
Abstract
Acute lung injury (ALI) is the result of damage to the capillary endothelia and the alveolar epithelial cell caused by various direct and indirect factors, leading to significant pulmonary interstitial and alveolar edema and acute hypoxic respiratory insufficiency. A subset of ALI cases progresses to irreversible pulmonary fibrosis, a condition with fatal implications. Zafirlukast is a leukotriene receptor antagonist licensed for asthma prevention and long-term treatment. This study demonstrated a significant improvement in lung tissue pathology and a reduction in inflammatory cell infiltration in models of lipopolysaccharide (LPS)-induced ALI and bleomycin (BLM)-induced lung inflammation following zafirlukast administration, both in vivo and in vitro. Moreover, zafirlukast was found to suppress the inflammatory response of alveolar epithelial cells in vitro and lung inflammation in vivo by reducing the activation of the TLR4/NF-κB/NLRP3 inflammasome pathway. In conclusion, zafirlukast relieved lung injury and the infiltration of inflammatory cells in the lung by regulating the TLR4/NF-κB/NLRP3 pathway.
Similar content being viewed by others

Introduction
Lung injury is the injury of the lung parenchyma caused by different injury factors. It generally refers to a series of symptoms caused by various physical and chemical injuries that occur in the lung, thus leading to the impaired function of the lung [1, 2]. Acute lung injury is a significant form of pulmonary damage, with continuous irritation or acute inflammatory damage potentially leading to pulmonary fibrosis [3]. Currently, there is a notable lack of effective therapeutic drugs for acute lung injury and pulmonary fibrosis, as the existing drugs demonstrating minimal impact on reducing mortality and extending survival time. Therefore, there is a pressing requirement to carry out research and create new therapies for pulmonary fibrosis and acute lung injury [4].
Acute lung injury is a complex condition marked by the impairment of alveolar epithelial cells resulting due to various factors. This impairment results in compromised capillary endothelial cells [5, 6], diffuse pulmonary interstitial and pulmonary edema, ultimately leading to acute hypoxic respiratory insufficiency [7]. Furthermore, acute lung injury can advance to acute respiratory distress syndrome. The pathogenesis of acute lung injury is extremely complex and intricate, involving the dysregulation of inflammatory response, aquaporin regulation, coagulation and fibrinolysis system imbalance, apoptosis, pyroptosis, and various other factors [8]. It has been reported in the literature that alveolar epithelial cells have an important regulatory role in acute lung injury, and the damaged alveolar epithelial cells is a crucial cause of lung injury [9, 10]. During the stage of injury repair, ATII cells (type II alveolar epithelial cells) have the capability to undergo trans-differentiation into ATI cells (type I alveolar epithelial cells). which can compensate for the loss of ATI cells in the lung injury stage and is essential for the reconstruction and maintenance of intact alveolar epithelial structure and function [1].
Grid shading, local alveolar septal thickening, traction bronchiectasis, and inflammatory cell infiltration are common signs of early pulmonary fibrosis progression [11, 12]. Pulmonary fibrosis is a severe lung condition marked by the excessive growth of fibroblasts and the buildup of a significant amount of extracellular matrix. The process is accompanied by inflammatory damage and structural destruction [13], leading to abnormal repair of damaged alveolar tissue and resulting in structural abnormalities [6]. The disease has a considerable impact on respiratory function, causing symptoms like dry cough and gradual dyspnea. As the disease and lung injury advance, patients undergo a decline in respiratory function [14]. At present, there are few drugs to treat pulmonary fibrosis, and none of them can alleviate the progress of the lesion, which is yet irreversible. Therefore, early intervention on inflammatory reaction caused by early pulmonary fibrosis can provide a basis for preventing the progress of pulmonary fibrosis [11].
Zafirlukast is a leukotriene receptor antagonist approved by FDA for the prevention and long-term treatment of asthma. Leukotrienes are a group of inflammatory mediators produced by arachidonic acid metabolism through 5- lipoxygenase pathway [14]. In vitro experiments show that it has a contractile effect on human bronchial smooth muscle about 1000 times stronger than histamine and platelet activating factor [8], and has a powerful effect of causing bronchial smooth muscle spasm, dilating blood vessels, improving microvascular permeability, leading to bronchial mucosal edema, up-regulating mucosal secretion function and increasing airway hyperresponsiveness. Many evidences show that leukotriene receptors regulate a variety of cell-mediated inflammatory reactions [15]. This study has assessed the effectiveness of zafirlukast in the treating acute lung injury and an early inflammatory model of pulmonary fibrosis using both in vivo and in vitro models. The objective was to identify potential drugs for these conditions [16].
Materials and methods
Animal models
Male C57BL/6 J mice aged 6–8 weeks were obtained from Charles River Laboratories in Beijing, China. All animal care and experimental procedures were conducted in accordance with the ethical guidelines established by Nankai University's Institutional Animal Care and Use Committee (IACUC) (Permit No. 2024-SYDWLL-000001), and compliance with the ARRIVE guidelines. All effort were made to minimize animal suffering. The mice were kept in a controlled environment with temperature (22–26 °C), humidity, and a 12 h light–dark cycle, with ad libitum access to water and food throughout the study. The mice in all experiments were euthanized using an intraperitoneal injection of pentobarbital sodium at a dosage of 130 mg/kg body weight, which was determined based on the recommendations of the IACUC and prior dose-tolerance studies.
The mice were given a 1% solution of pentobarbital sodium to sedate them before being given an intratracheal injection of LPS at a concentration of 3 mg/kg. The control group received the same procedure but with an equivalent volume of saline. Thirty mice were randomly allocated into five groups, each comprising six mice: control, LPS, LPS + Dexamethasone (10 mg/kg), LPS + Zafirlukast (2.5 mg/kg), and LPS + Zafirlukast (5 mg/kg). Dexamethasone was employed as the positive control. After 24 h, the mice were euthanized for the extraction of lung tissues for analysis.
