
Abstract
Background
Receptor-type tyrosine-protein phosphatase T (PTPRT) is a transmembrane protein that is involved in cell adhesion. We previously found that PTPRT was downregulated in multiple cancer types and the mutation of PTPRT was associated with cancer early metastasis. However, the impacts of PTPRT downregulation on tumour proliferation, invasion, and clinical interventions such as immune checkpoint inhibitor (ICI) therapies remained largely unknown.
Methods
Gene expression data of non-small cell lung cancer (NSCLC) samples from The Cancer Genome Atlas database were downloaded and used to detect the differential expressed genes between PTPRT-high and PTPRT-low subgroups. Knockdown and overexpress of PTPRT in lung cancer cell lines were performed to explore the function of PTPRT in vitro. Western blot and qRT-PCR were used to evaluate the expression of cell cycle-related genes. CCK-8 assays, wound-healing migration assay, transwell assay, and colony formation assay were performed to determine the functional impacts of PTPRT on cell proliferation, migration, and invasion. KM-plotter was used to explore the significance of selected genes on patient prognosis.
Results
PTPRT was found to be downregulated in tumours and lung cancer cell lines compared to normal samples. Cell cycle-related genes (BIRC5, OIP5, and CDCA3, etc.) were specifically upregulated in PTPRT-low lung adenocarcinoma (LUAD). Modulation of PTPRT expression in LUAD cell lines affected the expression of BIRC5 (survivin) significantly, as well as the proliferation, migration, and invasion of tumour cells. In addition, low PTPRT expression level was correlated with worse prognosis of lung cancer and several other cancer types. Furthermore, PTPRT downregulation was associated with elevated tumour mutation burden and tumour neoantigen burden in lung cancer, indicating the potential influence on tumour immunogenicity.
Conclusion
Our findings uncovered the essential roles of PTPRT in the regulation of proliferation, migration, and invasion of LUAD, and highlighted the clinical significance of PTPRT downregulation in lung cancer.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Receptor-type tyrosine-protein phosphatase T (PTPRT) is a transmembrane protein encoded by the PTPRT gene [1]. Previous studies have shown that PTPRT was involved in cell adhesion and phosphorylation of signal transducer and activator of transcription (STAT) [2, 3]. Furthermore, as a well-known tumour suppressor gene, mutations in PTPRT were associated with the occurrence of cancer [4]. PTPRT was found to be frequently mutated in multiple cancer types, especially in cutaneous melanoma, gastric adenocarcinoma, endometrial cancer, and lung cancer [5, 6]. Through the analysis of tumour genomic and clinical data, our previous study also supported that mutation events in PTPRT might serve as candidate markers of tumour early metastasis [7].
In recent years, immune checkpoint inhibitors (ICIs) play important roles in the treatment of cancers [8,9,10,11], while tumour mutation burden (TMB) was considered to be a prominent marker for responses to ICIs therapy [12,13,14,15]. Previous studies have shown that PTPRT mutations were associated with higher TMB and can predict the efficacy of ICIs [16, 17]. They also found that the mutation of PTPRT was associated with enhanced infiltration of CD8+ T cells and M1-macrophages [17]. These studies suggested that the dysfunction of PTPRT may be closely related to tumour immunogenicity. Our previous study have found that PTPRT was downregulated in a variety of cancers [7], but the effects and mechanisms of PTPRT downregulation on proliferation, migration, invasion, and tumour immunogenicity of lung cancer are still poorly understood.
In this study, through analysis of the gene expression data of TCGA lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) from The Cancer Genome Atlas (TCGA) database, we depicted the genes and pathways associated with PTPRT downregulation. We then validated the functional impacts of PTPRT expression on cell proliferation, migration and invasion through in vitro experiments using lung cancer cell lines. We also highlighted the role of PTPRT in modulating the expression of cell cycle-related genes, especially for BIRC5. Finally, the potential value of PTPRT as a biomarker for the prognosis of lung cancer was explored.
Materials and methods
Samples and data collection
The expression data of TCGA pan-cancer samples were obtained from the UCSC Xena database (https://toil-xena-hub.s3.us-east-1.amazonaws.com/download/tcga_RSEM_gene_tpm.gz). The clinical information of TCGA samples ("TCGA-CDR-Supplemental Table S1.xlsx”), containing the tumour types (such as LUAD and LUSC) was downloaded from the PanCanAtlas database (https://gdc.cancer.gov/about-data/publications/pancanatlas). Then transcripts per million (TPM) values of 509 LUAD and 479 LUSC samples were extracted. Four LUAD cell lines (A549, H1299, H1975, H838) and human normal lung epithelial cells BEAS-2B were purchased from the American Type Culture Collection (ATCC) for in vitro experiments.
Detection of differentially expressed genes (DEGs)
The DEGs between the PTPRT-high and PTPRT-low subgroups were detected by the limma package (v3.52.3). We divided the samples into PTPRT-high and PTPRT-low subgroups using upper and lower quartile TPM values of PTPRT in the TCGA-LUAD (upper quartile: 0.37; lower quartile: 0.02) or TCGA-LUSC (upper quartile: 0.94; lower quartile: 0.04) cohort. The parameter “decideTestsDGE(adjust.method = "fdr", p.value = 0.01,lfc = 1)” was used to detect DEGs and ggplot2 (version 3.3.6) was used for visualization.
