
Abstract
Melanocortin-3 receptors (MC3R) have a contextual role in appetite control that is amplified with hypocaloric conditioning. C57BL/6J (B6) mice subjected to hypocaloric feeding schedules (HFS) exhibit compulsive behavioral responses involving food anticipatory activity (FAA) and caloric loading following food access. These homeostatic responses to calorie-poor environs are attenuated in B6 mice in which Mc3r transcription is suppressed by a lox-stop-lox sequence in the 5’UTR (Mc3rTB/TB ). Here, we report that optimization of caloric loading in B6 mice subject to HFS, characterized by increased meal size and duration, is not observed in Mc3rTB/TB mice. Analysis of hypothalamic and neuroendocrine responses to HFS throughout the light-dark cycle suggests uncoupling of hypothalamic responses involving appetite-stimulating fasting-responsive hypothalamic neurons expressing agouti-related peptide (AgRP) and neuropeptide Y (Npy). Rescuing Mc3rs expression in Nkx2.1(+ve) neurons is sufficient to restore normal hypothalamic responses to negative energy balance. In addition, Mc3rs expressed in Nkx2.1(+ve) neurons are also sufficient to restore FAA and caloric loading of B6 mice subjected to HFS. In summary, MC3Rs expressed in Nkx2.1(+ve) neurons are sufficient to coordinate hypothalamic response and expression of compulsive behavioral responses involving meal anticipation and consumption of large meals during situations of prolonged negative energy balance.
Similar content being viewed by others

Introduction
Neural nutrient-sensing networks projecting from control nodes in the hypothalamus and brainstem increase chances of surviving nutrient scarcity by increasing feeding-related motivation while conserving energy1,2. Studying these systems is clinically relevant, as the appetitive responses controlled by these neurons limit the efficacy of obesity treatments using self-governed hypocaloric diets and bariatric surgery3,4. Central nervous melanocortin system neurons in the hypothalamus and brainstem are focal points for internal cues of metabolic condition5. Much attention has been given to how neural melanocortin-4 receptors (MC4R) optimize appetite and energy expenditure to conserve body mass in hypo- and hyper-caloric dietary conditions6. However, melanocortin-3 receptors (MC3R) expressed in hypothalamic and limbic structures also control appetitive behaviors7,8,9,10,11,12,13,14,15,16,17,18. Food anticipatory activity (FAA), a progressive increase in movement anticipating nutrient availability, is attenuated in Mc3r-deficient mice7,8,10,11. FAA indicates optimization of foraging behavior during times of nutrient scarcity, and involves synchronizing peak rhythms in activity with nutrient availability using circadian (“time-keeping”) functions19. MC3Rs expressed in limbic structures regulate feeding-related motivational responses during weight loss14, and exert sex-specific control over reward-related behaviors13. Hypothalamic and adrenal responses to fasting are also attenuated in Mc3r-deficient mice12. While MC4R signaling also regulates appetite6, it cannot compensate for loss of MC3Rs in hypocaloric situations.
While the mechanisms underlying the behavioral phenotype of Mc3r-deficient mice are unclear, abnormal responses of ‘first-order’ melanocortin neurons to sensory inputs may be a critical factor10,12. Fasting normally activates GABAergic neurons residing in the arcuate nucleus of the hypothalamus (ARC) co-expressing the orexigenic neuropeptide agouti-related peptide (AgRP) and neuropeptide y (Npy). AgRP is an MC3R/MC4R inverse agonist/antagonist; both AgRP and NPY are potent orexigens. Ablating AgRP/Npy neurons in adult mice causes anorexia20,21,22, and compromises adaptation to hypocaloric conditioning23. On the other hand, chemo- or opto-genetic activation rapidly induces feeding in mice24,25. Increased AgRP and Npy expression in response to fasting12 and hypocaloric conditioning10 is markedly attenuated in Mc3r-deficient mice. While it has not been determined whether this correlates with reduced neural activity, this result could nevertheless imply a complete or partial desensitization to internal cues of metabolic condition. Desensitization of AgRP/Npy neurons in Mc3r-deficient mice would also explain lack of compensation by MC4Rs, and other orexigenic neuropeptides/neurotransmitters released by AgRP/Npy neurons26, in hypocaloric situations.
Here we further investigated the role of neural MC3Rs in the expression of adaptive responses to hypocaloric conditions. Analysis of gross food intake suggests compulsive behavioral responses involving consumption of large meals in hypocaloric situations requires MC3Rs9. We therefore compared meal structure in Mc3r-deficient mice and controls in situations of ad libitum and hypocaloric feeding conditions. Mc3r expression is concentrated in hypothalamic and limbic structures13,14,27. Rescuing Mc3r expression in mesolimbic dopamine neurons only partially rescues the expression of homeostatic responses to hypocaloric conditions, improving motivation to self-administer food rewards without affecting gorging14. To decipher the contribution of MC3Rs expressed within hypothalamic nuclei to these responses, we rescued Mc3r transcription in LoxTBMc3r mice8 using transgenic mice expressing Cre in Nkx2.1(+ve) neurons28,29. Nkx2.1 (also known as thyroid transcription factor 1) is a transcription factor controlling development of the ventral hypothalamus and telencephalon in the CNS, and the thyroid, pituitary and lung in the periphery30,31. The Nkx2.1-Cre line has previously been used by others to investigate nutrient-responsive signaling pathways in hypothalamic and preoptic areas32,33,34. Our results suggest that MC3Rs expressed in Nkx2.1(+ve) neurons are sufficient for coordinating appetitive and hypothalamic responses to internal cues of metabolic state, but are insufficient for normal nutrient partitioning.
Results
Optimizing caloric loading performance during HFS requires MC3Rs
Previous studies examining feeding behavior in Mc3r-deficient mice reported gross intake, but did not assess meal structure9,12,14,15,35,36,37,38. We therefore first compared the effect of Mc3r-deficiency on meal structure, using male Mc3rTB/TB mice and WT controls. At baseline, Mc3rTB/TB mice weighed more than controls (30.2 ± 0.9 vs. 25.7 ± 0.7 g, p < 0.01) due to increased fat mass (FM) and a tendency for reduced fat-free mass (FFM) (FM 8.5 ± 0.5 vs. 6.0 ± 0.6 g, p < 0.05; FFM 14.8 ± 0.5 vs. 16.6 ± 0.5 g, values are estimated marginal means derived from ANCOVA using total body mass as a covariate). Baseline food intake in the ad libitum condition was similar when expressed in g/d (3.1 ± 0.3 vs. 3.0 ± 0.3 g), or adjusted for body mass and/or body composition (data not shown). Meal frequency, size and duration were not affected by genotype in the ad libitum condition (Fig. 1A–C), although bout frequency was significantly lower in Mc3rTB/TB mice (Fig. 1D, p < 0.001). While WT and Mc3rTB/TB mice ate the same total amount of food each day during HFS (data not shown), their feeding patterns were markedly different (Fig. 1E–G). WT mice responded to HFS by gorging, reducing meal frequency (Fig. 1E) and increasing meal size (Fig. 1F) and duration (Fig. 1G); these responses were all markedly attenuated in Mc3rTB/TB mice (Fig. 1E–G).

