
Abstract
Background
Newcastle disease (ND) is one of the most contagious and devastating diseases to poultry in the world. The causative agents are virulent strains of Newcastle disease virus (NDV), which belong to the genus Avulavirus, sub-family Paramyxoviridae, family Paramyxovirinae. Knowing the genomic and antigenic characteristics of virulent NDVs might contribute to ND control in China.
Results
The results showed that all of the virulent strains belonged sub-genotype VIId shared the same cleavage site 112RRQKR/F117 in the fusion protein. At least 69 % (38 of 55) of the NDV strains possessed E347K variation in the hemagglutinin-neuraminidase protein. The cross-neutralization tests confirmed that the strains harboring 347 K showed lower antigenic relatedness with LaSota. Furthermore, the immune-challenge experiment indicated that LaSota could not provide complete protection against infection with the E347K variant NDVs as the vaccinated birds were still able to be infected and shed virulent challenge viruses.
Conclusions
Currently, sub-genotype VIId NDVs are the prevalent virulent strains circulating among vaccinated chicken flocks in Eastern China. Our findings indicated that the E347K variation in HN gene would expand the antigenic difference with LaSota, which may be responsible for the increasing isolation rate of these strains from vaccinated chickens.
Similar content being viewed by others


Background
Newcastle disease (ND) is one of the most contagious and devastating diseases to poultry in the world [1]. The causative agents are virulent strains of Newcastle disease virus (NDV), which belong to the genus Avulavirus, sub-family Paramyxoviridae, family Paramyxovirinae [2, 3]. The viral RNA genome is approximately 15 kb in length and encodes six major virus proteins: the nucleocapsid protein (NP), phosphoprotein (P), matrix protein (M), fusion protein (F), hemagglutinin–neuraminidase (HN) and large polymerase protein (L). In addition, the HN protein is an important multifunctional surface glycoprotein that consists of a cytoplasmic domain, a transmembrane region, a stalk region and a globular head, in which the receptor–binding site, the site responsible for neuraminidase activity and all of the antigenic sites reside [4]. It has been confirmed that there are seven overlapping antigenic sites in the HN protein, whereas residues 345 to 353 constitute the only linear epitope identified in the HN gene, which is susceptible to immune pressure to generate antigenic variation [5]. Researchers have recently confirmed that the variation in the linear epitope in the HN protein would intensify the antigenic difference [6]. Previously, seven neutralizing epitopes positioned at residues 72, 74, 75, 78, 79, 157 to 171, and 343 of the F protein have been identified.
The strains of NDV can be divided into two distinct clades: class I and class II, and based on the older classification system, both clades could be divided into 9 genotypes, and the class II clade can be divided into fifteen genotypes when based on the new classification system, while the class I clade can be divided into three sub-genotypes. To date, a strict vaccination policy has been implemented in China and the vaccine strain LaSota which belonged to genotype II has been widely used in China for over 40 years. However, infections of genotype VIId NDVs have still frequently occurred in China and other Asian countries [7–13]. Meanwhile, there are also some virulent genotypes (VIb, IX) circulating among avian species in China [14, 15].
To elucidate the circulation of virulent NDVs among chickens in Eastern China, 55 virulent NDVs isolated from chicken flocks, including broilers and layers, from 2011 to 2013 were phylogenetically characterized.
Results
Isolation and identification of NDVs from clinical samples
Fifty-five NDVs from clinical samples of different chicken flocks in Eastern China were isolated from 2011 to 2013, identified and plaque-purified, and the details of the NDV isolates are shown in Table 1.
Phylogenetic analysis
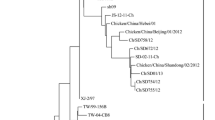
The phylogenetic analysis of the 55 NDVs based on the whole F gene sequences showed that all strains were clustered into sub-genotype VIId and they could be divided into two separate groups, namely VIId1 (n = 3) and VIId2 (n = 52), as previously described (Fig. 1) [7]. All of the strains isolated in 2012–2013 belonged to VIId2, and only three of the twenty one strains isolated in 2011 belonged to VIId1. All of the NDV isolates possessed the virulent F protein cleavage site motif 112RRQKR/F117. And over 70 % (39/55) of the isolates shared K78R variation on the F gene.

