
Abstract
Background
The sexual dimorphism represents one of the triggers of the metabolic disparities while the identification of sex-specific metabolites in the elderly has not been achieved.
Methods
A group of aged healthy population from Southwest China were recruited and clinical characteristics were collected. Fasting plasma samples were obtained and untargeted liquid chromatography-mass spectrometry-based metabolomic analyses were performed. Differentially expressed metabolites between males and females were identified from the metabolomic analysis and metabolite sets enrichment analysis was employed.
Results
Sixteen males and fifteen females were finally enrolled. According to clinical characteristics, no significant differences can be found except for smoking history. There were thirty-six differentially expressed metabolites between different sexes, most of which were lipids and lipid-like molecules. Twenty-three metabolites of males were increased while thirteen were decreased compared with females. The top four classes of metabolites were fatty acids and conjugates (30.6%), glycerophosphocholines (22.2%), sphingomyelins (11.1%), and flavonoids (8.3%). Fatty acids and conjugates, glycerophosphocholines, and sphingomyelins were significantly enriched in metabolite sets enrichment analysis.
Conclusions
Significant lipid metabolic differences were found between males and females among the elderly. Fatty acids and conjugates, glycerophosphocholines, and sphingomyelins may partly account for sex differences and can be potential treatment targets for sex-specific diseases.
Similar content being viewed by others

Background
Sexual dimorphism is a common biological phenomenon in the nature. Robust differences are found between males and females in disease incidence, disease severity, metabolism, and pharmacodynamics of interventions. The prevalence rates of coronary heart disease, heart failure, stroke, and various metabolic syndromes are significantly higher among the males while the occurrence and severity of knee osteoarthritis are also influenced by sex, with older females affected to a greater degree by the disease compared to age-matched males [1]. Sex difference is also found in lipid and cholesterol metabolism. At homeostasis, the female is prone to incorporate free fatty acids into triglycerides whereas the male likely oxidizes circulating free fatty acids [2]. It is reported that sexual dimorphism is due to sex chromosome and the following sex-specific hormone action.
Recognition and identification of sex differences are important for researchers to develop new treatments and physicians to deal with sex-specific diseases. Metabolomic is a systematic analysis of all the metabolites in a biological sample. The metabolites identified by metabolomic include amino acids, peptides, oligonucleotides, carbohydrates, organic acids, ketones, aldehydes, lipids, steroids, alkaloids, xenobiotics, and any other small molecules deriving from biological processes. The plasma metabolome of healthy individuals has already been analyzed and it is reported that sex differences are mainly correlated with amino acids and acylcarnitines, including creatine [3]. Sex can also affect the metabolome of biological fluids in an age-dependent way and previous publications identified the interaction between sex and age [4]. However, little publications emphasized aged population, as metabolism changes following the sex-specific hormones in postmenopausal females. Furthermore, the metabolic fingerprint of sex differences also varies between different races [5]. The published researches are mainly from western countries including Caucasian, African-American, Hispanic, and so on [6].
Herein, we adopted untargeted liquid chromatography-mass spectrometry (LC–MS)-based metabolomics to analyze sex-specific metabolic changes in a Chinese aged population. Plasma is chosen in the metabolomic analysis as it is a relatively accessible, stable, and informative biofluid.
Methods
Participants
A group of aged healthy population were recruited in Department of Physical Examination, the First Affiliated Hospital of Chongqing Medical University, from April 2016 to February 2017. All the participants were more than 50 years old and all the included females should be postmenopausal. The participants were without any acute illness or in acute state of chronic diseases at the enrollment.
This study was approved by the ethics committee of the First Affiliated Hospital of Chongqing Medical University and performed in accordance with Declaration of Helsinki. Statements of informed consent were obtained from all the participants prior to inclusion in this study. Clinical characteristics and metabolomic analysis were blindly collected or performed, separately [7].
Clinical characteristics
Clinical characteristics of all the participants were collected, including age, smoking history, alcohol consumption, hypertension, diabetes mellitus, hypercholesterolemia, and coronary heart disease. Fasting plasma samples were obtained by puncture of the median cubital vein at 6:00 am. The levels of total cholesterol, triglyceride, high-density lipoprotein cholesterol, low-density lipoprotein cholesterol, apolipoprotein A1, apolipoprotein B, and lipoprotein a were also determined using a Cobas Integra 400 plus automatic biochemical analyzer with matched reagent kits (Roche, Basel, Switzerland) [8].
