629–2639 (2011).
Karbin, S., Hagedorn, F., Dawes, M.A. & Niklaus, P.A. Treeline soil warming does not affect soil methane fluxes and the spatial micro-distribution of methanotrophic bacteria. Soil Biol. Biochem. 86, 164–171 (2015).
Tagesson, T. et al. Land-atmosphere exchange of methane from soil thawing to soil freezing in a high-Arctic wet tundra ecosystem. Global Change Biol. 18, 1928–1940 (2012).
Wickland, K.P., Striegl, R.G., Neff, J.C. & Sachs, T. Effects of permafrost melting on CO2 and CH4 exchange of a poorly drained black spruce lowland. J. Geophys Res 111, G02011 (2006).
Cantarel, A.A.M. et al. Four years of experimental climate change modifies the microbial drivers of N2O fluxes in an upland grassland ecosystem. Global Change Biol. 18, 2520–2531 (2012).
Bijoor, N.S., Czimczik, C.I., Pataki, D.E. & Billings, S.A. Effects of temperature and fertilization on nitrogen cycling and community composition of an urban lawn. Global Change Biol. 14, 2119–2131 (2008).
Goldberg, S.D. & Gebauer, G. Drought turns a Central European Norway spruce forest soil from an N2O source to a transient N2O sink. Global Change Biol. 15, 850–860 (2009).
Brummell, M.E., Farrell, R.E. & Siciliano, S.D. Greenhouse gas soil production and surface fluxes at a high arctic polar oasis. Soil Biol. Biochem. 52, 1–12 (2012).
Stewart, K.J., Brummell, M.E., Coxson, D.S. & Siciliano, S.D. How is nitrogen fixation in the high arctic linked to greenhouse gas emissions? Plant Soil 362, 215–229 (2013).
Liu, Q.J., Li, X.R., Ma, Z.Q. & Takeuchi, N. Monitoring forest dynamics using satellite imagery—a case study in the natural reserve of Changbai Mountain in China. Forest Ecol. Manag. 210, 25–37 (2005).
Li, G., Bai, F. & Sang, W.G. Different responses of radial growth to climate warming in Pinus koraiensis and Picea jezoensis var. komarovii at their upper elevational limits in Changbai Mountain, China. Chinese J. Plant Ecol. 35(5), 500–511 (2011).
Zhuang, Q. et al. Net emissions of CH4 and CO2 in Alaska: implications for the region’s greenhouse gas budget. Ecol. Appl. 17, 203–212 (2007).
Sturtevant, C.S. & Oechel, W.C. Spatial variation in landscape-level CO2 and CH4 fluxes from arctic coastal tundra: influence from vegetation, wetness and the thaw lake cycle. Global Change Biol. 19, 2853–2866 (2013).
Allan, J. et al. Methanogen community composition and rates of methane consumption in Canadian High Arctic permafrost soils. Env. Microbiol. Rep. 6, 136–144 (2014).
Lu, M. et al. Responses of ecosystem carbon cycle to experimental warming: a meta-analysis. Ecology 94, 726–738 (2013).
Wang, X. et al. Soil respiration under climate warming: differential response of heterotrophic and autotrophic respiration. Global Change Biol. 20, 3229–3237 (2014).
Wang, X.J., Zhou, Y.M., Wang, X.X., Jiang, X.J. & Han, S.J. Responses of soil enzymes in activity and soil microbes in biomass to warming in tundra ecosystem on Changbai Mountains. Acta Pedologica Sin. 51, 166–175 (2014).
Allison, S.D., Wallenstein, M.D. & Bradford, M.A. Soil-carbon response to warming dependent on microbial physiology. Nat. Geosci. 3, 336–340 (2010).
Schindlbacher, A. et al. Experimental warming effects on the microbial community of a temperate mountain forest soil. Soil Biol. Biochem. 43, 1417–1425 (2011).
Streit, K. et al. Soil warming alters microbial substrate use in alpine soils. Global Change Biol. 20, 1327–1338 (2014).
Manzoni, S., Taylor, P., Richter, A., Porporato, A. & Ågren, G.I. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol. 196, 79–91 (2012).
Luo, Y.Q., Wan, S.Q., Hui, D.F. & Wallace, L.L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature 413, 622–625 (2001).
Hartley, I.P., Heinemeyer, A. & Ineson, P. Effects of three years of soil warming and shading on the rate of soil respiration: substrate availability and not thermal acclimation mediates observed response. Global Change Biol. 13, 1761–1770 (2007).
Tucker, C.L., Bell, J., Pendall, E. & Ogle, K. Does declining carbon-use efficiency explain thermal acclimation of soil respiration with warming? Global Change Biol. 19, 252–263 (2013).
Ferrari, A., Hagedorn, F. & Niklaus, P.A. Experimental soil warming and cooling alters the partitioning of recent assimilates: evidence from a 14C-labelling study at the alpine treeline. Oecologia (on line) 10.1007/s00442-015-3427-y (2016).
Briones, M.J.I., Poskitt, J. & Ostle, N. Influence of warming and enchytraeid activities on soil CO2 and CH4 fluxes. Soil Biol. Biochem. 36, 1851–1859 (2004).