Functional enrichment analysis
The DEGs (FDR < 0.01 & |log2(foldchange)|> 1) between the PTPRT-high and PTPRT-low subgroups were selected for functional enrichment analysis. The Gene Ontology (GO) and KEGG pathway enrichment analysis were performed using the DAVID database (https://david.ncifcrf.gov/tools.jsp) [18]. Gene symbols of DEGs were used as input, and functional terms with “FDR < 0.05” were defined as significant enrichment.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
Total RNA was extracted using the RNA extraction kit (#RNAfast200, Fastagen, China), and the reverse transcription and qPCR were conducted using ReverTra Ace qPCR RT Kit and SYBR Green Realtime PCR Master Mix (#FSQ-101, #QPK-212, Toyobo Life Science, China) according to the manufacturer's instructions. The relative gene expression was examined by the comparative Ct method using 2−△△Ct. Primers were listed in supplementary Table S1.
Western blot
Cell samples were lysed with cold RIPA lytic buffer (#P0013B, Beyotime, China), and the extracted proteins were quantified using Pierce BCA Protein Assay Kit (#P0009, Beyotime, China). After electrophoresis was performed on a sodium dodecyl sulphate polyacrylamide gel (SDS-PAGE), the blots were transferred to a polyvinylidene difluoride membrane (#IPVH00010, Millipore, America). The membranes were incubated with diluted primary antibodies (survivin, #10,508–1-AP, 1:1000, Proteintech; PTPRT, #PA5-18,304, 1:500, Invitrogen) and GAPDH (#60,004–1-Ig, 1:16,000, Proteintech) at 4 °C overnight, and then the second antibody was added and incubated at room temperature for 30 min. ECL reagent (#RM00020, ABclonal, China) was added to the membranes to visualize the immunoreactive protein bands. Notably, the marker (Shanghai Yase Biomedical Technology Co., LTD., WJ102) was not able to be developed. Therefore, the position of the marker on the Kodak film after exposure was determined by comparing the color bands on the acetate fiber film using SDS-PAGE horizontal electrophoresis (Supplementary Fig. 1– 4).
RNA interference
To knock down PTPRT expression, different siRNAs (si-PTPRT-92, si-PTPRT-305, si-PTPRT-921) were designed and synthesized (GenePharma). A scrambled siRNA was used as the negative control (si-NC). The sequences of siRNAs were listed in Table S2. Cells were transiently transfected with 100 pmol of siRNAs using Lipofectamine 3000 (Invitrogen). After transfection for 48 h, the cells were collected for further experiments.
Overexpression of PTPRT
The coding sequence of PTRRT was synthesized and inserted into pcDNA™ 3.1Zeo ( +) plasmid for overexpression of PTPRT (OE-PTPRT). Empty pcDNA™ 3.1Zeo ( +) plasmid-transfected cells as a control (OE-NC). Transfected cells were collected 24h later, and total RNA was extracted, followed by detection of PTRRT expression by qRT-PCR.
CCK-8 and colony formation assay
The treated cells were digested and counted, and 1000 cells were inoculated into 96-well plates per well, cultured in a 37℃, 5% CO2 incubator for 0, 48h, and 72h. CCK-8 solution (#CA1210, Solarbio Life Science, China) was added to each well and was further incubated for 1-2h. Then the absorbance at 450nm was measured with a microplate reader (#ST-360, Kehua Bio-engineering Co., China) to compare the cell proliferation rates. For the colony formation assay, 400–1000 cells were seeded into 6-well plates and cultured for 10–14 days. Cells were then fixed with 4% paraformaldehyde (#30,188,928, Sinopharm, China) and stained with crystal violet staining buffer (#C0121-100ml, Beyotime, China) to allow the counting of cell colonies.
Wound-healing migration assay
About 5 × 105 cells were cultured in 6-well plates, and after monolayers of cells were formed, they were then gently scratched with a sterilized pipette tip. Then the cells were washed three times with PBS to remove the scratched cells and continued in culture with fresh medium added at 37℃ and 5% CO2 for 24 h. Photographs were taken in three random microscopic fields at 0 h, 24 h, 48h, and 72h.
Transwell assay
Transwell assays were conducted as previously described with modification [19]. Briefly, 1 × 104 cells were resuspended in a 10% FBS medium and inoculated into the upper wells of transwells. After incubation at 37℃ and 5% CO2 for 24 h, invasive cells to the lower wells were fixed with 4% paraformaldehyde and stained with 1% crystal violet (#DL22040, DLM, China) for 20 min. The number of invasive cells was counted in three microscopic fields.
Survival analysis
The survival analysis between PTPRT-high and PTPRT-low subgroups was performed on the “mRNA RNA-seq: Pancancer” module of the KM-plotter (http://kmplot.com) [20], “Auto select best cutoff” and “Follow up threshold: 60 months” were used to calculate the prognostic curves for these two subgroups.
The correlation between PTPRT and tumour immunogenicity
The mutation data of the TCGA pan-cancer cohort was downloaded from the PanCanAtlas database (https://gdc.cancer.gov/about-data/publications/pancanatlas). TMB was defined as the number of non-silent mutations (missense, nonsense, insertion/deletion, splice-site) per sample. The LUAD and LUSC samples were extracted according to tumour types. The neoantigen data of LUAD and LUSC samples was acquired from the Cancer Immunome Atlas (TCIA) database (https://www.tcia.at/home). TNB was defined as the number of neoantigens per sample.