Metabolomic analysis
The detailed procedure of metabolomic analysis was described in the former research [9]. Firstly, plasma samples stored at -80℃ were gradually thawed on ice, 2-chloro-1-phenylalanine dissolved in methanol (0.3 mg/mL) was served as internal standard. In a 1.5 mL Eppendorf tube, 50μL sample and 10μL internal standard were added and then vortexed for 10 s. Subsequently, 150μL ice-cold mixture of methanol and acetonitrile (2/1, vol/vol) were added. The mixtures were vortexed for 1 min, ultrasonicated at ambient temperature (25℃) for 5 min, placed at -20℃ for 10 min, and centrifuged at 15000 rpm at 4℃ for 10 min. 100μL of the supernatants from each tube were collected, filtered through 0.22 μm microfilters, and transferred to LC vials. The vials were stored at -80℃ until LC–MS analysis. Quality control sample was obtained by mixing all the samples equally as a pooled sample, and then processed using the above method with the analytic reagents. Fifteen quality controls and the whole samples were randomly injected throughout the analytical run to provide a set of data from which repeatability can be assessed.
We adopted a Waters UPLC I-class system equipped with a binary solvent delivery manager and a sample manager, coupled with a Waters VION IMS Q-TOF Mass Spectrometer equipped with an electrospray interface (Waters Corporation, Milford, USA) to perform the untargeted LC–MS metabolomics. Acquity BEH C18 column (100 mm × 2.1 mm i.d., 1.7 μm; Waters Corporation) was used and the column temperature was maintained at 45℃. The separation process was achieved using the following gradient: 5% B—25% B over 1–1.5 min, 25% B -100% B over 1.5–10.0 min, 100% B – 100% B over 10.0 – 13.0 min; 100% B – 5% B over 13.0 – 13.5 min, and 13.5 – 14.5 min holding at 5% B at a flow rate of 0.4 mL/min, where B is acetonitrile (0.1% (vol/vol) formic acid) and A is aqueous formic acid (0.1% (vol/vol) formic acid). Injection volume was 3μL.
The mass spectrometric data was collected using the Waters mass spectrometer operating in either positive or negative ion mode. The source temperature and desolvation temperature was set at 120℃ and 500℃, respectively, with a desolvation gas flow of 900 L/h. Centroid data was collected from 50 to 1,000 m/z with a scan time of 0.1 s and interscan delay of 0.02 s over a 13 min analysis time. Centroid data was collected from 50 to 1,000 m/z with a scan time of 0.1 s and interscan delay of 0.02 s over a 13 min analysis time. The obtained data were processed by baseline filtering, peak identification, integration, retention time correction, peak alignment and normalization using the build-in metabolomic software Progenesis QI (Waters Corporation). Retention time ranged from 0.5 to 14.0 min, mass ranged from 50 to 1000 Da, and mass tolerance was 0.01 Da. Isotopic peaks were excluded for analysis, noise elimination level was set at 10.00, minimum intensity was set to 15% of base peak intensity, and retention time tolerance was set at 0.01 min.
After that, data sets including m/z, peak retention time, and peak intensity of each ion were obtained, and further reduced by removing any peaks with missing values in more than 60% of the total samples. The internal standard was used for data quality control. Metabolite identification was performed based on the following two steps. First, we used our self-constructed metabolite databank, which contains chemical standards and a manually curated compound list based on accurate mass (m/z, ± 5 ppm), retention time, and spectral patterns. Second, further metabolites were identified based on accurate mass, isotope pattern and MS/MS spectra against public databanks, including Metlin (https://metlin.scripps.edu), Human Metabolome Database (HMDB, http://www.hmdb.ca) and so on. The peak intensity was deemed as expression level of a metabolite [10].