Baseline data are averaged over 2 d after 10d acclimation (A–D), or during HFS (2.2 g/d presented at ZT4) (E–G). (A) Meal frequency, (B) meal size, (C) meal duration and (D) number of bouts data were averaged from ad libitum feeding data recorded on day 11 and 12, presented as mean ± SEM. In (A–D), black bars = WT mice, n = 6; white bars = Mc3rTB/TB mice, n = 7. (E–G). After acclimation and measurement of baseline data, food was removed and mice subject to HFS. (E) Meal frequency, (F) meal size and (G) meal duration are shown for each day during the HFS. For (E–G), Two-way ANOVA with repeated measures indicated significant genotype effects on meal number, size and duration (all p < 0.05); a significant effect of time (# days on HFS) on meal frequency (p < 0.01), size and duration (both p < 0.001); and an interaction between genotype and time for meal number (p < 0.05), size and duration (both p < 0.001). (H,I) Food self-administration by lever pressing in WT and Mc3rTB/TB mice trained during the lights-on period (H) or dark period (I). Food self-administration is shown for mice subjected to HFS. Two-way ANOVA with repeated measures revealed a significant genotype effect (lights-on period, p < 0. 001; dark period, p < 0. 05) and training condition (lights-on or dark) effect on active lever pressings (lights-on period, p < 0. 001; dark period, p < 0. 001), with no interaction between genotype and training condition. *p < 0.05. **p < 0.01. ***p < 0.001. Black circles = WT mice; white circles = Mc3rTB/TB mice, n = 8/group.
We also assessed food self-administration by lever pressing in WT and Mc3rTB/TB mice maintained in situations of negative energy balance. As previously reported14, Mc3rTB/TB mice exhibited reduced self-administration of food rewards during training sessions in the lights-on period (Fig. 1H). To control for possible circadian-related phenotype effects7,11, a separate group of mice trained during the dark period was examined; the phenotype was not affected by lighting schedule (Fig. 1I). Reduced appetite observed in Mc3r-deficient mice thus appears to be independent of the time of day at which food is available.
Neuroendocrine responses to HFS are preserved in Mc3r-deficient mice
We next assessed whether Mc3r genotype alters circadian and non-circadian responses to HFS. The response of neuroendocrine hormones to phase shifts in caloric intake exhibits circadian and non-circadian (meal-related) patterns19. Changes in hormonal levels preceding meal time may signal impending caloric load, priming the digestive system to minimize disruption of nutrient homeostasis. We bred large numbers of age and sex-matched Mc3rTB/TB and littermate controls to allow for a comparison daily rhythms in major neuroendocrine hormones that control ingestive behavior and metabolism, and to compare hypothalamic gene expression as a measure of the neuroendocrine responses to altered metabolic condition associated with HFS.
At baseline, there was a modest but highly significant effect of genotype on body mass (WT mice, 26.96 ± 0.21 g, n = 77; Mc3rTB/TB mice, 29.93 ± 0.36 g, n = 79, p < 0.001). Unadjusted ad libitum food intake was similar between genotypes (WT, 2.95 ± 0.03 g/d, Mc3rTB/TB mice, 2.95 ± 0.04 g/d). However, when including body mass and energy balance (weight gain or loss during the course of measurement) as covariates, Mc3rTB/TB mice consume 6% fewer calories (2.86 ± 0.03 vs. 3.04 ± 0.03 g/d, p < 0.001).
A portion of the WT (n = 48) and Mc3rTB/TB mice (n = 49) were then assigned to HFS and provided 2.20 ± 0.02 g/d of food (70–75% of their habitual calorie intake) at ZT4; food remaining in the hopper was weighed at ZT8 on day 2, 4 and 6 to estimate speed of food consumption. After 6 days of HFS, WT mice consumed 100% of food provided within 4 h of food presentation. In contrast, Mc3rTB/TB mice had only consumed a portion (58.72 ± 3.94%) of the food after 4 h. This did not affect weight loss during the HFS (weight loss in g for WT, 2.88 ± 0.16 g; Mc3rTB/TB mice, 2.83 ± 0.13 g) as food remaining in cages housing Mc3rTB/TB mice (0.91 ± 0.09 g) was returned and consumed later.
As predicted, HFS had a significant effect on all three hormones measured (P < 0.001), with a significant effect of time of day (p < 0.05 for leptin, p < 0.001 for acyl-ghrelin and corticosterone, or CORT) and interaction between diet (HFS or ad libitum) and time of day (all p < 0.001) due to changes in feeding-related rhythms in circulating concentrations (Fig. 2A–D). HFS was associated with lower circulating leptin levels prior to the meal, with levels rising after food consumption (Fig. 2B). Acyl-ghrelin and CORT levels increased with HFS (Fig. 2C,D). Overall, these responses were retained in Mc3rTB/TB mice (Fig. 2A–D). For leptin, there was however a significant interaction between all three variables (genotype, time and feeding condition) (p < 0.01). Mc3rTB/TB mice are hyperleptinemic owing to obesity, however a decline in plasma leptin concentrations during HFS was still observed (Fig. 2A,B); meal-related increases were observed irrespective of genotype, but were more pronounced in Mc3rTB/TB mice.

P values for the effects of genotype (G), diet condition (D) and zeitgeber time (T) and their interactions determined by univariate analysis (A). Significant P values are in red. Leptin (B), acyl-ghrelin (C), and corticosterone (CORT) (D) plasma levels at different time points during AL (square) or HFS (circle) conditions (n = 7–8). The dotted lines represent the time of meal presentation under HFS; dark period is indicated by grey shading. Black circles = WT mice; White circles = Mc3rTB/TB mice. Note that for the AL condition, samples were not collected at ZT19 and ZT7.
The interaction between all three variables (genotype, time and feeding condition) was not significant for acyl-ghrelin and CORT. Plasma acyl-ghrelin concentrations increased with HFS (diet effect, p < 0.001) at the time points preceding food presentation but were normal in the post-prandial period, irrespective of genotype (Fig. 2A,C). There was a strong trend for an effect of genotype (p = 0.06), perhaps due to lower levels pre-meal (Fig. 2A,C), and a significant effect of zeitgeber time (p < 0.001). However, irrespective of genotype acyl-ghrelin levels generally exhibited a clear peak at ZT23, and had begun to decline at ZT3 (1 h prior to food presentation) to normal levels.
HFS increased the amplitude of the daily rhythm of CORT, irrespective of genotype (Fig. 2A,D). For mice on HFS, a bimodal pattern of response was observed with a minor peak anticipating meal presentation and a second peak at the onset of the dark phase. While CORT levels began to increase in anticipation of mealtime, the peak may be an artifact induced by suppression following the meal at ZT4. There was a strong trend for an effect of genotype (p = 0.07), which may be due to lower CORT levels in Mc3rTB/TB mice, particularly in the time points prior to feeding time (ZT23), and a highly significant effect of time (p < 0.001) due to the daily rhythm.
Hypothalamic responses to HFS are attenuated in Mc3r-deficient mice
We previously published that hypothalamic AgRP/Npy expression is lower prior to meal time (ZT3) in Mc3rTB/TB under HFS, and that hypothalamic AgRP/Npy expression correlates with FAA10. To further investigate meal-related responses to HFS, we measured expression of neuropeptides involved in appetite regulation expressed in ARC neurons (AgRP, Npy, Pomc and Cart) throughout the light/dark cycle in the mice used to assess neuroendocrine responses. We also examined genes linked to the stress response (Fkb5, Cdkn1a, Nr3c1, Nr13c2) and leptin signaling (LepRb, Socs3).
As expected, HFS markedly increased expression of AgRP and Npy mRNA in the hypothalamus in WT mice (Fig. 3A–C). HFS also reduced expression of Pomc and Cart mRNA encoding anorexigenic neuropeptides in WT mice (Fig. 3A,D–E). In Mc3rTB/TB mice, AgRP and Npy expression were not responsive to HFS (Fig. 3A,B), while suppression of Pomc and Cart mRNA were attenuated (Fig. 3D,E).