Mice were received a 1% pentobarbital sodium intraperitoneal sedative injection, followed by a 2U bleomycin (BLM; Hanhui Pharmaceuticals CO., LTD, China) intratracheal injection. The control group underwent the same procedure with an equivalent amount of saline intratracheally. Thirty mice were randomly divided into five groups of six: the control group, the BLM group, the BLM + Nintedanib group (60 mg/kg), the BLM + Zafirlukast group (2.5 mg/kg), and the BLM + Zafirlukast group (5 mg/kg). Nintedanib was used as the positive control. The mice were orally administered Zafirlukast (Dalian Melium Biotechnology Co.Ltd) and Nintedanib (Shanghai Acmec Biochemical Co.ltd, China) once daily from the first to the seventh day, and then euthanized sacrificed on the seventh day for assessment.
For the BLM-induced pulmonary fibrosis 21-day model, mice were received a 1% pentobarbital sodium intraperitoneal sedative injection, followed by a 2U BLM intratracheal injection. The control group underwent the same procedure with an equivalent amount of saline intratracheally. Forty mice were randomly divided into five groups of eight (to reduce experimental error): the control group, the BLM group, the BLM + Nintedanib group (60 mg/kg), the BLM + Zafirlukast group (2.5 mg/kg), and the BLM + Zafirlukast group (5 mg/kg). Nintedanib was used as the positive control. The mice were orally administered Zafirlukast (Dalian Melium Biotechnology Co.Ltd) and Nintedanib (Shanghai Acmec Biochemical Co.ltd, China) once daily from the first to the seventh day, and then euthanized sacrificed on the 21th day for assessment. Before euthanizing.
The dosages used in our study were based on widely reported safe doses as documented in the literature [17, 18]. Furthermore to ensure statistical independence, each experiment utilized a single specimen from each mouse. Consequently, each specimen was regarded as a distinct experimental unit. For each subject, a trio of distinct researchers participated in the process: the first researcher administered the treatment in accordance with the randomized schedule, being the sole individual privy to the group assignments. A second researcher took charge of sample collection. Ultimately, a third researcher, who was also uninformed of the treatment details, evaluated the expression levels across all groups.
Cell culture
Human alveolar adenocarcinomas basal epithelial cells (A549, acquired from ATCC) were cultured in RPMI-1640 media (Solarbio, USA). Mouse lung epithelial cells (MLE-12, acquired from ATCC) were cultured in DMEM/F-12 medium (Solarbio, USA). Both types of cells were incubated at 5% CO2 and 37 °C.
Western blot
The protein was extracted from cells and tissues using RIPA buffer (Beyotime, China). The protein was isolated by SDS electrophoresis, sealed with 5% skim milk for 1 h after transfer, then treated with primary antibody at 4 °C overnight, followed by secondary antibody (Abcam, UK) at room temperature for 2 h, and finally analyzed with chemiluminescent reagent (Affinity, USA). The protein expression levels were determined using the specified primary antibodies. And more information of all antibody used in this study is listed as following (Table 1).
Real-time quantitative PCR
RNA isolation with Trizol reagent (Thermo Fisher Scientific, USA) and reverse transcription with UNICON® qPCR SYBR Green Master Mix kit (Yeasen Biotech, Shanghai, China) were performed following with the manufacturer's guidelines. The gene primers were shown as below.
GADPH-Mouse | AGGTCGGTGTGAACGGATTTG (forward), |
TGTAGACCATGTAGTTGAGGTCA (reverse). | |
GADPH-Human | GGAGCGAGATCCCTCCAAAAT (forward), |
GGCTGTTGTCATACTTCTCATGG (reverse). | |
IL-6-Mouse | GGCGGATCGGATGTTGTGAT (forward), |
GGACCCCAGACAATCGGTTG (reverse). | |
IL-6 Human | ACTCACCTCTTCAGAACGAATTG (forward), |
CCATCTTTGGAAGGTTCAGGTTG (reverse). | |
IL-1β-Mouse | GAAATGCCACCTTTTGACAGTG (forward), |
TGGATGCTCTCATCAGGACAG (reverse). | |
IL-1β-Human | ATGATGGCTTATTACAGTGGCAA (forward), |
GTCGGAGATTCGTAGCTGGA (reverse). | |
NLRP3-Mouse | ATTACCCGCCCGAGAAAGG (forward), |
TCGCAGCAAAGATCCACACAG (reverse). | |
NLRP3-Human | GATCTTCGCTGCGATCAACAG (forward), |
CGTGCATTATCTGAACCCCAC (reverse). | |
TNF-α-Mouse | GGAACACGTCGTGGGATAATG (forward), |
GGCAGACTTTGGATGCTTCTT (reverse). | |
TNF-α-Human | CCTCTCTCTAATCAGCCCTCTG (forward), |
GAGGACCTGGGAGTAGATGAG (reverse). | |
MCP-1-Mouse | GATGCAATCAATGCCCCAGTC (forward), |
TTTGCTTGTCCAGGTGGTCCAT (reverse). | |
MCP-1-Human | CAGCCAGATGCAATCAATGCC (forward), |
TGGAATCCTGAACCCACTTCT (reverse). |
Bronchoalveolar lavage fluid
After endotracheal intubation, mice were thoroughly rinsed with 0.8 ml PBS for 3 times, bronchoalveolar-lavage fluid (BALF) was collected, and then centrifuged at a low temperature of 3000 rpm at 4 °C for 10 min. The supernatant after centrifugation could be packaged and frozen in the refrigerator at -80 °C for ELISA assay. Cell precipitation was suspended with 200 μl PBS, then 1 ml of red cell lysate was added, and the suspension was prepared with PBS after centrifugation for cell viability count and smear.
Enzyme linked immunosorbent assay (ELISA)
The ELISA analysis of IL-1β, IL-6, and TNF-α in BALF and cell supernatant were analyzed by using ELISA kits following the manufacturer’s guidelines (Jianglaibio, China).