Phylogenetic tree of 55 sub-genotype VIId NDV strains based on the whole region of the F gene
Additionally, a phylogenetic analysis was also performed based on the whole HN gene (Fig. 2). As determined from the phylogenetic tree based on the HN gene, all of the NDV isolates showed a distribution similar to that obtained from the tree based on the F genes: three in VIId1 and 52 in VIId2. Three residues, namely T102, A118 and T443, were unique for VIId1, whereas I102, E118 and M443 were characteristic residues of VIId2. All of the NDV isolates shared 96.3 % to 100 % nucleotide identity and 96.2 % to 99.8 % amino acid identity. Only two isolates (JS-08-11-Ch and JS-10-11-Ch) shared the E347G variation. More than 42.8 % of the isolates in 2011 (9/21) exhibited the E347K variation in HN, whereas 81.5 % (22 of 27) of those isolated in 2012 possessed this variation. The latter percentage is nearly two-fold higher than that found in 2011, and 100 % (7 of 7) of the strains isolated in 2013 possessed this variation. In addition, the layer-origin stains isolated in 2011, 2012 and 2013 shared 67 % (6/9), 75 % (6/8) and 100 % (3/3) of the E347K variation in the HN gene, respectively. Furthermore, we found that all of the strains harboring 347 K on HN also shared a G362A mutation. However, the role of this mutation has not been well elucidated. The 13 cysteine residues were conserved as described previously, and with the exception of the loss of a glycosylation site at 508 of JS-02-11, all of the other NDVs possessed six well-identified glycosylation sites.

Phylogenetic tree of 55 sub-genotype VIId NDV strains based on the complete HN gene
Pathogenicity tests
As is shown in Table 2, all the ten isolates were highly virulent, which were consistent with the tyipical virulent motif 112RRQKRF117 at the F cleavage sites.
Cross hemagglutination inhibition (HI) test and virus neutralization test
The HI titers of the anti-LaSota serum for the ten isolates were 4-8-fold lower than that for the homologous strain LaSota, displaying the significance of the antigenic difference between the NDV isolates and the vaccine strain LaSota. As determined from the cross HI test, the R values between the predominant strains and LaSota were all lower than 0.5, indicating the existence of a significant antigenic difference between the strains. Meanwhile the R values between the E347K-variant strains and LaSota were lower than that found for the 347E/G strains, which indicate that the E/G347K variation would expand the antigenic difference with the vaccine strain. And the significant antigenic difference was also confirmed by the R value obtained from cross-neutralization test between LaSota and the predominant strains (Table 3).
Test of the protective efficacy of the LaSota strain against the variant strains
As is shown in Table 4, none of the birds vaccinated with the LaSota vaccine showed clinical signs post challenge with JS-22-11-Ch or JS-14-12-Ch. In contrast, 100 % of the unvaccinated birds challenged with JS-22-11-Ch or JS-14-12-Ch died within five days post challenge. All of the unchallenged birds remained normal throughout the experiment.
As is shown in Table 5, tracheal and cloacal swabs were collected from the birds at days 3, 5, and 7 post challenge and subjected to virus isolation tests. The virus isolation rate from the tracheal swabs was 100 % (5/5) at day 3 post challenge in both the control-JS22 and control-JS14 groups, whereas the isolation rates for the La-JS22 and La-JS14 groups were 70 % (7/10) and 100 % (10/10), respectively. And the virus isolation rate from the tracheal swabs was 50 % (5/10) and 70 % (7/10) at day 5 post challenge in La-JS22 and La-JS14 groups, whereas the isolation rates at day 7 post challenge were 10 % (1/10) and 30 % (3/10), respectively.