The comparison of TMB between PTPRT-high and PTPRT-low subgroups (Fig. 7A, B, E, F) was conducted using the Comprehensive Analysis on Multi-Omics of Immunotherapy in Pan-cancer (CAMOIP) database (https://www.camoip.net/). The correlation of PTPRT expression and TMB/TNB was calculated with the “Spearman” method, using samples with both TMB/TNB and TPM values (Fig. 7C, D, G, H).
Statistical analysis
R software (v4.2.1, http://www.r-project.org) was used for all statistical analyses. The correlation between PTPRT and other genes was calculated with the “cor.test” function, and the “Spearman” method was used to calculate the correlation coefficient. P < 0.05 was considered statistically significant if not otherwise specified.
Results
Differential expressed genes (DEGs) and pathways associated with PTPRT
To reveal genes and pathways associated with the downregulation of PTPRT, we downloaded the gene expression data of LUAD (N = 509) and LUSC (N = 479) samples from TCGA database, which were divided into PTPRT-high and PTPRT-low subgroups according to the upper and lower quartile TPM values of PTPRT, respectively. DEGs between PTPRT-high and PTPRT-low subgroups were identified with the limma package (v3.52.3) (Fig. 1A-C, Table S3- 4). More DEGs were found in the TCGA-LUAD group, with 1376 DEGs unique to the TCGA-LUAD group alone (Fig. 1D). Interestingly, some cell adhesion-associated DEGs (such as ITGA8, CD22, and CLDN18) were found in both TCGA-LUAD and TCGA-LUSC cohorts and enriched in PTPRT-high subgroup (Fig. 1D), which was consistent with the previously reported function of PTPRT in cell adhesion [21]. On the other hand, many cell cycle-related DEGs (such as BIRC5, OIP5, NUF2, MCM10, MKI67, and TOP2A) were detected only in the TCGA-LUAD cohort and enriched in the PTPRT-low subgroup (Fig. 1D). To further explore the function of these DEGs, we conducted GO and KEGG pathway enrichment analysis [22]. Not surprisingly, DEGs specific to LUAD were enriched in cell cycle pathway or cell cycle-related terms, such as cell division, chromosome segregation, mitotic spindle organization, mitotic spindle, and mitotic spindle assembly checkpoint (Fig. 1E, F, Table S5- 6). Given that there are more DEGs associated with PTPRT in LUAD than in LUSC, and these DEGs are enriched in cell cycle-related pathways, we hypothesized that the downregulation of PTPRT in LUAD may have an impact on the cell proliferation of cancer cells. Therefore, we next focused on the influence of downregulation of PTPRT on the progression of LUAD.

The differential expressed genes (DEGs) between PTPRT-high and PTPRT-low subgroups in TCGA-LUAD and TCGA-LUSC cohorts and functional enrichment analysis. A DEGs between PTPRT-high and PTPRT-low subgroups in TCGA-LUAD cohort. B DEGs between PTPRT-high and PTPRT-low subgroups in TCGA-LUSC cohort. C DEGs number in TCGA-LUAD and TCGA-LUSC cohorts. D Comparison of DEGs between TCGA-LUAD and TCGA-LUSC cohorts. Top, Venn diagram showing DEGs between TCGA-LUAD and TCGA-LUSC cohorts. Middle, Venn diagram showing DEGs enriched in PTPRT-high subgroup between TCGA-LUAD and TCGA-LUSC cohorts. Bottom, Venn diagram showing DEGs enriched in PTPRT-low subgroup between TCGA-LUAD and TCGA-LUSC cohorts. E KEGG pathway enrichment of DEGs specific to LUAD. F Gene ontology enrichment of DEGs specific to LUAD. The terms marked in red in E and F are cell cycle or cell division-related pathways/biological processes
We further explored the expression correlation between PTPRT and genes involved in the cell adhesion or cell cycle pathways. As expected, genes in the cell adhesion pathway (such as CD22, ITGA8, CLDN18) were significantly positively correlated with PTPRT (all with R > 0.4, P < 0.01; Fig. 2A) Specifically, DEGs upregulated in the PTPRT-low subgroup specific to the TCGA-LUAD cohort were negatively correlated with PTPRT (Fig. 2B) in TCGA-LUAD, but not in TCGA-LUSC, including BIRC5 (R = -0.41, P < 0.001), NUF2 (R = -0.4, P < 0.001), OIP5 (R = -0.39, P < -0.001), CENPA (R = -0.39, P < 0.001), CDC20 (R = -0.39, P < 0.001), and CDCA3 (R = -0.39, P < 0.001), which were genes associated with the cell cycle pathway. The essential antiapoptotic gene BIRC5 (also known as survivin) was abundantly expressed in adenocarcinoma and involved in promoting cell proliferation and preventing cell apoptosis [23]. NUF2, OIP5 and CDC20 were key players in the regulation of cell division [24,25,26]. Together, these results showed that PTPRT expression was significantly correlated with multiple genes in the cell adhesion and cell cycle pathways in lung cancer, especially in LUAD.