Hematoxylin–Eosin (H&E) staining
The mouse's left lung was fixed in 10% formalin overnight and subsequently encased in paraffin. The lung slices (5 μm) were made for histological examination and then stained with hematoxylin–eosin (H&E) staining (Solarbio, China).
Immunohistochemistry
Lung tissue sections fixed in paraffin were deparaffinized with xylene and healed for two minutes at a high temperature. Slices samples should be treated with the pre-prepared UltraSensitiveTM SP immunohistochemical kit (Maxim, China) following the provided guidelines, and the colour reaction should be developed using DAB staining solution from Solarbio (China). Images were captured by using a light microscope from Nikon (Japan).
Immunofluorescence
The cells were treated using 4% paraformaldehyde for fixation, permeabilized with 0.2% Triton X-100, and blocked with 5% BSA in preparation for immunofluorescence. Subsequently, the cells were incubated with the primary antibody at 4 °C overnight. Following this, cells were exposed to a secondary antibody conjugated with FITC or TRITC. DAPI was used to label the nuclei (Solarbio, China). The confocal microscope (Japanese manufacturer Nikon) was used to investigate the fluorescence.
Data analysis
The data underwent analysis with the statistical software SPSS 17.0 and GraphPad Prism 7. Mean and standard deviation were utilized to represent the measurement data. The Student's t test was employed to assess differences between the experimental and control groups. Significant variances between groups were determined through one-way analysis of variance and a subsequent Bonferroni correction. Statistical significance was established at P < 0.05.
Results
Zafirlukast alleviates LPS-induced acute lung injury in mice
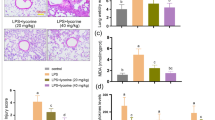
We developed the LPS-induced acute pulmonary damage model in C57BL/6 J mice to test the therapeutic efficacy of zafirlukast in treating acute lung injury. Prior to the tracheal injection of LPS, the mice received dexamethasone (10 mg/kg, intraperitoneal), zafirlukast (2.5 mg/kg and 5 mg/kg, intragastric), or saline (intragastric) one hour before. To study the effect of zafirlukast on the initial inflammatory phases of ALI, lung tissues and BALF were gathered 24 h after LPS administration. According to H&E staining, 24 h after LPS injection, considerable alveolar structural destruction and a significant inflammatory cell infiltration in BALF were caused by LPS. Zafirlukast can markedly decrease the influx of inflammatory cells and lung alveolar damage caused by LPS (Fig. 1A), and the inflammatory score of H&E images were showed in supplementary Fig. 1A. The total cell count in BALF indicates that zafirlukast, can reduce the overall cell count in the BALF compared to the LPS group (Fig. 1B), and the count of leukocytes, neutrophils, and macrophages was showed separately in (Fig. 1C-E). LPS treatment exacerbated protein leakage into the lungs, while zafirlukast can reduce the amount of protein in BALF (Fig. 1F). Furthermore, the mRNA levels of the inflammatory markers MCP-1, IL-1β, IL-6, and TNF-α in lung tissues showed a significantly decrease following zafirlukast treatment (Fig. 1G-J). Zafirlukast reduced the protein concentration of inflammatory factors IL-1β, IL-6, and TNF-α in BALF (Fig. 1K-M). The leukotrienes level of BALF in the LPS-induced 7-day pulmonary inflammation murine model by ELISA also showed that zafirlukast could significantly reduce the leukotrienes level of BALF dose-dependently (Supplementary Fig. 1B). These results indicate that zafirlukast may hinder inflammatory cell infiltration and decrease lung inflammation, thus averting acute lung injury induced by LPS.

Zafirlukast attenuates acute lung injury induced by LPS in mice. A H&E staining of lung tissue segments (Scale: 50 μm) and the BALF cell of mice (Scale: 50 μm). B Total cell counts in BALF of mice. C-E Number of inflammatory cells, lymphocytes, neutrophils, and macrophages. F Protein concentration in BALF of mice. G–J MCP-1, IL–1β, IL-6 and TNF-α mRNA expression in lung tissues of mice. K-M Measure the protein level of inflammatory factors IL–1β, IL-6 and TNF-α in BALF using ELISA. The data which presented in this study are expressed as mean ± SD (n = 3). the statistical analysis revealed significant differences when compared with the control group, denoted as #P < 0.05, ##P < 0.01, ###P < 0.001, *P < 0.05, **P < 0.01, ***P < 0.001 as compared with model group
Zafirlukast reduces lung inflammation in mice induced by BLM
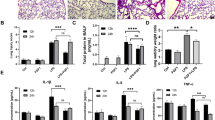
We further developed a model of lung injury induced by BLM in C57BL/6 J mice to evaluate the effect of zafirlukast on lung injury and inflammation. BLM (2 U/kg) was administered into the trachea from day 0 to day 6, and nintedanib (60 mg/kg, ig), zafirlukast (2.5 mg/kg and 5 mg/kg, ig), or saline (ig) were given daily. Lung tissues and BALF were collected 7 days after BLM injury. H&E staining of lung tissue slices and cells from the BALF showed that the stretching of the alveolar septum by BLM resulted in the infiltration of inflammatory cells., while zafirlukast was also able to repair alveolar damage (Fig. 2A), and the inflammatory score of H&E images were showed in supplementary Fig. 1C.Zafirlukast also decreased the total cell count and protein levels in BALF compared to the model group (Fig. 2B,2F), and the count of neutrophils, leukocytes and macrophages was showed separately in (Fig. 2C-E). The leukotrienes level of BALF in the BLM-induced 7-day pulmonary inflammation murine model by ELISA also showed that zafirlukast could significantly reduce the leukotrienes level of BALF dose-dependently (Supplementary Fig. 1D). Zafirlukast significantly decreased inflammatory cytokines expression in lung tissues (Fig. 2G-J) and BALF (Fig. 2K-M). These findings imply that zafirlukast can prevent persistent lung inflammation induced by BLM. To substantiate our approach, we have extended our investigation to 21 days with zafirlukast and positive drug nintedanib administration for the first 7 days and the result has been shown in the supplementary Fig. 2. This time point is selected to capture the evolution of fibrosis and to evaluate the efficacy of early intervention with zafirlukast. Our data reveal that the initiation of zafirlukast treatment during the early inflammatory phase significantly attenuates the fibrotic response at the 21-day mark.