The positive and negative peak data were merged and multivariate statistical analyses were performed by the SIMCA-P 13.0 software package (Umetrics, Umea, Sweden). The quality control samples were used to validate the stability of the metabolomic analysis. The unsupervised principal component analysis (PCA) was used to observe the data distribution. The orthogonal partial least squares-discriminant analysis (OPLS-DA) model with sevenfold cross validation was constructed to show statistical differences and recognize differentially expressed metabolites between the two groups. The constructed model was validated by a response permutation test with 200 iterations. Metabolites with variable influence on projection values (obtained from the OPLS-DA model) of greater than 1.0, fold change values of greater than 1.5 or lower than 0.67, and p values (obtained from Student t test) of less than 0.05 were recognized to be differentially expressed. Partial least squares-discriminant analysis (PLS-DA) with sevenfold cross validation was further constructed based on the above differentially expressed metabolites to visualize their differential ability between the male and females. The differentially expressed metabolites and their quantities were then exhibited as clustering heatmap. Metabolite sets enrichment analysis was further performed based on the above metabolites using MetaboAnalyst 5.0 (metaboanalyst.ca) [11].
Statistical analysis
Statistical analyses were completed using a commercially available software package (IBM SPSS version 22.0, New York, USA). Continuous data were expressed as means ± standard deviation and compared using Student t tests. Categorical data were exhibited as absolute numbers and percentage (%), and analyzed using Pearson χ2-tests or Fisher exact tests. P values less than 0.05 were considered as statistical significances [12].
Results
Clinical characteristics
A total of sixteen males and fifteen females were finally included in this study. No significant difference can be found in the mean age of these participants (62.56 ± 2.43 versus 65.73 ± 1.96). The males had a significant higher rate of smoking history compared with the females. There were no significant differences in the other clinical characteristics, indicating the two groups of participants were comparable. No statistical significances were found in total cholesterol, triglyceride, high-density lipoprotein cholesterol, low-density lipoprotein cholesterol, apolipoprotein A1, apolipoprotein B, and lipoprotein a (Table 1).
Metabolomic analysis
After excluding internal standards, a total of 10,403 individual peaks, including 6040 positive and 4363 negative peaks, were detected in approximately 98.8% of samples in each group. Based on these peaks, score plots from PCA and OPLS-DA analysis were performed and the results showed separations between the two groups (R2X = 0.129, R2Y = 0.855, Q2 = 0.084) (Fig. 1).

Multivariate statistical analyses of the liquid chromatography-mass spectrometry-based metabolomics between males and females included in this study. A PCA score plot of all the participants included seven components with a R2X value of 0.661 and a Q2 value of 0.219. B OPLS-DA score plot of all the participants included two components with a R2X value of 0.129, R2Y value of 0.855, and Q2 value of 0.084. PCA, principal component analysis; OPLS-DA, orthogonal partial least squares-discriminant analysis
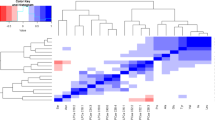
There were thirty-six differentially expressed metabolites between the two sexes, most of which were lipids and lipid-like molecules. Twenty-three metabolites were increased in the males while thirteen were decreased (Table 2). The differentially expressed metabolites and the corresponding quantities of each sample were exhibited in clustering heatmap (Fig. 2). PLS-DA score plot indicated clear separations between two sexes based on the above differentially expressed metabolites. The score plot included two components with a R2X value of 0.516, R2Y value of 0.748, and Q2 value of 0.65, indicating the model was stable and reliable. Meanwhile, the corresponding response permutation test indicated the PLS-DA model was not over-fitting (R2 = (0.0, 0.311), Q2 = (0.0, -0.237)) (Fig. 3).

Clustering heatmap of differentially expressed metabolites between males and females

PLS-DA score plot and its corresponding response permutation test based on the differentially expressed metabolites between males and females in this study. A PLS-DA score plot of all the participants included two components with a R2X value of 0.516, R2Y value of 0.748, and Q2 value of 0.65, indicating the model was stable and reliable. B Response permutation test indicated the constructed PLS-DA model was not over-fitting (R2 = (0.0, 0.311), Q2 = (0.0, -0.237)). PLS-DA, partial least squares-discriminant analysis
The differentially expressed metabolites can be categorized into fatty acids and conjugates, glycerophosphocholines (GPCs), sphingomyelins, macrolides and analogues, steroid conjugates, flavonoids, fatty amides, octadecanoids, docosanoids, glycerophosphoserines, glycerophosphoinositols, glycerophospholipids, glycerolipids, and so on. The top four main classes of metabolites were fatty acids and conjugates (11, 30.6%), GPCs (8, 22.2%), sphingomyelins (4, 11.1%), and flavonoids (3, 8.3%) (Fig. 4). According to the metabolite sets enrichment analysis, fatty acids and conjugates, GPCs, and sphingomyelins were significantly enriched in the over-representation analysis (Table 3).