Genes significantly correlated with PTPRT in TCGA-LUAD. A The cell adhesion-related genes were positively correlated with PTPRT. B The cell cycle-related genes were negatively correlated with PTPRT. Spearman correlation was determined
PTPRT inhibited the cell proliferation and expression of BIRC5 in vitro
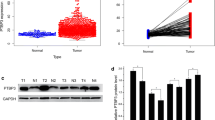
We then performed cell-based assays to explore the roles of PTPRT on cell proliferation, using four LUAD cell lines (A549, H1975, H1299, and H838). We first confirmed that the expression of PTPRT in those LUAD cells was significantly lower compared to the normal BEAS-2B bronchial epithelial cells (Fig. 3A). Among the four tumour cell lines, the expression of PTPRT was the highest in A549 and the lowest in H1975. Thus, knockdown or overexpression of PTPRT was conducted in A549 or H1975 cells (Fig. 3B, C). As CCK-8 assays showed, PTPRT-knocked-down A549 cells displayed enhanced cell proliferation, while PTPRT-overexpressing H1975 cells had decreased cell proliferation (Fig. 3D), suggesting an inhibitory role of PTPRT in cancer cell proliferating. Furthermore, cell colony formation was increased after the knockdown of PTPRT in non-malignant lung epithelial cells (Fig. 3E) or multiple LUAD cells (Fig. 3F, G). These data supported that PTPRT functioned as a negative regulator of cell proliferation.

In vitro validation of the influence of PTPRT with cell proliferation in LUAD cell lines. A qRT-PCR of PTPRT in four LUAD cell lines and a human bronchial epithelial cell lines BEAS-2B. B Knockdown of PTPRT (si-PTPRT-921) in A549 cells. C Overexpress of PTPRT (OE-PTPRT) in H1975 cells compared with negative control (OE-NC). D Cell proliferation curves after 4 different treatments (si-NC, si-PTPRT, OE-NC, and OE-PTPRT). Data were shown as the mean ± SD (n = 3). * P < 0.05, ** P < 0.01, *** P < 0.001 by unpaired Student’s t test. E–G The changes of colony formation efficiency upon PTPRT knockdown in non-malignant lung epithelial cells (E, BEAS-2B) and lung cancer cells (F, A549; G, H838). H-I The changes of cell cycle-related genes upon PTPRT knockdown (H) and overexpression (I)
To evaluate whether the cell cycle-related genes were regulated by PTPRT, we examined the expression of various genes (BIRC5, OIP5, CDC20, CDCA3, CENPA, and NUF2) by qRT-PCR after PTPRT knockdown or overexpression in A549 cells. As expected, the expression of BIRC5, CDCA3, and OIP5 was significantly upregulated upon PTPRT knockdown (Fig. 3H), while overexpression of PTPRT resulted in the downregulation of BIRC5, CDC20, CDCA3, NUF2, and OIP5 (Fig. 3I). These results further suggested the role of PTPRT in regulating the cell cycle pathway.
Due to the unique role of BIRC5 in adenocarcinoma [27, 28] and its strongest negative correlation with PTPRT expression, we then focused on the influence of modulation of PTPRT expression on BIRC5. Notably, the protein levels of BIRC5 were decreased upon PTPRT overexpression in LUAD cells (Fig. 4A, B, Supplementary Fig. 1– 2), while the expression of BIRC5 was upregulated at both mRNA (Fig. 4C) and protein (Fig. 4D, Supplementary Fig. 3) levels upon PTPRT knockdown in multiple cell lines. In addition, high expression of BIRC5 was significantly correlated with a worse prognosis in the TCGA-LUAD cohort (HR = 1.75, 95% CI 1.31 ~ 2.35, P < 0.0001; Fig. 4E). After multivariable Cox regression analysis, BIRC5 remained an independent risk factor for patients with LUAD (HR = 1.73, 95% CI 1.27 ~ 2.36, P = 0.0005, Fig. 4F). Furthermore, PTPRT was negatively correlated with BIRC5 in other cancer types such as breast carcinoma (BRCA, R = -0.497, P < 0.0001; Fig. 4G), pancreatic adenocarcinoma (PAAD, R = -0.371, P < 0.0001; Fig. 4G), stomach adenocarcinoma (STAD, R = -0.279, P < 0.0001; Fig. 4G), and mesothelioma (MESO, R = -0.465, P < 0.001; Fig. 4G). Collectively, our in-vitro results underscored the essential roles of PTPRT in inhibiting cell proliferation by affecting the expression of cell cycle-related genes like BIRC5, indicating that PTPRT downregulation could promote the malignant proliferation of tumour cells.

Regulation of PTPRT on BIRC5 and its effect on tumour prognosis. A, B Overexpression of PTPRT led to decreased expression of BIRC5 (survivin) at protein level (P = 0.029, two-sided Student's t-test) in A549 (A) and H1975 (B) cells. Three replicates of Western blots were performed, full-length gels and blots were shown in Supplementary Fig. 1– 2. C Increased BIRC5 mRNA expression in si-PTPRT compared to si-NC cells. D Increased BIRC5 protein level in si-PTPRT compared to si-NC cells, full-length gels and blots were shown in Supplementary Fig. 3. E The upregulation of BIRC5 was associated with a worse prognosis for patients with LUAD. F Univariable and multivariable Cox regression analysis showed that BIRC5 was an independent risk factor of LUAD. G. The inverse correlation of PTPRT and BIRC5 in multiple cancers. Data were shown as the mean ± SD (n = 3). * P < 0.05, ** P < 0.01, *** P < 0.001 by unpaired Student’s t test
PTPRT suppressed cell migration and invasion
We previously showed that the mutations in PTPRT were associated with the cancer metastasis of multiple cancer types [7]. To investigate whether downregulation of PTPRT was associated with cell migration and invasion, we performed wound healing and transwell assays using PTPRT-knocked-down or overexpressed tumour cells. Wound healing assays showed that PTPRT knockdown (si-PTPRT) promoted the migration capabilities of A549 cells (Fig. 5A; P = 8.059E-05). On the other hand, overexpression of PTPRT (OE-PTPRT) in A549 and H1975 cells significantly attenuated their migration capabilities (Fig. 5B-D). Consistently, transwell assays showed that PTPRT knockdown enhanced the invasion capabilities of A549 cells (Fig. 5E; P = 0.0009), and PTPRT overexpression significantly reduced the invasion capabilities of H1975 cells (Fig. 5F; P = 0.0005).