Zafirlukast has been shown to effectively reduce acute lung injury induced by BLM in mice. A H&E staining of lung tissue segments (Scale: 50 μm) and the BALF cell of mice (Scale: 50 μm). B Total cell counts in BALF of mice. C-E Number of inflammatory cells, lymphocytes, neutrophils, and macrophages. F Protein concentration in BALF of mice. G–J MCP-1, IL–1β, IL-6 and TNF-α mRNA expression in lung tissues of mice. K–M Quantify the protein levels of the inflammatory factors IL-1β, IL-6, and TNF-α in BALF using ELISA. The data presented in this study are expressed as mean ± SD. (n = 3). Statistical analysis revealed significant differences when compared with the control group, denoted as #P < 0.05, ##P < 0.01, ####P < 0.0001, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 as compared with model group
Zafirlukast inhibits epithelial cells inflammation induced by LPS via regulating TLR4/NF-KB/NLRP3/inflammasome pathway
We conducted a detailed analysis of exploring the molecular mechanisms underlying the anti-inflammatory effects of zafirlukast through an in vitro model. A549 or MLE-12 cells were exposed to LPS and/or zafirlukast for 24 h. Subsequently, the cells were gathered to assess the levels of inflammatory factors and the activity of the TLR4/NF-B/NLRP3/inflammasome pathway. The ELISA results demonstrated that zafirlukast decreased the secretion of inflammatory factors IL-1β, IL-6 and TNF-α in LPS-stimulated A549 cells (Fig. 3A). The results of qRT-PCR further validated that zafirlukast could suppress the mRNA expression levels of inflammatory factors IL-1β, IL-6, TNF-α and MCP-1 in A549 cells stimulated by LPS (Fig. 3B). In MLE-12 cells stimulated by LPS, zafirlukast also decreased the secretion of inflammatory factors IL-1β, IL-6, TNF-α and the mRNA expression levels of IL-1β, MCP-1, TNF-α and IL-6 (Fig. 3C, D). The results indicated that zafirlukast could inhibits inflammation in epithelial cells induced by LPS in vitro.

Zafirlukast effectively mitigates inflammation induced by LPS in epithelial cells in vitro. A A549 cells were subjected to LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) lasted for 24 h, and the level of IL-1β, IL-6 and TNF-α in each group’s cell supernatant was quantified using ELISA. B A549 cells were subjected to LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) last for 12 h, and the mRNA expression levels of IL-1β, IL-6, TNF-α and MCP-1 were evaluated through RT-qPCR. C MLE-12 cells were exposed to LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) for 24 h, the IL-1β, IL-6, and TNF-α levels in the cell supernatant of each group were quantified using ELISA. D MLE-12 cells subjected to LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) for 12 h, then the mRNA expression levels of IL-1β, IL-6, TNF-α and MCP-1 were assessed using RT-q PCR. Data are shown as mean ± SD (n = 3). Statistical significance was denoted as follows: #P < 0.05, ##P < 0.01, ###P < 0.001, ####P < 0.0001 compared with the control group, and *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 compared with model group
The TLR4/NF-κB/NLRP3 inflammasome pathway activity significantly increased in LPS-treated epithelial cells, contributing to the lung injury development [19].Western blot results showed significant reduction in MyD88, TLR4, NLRP3, p-p65, cleaved-IL-1β, and cleaved-caspase1 protein levels after zafirlukast treatment in LPS-stimulated A549 cells (Fig. 4A, B) and MLE-12 cells (Fig. 4E, F). Further immunofluorescence results confirmed zafirlukast’s ability to inhibit the TLR4/NF-κB/NLRP3/inflammasome pathway in LPS-treated A549 cells (Fig. 4C, D) and MLE-12 cells (Fig. 4G, H). These findings suggest that the TLR4/NF-κB/NLRP3/inflammasome pathways were activated in epithelial cells following LPS treatment, and zafirlukast was able to suppress this pathway activation thereby protecting epithelial cells and reducing the inflammatory response.

Zafirlukast effectively inhibits the LPS-induced inflammation of epithelial cells through TLR4/NF-KB/NLRP3 pathway in vitro. A-B A549 cells were subjected to different concentration of LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) for a duration of 24 h, Western blotting was then performed for assessing the protein expression levels of MyD88, TLR4, P65, P-P65 as well as inflammasome pathway-related protein NLRP3, caspase1, Cleaved-caspase1, IL-1β and Cleaved-IL-1β in the cells (A). The quantification of optical density was also determined (B). C-D Additionally, the activation of MYD88 (C) and P-P65 (D) by immunofluorescence in A549 cells subjected to LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) for 24 h (Scale: 50 μm). (E–F) MLE-12 cells were subjected to LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) for 24 h, and the Western blotting was used to assess the MyD88, TLR4, P65, P-P65 and the inflammasome pathway related protein NLRP3, caspase1, Cleaved-caspase1, IL-1β and Cleaved-IL-1β protein expression levels in the cells (E). The optical density was quantified and presented (F). G-H MLE-12 cells were subjected to LPS or Zafirlukast (2.5 μM, 5 μM, 10 μM, 20 μM) for 24 h, and the activation of MYD88 (G) and P-P65 (H) was assessed through immunofluorescence (Scale: 50 μm). Data are shown as mean ± SD (n = 3). Statistical significance was denoted as follows: #P < 0.05, ##P < 0.01, ###P < 0.001 compared with control group, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 compared with model group
Zafirlukast effectively suppresses LPS-induced lung inflammation by regulating TLR4/NF-κB/NLRP3/inflammasome pathway in vivo
We also tested zafirlukast's anti-inflammatory molecular pathways in vivo. Total protein was isolated from the lung tissues of mice that had been given a 24 h course of treatment with LPS, dexamethasone, zafirlukast, or saline. According to the results of western blot, mice suffering from acute lung injury exhibited elevated levels of TLR4, MyD88, NLRP3, p-p65, cleaved-IL-1β, and cleaved-caspase 1 in their lung tissues. The associated protein expression saw a significant reduction after zafirlukast treatment. (Fig. 5A, B). The immunohistochemistry results showed that zafirlukast can decrease the levels of MYD88 and p-p65 in lung tissues (Fig. 5C). These findings indicate that zafirlukast can inhibit the activation of the TLR4/NF-κB/NLRP3/inflammasome pathways and reduce LPS-induced acute lung damage in vivo.