Pie chart depicting the classification of differentially expressed metabolites
Discussion
Lipid metabolic disturbances are found in various diseases, including metabolic syndrome, cardiovascular disease, heart failure, cerebrovascular disease, and Guillain–Barre syndrome [13]. Lipid metabolism is correlated with Parkinson’s disease and related neuropsychiatric symptoms according to our former research [14]. The occurrence and development of diabetic peripheral neuropathy can be delayed by regulating lipid metabolism [15]. Sex differences were also found in very-low-density lipoproteins triglyceride and low-density lipoprotein cholesterol with age dependence [1]. We were the first to perform LC–MS-based metabolomics in Chinese aged population to clarify sex differences and found some specific lipid changes as follows.
Fatty acids and conjugates
The group of fatty acids and conjugates is a subclass of fatty acyls, which is among the eight categories of lipids in current classification system. It is associated with the occurrence and development of many diseases. The plasma levels of long-chain omega-3 and long-chain omega-6 fatty acids were associated with a lower risk of schizophrenia while short-chain omega-3 and short-chain omega-6 fatty acids were associated with an increased risk of schizophrenia [16]. The supplementation of omega-3 fatty acid can reduce major adverse cardiovascular events, cardiovascular death, and myocardial infarction [17]. It is reported that in tambaqui (Colossoma macropomum) elongation of very long-chain fatty acids enzymes and fatty acid metabolism played important roles in sexual differentiation [18]. A former gas-chromatography mass-spectrometry-based metabolomic analysis with urine samples from healthy males and females indicated that saturated fatty acids were significantly correlated with sex [19]. In this research, eleven differentially expressed fatty acids and conjugates were found and nine were significantly increased in males. These nine metabolites varied in molecular structures without obvious characteristics, including dicarboxylic acid, aromatic compound, monounsaturated fatty acid, polyunsaturated fatty acid, and the carbon numbers of those molecules ranged from eight to twenty-three. The sex-specific expression mode of fatty acids and conjugates may contribute to the differential incidences of schizophrenia and cerebral vascular diseases in males and females.
Glycerophosphocholines
GPCs are glycerophospholipids in which a phosphorylcholine moiety occupies a glycerol substitution site. GPCs are mainly exogenous and abundant in egg, soybean, beef, shrimp, and any other foods. The preferences and consumption structures of food between the two sexes lead to the different content levels of GPCs. GPCs can cross the blood–brain barrier and serve as contributors of choline and phospholipid in central nervous system. GPCs are reportedly involved in depression, anxiety, dementia, and many other neurological disorders. They can be hydrolyzed by the enzyme phospholipase A2 into lysophosphatidylcholines (LysoPCs) while lysoPCs can specifically bind to the G protein-coupled receptor family (GPR119, GPR40, GPR55 and GPR4), and induce intracellular calcium mobilization leading to increased glucose-stimulated insulin secretion. LysoPCs also have several protective or anti-inflammatory effects and serve as dual-activity ligand molecules in the innate immune system [20]. A total of eight GPCs had been found differentially expressed between males and females. The contents of LysoPCs were significantly higher in males while the lipid chains of males were significantly longer than females.
Sphingomyelins
Sphingomyelins help form lipid rafts in cell membranes and are involved in signal transduction and transportation of lipids and proteins [21]. It can be hydrolyzed by sphingomyelinases into ceramides, which are important second messengers in cell proliferation, differentiation, proliferation, and apoptosis [22]. The balance of sphingomyelins is essential for normal neuronal function and the deficiencies in enzymes of sphingomyelins metabolism can lead to various severe brain disorders. Some researches indicated blood-based sphingomyelins played crucial roles in dementia but the conclusions were inconsistent [23]. A longitudinal cohort provided evidence for sex-specific associations between sphingomyelins and dementia, which might account for the above inconsistence [24]. The changes of plasma sphingomyelins with age were statistically different by sex, and sphingomyelins decreased in males but increased in females with age [25]. Sex differences of sphingomyelins were also found in the development of stress-induced depression [26].