Hu, Y. et al. Effects of warming and grazing on N2O fluxes in an alpine meadow ecosystem on the Tibetan plateau. Soil Biol. Biochem. 42, 944–952 (2010).
Lamb, E.G. et al. A High Arctic soil ecosystem resists long-term environmental manipulations. Global Change Biol. 17, 3187–3194 (2011).
Laganière, J., Paré, D., Bergeron, Y. & Chen, H.Y.H. The effect of boreal forest composition on soil respiration is mediated through variations in soil temperature and C quality. Soil Biol. Biochem. 53, 18–27 (2012).
Tang, X.L., Liu, S.G., Zhou, G.Y., Zhang, D.Q. & Zhou, C.Y. Soil-atmospheric exchange of CO2, CH4 and N2O in three subtropical forest ecosystems in southern China. Global Change Biol. 12, 546–560 (2006).
Carbone, M.S. et al. Seasonal and episodic moisture controls on plant and microbial contributions to soil respiration. Oecologia 167, 265–278 (2011).
Sjögersten, S. & Wookey, P.A. Spatio-temporal variability and environmental controls of methane fluxes at the forest-tundra ecotone in the Fennoscandian mountains. Global Change Biol. 8, 885–894 (2002).
Yang, G. et al. Effects of soil warming, rainfall reduction and water table level on CH4 emissions from the Zoige peatland in China. Soil Biol. Biochem. 78, 83–89 (2014).
Topp, E. & Pattey, E. Soils as sources and sinks for atmospheric methane. Can. J. Soil Sci. 77, 167–178 (1997).
Dijkstra, F.A., Morgan, J.A., Follett, R.F. & Lecain, D.R. Climate change reduces the net sink of CH4 and N2O in a semiarid grassland. Global Chang Biol. 19, 1816–1826 (2013).
Smith, K.A. et al. Exchange of greenhouse gases between soil and atmosphere: interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 54, 779–791 (2003).
Yrjälä, K. et al. CH4 production and oxidation processes in a boreal fen ecosystem after long-term water table drawdown. Global Change Biol. 17, 1311–1320 (2011).
Wu, X. et al. Effects of soil moisture and temperature on CO2 and CH4 soil–atmosphere exchange of various land use/cover types in a semi–arid grassland in Inner Mongolia, China. Soil Biol. Biochem. 42, 773–787 (2010).
Carter, M.S. et al. Effects of elevated atmospheric CO2, prolonged summer drought and temperature increase on N2O and CH4 fluxes in a temperate heathland. Soil Biol. Biochem. 43, 1660–1670 (2011).
Maag, M. & Vinther, F.P. Effects of temperature and water on gaseous emissions from soils treated with animal slurry. Soil Sci. Soc. Am. J. 63, 858–865 (1999).
Xu, Q.Q., Liu, Q.J. & Zhang, G.C. Soil nitrogen dynamics and productivity of snowpack Sanguisorba sitchensis community in alpine tundra of Changbai Mountain, China. Chinese J. Plant Ecol. 35(2), 159–166 (2011).
Song, C.C., Xu, X.F., Tian, H.Q. & Wang, Y.Y. Ecosystem–atmosphere exchange of CH4 and N2O and ecosystem respiration in wetlands in the Sanjiang Plain, Northeastern China. Global Change Biol. 15, 692–705 (2009).
Acknowledgements
This research was supported by National Natural Science Foundation of China(31170461) and by Open Research Fund Program of Changbai Mountain Academy of Sciences (201504). We would like to thank Professor Shijie Han, the director of Changbai Mountain Station, Chinese Academy of Sciences for the help in facilitating field trip to experimental sites. We also thank Qingjun Lai for his assistance in the field and laboratory measurements.
Author information
Authors and Affiliations
Contributions
Y.Z. wrote the manuscript text and executed statistical analysis. Y.Z. and M.H.L. designed the experiment. F.H. and M.H.L. revised the manuscript text and analysis methods. C.Z., X.J. and X.W. contributed to field experiments and gas measurements. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhou, Y., Hagedorn, F., Zhou, C. et al. Experimental warming of a mountain tundra increases soil CO2 effluxes and enhances CH4 and N2O uptake at Changbai Mountain, China. Sci Rep 6, 21108 (2016). https://doi.org/10.1038/srep21108
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep21108
- Springer Nature Limited
This article is cited by
-
Modelling impacts of lateral N flows and seasonal warming on an arctic footslope ecosystem N budget and N2O emissions based on species-level responses
Biogeochemistry (2022)
-
Effects of mixing biochar on soil N2O, CO2, and CH4 emissions after prescribed fire in alpine meadows of Wugong Mountain, China
Journal of Soils and Sediments (2020)
-
Biocrusts Modulate Responses of Nitrous Oxide and Methane Soil Fluxes to Simulated Climate Change in a Mediterranean Dryland
Ecosystems (2020)
-
Nitrous oxide emissions from three temperate forest types in the Qinling Mountains, China
Journal of Forestry Research (2019)
-
Impacts of precipitation, warming and nitrogen deposition on methane uptake in a temperate desert
Biogeochemistry (2019)