Zafirlukast effectively mitigates lung inflammation induced by LPS by the regulation of the TLR4/NF-KB/NLRP3 pathway in vivo. A-B MYD88, TLR4, P65, P-P65, NLRP3, caspase1, Cleaved-caspase1, IL-1β and Cleaved-IL-1β protein levels in lung homogenate were assessed using Western blot (A), Below is the analysis of the corresponding optical density. B. C Immunohistochemical analysis was performed to assess the expression levels of MYD88 and P-P65 in lung tissue sections, and quantitative results were obtained. Data are expressed as mean ± SD (n = 3). Statistical significance was denoted as #P < 0.05, ##P < 0.01 compared with control group, *P < 0.05, **P < 0.01 compared with model group
Zafirlukast effectively suppresses BLM-induced lung inflammation by controlling the TLR4/NF-κB/NLRP3/inflammasome pathway in vivo
Then we tested zafirlukast's in vivo anti-inflammatory molecular pathways in BLM model. Total protein was taken from the lung tissues of mice that had been given BLM, nintedanib, zafirlukast, or saline treatment for 7 days. The Western blot analysis indicated that animals with BLM-induced inflammation exhibited higher levels of TLR4, MyD88, NLRP3, p-p65, cleaved-IL1β and cleaved-caspase 1in the lung tissues. The corresponding protein expression notably decreased after zafirlukast therapy (Fig. 6Α, Β). In the immunohistochemical experiments, zafirlukast was observed to reduce the protein levels of MYD88 and p-p65 in lung tissues (Fig. 6C). It has been demonstrated that zafirlukast has the ability to suppress the activation of TLR4/NF-κB/NLRP3/inflammasome pathways, consequently inhibiting the inflammatory response and alleviating BLM-induced inflammation in vivo.

Zafirlukast effectively reduces lung inflammation induced by BLM by regulating the TLR4/NF-KB/NLRP3 pathway in vivo. A-B The Western blot method was used to assess the protein levels of MYD88, TLR4, P65, P-P65, NLRP3, caspase1, Cleaved-caspase1, IL-1β, and Cleaved-IL-1β in lung homogenate. A The following analysis of optical density is presented below. B. C Identification via immunohistochemistry and quantification of MYD88 and P-P65 expression levels in sections of lung tissue. Data are expressed as mean ± SD (n = 3). #P < 0.05, ##P < 0.01, ###P < 0.001 compared with control group, *P < 0.05, **P < 0.01, ***P < 0.001 compared with model group
Discussion
The lung plays a crucial role as a respiratory organ within the human body. In clinic, lung damage is still rather common. This is a dangerous lung disease that can lead to respiratory failure, pulmonary edema, atelectasis, and other health problems. Pulmonary fibrosis is mostly a disease of the interstitial lung, alveoli, or bronchioles that can impair the respiratory, neurological, and circulatory systems [2]. Both acute lung injury and pulmonary fibrosis currently lack effective medications and treatment approaches, making it critical to discover new drugs [20]. In this investigation, we discovered that zafirlukast can alleviate LPS-induced acute lung damage and BLM-induced early inflammation of pulmonary fibrosis in mice. The mechanistic study demonstrates that zafirlukast can reduce the inflammatory response via TLR4/NF-B/NLRP3, and so ameliorate acute lung injury and lung inflammation [21].
Lipopolysaccharide (LPS), the main component of the outer membrane of gram-negative bacteria, is associated with many diseases, such as sepsis and pneumonia. Therefore, LPS is often used to establish animal models of acute lung injury, and non-invasive infusion of LPS through the trachea leads to infiltration of inflammatory cells in the lungs of mice, causing acute inflammation within 24 h [22]. Bleomycin (BLM), as a clinical chemotherapy drug for the treatment of malignant tumors, can also induce lung inflammatory damage and fibrosis when administered in large doses. Thus, in animal models, BLM is often used to cause lung damage and fibrosis [23]. The damage of lung tissue after BLM modeling was time dependent. BLM can cause pulmonary inflammation in the first week of modeling, after administration of BLM, an increase in collagen and inflammatory cell infiltration can be observed significantly, in particular in perialveolar areas, by day 7, tissues showed progressive morphological changes and a reginal interstitial fibrosis [24, 25], and the pathological changes include alveolar epithelial cell injury, inflammatory cell infiltration and increased levels of pro-inflammatory factors. Pulmonary fibrosis progressed gradually during the second to third week after BLM modeling, and pathological changes included abnormal activation of myofibroblasts and massive extracellular matrix deposition [10, 26]. Therefore, in our study, we induced inflammatory damage in mouse lung tissue using LPS for 24 h or BLM for 7 days. To substantiate our approach, we have extended our investigation to encompass a 21-day post-bleomycin treatment model. This finding is consistent with the hypothesis that modulating the immune response during the initial stages of pulmonary fibrosis can alter the disease trajectory.