There were several limitations to this study. Firstly, the sample number of included participants was relatively small and further confirmation is needed. Secondly, more experimental methods should be performed to extensively identify sex differences of metabolism in plasma, including gas-chromatography mass-spectrometry and nuclear magnetic resonance profiling.
Conclusions
Significant lipid metabolic differences were found between the two sexes among the elderly. Fatty acids and conjugates, glycerophosphocholines, and sphingomyelins may partly account for sex differences and can be potential treatment targets for sex-specific diseases.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author (Mei-Xue Dong, Email:dong_meixue@whu.edu.cn) on reasonable request.
Abbreviations
- UPLC:
-
Ultra Performance Liquid Chromatography
- PCA:
-
Principal component analysis
- OPLS-DA:
-
The orthogonal partial least squares-discriminant analysis
- GPCs:
-
Glycerophosphocholines
- LysoPCs:
-
Lysophosphatidylcholines
References
Bell JA, Santos Ferreira DL, Fraser A, Soares A, Howe LD, Lawlor DA, et al. Sex differences in systemic metabolites at four life stages: cohort study with repeated metabolomics. Bmc Med. 2021;19(1):58.
Uranga AP, Levine J, Jensen M. Isotope tracer measures of meal fatty acid metabolism: reproducibility and effects of the menstrual cycle. Am J Physiol Endocrinol Metab. 2005;288(3):E547–55.
Costanzo M, Caterino M, Sotgiu G, Ruoppolo M, Franconi F, Campesi I. Sex differences in the human metabolome. Biol Sex Differ. 2022;13(1):30.
Vignoli A, Tenori L, Luchinat C, Saccenti E. Age and sex effects on plasma metabolite association networks in healthy subjects. J Proteome Res. 2018;17(1):97–107.
Lau CE, Siskos AP, Maitre L, Robinson O, Athersuch TJ, Want EJ, et al. Determinants of the urinary and serum metabolome in children from six European populations. Bmc Med. 2018;16(1):202.
Lawton KA, Berger A, Mitchell M, Milgram KE, Evans AM, Guo L, et al. Analysis of the adult human plasma metabolome. Pharmacogenomics. 2008;9(4):383–97.
Dong MX, Hu L, Huang YJ, Xu XM, Liu Y, Wei YD. Cerebrovascular risk factors for patients with cerebral watershed infarction: A case-control study based on computed tomography angiography in a population from Southwest China. Medicine. 2017;96(28): e7505.
Dong MX, Xu XM, Hu L, Liu Y, Huang YJ, Wei YD. Serum Butyrylcholinesterase activity: a biomarker for Parkinson’s disease and related dementia. Biomed Res Int. 2017;2017:1524107.
Hu L, Dong MX, Huang YL, Lu CQ, Qian Q, Zhang CC, et al. Integrated metabolomics and proteomics analysis reveals plasma lipid metabolic disturbance in patients with Parkinson’s disease. Front Mol Neurosci. 2020;13:80.
Liang L, Rasmussen MH, Piening B, Shen X, Chen S, Röst H, et al. Metabolic dynamics and prediction of gestational age and time to delivery in pregnant women. Cell. 2020;181(7):1680–92.
Dong MX, Wei YD, Hu L. Lipid metabolic dysregulation is involved in Parkinson’s disease dementia. Metab Brain Dis. 2021;36(3):463–70.
Dong MX, Li CM, Shen P, Hu QC, Wei YD, Ren YF, et al. Recombinant tissue plasminogen activator induces long-term anxiety-like behaviors via the ERK1/2-GAD1-GABA cascade in the hippocampus of a rat model. Neuropharmacology. 2018;128:119–31.
Tang HY, Chiu DT, Lin JF, Huang CY, Chang KH, Lyu RK, et al. Disturbance of plasma lipid metabolic profile in Guillain-Barre syndrome. Sci Rep-Uk. 2017;7(1):8140.