Discussion
The genotype VII NDVs have become the most prevalent strains in China since the 1990s, and the sub-genotype VIId NDVs are mainly responsible for the present ND epizootic in China [7, 13, 16]. In this study, virulent NDVs from the vaccinated chicken flocks were characterized as sub-genotype VIId and could be clustered into sub-genotype VIId1 and VIId2 based on the phylogenetic trees based on both F and HN genes [7]. T102, A118, and T443 of the HN gene were unique residue substitutions found in the VIId1 NDVs, and I102, E118, and M443 of the HN gene were characteristic residues of the VIId2 NDVs. In this study, we found that sub-genotype VIId2 remains the predominant genotype VII NDVs from 2011 to 2013, and all of the NDVs isolated from 2012 to 2013 were located in this clade.
The HN protein of NDV plays an important role in immune protection against virus infection, and the variation of the antigenic epitope would expand the antigenic difference. And analysis of the role of site 347 in antigenicity has confirmed that the E347K variation would intensify the antigenic difference and may increase the risk of vaccine breakdown [17, 18]. The isolation rate of strains with the E347K variation is increasing yearly, as previous reported. More than 36 % of the virus strains (20/55) were isolated in commercial layer flocks, which have resulted in losses in egg production. Cho et al. [18] found that the Korean variation strains have also caused losses in egg production. Moreover, more than 75 % (15/20) of the NDV strains isolated from layer flocks were the E347K-variation strains. The high isolation rate indicates that the E347K-variation strains can infect chickens under immune pressure because the layers were all well-vaccinated in China. Both site 347 and site 362 locate around the receptor-binding pocket, and we found of E347K, G362A co-variant strains, whereas Cho et al. also identified two strains with the E347K and G362R co-variant ones. However, the role of the co-variation remains unknown [6].
At present, LaSota is widely and frequently used to prevent NDV infection in the poultry industries in China; however, virulent NDV infections continue to occur in the well-vaccinated chicken flocks [12, 19]. Moreover, both cross HI and cross-neutralization tests were performed, and the R values of all of the strains were lower than 0.5. However, the R values between the E347K-variant strains and LaSota were lower than that found for the 347E/G strains, which indicate that the E347K variation would expand the antigenic difference with the vaccine strain. Therefore, the results of the cross-neutralization and cross HI experiments may also be consistent with the higher isolation rate of the E347K-variant strains, which demonstrated significant antigenic differences between the vaccine strain and the predominant variant strains should be responsible for the outbreaks of ND in China.
In addition, the protection efficiency of LaSota against the E347K variation strains in China is not well-elucidated. In this study, the vaccinated SPF chickens were fully protected against morbidity and mortality, but virus shedding was not stopped. Our data suggest that the LaSota inactivated vaccine cannot protect chickens from virus shedding when infected with the variant NDVs, and the variant strain challenged group showed higher virus isoation rate which may be one of the reasons for the circulation of sub-genotype VIId variation strains in vaccinated chicken flocks.
In conclusion, our findings indicate that the sub-genotype VIId NDVs with E347K variation in the HN protein are predominant in eastern China and that this variation would intensify the antigenic difference with the vaccine strains. Moreover, the vaccine strain LaSota has been used for more than 40 years and can hardly protect chickens effectively from virus shedding when infected with the circulation strains. Therefore, new NDV vaccines closely related to the prevalent genotype VIId viruses should be developed to control ND in China.
Conclusions
Currently, sub-genotype VIId NDVs are the prevalent virulent strains circulating among vaccinated chicken flocks in Eastern China. Our findings indicate that the E347K variation in HN protein would expand the antigenic difference with LaSota, which may be responsible for the increasing isolation rate of these strains from vaccinated chickens.
Methods
Virus isolation and antiserum preparation
Fifty-five virulent NDVs were isolated from 190 clinical samples from different flocks in Eastern China from 2011 to 2013 under the permission of the flock owners. All of the viruses were plaque-purified three times on primary chicken embryo fibroblasts (CEF) and proliferated in 9-10-day-old specific-pathogen-free (SPF) chicken embryos [20]. The virus stocks grown in allantoic fluids were stored at −70 °C until use.