Wound healing assays of cell migration in A549 and H1975 cells. A Wound healing assays of si-NC and si-PTPRT in A549 cells. B Western blot images of PTPRT overexpression in A549 and H1975 cells, full-length gels and blots were shown in Supplementary Fig. 4. C Wound healing assays of Vector (OC-NC) and OE-PTPRT in A549 cells. D Wound healing assays of Vector (OC-NC) and OE-PTPRT in H1975 cells. Wound healing was recorded and quantitated for three microscopic fields. E Invasion of si-NC and si-PTPRT A549 cells were stained and counted under a light microscope. F Invasion of Vector (OE-NC) and OE-PTPRT H1975 cells were stained and counted under a light microscope. Scale bar, 100 µm. Data were shown as the mean ± SD (n = 3). * P < 0.05, ** P < 0.01, *** P < 0.001 by unpaired Student’s t test
PTPRT expression was associated with the prognosis of LUAD and other cancer types
As PTPRT showed regulatory effects on the proliferation, migration, and invasion of LUAD cells, we speculated that the expression level of PTPRT might be predictive for the prognosis of patients with LUAD. Using KM-plotter [20], a web-based survival analysis tool tailored for medical research, we found that high expression of PTPRT was significantly correlated with better survival rates of patients with LUAD (HR = 0.71, %95 CI 0.51 ~ 0.97, log-rank P = 0.033; Fig. 6A). Similarly, high expression of PTPRT was also significantly correlated with favourable prognosis of patients with LUSC (HR = 0.7, 95% CI 0.52 ~ 0.95, P = 0.02; Fig. 6B), breast carcinoma (HR = 0.44, 95% CI 0.29 ~ 0.65, P = 3.9E-5; Fig. 6C), head and neck squamous cell carcinoma (HNSCC, HR = 0.73, 95% CI 0.55 ~ 0.97, P = 0.029; Fig. 6D), pancreatic adenocarcinoma (PAAD, H = 0.38, 95% CI 0.25 ~ 0.59, P = 6E-6; Fig. 6E), and rectum adenocarcinoma (READ, HR = 0.36, 95% CI 0.16 ~ 0.8, P = 0.0094; Fig. 6F).

Kaplan–Meier survival curve of human cancers with high and low PTPRT expression. A LUAD, lung adenocarcinoma. B LUSC, squamous cell carcinoma of lung. C BRCA, breast carcinoma. D HNSCC, head and neck squamous cell carcinoma. E PAAD, pancreatic adenocarcinoma. F READ, Rectal adenocarcinoma. The survival analysis between PTPRT-high and PTPRT-low subgroups was performed on the “mRNA RNA-seq: Pancancer” module of the KM-plotter (http://kmplot.com)
PTPRT expression was associated with elevated TMB and TNB in lung cancer
Previous studies found that the occurrence of PTPRT mutation was associated with a high TMB and better responses to ICI therapies [16]. To explore whether the expression level of PTPRT was also associated with TMB, we divided the TCGA-LUAD cohort into the PTPRT-high subgroup and PTPRT-low subgroup using the median expression of PTPRT as a cut-off, and found that the PTPRT-high subgroup had a lower TMB and TNB than the PTPRT-low subgroup (Fig. 7A, B). The PTPRT expression level was also negatively correlated with TMB and TNB, respectively (Fig. 7C, D, Table S7- 8). Similar analysis was performed with the TCGA-LUSC cohort, and the results were in accordance with those in LUAD (Fig. 7E–H, Table S9- 10). Together, these findings suggested that the expression level of PTPRT was associated with tumour immunogenicity in lung cancer, and might be predictive of the responses to ICI therapies.

The association of PTPRT expression with TMB and TNB. TMB (A) and TNB (B) difference between PTPRT-high and PTPRT-low samples in TCGA-LUAD. TMB (C) and TNB (D) were negatively correlated with PTPRT expression level in TCGA-LUAD. TMB (E) and TNB (F) difference between PTPRT-high and PTPRT-low samples in TCGA-LUSC. TMB (G) and TNB (H) were negatively correlated with PTPRT expression levels in TCGA-LUSC. Spearman correlation was determined
Discussion and conclusion
Previous studies have shown that PTPRT functioned as a tumour suppressor involved in cell–cell aggregation and tumour progression [29,30,31]. Peyser et al. found that hypermethylation in the promoter region of PTPRT as well as mutations in PTPRT promoted the activation of the STAT3 pathway [1, 32]. In our previous studies, we found that PTPRT mutations were associated with cancer metastasis and PTPRT expression was downregulated in a variety of cancers, including lung cancer [7, 33]. However, the specific effects of PTPRT downregulation remained largely unknown.