Dong MX, Wei YD, Hu L. The disturbance of lipid metabolism is correlated with neuropsychiatric symptoms in patients with Parkinson’s disease. Chem Phys Lipids. 2021;239: 105112.
Wang W, Li X, Ren Y. Correlation analysis and intervention study on disturbance of lipid metabolism and diabetic peripheral neuropathy. Comput Math Method M. 2022;2022:2579692.
Jones HJ, Borges MC, Carnegie R, Mongan D, Rogers PJ, Lewis SJ, et al. Associations between plasma fatty acid concentrations and schizophrenia: a two-sample Mendelian randomisation study. The lancet Psychiatry. 2021;8(12):1062–70.
Shen S, Gong C, Jin K, Zhou L, Xiao Y, Ma L. Omega-3 fatty acid supplementation and coronary heart disease risks: a meta-analysis of randomized controlled clinical trials. Front Nutr. 2022;9: 809311.
Ferraz RB, Paixão RV, Lopes-Marques M, Machado AM, Salaro AL, Castro L, et al. The repertoire of the elongation of very long-chain fatty acids (Elovl) protein family is conserved in tambaqui (Colossoma macropomum): Gene expression profiles offer insights into the sexual differentiation process. Comparative Biochem Physiol Part B, Biochem Mol Biol. 2022;261: 110749.
Fan S, Yeon A, Shahid M, Anger JT, Eilber KS, Fiehn O, et al. Sex-associated differences in baseline urinary metabolites of healthy adults. Sci Rep-Uk. 2018;8(1):11883.
Knuplez E, Marsche G. An Updated Review of Pro- and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System. Int J Mol Sci. 2020;21(12):4501.
Brown DA, London E. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J Biol Chem. 2000;275(23):17221–4.
Hannun YA, Obeid LM. Sphingolipids and their metabolism in physiology and disease. Nat Rev Mol Cell Bio. 2018;19(3):175–91.
Loft L, Moseholm KF, Pedersen K, Jensen MK, Koch M, Cronje HT. Sphingomyelins and ceramides: possible biomarkers for dementia? Curr Opin Lipidol. 2022;33(1):57–67.
Mielke MM, Haughey NJ, Han D, An Y, Bandaru V, Lyketsos CG, et al. The association between plasma ceramides and sphingomyelins and risk of alzheimer’s disease differs by sex and APOE in the Baltimore longitudinal study of aging. J Alzheimer’s Dis : JAD. 2017;60(3):819–28.
MohammadzadehHonarvar N, Zarezadeh M, Molsberry SA, Ascherio A. Changes in plasma phospholipids and sphingomyelins with aging in men and women: A comprehensive systematic review of longitudinal cohort studies. Ageing Res Rev. 2021;68: 101340.
Leskanicova A, Babincak M, Mochnacky F, PipovaKokosova N, Kukelova D, Urbanska N, et al. Sex-dependent differences in stress-induced depression in Wistar rats are accompanied predominantly by changes in phosphatidylcholines and sphingomyelins. J Physiol Pharmacol : J Polish Physiol Soc. 2021;72(4):72.
Acknowledgements
Not applicable.
Funding
This work was supported by The Open Fund of Hubei Key Laboratory of Renmin Hospital of Wuhan University (2021KFY040).
Author information
Authors and Affiliations
Contributions
Mei-Xue Dong and You-Dong Wei designed the study. Yuan-Jun Huang and Ling Hu collected and analyzed clinical characteristics. Mei-Xue Dong and You-Dong Wei performed experiments. The first draft of the manuscript was written by Mei-Xue Dong and Wei Ke. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Ethical approval was approved by the Ethics Committee of the First Affiliated Hospital of Chongqing Medical University and all the procedures were performed in accordance with Declaration of Helsinki. Written informed consents were obtained from all the participants prior to inclusion.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Huang, YJ., Ke, W., Hu, L. et al. Liquid chromatography-mass spectrometry-based metabolomic profiling reveals sex differences of lipid metabolism among the elderly from Southwest China. BMC Geriatr 23, 156 (2023). https://doi.org/10.1186/s12877-023-03897-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12877-023-03897-z