
Abstract
Background
Host-associated gut microbial communities are key players in shaping the fitness and health of animals. However, most current studies have focused on the gut bacteria, neglecting important gut fungal and archaeal components of these communities. Here, we investigated the gut fungi and archaea community composition in Large White piglets using shotgun metagenomic sequencing, and systematically evaluated how community composition association with gut microbiome, functional capacity, and serum metabolites varied across three weaning periods.
Results
We found that Mucoromycota, Ascomycota and Basidiomycota were the most common fungi phyla and Euryarchaeota was the most common archaea phyla across individuals. We identified that Methanosarcina siciliae was the most significantly different archaea species among three weaning periods, while Parasitella parasitica, the only differential fungi species, was significantly and positively correlated with Methanosarcina siciliae enriched in day 28 group. The random forest analysis also identified Methanosarcina siciliae and Parasitella parasitica as weaning-biased archaea and fungi at the species level. Additionally, Methanosarcina siciliae was significantly correlated with P. copri and the shifts of functional capacities of the gut microbiome and several CAZymes in day 28 group. Furthermore, characteristic successional alterations in gut archaea, fungi, bacteria, and serum metabolites with each weaning step revealed a weaning transition coexpression network, e.g., Methanosarcina siciliae and P. copri were positively and significantly correlated with 15-HEPE, 8-O-Methyloblongine, and Troxilin B3.
Conclusion
Our findings provide a deep insight into the interactions among gut archaea, fungi, bacteria, and serum metabolites and will present a theoretical framework for understanding gut bacterial colonization and succession association with archaea during piglet weaning transitions.
Similar content being viewed by others


Background
The gut microbiome is a complex ecosystem comprised of interacting communities of bacteria, fungi, viruses, and archaea [1]. There is a great deal of evidence, for instance, that the intestinal microbiome plays crucial roles in pig health, and is linked with daily growth and body weight [2], nutrient processing and energy harvesting [3], and fat storage [4]. However, the majority of these findings attributed changes in porcine health to bacteria, which are not the only components of gut microbiomes. To date, very little work has been done on the role of the fungal and archaeal microbiota components.
There are over 3.8 million species of fungi, which display an immense diversity of life forms, nutritional strategies, and associations with other organisms [5, 6]. For example, infections by parasitic eukaryotes result in decreased allergic and autoimmune disease prevalence [7] and have been used for therapeutic interventions in that context [8, 9]. The fungus Candida can decompose starch (i.e., carbohydrates in foods) into monosaccharides, providing the raw materials for bacterial fermentation [10]. Furthermore, recent studies have demonstrated that some fungi, like Candida albicans, can cause disease under the right circumstances, including failure to thrive [11, 12]. However, we still know little about the role that eukaryotes play within the host microbiome and their impacts on the host health.
Archaea are single-celled prokaryotes with distinct cellular characteristics that separate them from bacteria and eukaryotes, such as lack of peptidoglycan and D-glycerol esters or fatty acids [13]. Some archaeal species are mesophilic [14] and others are stable commensals of the gastrointestinal tract where they perform functions such as methanogenesis, transformation of heavy metals, trimethylamine metabolism, and immune modulation [15,16,17,18]. The most common gut archaea is Methanobrevibacter smithii [19, 20], an obligate carbon dioxide (CO2) reducing species that produces methane from side products of bacterial fermentation [21]. Methanogens drive effective degradation of organic substances and consequentially, M. smithii plays a key role in interspecies hydrogen transfer by maintaining syntrophic relationships with bacterial populations [22].
In piglets, the process of weaning presents a unique challenge to porcine gut physiology [23]. To date, only a few studies have examined the development of the gut fungi and archaea during the suckling and weaning periods [12, 24,25,26]. However, relatively little is known about the gut microbiome interactions (e.g. fungi-archaea interactions) and host-microbial interactions (e.g. archaea-serum metabolites interactions) during the weaning transition. In this study, we collected the feces samples from piglets at three different ages, 14, 21, and 28 days during weaning periods. Then, shotgun metagenomic sequencing was performed on fecal samples to comprehensively characterize porcine gut fungi and archaea composition during weaning periods. Furthermore, we constructed a co-occurrence network based on the relative abundances of differential gut archaea, fungi and bacteria species and serum metabolites during the three weaning periods to further uncover how these signatures modulated host metabolism.