In this study, through analysis of TCGA gene expression data, we revealed that genes associated with the downregulation of PTPRT in LUAD were enriched in cell cycle pathways. In vitro functional verification using LUAD cell lines further demonstrated that modulation of PTPRT expression affected cell proliferation profoundly through the regulation of cell cycle-related genes. Notably, we showed that BIRC5, an oncogene specially involved in adenocarcinoma [27, 28], was significantly negatively correlated with PTPRT in LUAD, but not in LUSC. BIRC5 acted as an important regulator of cell division, through directing the movement of passage protein complex (CPC) to different locations from the inner centromere [34, 35]. The expression of BIRC5 was high during fetal development and in adenocarcinoma tissues, yet low in adult tissues [23, 36]. Our results showed that knockdown of PTPRT elevated the expression of BIRC5 and promoted the proliferation, migration, and invasion of LUAD cells. On the other hand, overexpression of PTPRT suppressed BIRC5 expression as well as the proliferation, migration, and invasion of LUAD cells, suggesting that PTPRT downregulation in tumours could promote the malignant growth or metastasis of lung cancer.
In recent years, ICI therapy have emerged as potent treatment strategies for various solid cancers [37, 38]. Many studies have been devoted to exploring biomarkers for predicting therapeutic responses to ICIs, among which TMB, PD-L1, and microsatellite instability (MSI) have been proven to be useful in practice [13, 39, 40]. Previous studies have shown that PTPRT mutation was positively correlated with the level of TMB, and patients with PTPRT mutation had higher response rates to ICIs than those with wild-type PTPRT [16, 17]. In this study, we found that PTPRT downregulation correlated with higher TMB and TNB in lung cancer, suggesting its role as a candidate biomarker for ICI therapy efficacies.
In conclusion, our findings uncovered the central role of PTPRT in regulating the expression of cell cycle-related genes including BIRC5. PTPRT downregulation promoted the proliferation, migration, and invasion of LUAD cells, while PTPRT overexpression exerted opposite effects. Additionally, we emphasized the predictive value of PTPRT expression on the clinical outcomes as well as tumour immunogenicity of lung cancer. Our work suggested that PTPRT expression might stratify patients with lung cancer for the use of ICIs, and new treatment tools might be developed through increasing PTPRT expression in LUAD.
Availability of data and materials
The data reported in this study can be found in supplementary materials, and additional data can be obtained by contacting corresponding authors. The code used in this manuscript is available at: https://github.com/gkdsuperchan/PTPRT-expression.
References
Lui VW, Peyser ND, Ng PK, Hritz J, Zeng Y, Lu Y, Li H, Wang L, Gilbert BR, General IJ, et al. Frequent mutation of receptor protein tyrosine phosphatases provides a mechanism for STAT3 hyperactivation in head and neck cancer. Proc Natl Acad Sci USA. 2014;111(3):1114–9.
Huang Y, Zhang Y, Ge L, Lin Y, Kwok H. The Roles of Protein Tyrosine Phosphatases in Hepatocellular Carcinoma. Cancers. 2018;10(3):82.
Zhang X, Guo A, Yu J, Possemato A, Chen Y, Zheng W, Polakiewicz RD, Kinzler KW, Vogelstein B, Velculescu VE, et al. Identification of STAT3 as a substrate of receptor protein tyrosine phosphatase T. Proc Natl Acad Sci USA. 2007;104(10):4060–4.
Julien SG, Dubé N, Hardy S, Tremblay ML. Inside the human cancer tyrosine phosphatome. Nat Rev Cancer. 2011;11(1):35–49.
Priestley P, Baber J, Lolkema MP, Steeghs N, de Bruijn E, Shale C, Duyvesteyn K, Haidari S, van Hoeck A, Onstenk W, et al. Pan-cancer whole-genome analyses of metastatic solid tumours. Nature. 2019;575(7781):210–6.
Bailey MH, Tokheim C, Porta-Pardo E. Comprehensive Characterization of Cancer Driver Genes and Mutations. Cell. 2018;173:371–85.
Chen C, Liu H, Xu Q, Zhang X, Mu F, Liu J. Association of PTPRT Mutations with Cancer Metastasis in Multiple Cancer Types. Biomed Res Int. 2022;2022:9386477.
Tang J, Yu JX, Hubbard-Lucey VM, Neftelinov ST, Hodge JP, Lin Y. The clinical trial landscape for PD1/PDL1 immune checkpoint inhibitors. Nat Rev Drug Discovery. 2018;17(12):854–5.
Chen L. Han XJTJoci: Anti–PD-1/PD-L1 therapy of human cancer: past, present, and future. J Clin Invest. 2015;125(9):3384–91.
Saada-Bouzid E, Defaucheux C, Karabajakian A, Coloma VP, Servois V, Paoletti X, Even C, Fayette J, Guigay J, Loirat D, et al. Hyperprogression during anti-PD-1/PD-L1 therapy in patients with recurrent and/or metastatic head and neck squamous cell carcinoma. Ann Oncol. 2017;28(7):1605–11.
Long J, Qi Z, Rongxin Z. PD-1/PD-L1 pathway blockade works as an effective and practical therapy for cancer immunotherapy. Cancer Biol Med. 2018;15(2):116–23.