In acute lung injury, alveolar epithelial cells play an important role in the innate immune response at injury initiation, such as involvement in neutrophil extracellular traps and the release of histones and inflammatory mediators [27]. Histones have the ability to activate cells, thus triggering an inflammatory response, and changing permeability of cells, ultimately causing lung injury. Our research findings has shown that zafirlukast has the potential to reduce alveolar protein leakage in lung-damaged mice, while also improving the extent of pathological damage to the lungs [28]. Furthermore, zafirlukast has been observed to decrease the expression of inflammatory mediators such as IL-1β, TNF-α, and MCP-1 in lung tissues as well as reduce the infiltration of the inflammatory cell in mouse alveoli. These results suggest that zafirlukast may act as preventative measure against lung injury and the subsequent inflammatory cascade induced by BLM or LPS, thereby indicating the drug's potential efficacy in treating lung injury illnesses [29].Besides, we designed further experiment to test if zafirlukast could inhibit the expression of leukotriene in both LPS-induced and BLM-induced model as zafirlukast is a leukotriene receptor antagonist approved by FDA, the specific results have been shown in supplementary Fig. 1B and 1D. Our findings indicate that leukotriene levels were significantly elevated in both models, which is consistent with the known inflammatory effects of LPS and BLM [30,31,32,33]. Importantly, treatment with zafirlukast led to a significant reduction in leukotriene levels compared to the untreated groups, suggesting that zafirlukast may exert its anti-inflammatory effects in both murine models as we expected.
The TLR4, a member of the pattern recognition receptor family, is an integral transmembrane protein. Upon activation, TLR4 induces an upregulation of NF-κB, a transcription factor pivotal for the synthesis of proinflammatory cytokines and its downstream signals [34,35,36]. These cytokines are instrumental in the activation of the innate immune response, a critical component in the etiology of pulmonary fibrosis [37]. Furthermore, TLR4 has been implicated in the exacerbation of fibrosis in lung injury induced by bleomycin, as evidenced by the heightened expression of TLR4-related proteins, which are hallmarks of fibrotic processes [19, 38, 39]. Consistent with these findings, the current study demonstrates that bleomycin administration results in an elevated expression of TLR4, correlating with pulmonary inflammation [40]. The therapeutic intervention with zafirlukast has been shown to attenuate this effect by inhibiting the TLR4/NF-κB signaling cascade, thereby mitigating the ensuing pulmonary fibrosis. Moreover, research has been shown that LPS can directly bind to TLR4, leading to the activation of NF-κB and the initiation of genes associated with the inflammatory response [41,42,43,44]. In addition, the significant effect of zafirlukast in both lung inflammatory model may suggest that TLR4 inhibitors could suppress the pulmonary inflammation in the same way as zafirlukast, which is in accordance with Zhou J, et al. [45] and Liu Y, et al. [46] who found that TLR4-antagonist treatment can alleviate pulmonary inflammation induced by LPS or BLM.
Studies suggest that the activation of NF-κB in lung epithelial cells can induce neutrophil recruitment, arterial hypoxemia, pulmonary edema, and even mortality without infection or external stimuli. It is important to note that the emergence of the NLRP3 inflammasome and the subsequent activation of IL-1β have been linked to acute lung injury as well as chronic lung conditions in the long term [28]. The impact of zafirlukast on the TLR4/NF-κB/NLRP3 inflammasome pathway in the lung epithelial cells and tissues was investigated, and the findings demonstrated its ability to suppress the expression of MYD88, TLR4, and NF-κB induced by LPS and BLM. Furthermore, it was observed to reduce the production of NLRP3 inflammasome-associated proteins and the generation of cleaved-IL-1β, consequently mitigating the inflammatory response [47].
Conclusion
In conclusion, research on animal pharmacodynamics and pharmacology of zafirlukast shown its potential to effectively alleviate the advancement of different stages of lung damage, such as acute lung injury and early fibrosis. Nonetheless, further investigation is elucidated the specific targets of this medication. Zafirlukast has a certain degree of clinical safety verification as a marketed medicine, which allows for a significant reduction in the amount of time needed to conduct research and create additional indications. This study offers prospective compounds for research on lung damage disease.
Availability of data and materials
The datasets generated and/or analyzed during the current study and information about unique materials used in this study are available from the corresponding author upon reasonable request. This includes the raw data files, statistical analysis outputs, and any other materials that were used to reach the conclusions of the study.
References
Robb CT, Regan KH, Dorward DA, et al. Key mechanisms governing resolution of lung inflammation. Seminars in Immunopathology. 2016;38(4):425–48.
Go RE, Lee SM, Shin YJ, et al. Time-dependent effect of inhaled cigarette smoke exposure in the bleomycin-induced lung injury rat model. Environ Toxicol. 2022;37(5):1231–43.
Gordon D, Rubenfeld MD, Caldwell E, et al. Incidence and outcomes of acute lung injury. New England J Med. 2005;353:1685–93.
Kolb P, Upagupta C, Vierhout M, et al. The importance of interventional timing in the bleomycin model of pulmonary fibrosis. Eur Respir J. 2020;55(6):1901105.
del Giudice MM, Pezzulo A, Capristo C, et al. Leukotriene modifiers in the treatment of asthma in children. Ther Adv Respir Dis. 2009;3(5):245–51.
Trinh HKT, Lee S-H, Cao TBT, et al. Asthma pharmacotherapy: an update on leukotriene treatments. Expert Rev Respir Med. 2019;13(12):1169–78.
Hughes KT, Beasley MB. Pulmonary manifestations of acute lung injury: more than just diffuse alveolar damage. Arch Pathol Lab Med. 2017;141(7):916–22.
Franco Amaral A, De Figueiredo Braga Colares P, Adib Kairalla R. Idiopathic pulmonary fibrosis: current diagnosis and treatment. J Br Pneumol. 2023;49(4):e20230085.
Theron AJ, Steel HC, Tintinger GR, et al. Cysteinyl leukotriene receptor-1 antagonists as modulators of innate immune cell function. J Immunol Res. 2014;2014:1–16.