Table representing for each gene studied P values of multiple way ANOVA for genotype (G), diet condition (HFS or ad libitum, D) and zeitgeber time (T) and their interactions (A). Significant P values are in red. Relative expression of AgRP (B) , Npy (C), Pomc (D), and Cart mRNA (E) at different time points during ad libitum (AL, square) or hypocaloric feeding schedule (HFS, circle) conditions (n = 7–8). The dotted lines represent the time of meal presentation under HFS; dark period is indicated by grey shading. Black circles = WT mice; White circles = Mc3rTB/TB mice. Note that for the AL condition, samples were not collected at ZT19 and ZT7. Scatterplots showing associations between plasma corticosterone (CORT) and hypothalamic AgRP (F), Npy (G) and Pomc (H) relative expression for WT and Mc3rTB/TB mice. Dark grey circles = WT mice; White circles = Mc3rTB/TB mice. Data shown are pooled from samples collected in the AL and HFS condition.
This experiment also allows for an analysis of meal-related responses of hypothalamic gene expression. Surprisingly, AgRP expression was not suppressed by feeding, with peak expression occurring after meal at the same time as CORT (ZT11, compare Fig. 2D and 3B). Indeed, of the three hormones measured, CORT showed the strongest association with hypothalamic gene expression (Suppl. materials Table 1). A highly significant strong positive association was observed between plasma CORT and hypothalamic AgRP and Npy expression in WT mice (Suppl. materials Table 1, Fig. 3F,G), there was also a weak negative association between plasma CORT and hypothalamic Pomc mRNA (Suppl. materials Table 1, Fig. 3H). While still significant, the associations between CORT and AgRP, Npy and Pomc expression were weakened in Mc3rTB/TB mice (Suppl. materials Table 1, Fig. 3F–H).
Genotype did not affect LepRb expression (Suppl. materials Fig. S1A,B). Socs3 expression was slightly increased in Mc3rTB/TB mice, consistent with hyperleptinemia (Fig. S1A–C). No major differences were also observed in hypothalamic Nr3c1 (glucocorticoid receptor, Suppl. materials Fig. S1A–D), or Nr3c2 (mineralocorticoid receptor; Suppl. materials Fig. S1A–E).
Fkbp5 and Cdkn1a are stress responsive genes regulated during food restriction39. Daily profiles in Fkbp5 and Cdkn1a expression were similar (Fig. 4A–C), and correlated strongly with plasma CORT concentrations (compare Figs 2D with 4B and C; Suppl. materials Table 1). During HFS, WT mice exhibited evidence of two peaks in Cdkn1a and Fkbp5 expression coinciding with peaks in CORT; a major peak at ZT15 during the dark period and minor peaks at ZT3 anticipating food presentation (Fig. 4B,C). In Mc3rTB/TB mice, peaks anticipating meal presentation was absent (Fig. 4B-C), correlating with a lower nadir and weakened pre-meal increase in circulating CORT (cf. Fig. 2D). An identical pattern of changes associated with genotype and feeding condition was observed for Fkbp5 in the cerebellum, suggesting systemic failure (Fig. 4A,E).

Table representing for each gene studied P values of multiple way ANOVA for genotype (G), diet condition (D) and zeitgeber time (T) and their interactions (A). Significant P values are in red. Relative expression of Fkbp5 (B) and Cdkn1a in hypothalamus (HYP) (C), and Fkbp5 mRNA in the cerebellum (Cer) (D) at different time points (n = 7–8/group). The dotted lines represent the time of meal presentation under HFS; dark period is indicated by grey shading. Black circles = WT mice; White circles = Mc3rTB/TB mice. Note that for the AL condition, samples were not collected at ZT19 and ZT7.
To rule out compromised glucocorticoid signaling in Mc3rTB/TB mice, we performed an additional study. Mc3rTB/TB mice and littermate control were treated with dexamethasone (DEX, 10 mg/kg ip.) at ZT3 and then euthanized at ZT7. Genotype did not significantly affect treatment induced weight loss (Suppl. materials Fig. S2A) or stimulation of glucocorticoid-regulated genes (Suppl. materials Fig. S2B–E). DEX treatment modestly but significantly increased AgRP mRNA, irrespective of genotype (Suppl. materials Fig. S2F). No treatment effects were observed for Npy, Pomc or Cart expression (Suppl. materials Fig. 2F).
As anticipatory responses to HFS exhibits a circadian component19, we also examined expression of core clock and clock-output genes. No major effects of feeding condition, or interactions between feeding condition and genotype were observed in the hypothalamus, although were significant effects of feeding condition on Bmal1 and Npas2 expression and of genotype on Npas2 and Nr1d1 expression (Suppl. materials Fig. S3A–D,I). The presence of multiple oscillators in the hypothalamus may have confounded this analysis40; we therefore analyzed clock gene expression in the cerebellum, a region of the brain linked to motor control that harbors a food-entrainable oscillator41. There was a significant interaction between genotype, feeding condition and time (Suppl. materials Fig. S3E–I). Interestingly, only Per2 was clearly affected by genotype around meal-time, with WT but not Mc3rTB/TB mice exhibiting an increase anticipating food presentation (Suppl. materials Fig. S3H–I).
Altogether, the neuroendocrine response to HFS does not appear to be markedly compromised by loss of MC3Rs. In contrast, meal-related patterns in the expression of hypothalamic neuropeptides involved in regulating appetite are clearly disturbed in Mc3rTB/TB mice. While the expression of stress-regulated genes in Mc3rTB/TB mice appears overall to be responsive to HFS, there is evidence for a specific meal-related event that is Mc3r-dependent. Our findings thus suggest loss of MC3R-signaling causes a hypothalamic defect in the capacity to respond to fluctuations of circulating signals of energy status. Finally, the response of AgRP and Npy expression to HFS differs suggesting independent regulation. It is also worth noting that food intake did not suppress AgRP expression (Fig. 3B), with expression continuing to rise after the meal in line with CORT (Fig. 2D).
Dissociation of feeding-related and nutrient-partitioning roles of MC3Rs
We next examined whether neural MC3Rs are sufficient to restore normal appetitive responses during HFS. We first used nestin-Cre to broadly target neural transcription in Mc3rTB/TB mice (NES-MC3R), as previously described8. While the nestin-Cre strain is not an ideal tool for selectively targeting the CNS42, we have observed strong Cre activity relative to another Cre strain targeting the CNS (synapsin-Cre) when using reporter mice (Girardet and Butler, unpublished observations). We therefore used it in proof-of-principle studies to determine whether rescuing Mc3r transcription in the brain would restore FAA. Analysis of Mc3r expression in the hypothalamus and medial habenula (MHab) indicated partial rescue in NES-MC3R relative to controls (Fig. 5A).

(A) Assessment of Mc3r transcription in the hypothalamus (HYP) and medial habenula (MHab) of NES-MC3R mice. (B) Body composition showing averaged values by genotype (left panel) and scatterplots showing the relationship between fat mass (FM, center panel) and fat-free mass (FFM, right panel) and total body mass. (C) Wheel running data for WT (black) and Mc3rTB/TB (blue) mice fed ad libitum, and then after 3, 4, 6 or 7d of hypocaloric feeding. (D) Wheel running data for Nestin-Cre (green) and NES-MC3R (red) mice fed ad libitum (AL) and then after 3, 4, 6 or 7d of hypocaloric feeding (HFS). (E) Averaged FAA data on days 3 and 4 of HFS; (F) Averaged FAA data on days 6 and 7 of HFS. In (A,B,E and F) columns not sharing letters are significantly different, p < 0.05.