Endris V, Buchhalter I, Allgauer M, Rempel E, Lier A, Volckmar AL, Kirchner M, von Winterfeld M, Leichsenring J, Neumann O, et al. Measurement of tumor mutational burden (TMB) in routine molecular diagnostics: in silico and real-life analysis of three larger gene panels. Int J Cancer. 2019;144(9):2303–12.
Samstein RM, Lee CH, Shoushtari AN, Hellmann MD, Shen R, Janjigian YY, Barron DA, Zehir A, Jordan EJ, Omuro A, et al. Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat Genet. 2019;51(2):202–6.
Cristescu R, Mogg R, Ayers M, Albright A, Murphy E, Yearley J, Sher X, Liu XQ, Lu H, Nebozhyn M, et al. Pan-tumor genomic biomarkers for PD-1 checkpoint blockade-based immunotherapy. Science. 2018;362(6411)::eaar3593.
Hellmann MD, Nathanson T, Rizvi H, Creelan BC, Sanchez-Vega F, Ahuja A, Ni A, Novik JB, Mangarin LMB, Abu-Akeel M, et al. Genomic Features of Response to Combination Immunotherapy in Patients with Advanced Non-Small-Cell Lung Cancer. Cancer Cell. 2018;33(5):843–852.e844.
He Z, Li A, Lin D, Gu Y, Chen Y, Ou Q, Li L, Yao H, Yu Y. Association of immune checkpoint inhibitor with survival in patients with cancers with protein tyrosine phosphatase receptor T mutation. Clin Transl Med. 2020;10(6):e214.
Zhang W, Shi F, Kong Y, Li Y, Sheng C, Wang S, Wang Q. Association of PTPRT mutations with immune checkpoint inhibitors response and outcome in melanoma and non-small cell lung cancer. Cancer Med. 2022;11(3):676–91.
Sherman BT, Hao M, Qiu J, Jiao X, Baseler MW, Lane HC, Imamichi T, Chang W. DAVID: a web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022;50(W1):W216–w221.
Wang L, Yang H, Lei Z, Zhao J, Chen Y, Chen P, Li C, Zeng Y, Liu Z, Liu X, et al. Repression of TIF1γ by SOX2 promotes TGF-β-induced epithelial-mesenchymal transition in non-small-cell lung cancer. Oncogene. 2016;35(7):867–77.
Lánczky A, Győrffy B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J Med Internet Res. 2021;23(7):e27633.
Yu J, Becka S, Zhang P, Zhang X, Brady-Kalnay SM, Wang Z. Tumor-Derived Extracellular Mutations of PTPRT/PTP Are Defective in Cell Adhesion. Mol Cancer Res. 2008;6(7):1106–13.
Kanehisa M, Furumichi M, Sato Y, Kawashima M, Ishiguro-Watanabe M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023;51(D1):D587–d592.
Xu L, Yu W, Xiao H, Lin K. BIRC5 is a prognostic biomarker associated with tumor immune cell infiltration. Sci Rep. 2021;11(1):390.
Deng Y, Li J, Zhang Y, Hu H, Wan F, Min H, Zhou H, Gu L, Liao X, Zhou J, et al. NUF2 Promotes Breast Cancer Development as a New Tumor Stem Cell Indicator. Int J Mol Sci. 2023;24(4):4226.
Pan M, Wang Y, Wang Z, Duan H, Shao C, Ding P, Lei J, Zhao J, Ma Z, Zhang F, et al. The mitosis-related gene OIP5 is a potential biomarker in pan-cancer. Ann Transl Med. 2023;11(2):117.
Xian F, Yang X, Xu G. Prognostic significance of CDC20 expression in malignancy patients: A meta-analysis. Front Oncol. 2022;12:1017864.
Zhang Q, Chen X, Hu Y, Zhou T, Du M, Xu R, Chen Y, Tang P, Chen Z, Li J. BIRC5 Inhibition Is Associated with Pyroptotic Cell Death via Caspase3-GSDME Pathway in Lung Adenocarcinoma Cells. Int J Mol Sci. 2023;24(19):14663.
Bugide S, Edwards YJK, Gupta R, Green MR, Wajapeyee N. CBX5 loss drives EGFR inhibitor resistance and results in therapeutically actionable vulnerabilities in lung cancer. Proc Natl Acad Sci USA. 2023;120(4):e2218118120.
Hsu H-C, Lapke N, Chen S-J, Lu Y-J, Jhou R-S, Yeh C-Y, Tsai W-S, Hung H-Y, Hsieh JC, Yang T-S, et al. PTPRT and PTPRD Deleterious Mutations and Deletion Predict Bevacizumab Resistance in Metastatic Colorectal Cancer Patients. Cancers. 2018;10(9):314–28.
Zhang P, Becka S, Craig SE, Lodowski DT, Brady-Kalnay SM, Wang Z. Cancer-derived mutations in the fibronectin III repeats of PTPRT/PTPrho inhibit cell-cell aggregation. Cell Commun Adhes. 2009;16(5–6):146–53.
Wang Z, Shen D, Parsons DW. Alberto, Bardelli: Mutational Analysis of the Tyrosine Phosphatome in Colorectal Cancers. Science. 2004;304:1164–6.