SPF chickens (five in each group) were vaccinated with inactivated, oil-emulsion derived from LaSota and the strains listed in Table 2. The serum for the five birds vaccinated with the same vaccine were pooled together at three weeks post-vaccination and stored at −70 °C until use.
All of the SPF chicken embryos and the SPF chickens used in this study were bought from Beijing Merial Vital Laboratory Technology Co., LTD.
Viral RNA purification, RT-PCR, and sequencing of the F and HN genes
Virus preparation, viral RNA extraction and the initial RT reaction were performed as described previously [21]. Based on the published F and HN gene sequences in the GenBank database, we designed two pairs of primers (Table 6) for amplifying the F and HN genes of the virulent NDV isolates. And we also designed a pair of primers based one the same region of all NDV F genes which could be used for NDV identification. PCR products possessing the expected length were purified and sequenced by Sangon (Shanghai, China).
Sequence analysis
Nucleotide sequence editing, analysis and prediction of the amino acid sequences for both the F and HN proteins and alignments were conducted using the Clustal W multiple alignment method in the MegAlign program of the Lasergene package (DNASTAR Inc., Madison, WI, USA). Phylogenetic trees based on the F and HN genes were constructed using the MEGA 5 program (Version 5.2) with the maximum likelihood method algorithm [22]. In addition to the 55 strains collected in this study, 31 previously reported F and HN gene sequences representing different genotypes were included for comparison, and the accession numbers of each of these NDVs are shown in the phylogenetic trees [7, 16, 21].
Pathogenicity tests
The intracerebral pathogenicity index in 1-day-old chicks and the mean death time in 9 to 11-day-old SPF chicken embryos were determined for some of the isolates in this study as Liu did previously [7].
Cross hemagglutination inhibition (HI) test and virus neutralization test
To measure the antigenic difference between the vaccine strain LaSota and the isolated strains, we selected ten strains (One harboring 347G, four harboring 347E, and five harboring 347 K) and performed cross HI and virus neutralization test as describe by Cho and Li [8, 23]. Briefly, virus-serum mixtures were inoculated into the allantoic cavity of 10-day-old SPF chicken embryos 0.2 mL per sample. At the same time, blank controls and no-serum control samples were prepared by inoculating with an equal volume of PBS or virus. Eight days after incubation, the virus present in the chorioallantoic sac was titrated by calculating the 50 % virus neutralization endpoint. Virus neutralizing activity was determined relative to the no-serum control. The antigenic relatedness of the predominant strains and LaSota strain was expressed using the R value, as described by Archetti and Horsfall [24]. The following formula was used: \( \mathrm{R}\kern0.5em =\kern0.5em \sqrt{r1*r2} \), where r1 is the titer of strain A with antiserum B divided by the titer of strain A with antiserum A, and r2 is the titer of strain B with antiserum A divided by the titer of strain B with antiserum B. A result of 0.67 ≤ R ≤ 1.5 indicates no significant antigenic difference between the two viruses, whereas 0.5 ≤ R ≤ 0.67 indicates a minor antigenic difference between the two viruses. An R value of R < 0.5 indicates a major antigenic difference between the two virus strains.
Test of the protective efficacy of the LaSota strain against the variant strains
To determine whether the vaccine strain LaSota with a low antibody level can fully protect against the variant strains, two strains, namely, JS-22-11-Ch (347E, 362G) and JS-14-12-Ch (347 K, 362A), were selected, and a locally produced, inactivated, oil-emulsion vaccine derived from the LaSota strain (109.3EID50) was used in this study [25]. Twenty-five birds were immunized via the intramuscular route with a single dose (0.1 ml) of the LaSota vaccine. Fifteen birds were not vaccinated and served as a control group. Three weeks post-vaccination, 20 vaccinated birds were chosen and divided into two groups of 10. The birds in the La-JS22 group were challenged with 106.0 EID50/100 μl JS-22-11-Ch (n = 10) via the eye drop and intranasal route (ED/IN route; 100 μl), and those in the La-JS14 group were challenged with JS-14-12-Ch (n = 10) via same (100 μl) route (ED/IN route). The unvaccinated birds were also challenged with JS-22-11-Ch (n = 5) or JS-14-12-Ch (n = 5) as described above and were grouped in the Control-JS22 and Control-JS14 groups. Ten birds in the Control-La and Control-PBS groups (five LaSota-vaccinated birds and five unvaccinated birds) were challenged with the same volume (100 μl) of PBS via the same route. All of the birds were monitored daily for overt clinical signs (depression, respiratory signs, diarrhea, etc.) and mortality. Tracheal and cloacal swabs were collected from birds at days 3, 5, and 7 post-challenge (pc) for virus isolation.