Nestin-Cre mice exhibit a mild hypopituitarism phenotype with reduced growth hormone secretion and altered body mass42. Body composition data are shown in Fig. 5B. In our laboratory condition, nestin-Cre mice exhibit a nutrient partitioning phenotype (FM for nestin-Cre negative, 1.74 ± 0.13 g; nestin-Cre positive, 3.13 ± 0.16 g; p < 0.001; FFM for nestin-Cre negative,16.07 ± 0.08 g; nestin-Cre positive, 15.17 ± 0.10 g, p < 0.001; values are estimated marginal means adjusted for total body mass, n = 33 for nestin-Cre negative;Mc3rTB/TB, n = 23 for nestin-Cre positive;Mc3rTB/TB or wt/wt). The anticipated effect of Mc3r-deficiency on adiposity was also observed (FM for Mc3rwt/wt, 1.95 ± 0.18 g; Mc3rTB/TB, 2.93 ± 0.16 g; p < 0.01; FFM for Mc3rwt/wt,15.88 ± 0.12 g; Mc3rTB/TB , 15.35 ± 0.10 g, p < 0.01; n = 25 for nestin-Cre (+ve or −ve); Mc3rwt/wt; n = 31 for nestin-Cre (+ve or −ve); Mc3rTB/TB). For both FM and FFM, there was a significant interaction between nestin-Cre and Mc3r genotype (P < 0.05), with the predicted nutrient partitioning phenotype associated with Mc3r-deficiency observed in nestin-Cre (−ve) but not in nestin-Cre (+ve) mice (Fig. 5B).
Comparison of FAA in “WT”, which are nestin-Cre (−ve) mice with normal Mc3r expression, and Mc3rTB/TB mice shows the previously observed attenuated response (Fig. 5C,E,F). However, comparison of nestin-Cre positive and NES-MC3R mice shows no difference (Fig. 5D,E,F). FAA thus appears to be normalized in NES-MC3R mice compared to nestin-Cre controls (Fig. 5D,F) that are matched for adiposity, suggesting the role of MC3Rs in regulating food-seeking behaviors is dissociated from, and independent of, effects on adiposity and nutrient-partitioning.
MC3R expressed in Nkx2.1(+ve) neurons are sufficient for FAA and hypothalamic responses during HFS
Nkx2.1-Cre targets transcription in regions of the forebrain and hypothalamus28,29. We confirmed Cre activation in our laboratory condition using an inducible tdTomato reporter mouse (Fig. 6A). Using qRT-PCR, rescue of Mc3r transcription was observed in the hypothalamus of nkx2.1-Cre;Mc3rTB/TB (NKX-MX3R) mice at levels comparable to that observed for nestin-Cre, with no rescue in the MHab (Fig. 6B). Regression analysis of body composition data shows the anticipated preferential deposition of FM over FFM when expressed as estimated marginal means adjusted for body mass in Mc3rTB/TB mice (Fig. 6C–H). NKX-MC3R mice exhibited a modest rescue of the nutrient partitioning phenotype associated with Mc3r-deficiency in males (Fig. 6C–E) and females (Fig. 6F–H). NKX-MC3R mice accumulated 1–2 g less FM as a function of total body mass compared to Mc3rTB/TB mice, irrespective of sex (Fig. 6D,G). In males and female mice, the impact of MC3Rs expressed in Nkx2.1(+ve) neurons on normalizing FFM was also modest (Fig. 6E,H).

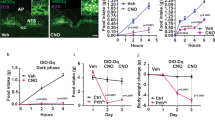
(A) Confocal image obtained from a reporter Nkx2.1Cre; ROSA-tdTomato mouse showing Nkx2.1-expressing cells at the level of the mediobasal hypothalamus. (B) Assessment of hypothalamic Mc3r expression by qRT-PCR revealed a recombination efficiency of 16% of Nkx2.1-Cre, but no rescue in the MHab. (C–H) Regression analysis comparing body composition in males (C–E) and females (F–H). Body composition (FM, FFM and H2O content) adjusted for total body mass are shown for males (C) and females (F). The Nkx2.1-Cre transgenic mice had normal values, and were therefore combined into one control group (Cre+ or −); values not sharing letters are significantly different, p < 0.01 for FM, FFM, p < 0.05 for H2O. Scatterplots showing the impact of genotype on relationship between body mass and FM (D,G) or FFM (E,H) are shown for males (D,E) and females (G,H). In (B,C and F) columns not sharing letters are significantly different, p < 0.05.
We next compared responses of NKX-MC3R and Mc3rTB/TB to situations of negative energy balance. Measurement of 4 h food intake during time restricted feeding revealed lower intake in Mc3rTB/TB mice compared to WT controls (Fig. 7A); this difference was not observed when NKX-MC3R mice were compared to Nkx2.1-Cre mice (Fig. 7B). An analysis of hypothalamic gene expression in mice fed ad libitum or subject to HFS for 6d indicated that the response of AgRP/Npy neurons was restored in NKX-MC3R mice (Fig. 7C,D). In this experiment, there was no significant effect of timed restricted feeding or genotype on Pomc expression (data not shown), perhaps due to differences in feeding protocol and/or sex (females were used for this experiment versus males used for the experiment shown in Fig. 3). Finally, analysis of FAA indicated impaired behavioral adaptation to daytime feeding in Mc3rTB/TB mice was rescued in NKX-MC3R mice (Fig. 8A–C). These results suggest that MC3Rs expressed in Nkx2.1(+ve) neurons are sufficient to restore the response of AgRP/Npy gene expression to HFS and for expression of FAA.

Food intake in the 4 h period following time-restricted feeding (TRF) is compared (A) Hypothalamic expression (relative to WT controls fed ad libitum) is shown for AgRP (B) and Npy (C) in female mice fed ad libitum (AL) or after 6 d of restricted feeding (n = 4–6/group). Restricted feeding significantly increased expression of AgRP and Npy in WT, Nkx2.1-Cre and NKX-MC3R mice, but not in Mc3rTB/TB mice. In A, *p < 0.05 compared to controls (WT vs. Mc3rTB/TB ). In (B and C) *p < 0.05 compared to AL condition; **p < 0.01 compared to AL condition.

(A) Wheel running data is shown for controls, Mc3rTB/TB and NKX-MC3R mice at baseline in ad libitum condition (AL), and after 3, 4, 6 and 7 days of HFS (mice fed at ZT4). Averaged FAA data for days 3 and 4 (B), and days 6 and 7 (C), are shown. The FAA data shown in B and C are activity in the 3 h window prior to food presentation (ZT1-ZT4), and are adjusted for 24 h activity. (Mc3rTB/TB mice, n = 20; NKX-MC3R mice, n = 19; controls, n = 38). Controls are pooled WT and Nkx2.1-Cre mice which exhibited similar FAA responses. *p < 0.05 compared to controls; **p < 0.01 compared to controls, p < 0.05 compared to NKX-MC3R mice.
Discussion
These studies provide important new information on the importance of MC3R signaling in driving homeostatic appetitive responses to hypocaloric conditioning. It is well known that weight loss is associated with homeostatic responses involving increased appetite. The current results confirm that intact MC3R signaling is crucial for compulsive behavioral responses involving consumption of large meals in situations of prolonged negative energy balance. This phenotype may be at least partially explained by the failure of crucial “fasting-responsive” neurons in the ARC to respond to internal cues of metabolic condition.
While the analysis of hypothalamic expression of orexigenic neuropeptides is not novel, the data nevertheless reinforce previous observations by our laboratory10 and others12. Moreover, combining a rigorous analysis of daily patterns and meal effects in neuroendocrine stress-related hormones and hypothalamic expression of stress-responsive genes, provides novel insights into the relationship between CORT and stress-responsive genes in the nervous system. Finally, the observation that MC3Rs expressed in Nkx2.1(+ve) neurons are sufficient to restore appetitive responses to hypocaloric conditioning is novel, and highly significant. This important data further refines our understanding of how MC3R signaling neurons may exert strong regulatory control over local and peripheral nutrient sensor signals on the cluster of fasting-responsive neurons that govern appetite.