Peyser ND, Freilino M, Wang L, Zeng Y, Li H, Johnson DE, Grandis JR. Frequent promoter hypermethylation of PTPRT increases STAT3 activation and sensitivity to STAT3 inhibition in head and neck cancer. Oncogene. 2016;35(9):1163–9.
Chen C, Shi C, Huang X, Zheng J, Zhu Z, Li Q, Qiu S, Huang Z, Zhuang Z, Wu R, et al. Molecular Profiles and Metastasis Markers in Chinese Patients with Gastric Carcinoma. Sci Rep. 2019;9(1):13995.
Yamagishi Y, Honda T, Tanno Y, Watanabe Y. Two histone marks establish the inner centromere and chromosome bi-orientation. Science. 2010;330(6001):239–43.
Wang H, Holloway MP, Ma L, Cooper ZA, Riolo M, Samkari A, Elenitoba-Johnson KS, Chin YE, Altura RA. Acetylation directs survivin nuclear localization to repress STAT3 oncogenic activity. J Biol Chem. 2010;285(46):36129–37.
Moriai R, Tsuji N, Moriai M, Kobayashi D, Watanabe N. Survivin plays as a resistant factor against tamoxifen-induced apoptosis in human breast cancer cells. Breast Cancer Res Treat. 2009;117(2):261–71.
Rosenberg SA, Restifo NPJS. Adoptive cell transfer as personalized immunotherapy for human cancer. Science. 2015;348(6230):62–8.
Nadal E, Massuti B, Dómine M, García-Campelo R, Cobo M, Felip E. Immunotherapy with checkpoint inhibitors in non-small cell lung cancer: insights from long-term survivors. Cancer Immunol Immunother. 2019;68(3):341–52.
Goodman AM, Kato S, Bazhenova L, Patel SP, Frampton GM, Miller V, Stephens PJ, Daniels GA, Kurzrock R. Tumor Mutational Burden as an Independent Predictor of Response to Immunotherapy in Diverse Cancers. Mol Cancer Ther. 2017;16(11):2598–608.
Blons H, Garinet S, Laurent-Puig P, Oudart J-B. Molecular markers and prediction of response to immunotherapy in non-small cell lung cancer, an update. J Thorac Dis. 2019;11(S1):S25–36.
Acknowledgements
We sincerely thank the support provided by the Department of Pathology, Peking University Shenzhen Hospital. We would also like to thank Duolaimi Biotechnology (Wuhan) Co., Ltd. for their support in cytological experiments.
Funding
This research was supported by the Guangdong Basic and Applied Basic Research Foundation under Grant No. 2022A1515111138, the Science, Technology, and Innovation Commission of Shenzhen Municipality under Grant No. JCYJ20180228175531145, and the Open Fund Project of BGI-Shenzhen (BGIRSZ20200003).
Author information
Authors and Affiliations
Contributions
C.C. and J-X.L. had full access to all the data in the study and took responsibility for the experiment’s accuracy and data analysis. C.C. Y-L.L., Q-M.X., and H-Z.L. analyzed and interpreted the data. C.C., Q-M.X. and J-X.L. revised the manuscript. All authors have read and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Peking University Shenzhen Hospital Institutional Review Board, which waived additional informed consent because all data used in this study were obtained from TCGA databases or online tools/websites.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Supplementary Table S1. Gene primers used in this study. Supplementary Table S2. siRNA sequence used in this study. Supplementary Table S3. Different expressed genes (DEGs) between PTPRT-high and PTPRT-low subgroups in TCGA-LUAD cohort. Supplementary Table S4. DEGs between PTPRT-high and PTPRT-low subgroups in TCGA-LUSC cohort. Supplementary Table S5. KEGG enrichment of DEGs (N=1376) specific to LUAD cohort. Supplementary Table S6. Gene Ontology enrichment of DEGs (N=1376) specific to LUAD cohort. Supplementary Table S7. The TMB and TPM of each sample in TCGA LUAD cohort. Supplementary Table S8. The TNB and TPM of each sample in TCGA LUAD cohort. Supplementary Table S9. The TMB and TPM of each sample in TCGA LUSC cohort. Supplementary Table S10. The TNB and TPM of each sample in TCGA LUSC cohort.
Additional file 2:
Supplementary Figure 1. Overexpression of PTPRT in A549 cells led to decreased expression of BIRC5 (survivin) at the protein level. Three replicates of Western blots were performed. Notably, the marker (Shanghai Yase Biomedical Technology Co., LTD., WJ102) was not able to be developed. Therefore, the position of the marker on the Kodak film after exposure was determined by comparing the color bands on the acetate fiber film using SDS-PAGE horizontal electrophoresis. Supplementary Figure 2. Overexpression of PTPRT in H1975 cells led to decreased expression of BIRC5 at the protein level. Three replicates of Western blots were performed. Supplementary Figure 3. Knockdown of PTPRT led to decreased expression of BIRC5 at the protein level in A549, H838, and BEAS-2B cells. Three replicates of Western blots were performed in A549 cells and once each in H1299 and BEAS-2B. Supplementary Figure 4. Western blot image of PTPRT overexpression in A549 and H1975 cells. The experiment was repeated once in each cell line.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Chen, C., Liu, H., Li, Y. et al. Downregulation of PTPRT elevates the expression of survivin and promotes the proliferation, migration, and invasion of lung adenocarcinoma. BMC Cancer 24, 63 (2024). https://doi.org/10.1186/s12885-024-11840-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-024-11840-7