Abbreviations
CEF, chicken embryo fibroblasts; F, fusion protein; HI, hemagglutination inhibition; HN, hemagglutinin–neuraminidase; L, large polymerase protein; M, matrix protein; ND, Newcastle disease; NDV, Newcastle disease virus; NP, nucleocapsid protein; P, phosphoprotein; SPF, specific-pathogen-free
References
Aldous EW, Alexander DJ. Detection and differentiation of Newcastle disease virus (avian paramyxovirus type 1). Avian Pathol. 2001;30:117–28.
Mayo MA. A summary of taxonomic changes recently approved by ICTV. Arch Virol. 2002;147:1655–63.
Mayo MA. Virus taxonomy - Houston 2002. Arch Virol. 2002;147:1071–6.
Peeters BP, de Leeuw OS, Verstegen I, Koch G, Gielkens AL. Generation of a recombinant chimeric Newcastle disease virus vaccine that allows serological differentiation between vaccinated and infected animals. Vaccine. 2001;19:1616–27.
Iorio RM, Syddall RJ, Sheehan JP, Bratt MA, Glickman RL, Riel AM. Neutralization map of the hemagglutinin-neuraminidase glycoprotein of Newcastle disease virus: domains recognized by monoclonal antibodies that prevent receptor recognition. J Virol. 1991;65:4999–5006.
Cho SH, Kwon HJ, Kim TE, Kim JH, Yoo HS, Kim SJ. Variation of a newcastle disease virus hemagglutinin-neuraminidase linear epitope. J Clin Microbiol. 2008;46:1541–4.
Wu S, Wang W, Yao C, Wang X, Hu S, Cao J, Wu Y, Liu W, Liu X. Genetic diversity of Newcastle disease viruses isolated from domestic poultry species in Eastern China during 2005–2008. Arch Virol. 2010;156:253–61.
Cho SH, Kim SJ, Kwon HJ. Genomic sequence of an antigenic variant Newcastle disease virus isolated in Korea. Virus Genes. 2007;35:293–302.
Choi KS, Kye SJ, Kim JY, To TL, Nguyen DT, Lee YJ, Choi JG, Kang HM, Kim KI, Song BM, Lee HS. Molecular epidemiology of Newcastle disease viruses in Vietnam. Trop Anim Health Prod. 2014;46:271–7.
Kwon HJ, Cho SH, Ahn YJ, Seo SH, Choi KS, Kim SJ. Molecular epidemiology of Newcastle disease in Republic of Korea. Vet Microbiol. 2003;95:39–48.
Umali DV, Ito H, Suzuki T, Shirota K, Katoh H, Ito T. Molecular epidemiology of Newcastle disease virus isolates from vaccinated commercial poultry farms in non-epidemic areas of Japan. Virol J. 2013;10:330.
Zhang YY, Shao MY, Yu XH, Zhao J, Zhang GZ. Molecular characterization of chicken-derived genotype VIId Newcastle disease virus isolates in China during 2005–2012 reveals a new length in hemagglutinin-neuraminidase. Infect Genet Evol. 2014;21:359–66.
Rui Z, Juan P, Jingliang S, Jixun Z, Xiaoting W, Shouping Z, Xiaojiao L, Guozhong Z. Phylogenetic characterization of Newcastle disease virus isolated in the mainland of China during 2001–2009. Vet Microbiol. 2010;141:246–57.