Feeding protocols limiting caloric intake are a severe metabolic stressor for mice43,44. Most food consumption occurs within 1 h of food presentation; mice therefore adapt to repetitive cycles of gorging interrupted by prolonged fasting periods (>20 h)9,44. The gorging response is minimally affected by the method used to restrict calories. It is observed when food access is restricted to 4 h windows each day using mechanical barriers9, or when a hypocaloric meal is placed at 24 h intervals in food-hoppers44. An examination of meal structure (Fig. 1) clearly demonstrates the abnormal behavioral response of Mc3r-deficient mice to HFS. The predicted gorging behavior observed in control mice involves a progressive adaptation of feeding strategy leading to fewer larger meals. When populations are competing for limited nutrient supply, this response could be considered as adaptive. That this response is clearly and markedly attenuated in Mc3r-deficient mice provides further evidence of a crucial role for this receptor in the expression of altered feeding behavior (appetite) in hypocaloric situations.
The literature on the impact of Mc3r-deficiency on feeding behavior when nutrients are not limiting is ambiguous. Various groups have reported hypophagia38, normal intake8,9,37 or even modest hyperphagia15,36 in Mc3r-deficient models, suggesting sensitivity to background strain and environment. Here we presented data from two experiments in the ad libitum condition. The data from the largest study reported to date (N = 77–79/group) suggest a modest reduction in Mc3r-deficient mice when caloric intake is adjusted using body mass and energy balance as covariates. The result showing reduced frequency of bouts assessed using an automated system in the ad libitum condition is novel, and may indicate reduced interest in food.
The significance of these results for the human condition is unclear given the rarity of MC3R mutations inducing loss of function in the general population. However, two studies reported a “slowness-in eating” phenotype with MC3R mutations that alter signaling properties in cultured cells17,18. Our results suggest that the feeding phenotype associated with loss of MC3R signaling in mice is amplified with hypocaloric conditioning. Whether MC3R haploinsufficiency in humans would similarly affect appetite during situations of prolonged hypocaloric conditioning has not been explored. Our data also raise interesting questions about how selective MC3R compounds would affect feeding behavior.
Appetitive responses to weight loss can limit the effectiveness of voluntary reduction in caloric intake as an obesity therapy. The finding that rescuing Mc3r expression in Nkx2.1(+ve) neurons restores compulsive behavioral responses involving meal anticipation and consumption of large meals is therefore significant. Multiple populations of neurons are involved in the activation of motivational responses, with fasting responsive AgRP neurons forming a focal point for local (synaptic and nutrient sensing) and humoral inputs from the periphery45,46,47. These observations suggest a population of Nkx2.1(+ve);Mc3r(+ve) neurons that are instrumental in integrating multiple signals of metabolic condition to enhance appetite during hypocaloric situations.
Cre activity in Nkx2.1-Cre mice has been mapped to GABAergic, Npy+ve, Pomc+ve and TH+ve neurons in the ARC and glutamatergic neurons in the ventromedial hypothalamus (VMH)29. It is however important to note that Cre activity in this model is not restricted to the hypothalamus. Cre activity has been observed throughout the telencephalon, including the cerebral cortex, amygdala, olfactory bulb, striatum, globus pallidus, septum, nucleus basalis and hippocampus28. Mc3r expression has been observed in the amygdala, preoptic area and hippocampus27. While it is possible that MC3Rs expressed in hypothalamic nuclei support appetitive responses to hypocaloric conditions, further studies are required to identify which subset of MC3R(+ve);Nkx2.1(+ve) neurons is responsible for restoring appetite responses. MC3Rs expressed by steroidogenic factor-1 (SF1) neurons in the ventromedial hypothalamus appear to be insufficient to restore FAA8. Further investigation of the network of hypothalamic Nkx2.1(+ve);Mc3r(+ve) neurons populations is warranted.
Another limitation to the current study is that the examination of hypothalamic responses was limited to gene expression, which may not necessarily correlate with changes in neural activity48. Further studies assessing neural activity using other techniques are therefore needed. It is also unclear whether Nkx2.1(+ve);Mc3r(+ve) neurons are actively involved in the daily processing of metabolic cues, or affect maturation of hypothalamic circuits governing appetite.
Interestingly, while MC3Rs expressed in Nkx2.1(+ve) neurons appear to be linked strongly to neural circuits governing appetite control, they appear insufficient to restore normal nutrient partitioning. Based on the current results and previously published observations8,14, the identity and site of action for MC3Rs involved in regulating nutrient partitioning remains unclear.
Other significant advances provided by these experiments stem from the analysis of the neuroendocrine response to HFS. Neuroendocrine hormones and gut peptides exhibiting anticipatory responses to meal-time may have a role in preparing the nervous system for nutrient consumption, enhancing feeding efficiency when nutrients are limiting49,50,51. With HFS, acyl-ghrelin and CORT exhibited increased amplitude in the 24 h rhythm (Fig. 2). For acyl-ghrelin, the peak anticipates mealtime irrespective of genotype. This suggests that entrainment of acyl-ghrelin released to anticipate mealtime in response to food-entrainable oscillators in X/A-like cells of the stomach52 is retained in Mc3rTB/TB mice.
The impact of genotype and HFS on plasma leptin concentrations was predictable. As reported35,38,53, obesity associated with Mc3r-deficiency mice causes hyperleptinemia. HFS reduced plasma leptin concentrations during the premeal phase, with a marked postprandial increase observed irrespective of genotype. While plasma leptin concentrations in Mc3r-deficient mice were markedly reduced in the period leading up to food presentation (ZT23 and ZT3), the level did not fall below the values for controls fed ad libitum. This observation potentially confounds our interpretation of studies examining behavioral and metabolic adaptation in Mc3r-deficient mice. The decline in leptin is a powerful signal driving behavioral and metabolic adaptation to weight loss1,2. It is thus feasible that failure of leptin to fall below a threshold level may contribute to the phenotype observed in these experiments. However, the results from the NES-MC3R experiment suggest that the loss of food anticipatory behaviors associated with Mc3r-deficiency is not secondary to obesity, as FAA is restored independently of reductions in FM. Moreover, as shown in Fig. 5, NKX-MC3R mice exhibit only modest improvements in adiposity but have marked improvement in food anticipatory behaviors, gorging behavior and responsiveness of AgRP/Npy to signals of altered metabolic condition.
For CORT and the hypothalamo-pituitary-adrenal (HPA) axis, interpreting the interaction between genotype and feeding condition is more complex. While there was a marked increase in amplitude, the retention of the peak at ZT11 marking the transition between light and dark periods is surprising, as we anticipated a phase shift with peak levels occurring pre-meal based on the literature49. This discrepancy could be due to insufficient duration of HFS to produce a phase shift in the control of adrenal steroidogenesis by circadian oscillators54. However, work by other laboratories indicated that peripheral clocks adjust with 7d of restricted feeding55. Furthermore, the daily pattern of CORT fluctuations under restricted feeding also depends on the interval between meal presentation and activity onset and the severity of the caloric restriction56,57.
The current study consolidates evidence for dissociation between circadian oscillator-controlled stress responses responsive to light- and calorie-dependent inputs58,59. The response of glucocorticoid-regulated “stress response” genes in the hypothalamus and cerebellum suggests HFS has induced a peak in the activity of stress-regulated genes that anticipates food presentation. The response is likely driven by or responsive to systemic non-local inputs, as we could not detect Mc3r expression in the cerebellum, consistent with other reports5. The absence of the feeding-related peak in the hypothalamus and cerebellum of Mc3r-deficient mice suggests an independent stress-response governed by nutrient availability that is dependent on MC3R signaling. The second peak entrained to the transition between the light- and dark-phases is retained in Mc3r-deficient mice, and likely represents the light-entrained rhythm in adrenal steroidogenesis54.
Renquist et al. reported that the response of the HPA axis to a single bout of fasting is dependent on functional MC3Rs, however the response to negative energy balance associated with prolonged bouts of restricted feeding is maintained12. While our data are consistent with Renquist’s data, it differs in suggesting that a component of the stress response is attenuated in Mc3r-deficient mice. The timing of a meal-entrained stress response, indicated by peaks in expression of stress-responsive genes, coinciding with nutrient consumption is dependent on functional MC3Rs. This response is likely indirect and may involve a circadian component. Rhythms in the expression of core elements and circadian oscillator output genes that show entrainment in control mice in the cerebellum do not respond in Mc3r-deficient mice (Suppl. Fig. 2).