Results
The taxonomic characterization of swine gut fungi and archaea
The gut fungi and archaea of five Large White piglets (two males and three females) across three age strata were studied. Fresh fecal sampling continued for all five piglets at 14 days (day 14 group), 21 days (day 21 group, the day of weaning), and 28 days (day 28 group) of age. To taxonomically characterize the swine gut fungi and archaea, we performed shotgun metagenomic sequencing on all 15 fecal samples. In total, we generated ~ 3,232 million base-pairs (bp). A sequence assembly analysis of the samples produced 1,566,160 contigs with an average length of 2,090 bp and an average N50 and N90 length of 4,599 bp and 707 bp, respectively (Supplementary Table 1). We determined the phylogenetic composition of the fecal microbiota by blasting against the National Center for Biotechnology Information (NCBI) non-redundant (NR) database. The results showed that 98.46% non-redundant genes were assigned to the bacteria super kingdom, whereas 0.58% genes, 0.05% genes and 0.91% genes were annotated as viruses, eukaryotes and archaea, respectively.
We then identified the taxonomic composition of swine gut fungi and archaea at the phylum, family and species levels. The Mucoromycota (62.94%—97.56%), Ascomycota (0.89%—31.06%) and Basidiomycota (1.30%—7.63%) were the most common fungal phyla present across individuals (Supplementary Fig. 1A). The top 3 most common fungal families were Mucoraceae, Rhizopodaceae, and Neocallimastigaceae, with an average abundance across samples of 37.68%, 28.96% and 16.33%, respectively (Fig. 1). A total of 23 species were found to be present across all 15 samples, with Parasitella parasitica being the most abundant fungi species (Supplementary Fig. 2A). For archaea, we identified 6 phyla, 30 families and 36 species present in all 15 samples. Euryarchaeota was the most abundance phylum (Supplementary Fig. 1B), and Methanobacteriaceae, Methanosarcinaceae, and Candidatus Methanomethylophilaceae were the most abundant families (Fig. 2). Moreover, Methanobrevibacter smithii was the most abundance species, followed by Candidatus Methanomethylophilus alvus (Supplementary Fig. 2B).

Categories and relative abundance of gut fungi at the family level by shotgun metagenomic sequencing data for all tested samples

Categories and relative abundance of gut archaea at the family level by shotgun metagenomic sequencing data for all tested samples
Variations in the gut fungi and archaea community across three weaning periods
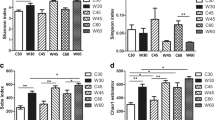
To identify the differences in gut fungi and archaea at the species level, we performed linear discriminant analysis (LDA) effect size (LEfSe) analysis on our metagenomic sequencing data. Results showed that four archaea species were significantly different in abundance across the three weaning periods (Fig. 3A), one of which was significantly elevated in the day 28 group and three of which were significantly elevated in the day 21 group. For example, Methanosarcina siciliae was enriched in the day 28 group, and Methanolobus psychrophilus, Thermoplasmatales archaeon-BRNA1, and Thermoplasmata archaeon were enriched in the day 21 group. On the other hand, only one fungi species, Parasitella parasitica, differed significantly in abundance across the three weaning periods, enriching in the day 28 group (Fig. 3B). A random forest analysis was then conducted to examine our ability to discriminate among the weaning periods based on fecal microbiota metagenomic sequencing of archaea (Fig. 3C) and fungi (Fig. 3D) species. As was found in LEfSe analysis, Methanosarcina siciliae, Methanolobus psychrophilus, and Thermoplasmata archaeon (archaea) and Parasitella parasitica (fungus) differed significantly among the three weaning periods and could therefore be used to distinguish among the three weaning periods with moderate to high diagnostic accuracy (area under the curve (AUC): 86% (Fig. 4A), 76% (Supplementary Fig. 3A), 60% (Supplementary Fig. 3B), and 84% (Fig. 4B), respectively).