Jenkins RG, Moore BB, Chambers RC, et al. An official american thoracic society workshop report: use of animal models for the preclinical assessment of potential therapies for pulmonary fibrosis. Am J Respir Cell Mol Biol. 2017;56(5):667–79.
Baratella E, Ruaro B, Giudici F, et al. Evaluation of correlations between genetic variants and high-resolution computed tomography patterns in idiopathic pulmonary fibrosis. Diagnostics. 2021;11(5):762.
Selman M, Pardo A. The leading role of epithelial cells in the pathogenesis of idiopathic pulmonary fibrosis. Cell Signal. 2020;66:109482.
Beers MF, Morrisey EE. The three R’s of lung health and disease: repair, remodeling, and regeneration. J Clin Investig. 2011;121(6):2065–73.
Yuksel H, Ozbilgin K, Coskun S, et al. 2003 protective effect of leukotriene receptor antagonist montelukast on smoking-induced lung injury in wistar rats. Acta Med Okayama. 2003;57:13–9.
Seth J, Kligerman MD, Teri J, Franks MD, Jeffrey R, et al. Organization and fibrosis as a response to lung injury in diffuse alveolar damage, organizing pneumonia, and acute fibrinous and organizing pneumonia. Radio Pathol Arch. 2013;33:1951–75.
Chen L, Yang Y, Li C-T, et al. CysLT 2 receptor mediates lipopolysaccharide-induced microglial inflammation and consequent neurotoxicity in vitro. Brain Res. 2015;1624:433–45.
Nayak BK, Kumar A, Kothiyal P. Pharmacological evaluation of Zafirlukast in experimentally induced global cerebral ischemia/reperfusion injury in mice. Int J Pharm Pharm Sci. 2018;10(2):30–6. https://doi.org/10.22159/ijpps.2018v10i2.21633.
Jain NK, Kulkarni SK, Singh A. Role of cysteinyl leukotrienes in nociceptive and inflammatory conditions in experimental animals. Eur J Pharmacol. 2001;423(1):85–92. https://doi.org/10.1016/S0014-2999(01)01083-4.
Gan W, Li X, Cui Y, Xiao T, Liu R, Wang M, Wei Y, Cui M, Ren S, Helian K, Ning W, Zhou H, Yang C. Pinocembrin relieves lipopolysaccharide and bleomycin induced lung inflammation via inhibiting TLR4-NF-κBNLRP3 inflammasome signaling pathway. Int Immunopharmacol. 2021;90:107230. https://doi.org/10.1016/j.intimp.2020.107230.
Xiaohe Li, Ling Ma, Yuli Wei,et al. Cabozantinib ameliorates lipopolysaccharide-induced lung inflammation and bleomycin--induced early pulmonary fibrosis in mice[J]. International Immunopharmacology, 2021:1567–5769.
Davino-Chiovattoa JE, Oliveira-Junior MC, MacKenzie B. Montelukast, leukotriene inhibitor, reduces lps-induced acute lung inflammation and human neutrophil activation. 2019:0300–2896.
Lin Y-C, Huang M-Y, Lee M-S, et al. Effects of montelukast on M2-related cytokine and chemokine in M2 macrophages. J Microbiol Immunol Infect. 2018;51(1):18–26.
Wang HM, Bodenstein M, Markstaller K. Overview of the Pathology of Three Widely Used Animal Models of Acute Lung Injury. Eur Surg Res. 2008;40(4):305–16.
Arezzini B, Vecchio D, Signorini C, Stringa B, Gardi C. F2-isoprostanes can mediate bleomycin-induced lung fibrosis. Free Radic Biol Med. 2018;115:1–9. https://doi.org/10.1016/j.freeradbiomed.2017.11.007.
Degryse AL, Lawson WE. Progress toward improving animal models for idiopathic pulmonary fibrosis. Am J Med Sci. 2011;341:444–9.
Carrington R, Jordan S, Pitchford SC, Page CP. Use of animal models in IPF research. Pulm Pharmacol Therapeu. 2018;51:73–8 ISSN 1094-5539.
Al-Amran FG, Hadi NR, Hashim AM. Cysteinyl leukotriene receptor antagonist montelukast ameliorates acute lung injury following haemorrhagic shock in rats†. Eur J Cardiothorac Surg. 2013;43(2):421–7.
Guo Z, Zeng X, Zheng Y. Zafirlukast ameliorates Docetaxel-induced activation of NOD-like receptor protein 3 NLRP3 inflammasome mediated by sirtuin1 SIRT1 in hepatocytes. Bioengineered. 2021;12(2):11030–40.
Lu Chen, Yi Yang, Chen-Tan Li,et al. CysLT2 receptor mediates lipopolysaccharideinduced microglial inflammation and consequentneurotoxicity in vitro. 2015,08,007:0006–8993.
Li X, Xie M, Lu C, Mao J, Cao Y, Yang Y, Wei Y, Liu X, Cao S, Song Y, Peng J, Zhou Y, Jiang Q, Lin G, Qin S, Qi M, Hou M, Liu X, Zhou H, Yang G, Yang C. Design and synthesis of Leukotriene A4 hydrolase inhibitors to alleviate idiopathic pulmonary fibrosis and acute lung injury. Eur J Med Chem. 2020;1(203):112614.
Failla M, Genovese T, Mazzon E, Gili E, Muià C, Sortino M, Crimi N, Caputi AP, Cuzzocrea S, Vancheri C. Pharmacological inhibition of leukotrienes in an animal model of bleomycin-induced acute lung injury. Respir Res. 2006;7: 137.
Lv J, Xiong Y, Li W, Yang W, Zhao L, He R. BLT1 mediates bleomycin-induced lung fibrosis independently of neutrophils and CD4+ T cells. J Immunol. 2017;198(4):1673–84.