In summary, the results of these experiments further support the crucial role of MC3R signaling in the expression of homeostatic adaptive behavioral responses to hypocaloric conditioning. Rescuing Mc3r expression in the hypothalamic and limbic structures improves appetitive responses to internal cues of metabolic condition, but at best has a minor inhibitory effect on adiposity. The primary site(s) and mechanisms of action of MC3Rs involved in nutrient partitioning therefore remain unclear. One interpretation of the results from studies of the hypothalamic and behavior responses to HFS is that critical fasting-responsive melanocortin neurons in the hypothalamus of Mc3r-deficient mice are at least partially uncoupled from signals of metabolic condition. However, caution must be applied as this interpretation is based exclusively on gene expression data that may not necessarily correlate with activity of ARC neurons48. The basis of the behavioral phenotype could involve ‘active’ regulatory functions of MC3Rs in setting the gain in the response of fasting-responsive neurons to signals of metabolic condition. Alternatively, MC3R signaling in Nkx2.1(+ve) neurons may be crucial for the development and maturation of hypothalamic circuitry involved in appetite control. When combined with recently published data14, MC3Rs expressed in hypothalamic and limbic neurons overall are suggested to have functions that promote feeding behavior and maintain behavioral flexibility in response to internal cues that control incentive salience and motivational state. Finally, the current studies further dissociate the paradoxical impact of Mc3r-deficiency to reduce feeding behavior while regulating nutrient partitioning to favor the development of an obese phenotype.
Methods
Animal husbandry
All of the experiments involving mice were performed in accordance to the guidelines and regulations provided by the Institutional Animal Care and Use Committees of the Scripps Research Institute and Saint Louis University School of Medicine, which reviewed and approved the studies.
The mice used in these studies were acclimated to single-housing at a constant temperature of 23C with food and water provided ad libitum under a 12:12 light-dark cycle, unless otherwise specified. In B6(Cg)-Mc3rtm1Butl/J (Mc3rTB/TB) mice, Mc3r transcription is inhibited by a loxP-flanked transcriptional blocker; these mice are primarily on the C57BL/6J background with a small contribution from/6N. The percentage of genes derived from C57BL/6J and/6N in each of our experimental mice is presently unknown. Mc3r transcription was rescued throughout the CNS (NES-MC3R mice) using B6.Cg-Tg(Nes-cre)1Kln/J (nes-cre)60 backcrossed on the Mc3rTB/TB background. Mc3r transcription was rescued using Nkx2.1-Cre (C57BL/6J-Tg(Nkx2–1-cre)2Sand/J) (NKX-MC3R mice)28.
B6.Cg-Gt(ROSA)26Sortm14(CAG-tdTomato)Hze/J was crossed with Nkx2.1-Cre to obtain Nkx2.1Cre;ROSA-tdTomato mice in order to visualize recombination pattern at the level of the mediobasal hypothalamus. All mice studied were littermates obtained from heterozygous breeding. Cre genotype was kept to heterozygosity and carried by the male for breeding purpose. Genotyping PCR using tail-tip DNA was used to assess germline recombination. Animals showing Cre-mediated recombination in the tail were then removed from the study. Note that for experiment involving Cre-mediated excision, Cre-positive mice were also studied in order to evaluate and account for the effect of Cre expression itself.
For consistency with previous experiments7,8,9,10,11,14,61,62, we used a refined low fat/high carbohydrate diet (LFD, 10% kcal from fat, 20% kcal/protein and 70% kcal/carbohydrates, D12450B; Research diets Inc., New Brunswick, NJ). Nuclear magnetic resonance (NMR, Bruker Minispec) was used to measure fat mass (FM), fat-free mass (FFM) and free H2O14.
Hypocaloric conditioning protocol
To assess circadian- and feeding-related adaptation to scheduled feeding, a simple protocol was used. A single meal, amounting to 70–75% of normal habitual food intake determined empirically, was provided daily at ZT4 (ZT, zeitgeber time; ZT 0 and ZT12 represent times of dark/light and light/dark transition). After 4 hours, the amount of food of remained was weighed on days 2, 4 and 6 to estimate consumption. Any remaining food was returned to the cage.
Neuroendocrine and hypothalamic responses were examined using adult (12–14wk) male wild type (WT, n = 77) and Mc3rTB/TB (n = 79) mice euthanized at 4 h intervals throughout the day with the HFS group receiving food. Red lights were used as needed to avoid disrupting daily rhythms.
Contribution of hypothalamic MC3Rs to hypothalamic responses was investigated in adult female mice (n = 4–6) euthanized at ZT3.
Assessment of food intake and Meal pattern analysis
Meal structure was examined using an automated system for continuous monitoring of food consumption (BiodaQ 2.3, Research Diets Inc., New Brunswick, NJ) and BiodaQ 2.3 software. ‘Bouts’ indicate disturbance of the hopper and instability in scale readings suggesting approach and investigation; actual changes in food weight were used to estimate meal size. Meals were defined as bouts occurring within 5 minutes of each other and consumption of ≥0.02 g.
Mice acclimated to single housing on bedding with no caloric value (aspen chips) and LFD for 3 wk were transferred to BiodaQ cages. After 10d of acclimation, baseline feeding behavior was established using 2d of recordings before food deprivation.
Adult male mice (8–10 wk) were then subjected to HFS for 6d with food provided at ZT4 with water provided ad libitum.
Nkx2.1-Cre male and female mice (20 wk) were then submitted to a timed food restriction protocol with food access granted access for 4 h (from ZT4 to ZT8)9.
Operant conditioning behavior
Self-administration experiments were performed as previously described using operant chambers (Coulbourn Instruments)14. Briefly, mice were trained in operant chambers housed in sound-attenuating cubicles (Coulbourn Instruments). Each chamber was equipped with two levers (predetermined to be “active” or “inactive”), a food pellet hopper and a house light. Prior to training, 3.5–4 months old male mice were subjected to 14 days of caloric restriction in order to decrease body weight to 85–90% of normal. Weight loss was then maintained throughout the study by feeding the mice a hypocaloric meal after the end of the training session. Mice were first habituated to the chambers with chocolate flavored food pellets (sucrose; Dustless Precision Pellets 20 mg, Bioserv) dropping every minute concomitantly with the activation of a light cue over the active lever. Following two days on this “magazine” schedule, mice were trained for 7 days in a Fixed Ratio 1 (FR1) schedule; each “active” lever pressing resulted in the delivery of a sucrose pellet and the activation of the light cue. Pressing the inactive lever had no outcome. The training session lasted 60 min with a 1 sec time out period following the delivery of the pellet. Two cohorts of WT and Mc3rTB/TB male mice maintained either under a normal 12 h light:12 h dark cycle or an inverted 12 h dark:12 h light cycle were studied respectively during their inactive (light phase) and active (dark phase) period. Note that all manipulation of the cohort maintained under the inverted light-dark cycle was performed under red light.
Assessment of locomotor activity
FAA was assessed using mice housed in cages with free access to running wheels as previously described10. Briefly, after a 1 wk acclimation period to wheel-equipped cages in a 12:12 light-dark (LD), food was measured 3 times over a 1 week period before all food was removed and mice were submitted to HFS protocol. Wheel running data was analyzed using Clocklab software (Actimetrics, Evanston, IL).