Changes in gut archaea and fungi composition based on metagenomic sequencing results across the three weaning periods. (A) Differences between gut archaea species. LDA score ≥ 2 was set as the threshold. (B) Differences between gut fungi species. LDA score ≥ 2 was set as the threshold. (C) Discrimination of samples from different weaning periods based on the gut archaea species level from a Random Forest analysis. (D) Discrimination of samples from different weaning periods based on the gut fungi species level from a Random Forest analysis

Receiver operating curve (ROC) for Methanosarcina siciliae and Parasitella parasitica. (A) Methanosarcina siciliae. The AUC was 86% with the 95% CI of 60.3 – 100%. (B) Parasitella parasitica. The AUC was 84% with the 95% CI of 56.84 – 100%
The archaea and eukaryotes associated with bacterial colonization and succession
In our previous study, 17 bacterial species (e.g. P. copri), 8 KEGG functional terms and 21 CAZymes were identified to have significantly different abundances among the three weaning periods, and the abundance of bacteria P. copri played an important role in mitigating piglet adaptation during the weaning transition and was correlated with KEGG pathways and CAZymes [27]. Here, we used a Spearman correlation analysis to determine whether the archaea and eukaryotes are associated with bacterial colonization and succession, especially the bacteria P. copri. However, only one archaea (Methanosarcina siciliae) was found to be positively correlated with P. copri (Fig. 5A). The abundance of Methanosarcina siciliae was markedly higher in the day 28 group (P < 0.001, FDR) and lower in day 14 group (P < 0.001, FDR) compared to that in day 21 group (Fig. 5B). Furthermore, Methanosarcina siciliae abundance was positively correlated with most differential KEGG pathways except cationic antimicrobial peptide (CAMP) resistance (ko01503) pathway (Supplementary Fig. 4A). The sulfur relay system (ko04122) was positively correlated with all of the differential archaea and fungi species (Supplementary Fig. 4A). In terms of CAZymes enrichment, most of differential CAZymes were positively correlated with Methanosarcina siciliae abundance (Supplementary Fig. 4B), indicating the contribution of the archaea and fungi species to changes in CAZymes.

Levels of gut archaea and fungi species found to be significantly correlated with gut bacterial species, and the absolute abundances of Methanosarcina siciliae. (A) The heat maps showing the gut archaea and fungi species relationship with gut bacterial species. The X-axis represents the bacterial species. The Y-axis indicates the gut archaea and fungi species. * P < 0.05, ** P < 0.01, and *** P < 0.005. (B) The absolute abundances of Methanosarcina siciliae based on metagenomic sequencing results among the three weaning periods. * P < 0.05, ** P < 0.01, and *** P < 0.005
Co-occurrence analysis among the gut bacteria, archaea, fungi and serum metabolites
Previously, we identified a total of 15 metabolite features showing distinct enrichment patterns among the three weaning periods, including15-HEPE, 8-O-Methyloblongine, and Troxilin B3 that were enriched in the day 28 group [27]. Here, we explored potential correlations in abundances among these differential gut archaea, fungi, bacterial species and serum metabolites using a co-occurrence analysis. Bacterial and archaea species were found to form strong and broad co-occurring relationships with serum metabolites; fungi species, on the other hand, displayed only mild correlations with bacterial species, archaea species, and serum metabolites (Fig. 6). Within this coexpression network, bacterial species were clustered in two primary groups (cluster 1 and cluster 3). Serum metabolites also clustered into two groups (cluster 5 and cluster 6). Of note, the P. copri, Methanosarcina siciliae and Parasitella parasitica were positively correlated with each other’s, and found in cluster 2, while gut archaea species were mostly found in cluster 4. Furthermore, P. copri and Methanosarcina siciliae were positively and significantly correlated with 15-HEPE, 8-O-Methyloblongine, and Troxilin B3, indicating a synergistic and niche-related relationship.