Davino-Chiovatto JE, Oliveira-Junior MC, MacKenzie B, Santos-Dias A, Almeida-Oliveira AR, Aquino-Junior JCJ, Brito AA, Rigonato-Oliveira NC, Damaceno-Rodrigues NR, Oliveira APL, Silva AP, Consolim-Colombo FM, Aimbire F, Castro-Faria-Neto HC, Vieira RP. Montelukast, leukotriene inhibitor, reduces LPS-induced acute lung inflammation and human neutrophil activation. Arch Bronconeumol (Engl Ed). 2019;55(11):573–80. https://doi.org/10.1016/j.arbres.2019.05.003. English, Spanish Epub 2019 Jun 27.
Wei Y, Qi M, Liu C, Li L. Astragalus polysaccharide attenuates bleomycin-induced pulmonary fibrosis by inhibiting TLR4/ NF-κB signaling pathway and regulating gut microbiota. Eur J Pharmacol. 2023;944:175594. https://doi.org/10.1016/j.ejphar.2023.175594. Epub 2023 Feb 17.
Li Y, Cai W, Jin F, Wang X, Liu W, Li T, Yang X, Liu H, Xu H, Yang F. Thalidomide alleviates pulmonary fibrosis induced by silica in mice by inhibiting ER stress and the TLR4-NF-κB pathway. Int J Mol Sci. 2022;23(10):5656. https://doi.org/10.3390/ijms23105656.PMID:35628464;PMCID:PMC9144898.
Wang L, Shao M, Jiang W, Huang Y. Resveratrol alleviates bleomycin-induced pulmonary fibrosis by inhibiting epithelial-mesenchymal transition and down-regulating TLR4/NF-κB and TGF-β1/smad3 signalling pathways in rats. Tissue Cell. 2022;79:101953. https://doi.org/10.1016/j.tice.2022.101953. Epub 2022 Oct 7 PMID: 36228366.
Kabel AM, Estfanous RS, Alrobaian MM. Targeting oxidative stress, proinflammatory cytokines, apoptosis and toll like receptor 4 by empagliflozin to ameliorate bleomycin-induced lung fibrosis. Respir Physiol Neurobiol. 2020;273:103316. https://doi.org/10.1016/j.resp.2019.103316. Epub 2019 Oct 7.
Jia L, Sun P, Gao H, Shen J, Gao Y, Meng C, Fu S, Yao H, Zhang G. Mangiferin attenuates bleomycin-induced pulmonary fibrosis in mice through inhibiting TLR4/p65 and TGF-β1/Smad2/3 pathway. J Pharm Pharmacol. 2019;71(6):1017–28.
Wang J, Wang H, Fang F, Fang C, Wang S, Lu C, Liu N. Danggui buxue tang ameliorates bleomycin-induced pulmonary fibrosis by suppressing the TLR4/NLRP3 signaling pathway in rats. Evid Based Complement Alternat Med. 2021;23(2021):8030143.
He Z, Zhu Y, Jiang H. Inhibiting toll-like receptor 4 signaling ameliorates pulmonary fibrosis during acute lung injury induced by lipopolysaccharide: an experimental study. Respir Res. 2009;10(1):126. https://doi.org/10.1186/1465-9921-10-126.
Hennessy EJ, Parker AE, O’Neill LAJ. Targeting Toll-like receptors: emerging therapeutics? Nat Rev Drug Discovery. 2010;9(4):293–307.
Gon Y. Toll-Like Receptors and Airway Inflammation. Allergol Int. 2008;57(1):33–7.
Zhang C, Wang X, Wang C, He C, Ma Q, Li J, Wang W, Xu YT, Wang T. Qingwenzhike prescription alleviates acute lung injury induced by LPS via inhibiting TLR4/NF-kB pathway and NLRP3 inflammasome activation. Front Pharmacol. 2021;23(12):790072.
Wang Z, Li F, Aga EB, Liang X, He C, Yin L, Xu F, Li H, Tang H, Lv C. “Pterocephalodes hookeri-Onosma hookeri” decoction protects against LPS-induced pulmonary inflammation via inhibiting TLR4/ NF-κB signaling pathway. J Ethnopharmacol. 2024;318(Pt A):116918.
Zhou J, Peng Z, Wang J. Trelagliptin alleviates lipopolysaccharide (LPS)-Induced Inflammation and oxidative stress in acute lung injury mice. Inflammation. 2021;44(4):1507–17. https://doi.org/10.1007/s10753-021-01435-w. Epub 2021 Mar 10 PMID: 33751359.
Liu Y, Chen H, Wu Y, Ai F, Li W, Peng H, Gui F, Yu B, Chen Z. Sinomenine attenuates bleomycin-induced pulmonary fibrosis, inflammation, and oxidative stress by inhibiting TLR4/NLRP3/TGFβ signaling. Inhal Toxicol. 2024;36(4):217–27. https://doi.org/10.1080/08958378.2024.2335193. Epub 2024 May 7 PMID: 38713814.
Chen WC, Yu WK, et al. NLRP3 inflammasome activates endothelial-to-mesenchymal transition via focal adhesion kinase pathway in bleomycin-induced pulmonary fibrosis. Int J Mol Sci. 2023;24:15813.
Acknowledgements
We would like to express our sincere gratitude to all members who participated in this study.
Funding
This study was supported by The National Natural Science Foundation of China [Grant 82270069, 82070060].
Author information
Authors and Affiliations
Contributions
T.X drafted the work, Q.Z and T.Z wrote the main manuscript, L.M and J.L prepared Figs. 1– 2, D.C and Y.L prepared Fig. 3– 4, Z.Y and R.J prepared Fig. 5– 6, Y.C and J.G interpreted the date, X.L, A.X and H.Z reviewed the manuscript. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal welfare and experimental protocols adhered to the approved guidelines of the Institutional Animal Care and Use Committee (IACUC) of Nankai University (Permit No. 2024-SYDWLL-000001).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Xue, T., Zhang, Q., Zhang, T. et al. Zafirlukast ameliorates lipopolysaccharide and bleomycin-induced lung inflammation in mice. BMC Pulm Med 24, 456 (2024). https://doi.org/10.1186/s12890-024-03273-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12890-024-03273-6