Zhang S, Wang X, Zhao C, Liu D, Hu Y, Zhao J, Zhang G. Phylogenetic and pathotypical analysis of two virulent Newcastle disease viruses isolated from domestic ducks in China. PLoS One. 2011;6, e25000.
Wang J, Liu H, Liu W, Zheng D, Zhao Y, Li Y, Wang Y, Ge S, Lv Y, Zuo Y, et al. Genomic Characterizations of Six Pigeon Paramyxovirus Type 1 Viruses Isolated from Live Bird Markets in China during 2011 to 2013. PLoS One. 2015;10, e0124261.
Ebrahimi MM, Shahsavandi S, Moazenijula G, Shamsara M. Phylogeny and evolution of Newcastle disease virus genotypes isolated in Asia during 2008–2011. Virus Genes. 2012;45:63–8.
Hu S, Wang T, Liu Y, Meng C, Wang X, Wu Y, Liu X. Identification of a variable epitope on the Newcastle disease virus hemagglutinin-neuraminidase protein. Vet Microbiol. 2010;140:92–7.
Cho SH, Kwon HJ, Kim TE, Kim JH, Yoo HS, Park MH, Park YH, Kim SJ. Characterization of a recombinant Newcastle disease virus vaccine strain. Clin Vaccine Immunol. 2008;15:1572–9.
Qin ZM, Tan LT, Xu HY, Ma BC, Wang YL, Yuan XY, Liu WJ. Pathotypical characterization and molecular epidemiology of Newcastle disease virus isolates from different hosts in China from 1996 to 2005. J Clin Microbiol. 2008;46:601–11.
Harper DR. Anovel plaque assaysystem for paramyxoviruses. J Virol Methods. 1989;25.
Liu XF, Wan HQ, Ni XX, Wu YT, Liu WB. Pathotypical and genotypical characterization of strains of Newcastle disease virus isolated from outbreaks in chicken and goose flocks in some regions of China during 1985–2001. Arch Virol. 2003;148:1387–403.
Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol. 2007;24:1596–9.
Li ZJ, Li Y, Chang S, Ding Z, Mu LZ, Cong YL. Antigenic variation between Newcastle disease viruses of goose and chicken origin. Arch Virol. 2010;155:499–505.
Archetti I, Horsfall Jr FL. Persistent antigenic variation of influenza A viruses after incomplete neutralization in ovo with heterologous immune serum. J Exp Med. 1950;92:441–62.
Hu Z, Hu S, Meng C, Wang X, Zhu J, Liu X. Generation of a genotype VII Newcastle disease virus vaccine candidate with high yield in embryonated chicken eggs. Avian Dis. 2011;55:391–7.
Acknowledgements
The authors would like to recognize Mrs. Liu Huimou, Ding Jihong and Gu kaihong for their significant contributions to the success of this study.
Funding
This work was supported by Chinese Special Fund for Agro-Scientific Research in the Public Interest (201303033, 201003012), the Earmarked Fund for Modern Agro-Industry Technology Research System (nycytx-41-G07), National Natural Science Foundation of China (31172338), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Availability of data and materials
All the data supporting the findings is contained within the manuscript.
Authors’ contributions
JZ and SH carried out the study design, phylogenetic analysis, sequence alignment, and drafted the manuscript. HX contributed for RNA preparation and RT-PCR. JL, ZZ participated in the whole procedure of the virus isolation and pathogenicity tests. XW contributed to the design of the study and revision of the manuscript. XL conceived of the study, provided consultation and coordination, and helped to draft the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent to publish
Not applicable.
Ethics approval and consent to participate
Experimental protocols were approved and conducted by the Laboratory Animal Monitoring Committee of Jiangsu province of China. All experiments involving live viruses and animals were housed in negative-pressure isolators with HEPA filters. All samples were collected under the permission of the flock owners.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Zhu, J., Hu, S., Xu, H. et al. Characterization of virulent Newcastle disease viruses from vaccinated chicken flocks in Eastern China. BMC Vet Res 12, 113 (2016). https://doi.org/10.1186/s12917-016-0732-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-016-0732-6