A co-occurrence network constructed from the relative abundances of differential bacterial species, archaea species, fungi species and serum metabolites. Purple, green and blue dots indicate the day 14 group, day 21 group and day 28 group, respectively. Edges between nodes indicate Spearman’s negative (light gray) or positive (light red) correlation
Discussion
Although gut fungi and archaea are essential member of the microbiota in animals, they have been neglected in the past years [28, 29]. Very limited number of previous studies have examined the composition of gut fungi and archaea. Of these, only a handful of studies have focused on how gut fungi and archaea communities develop and change during the piglet suckling and weaning periods [12, 24,25,26]. Here, we performed shotgun metagenomic sequencing to comprehensively characterize porcine gut fungi and archaea during the piglet weaning periods. Our results indicate that gut fungi and archaea communities are highly dynamic during suckling and weaning.
Consistent with previous studies, Ascomycota and Basidiomycota were the two most dominant phyla of fungi in pigs [30, 31], followed by Mucoromycota. Mucoromycota, characterized by the production of asexual aplanospores, fusion of gametangia to produce zygospores, and walls of chitin and chitosan [32, 33], is of interest because it is one of the few fungi interacting with bacteria. Fungal-bacteria interactions are the focus of a new field of microbiology with relevance to microbial ecology, host health and biotechnology [34]. For the archaea, Euryarchaeota was the most abundance phyla, followed by Methanobacteriaceae, Methanosarcinaceae, and Candidatus Methanomethylophilaceae. The significant role we found for Euryarcheaota in pig microbiomes suggests a previously underestimated role in human health and disease [35]. In addition, these sequences are similar to a number of clone sequences recovered from pig feces and cattle rumens [36], raising the possibility that a novel group of Euryarchaeota has adapted to live in the animal intestinal tract [35].
Methanosarcina siciliae and Parasitella parasitica were found to be the most significantly different archaea and fungi species among three weaning periods, respectively. Random forest analysis also identified Methanosarcina siciliae and Parasitella parasitica as a weaning-biased archaea and fungi at the archaea and fungi species level, respectively. Methanosarcina siciliae is known to be able to degrade dimethylsulfide and methane thiol to hydrogen sulfide, methane, and carbon dioxide [37, 38]. Parasitella parasitica, a facultative mycoparasite of zygomycetous fungi, forms cytoplasmic fusions with its hosts during infection [39], infecting many different mucoralean species from nearly all major genera [40, 41]. Moreover, Methanosarcina siciliae and Parasitella parasitica could be used to distinguish the three weaning periods with high diagnostic accuracy, suggesting that Methanosarcina siciliae and Parasitella parasitica are important species for mitigating piglet adaptation during the weaning transition.
The archaea were found to be ubiquitously present in the piglet gastrointestinal tract microbiome across the three weaning periods. In particular, Methanosarcina siciliae, the only differential archaea species, was significantly and positively correlated with the bacteria P. copri. A previous study focused on neonatal colonization found that archaea were transient and found almost exclusively only within the first few weeks of life [42]. Other studies suggest that archaea are core members of the adult gastrointestinal tract microbiome [43, 44]. Interesting, we also observed that KEGG pathways and CAZymes related to energy metabolism were positively associated with Methanosarcina siciliae. However, human archaeal commensal has a highly restricted energy metabolism [45], which makes it a specialized member of the gastrointestinal microbiome [44]. Furthermore, the P. copri and Methanosarcina siciliae were positively correlated with 15-HEPE, 8-O-Methyloblongine, and Troxilin B3. These serum metabolites may provide a mechanism for anti-inflammatory, and potentially act as a biomarker for food consumption and release of arachidonic acid and diacylglycerols [46, 47]. These results indicate that bacterial colonization and succession are associated with archaea and are thereby among the earliest colonizers of the piglet gastrointestinal tract microbiome.