Blood chemistries
Trunk blood was collected in the presence of AEBSF (1 mg/ml, Sigma, St Louis, MO). Plasma was collected after centrifugation (15 min, 2000 g, 4C). To measure acyl-ghrelin, a portion of plasma was acidified with HCl (0.05N). Mouse ELISA kits were used to measure acyl-ghrelin (Millipore, Billerica, MA), leptin (Crystal Chem, Downers Grove, IL) and corticosterone (CORT; Enzo Life Sciences Inc., Farmingdale, NY); intra- and the inter-assay CV were < 5%.
Drug treatment
Mice acclimated to single-housing were injected with dexamethasone 21-phosphate disodium salt (10 mg/kg ip., D1159, Sigma, St Louis, MO) or saline at ZT3 and returned to home cages for 4 h before sacrifice.
Gene expression analysis
The hypothalamus, medial habenula (MHab) and cerebellum were dissected from frozen brains on dry ice and total RNA extracted using Trizol (Invitrogen, Life Technologies). Total RNA was treated with a DNAfree kit (Ambion, Life Technologies) and 800 ng used for cDNA synthesis (Superscript III Reverse transcription kit, Invitrogen). Quantitative PCR was performed in 384-well plates using Taqman gene expression and QuantStudio 6 or 7 Detection Systems (Applied Biosystems, Life Technologies) with Tbp as the reference gene.
Statistical analysis
All data are presented as mean ± SEM. Statistical analyses were performed using GraphPad Prism 6 and IBM SPSS Statistics software. The effect of genotype of body composition was assessed by ANCOVA with total body mass as a covariate; FM, FFM and free H2O are presented as estimated marginal means adjusted for total body mass unless stated otherwise63,64. Differences between two genotypes were assessed by unpaired t test. For feeding studies, effects of Mc3r genotype and time were analyzed using 2-way ANOVA with repeated measures followed by Bonferroni’s post hoc test. In experiments examining the effect of feeding and genotype on 24 h rhythms, multi-factor ANOVA were performed with feeding (HFS or ad libitum), genotype and time as independent variables. Spearman correlation was used to evaluate relationships between neuropeptide expression and hormonal levels. A p value < 0.05 was considered significant.
Additional Information
How to cite this article: Girardet, C. et al. Melanocortin-3 receptors expressed in Nkx2.1(+ve) neurons are sufficient for controlling appetitive responses to hypocaloric conditioning. Sci. Rep. 7, 44444; doi: 10.1038/srep44444 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Rosenbaum, M., Kissileff, H. R., Mayer, L. E., Hirsch, J. & Leibel, R. L. Energy intake in weight-reduced humans. Brain research 1350, 95–102 (2010).
Maclean, P. S., Bergouignan, A., Cornier, M. A. & Jackman, M. R. Biology’s response to dieting: the impetus for weight regain. American journal of physiology . Regulatory, integrative and comparative physiology 301, R581–600 (2011).
Leibel, R. L. et al. Biologic Responses to Weight Loss and Weight Regain: Report From an American Diabetes Association Research Symposium. Diabetes 64, 2299–2309 (2015).
Ravussin, E. & Ryan, D. H. Energy expenditure and weight control: Is the biggest loser the best loser? Obesity 24, 1607–1608 (2016).
Cone, R. D. Anatomy and regulation of the central melanocortin system. Nature neuroscience 8, 571–578 (2005).
Krashes, M. J., Lowell, B. B. & Garfield, A. S. Melanocortin-4 receptor-regulated energy homeostasis. Nature neuroscience 19, 206–219 (2016).
Sutton, G. M. et al. The melanocortin-3 receptor is required for entrainment to meal intake. The Journal of neuroscience: the official journal of the Society for Neuroscience 28, 12946–12955 (2008).
Begriche, K. et al. Genetic dissection of the functions of the melanocortin-3 receptor, a seven-transmembrane G-protein-coupled receptor, suggests roles for central and peripheral receptors in energy homeostasis. The Journal of biological chemistry 286, 40771–40781 (2011).
Begriche, K. et al. Melanocortin-3 receptors are involved in adaptation to restricted feeding. Genes, brain, and behavior 11, 291–302 (2012).
Girardet, C., Mavrikaki, M., Southern, M. R., Smith, R. G. & Butler, A. A. Assessing interactions between Ghsr and Mc3r reveals a role for AgRP in the expression of food anticipatory activity in male mice. Endocrinology 155, 4843–4855 (2014).
Begriche, K., Sutton, G. M. & Butler, A. A. Homeostastic and non-homeostatic functions of melanocortin-3 receptors in the control of energy balance and metabolism. Physiology & behavior 104, 546–554 (2011).
Renquist, B. J. et al. Melanocortin-3 receptor regulates the normal fasting response. Proceedings of the National Academy of Sciences of the United States of America 109, E1489–1498 (2012).
Lippert, R. N., Ellacott, K. L. & Cone, R. D. Gender-specific roles for the melanocortin-3 receptor in the regulation of the mesolimbic dopamine system in mice. Endocrinology 155, 1718–1727 (2014).
Mavrikaki, M. et al. Melanocortin-3 receptors in the limbic system mediate feeding-related motivational responses during weight loss. Molecular metabolism 5, 566–579 (2016).
Lee, B. et al. A mouse model for a partially inactive obesity-associated human MC3R variant. Nature communications 7, 10522 (2016).
Savastano, D. M. et al. Energy intake and energy expenditure among children with polymorphisms of the melanocortin-3 receptor. The American journal of clinical nutrition 90, 912–920 (2009).
Aris, I. M. et al. MC3R gene polymorphisms are associated with early childhood adiposity gain and infant appetite in an Asian population. Pediatric obesity (2015).
Obregon, A. M. et al. Melanocortin-3 receptor gene variants: association with childhood obesity and eating behavior in Chilean families. Nutrition 26, 760–765 (2010).
Patton, D. F. & Mistlberger, R. E. Circadian adaptations to meal timing: neuroendocrine mechanisms. Frontiers in neuroscience 7, 185 (2013).
Luquet, S., Perez, F. A., Hnasko, T. S. & Palmiter, R. D. NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science 310, 683–685 (2005).
Gropp, E. et al. Agouti-related peptide-expressing neurons are mandatory for feeding. Nature neuroscience 8, 1289–1291 (2005).
Bewick, G. A. et al. Post-embryonic ablation of AgRP neurons in mice leads to a lean, hypophagic phenotype. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 19, 1680–1682 (2005).
Tan, K., Knight, Z. A. & Friedman, J. M. Ablation of AgRP neurons impairs adaption to restricted feeding. Molecular metabolism 3, 694–704 (2014).
Aponte, Y., Atasoy, D. & Sternson, S. M. AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nature neuroscience 14, 351–355 (2011).
Krashes, M. J. et al. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. The Journal of clinical investigation 121, 1424–1428 (2011).
Krashes, M. J., Shah, B. P., Koda, S. & Lowell, B. B. Rapid versus delayed stimulation of feeding by the endogenously released AgRP neuron mediators GABA, NPY, and AgRP. Cell metabolism 18, 588–595 (2013).
Roselli-Rehfuss, L. et al. Identification of a receptor for gamma melanotropin and other proopiomelanocortin peptides in the hypothalamus and limbic system. Proceedings of the National Academy of Sciences of the United States of America 90, 8856–8860 (1993).
Xu, Q., Tam, M. & Anderson, S. A. Fate mapping Nkx2.1-lineage cells in the mouse telencephalon. The Journal of comparative neurology 506, 16–29 (2008).
Yee, C. L., Wang, Y., Anderson, S., Ekker, M. & Rubenstein, J. L. Arcuate nucleus expression of NKX2.1 and DLX and lineages expressing these transcription factors in neuropeptide Y(+), proopiomelanocortin(+), and tyrosine hydroxylase(+) neurons in neonatal and adult mice. The Journal of comparative neurology 517, 37–50 (2009).
Kimura, S. et al. The T/ebp null mouse: thyroid-specific enhancer-binding protein is essential for the organogenesis of the thyroid, lung, ventral forebrain, and pituitary. Genes & development 10, 60–69 (1996).
Takuma, N. et al. Formation of Rathke’s pouch requires dual induction from the diencephalon. Development 125, 4835–4840 (1998).
Ring, L. E. & Zeltser, L. M. Disruption of hypothalamic leptin signaling in mice leads to early-onset obesity, but physiological adaptations in mature animals stabilize adiposity levels. The Journal of clinical investigation 120, 2931–2941 (2010).
Chong, A. C., Greendyk, R. A. & Zeltser, L. M. Distinct networks of leptin- and insulin-sensing neurons regulate thermogenic responses to nutritional and cold challenges. Diabetes 64, 137–146 (2015).
Chong, A. C., Vogt, M. C., Hill, A. S., Bruning, J. C. & Zeltser, L. M. Central insulin signaling modulates hypothalamus-pituitary-adrenal axis responsiveness. Molecular metabolism 4, 83–92 (2015).
Butler, A. A. et al. A unique metabolic syndrome causes obesity in the melanocortin-3 receptor-deficient mouse. Endocrinology 141, 3518–3521 (2000).
Butler, A. A. The melanocortin system and energy balance. Peptides 27, 281–290 (2006).
Sutton, G. M. et al. Diet-genotype interactions in the development of the obese, insulin-resistant phenotype of C57BL/6J mice lacking melanocortin-3 or -4 receptors. Endocrinology 147, 2183–2196 (2006).
Chen, A. S. et al. Inactivation of the mouse melanocortin-3 receptor results in increased fat mass and reduced lean body mass. Nature genetics 26, 97–102 (2000).
Guarnieri, D. J. et al. Gene profiling reveals a role for stress hormones in the molecular and behavioral response to food restriction. Biological psychiatry 71, 358–365 (2012).
Dibner, C., Schibler, U. & Albrecht, U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol 72, 517–549 (2010).
Mendoza, J., Pevet, P., Felder-Schmittbuhl, M. P., Bailly, Y. & Challet, E. The cerebellum harbors a circadian oscillator involved in food anticipation. The Journal of neuroscience: the official journal of the Society for Neuroscience 30, 1894–1904 (2010).
Harno, E., Cottrell, E. C. & White, A. Metabolic pitfalls of CNS Cre-based technology. Cell metabolism 18, 21–28 (2013).
Woods, S. C. The eating paradox: how we tolerate food. Psychological review 98, 488–505 (1991).
Bruss, M. D., Khambatta, C. F., Ruby, M. A., Aggarwal, I. & Hellerstein, M. K. Calorie restriction increases fatty acid synthesis and whole body fat oxidation rates. American journal of physiology. Endocrinology and metabolism 298, E108–116 (2010).
Krashes, M. J. et al. An excitatory paraventricular nucleus to AgRP neuron circuit that drives hunger. Nature 507, 238–242 (2014).
Sternson, S. M., Atasoy, D., Betley, J. N., Henry, F. E. & Xu, S. An Emerging Technology Framework for the Neurobiology of Appetite. Cell metabolism 23, 234–253 (2016).
Gautron, L., Elmquist, J. K. & Williams, K. W. Neural control of energy balance: translating circuits to therapies. Cell 161, 133–145 (2015).
Pedroso, J. A. et al. Changes in leptin signaling by SOCS3 modulate fasting-induced hyperphagia and weight regain in mice. Endocrinology. en20161038 (2016).
Tschop, M. H., Castaneda, T. R. & Woods, S. C. The brain is getting ready for dinner. Cell metabolism 4, 257–258 (2006).
Vahl, T. P., Drazen, D. L., Seeley, R. J., D’Alessio, D. A. & Woods, S. C. Meal-anticipatory glucagon-like peptide-1 secretion in rats. Endocrinology 151, 569–575 (2010).
Dailey, M. J., Stingl, K. C. & Moran, T. H. Disassociation between preprandial gut peptide release and food-anticipatory activity. Endocrinology 153, 132–142 (2012).
LeSauter, J., Hoque, N., Weintraub, M., Pfaff, D. W. & Silver, R. Stomach ghrelin-secreting cells as food-entrainable circadian clocks. Proceedings of the National Academy of Sciences of the United States of America 106, 13582–13587 (2009).
Trevaskis, J. L. et al. Role of adiponectin and inflammation in insulin resistance of Mc3r and Mc4r knockout mice. Obesity 15, 2664–2672 (2007).
Oster, H. et al. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell metabolism 4, 163–173 (2006).
Damiola, F. et al. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes & development 14, 2950–2961 (2000).
Honma, K. I., Honma, S. & Hiroshige, T. Critical role of food amount for prefeeding corticosterone peak in rats. The American journal of physiology 245, R339–344 (1983).
Garcia-Belenguer, S., Oliver, C. & Mormede, P. Facilitation and feedback in the hypothalamo-pituitary-adrenal axis during food restriction in rats. Journal of neuroendocrinology 5, 663–668 (1993).
Le Minh, N., Damiola, F., Tronche, F., Schutz, G. & Schibler, U. Glucocorticoid hormones inhibit food-induced phase-shifting of peripheral circadian oscillators. The EMBO journal 20, 7128–7136 (2001).
Sheward, W. J. et al. Entrainment to feeding but not to light: circadian phenotype of VPAC2 receptor-null mice. The Journal of neuroscience: the official journal of the Society for Neuroscience 27, 4351–4358 (2007).
Tronche, F. et al. Disruption of the glucocorticoid receptor gene in the nervous system results in reduced anxiety. Nature genetics 23, 99–103 (1999).
Sutton, G. M. et al. Central nervous system melanocortin-3 receptors are required for synchronizing metabolism during entrainment to restricted feeding during the light cycle. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 24, 862–872 (2010).
Girardet, C., Begriche, K., Ptitsyn, A., Koza, R. A. & Butler, A. A. Unravelling the mysterious roles of melanocortin-3 receptors in metabolic homeostasis and obesity using mouse genetics. Int J Obes (Lond) Supplement 4, S37–S44 (2014).
Allison, D. B., Paultre, F., Goran, M. I., Poehlman, E. T. & Heymsfield, S. B. Statistical considerations regarding the use of ratios to adjust data. International journal of obesity and related metabolic disorders: journal of the International Association for the Study of Obesity 19, 644–652 (1995).
Packard, G. C. & Boardman, T. J. The Misuse of Ratios, Indices, and Percentages in Ecophysiological Research. Physiological Zoology 61, 1–9 (1988).
Acknowledgements
We thank Jillian Krings (Saint Louis University) and Peter R. Levasseur (Papé Family Pediatric Research Institute) for outstanding technical assistance for these experiments. The authors acknowledge the editorial assistance provided by Prof. Terrance Egan, Department of Pharmacology & Physiology, Saint Louis University School of Medicine, during manuscript preparation. We also thank the Vivarium staff from TSRI-Florida and Comparative Medicine staff from St Louis University for excellent animal husbandry as well as their help during the course of these studies. Funding for this work was provided by the National Institutes of Health (DK073189 to A.A.B.).
Author information
Authors and Affiliations
Contributions
C.G., M.M. and A.B. designed experiments. C.G., M.M., J.S. and D.M. researched data. C.M. and A.B. provided research materials; C.A.M. also provided discussions on behavioral phenotyping of mice. C.G. and A.B. wrote the first draft of the manuscript; M.M. and D.M. reviewed and edited the manuscript. C.G. and A.B. are the guarantors of this work and, as such, had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Girardet, C., Mavrikaki, M., Stevens, J. et al. Melanocortin-3 receptors expressed in Nkx2.1(+ve) neurons are sufficient for controlling appetitive responses to hypocaloric conditioning. Sci Rep 7, 44444 (2017). https://doi.org/10.1038/srep44444
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep44444
- Springer Nature Limited