Conclusions
In this study, we found that gut fungi and archaea composition were significantly changed through time during the weaning processes. Methanosarcina siciliae and Parasitella parasitica were found to be the most significantly different archaea and fungi species, respectively, which were also identified by random forest analysis as weaning-biased archaea and fungi at the species level. Methanosarcina siciliae was significantly correlated with the changes of P. copri abundance, KEGG pathways and CAZymes. Furthermore, Methanosarcina siciliae and P. copri were positively and significantly correlated with 15-HEPE, 8-O-Methyloblongine, and Troxilin B3, indicating alterations of gut bacteria, archaea, fungi, and serum metabolites generated a characteristic weaning transition coexpression network. Overall, our results reveal important insights into the interactions among gut bacteria, archaea, fungi and serum metabolites and lead to beneficial understanding of gut bacterial colonization and succession associated with archaea.
Materials and methods
Experimental animals and sample collection
The gut microbiome of five Large White piglets (two males and three females) from three age strata was studied. All piglets stayed with their dams during the suckling period and were allowed to nurse freely until weaning began at 21 days old. Once weaned, piglets were transferred into the same pen and were provided with commercial formula diet and clean water ad libitum. According to previous studies [25, 48, 49], seven days before and after weaning day were chosen for other two periods. Fresh feces from all piglets were collected from each animal’s anus by rectal massage at 14 days of age (day 14 group), 21 days (the day of weaning, day 21 group), and 28 days of age (day 28 group). All piglets had no obvious disease or diarrhea and received no probiotic or antibiotic therapy during the period from birth to the end of this study. Fecal samples were immediately snap-frozen in liquid nitrogen for transportation and stored at − 80 °C for later use.
Microbial DNA extraction
Microbial DNA was extracted from fecal samples using the QIAamp Fast DNA Stool Mini Kit (Qiagen, Germany) according to the manufacturer’s instructions. The concentration of DNA was determined using a Nanodrop-1000 (Thermo Scientific, USA) and the DNA purity was determined by 0.8% (w/v) agarose gel electrophoresis. All DNA samples were stored at -20 °C until further processing.
Metagenomic sequencing analysis
Metagenomic sequencing of the 15 fecal microbial DNA samples was performed using a Novaseq-PE150 platform. Briefly, sequencing libraries were generated using NEB Next® Ultra™ DNA Library Prep Kit for Illumina (NEB, USA) by following manufacturer’s instructions, and index codes were added to attribute sequences to each sample. The clustering of the index-coded samples was performed on a cBot Cluster Generation System according to the manufacturer’s instructions. After cluster generation, the prepared libraries were sequenced on the Novaseq-PE150 platform and paired-end reads were generated. The raw data were preprocessed using Readfq in order to acquire clean data for subsequent analysis. As host pollution may exist in samples, the clean data was checked via blast against the host database using Bowtie2.2.4 software [50] to filter out reads that are of host origin. The clean data were then assembled and analyzed using SOAPdenovo software [51, 52]. The scaftigs were predicted the ORF using MetaGeneMark (V2.10) software, and length information shorter than 100 nt [51, 53] was filtered from the predicted results using the default parameters. For ORF predicted scaftigs, CD-HIT [54] software was used to reduce redundancy and create the unique initial gene catalogue. DIAMOND [55] software was used to blast the Unigenes with the bacterial, fungal, archaeal and viral sequences, all of which were extracted from the NCBI NR database. As each sequence may have multiple aligned results, we chose the aligned sequence result where the e value ≤ the smallest e value × 10 [56]. These sequences were used in the LCA algorithm which was applied to system classification of MEGAN [57] software to guarantee the species annotation information of sequences.
Statistical analysis
Sparse Correlation for Compositional data (SparCC) was employed to determine co-abundance (positive) and co-exclusion (negative) relationships among differential gut fungi, archaea, bacterial species, and serum metabolites based on their relative abundances [58]. Network analysis was performed and visualized using Cytoscape (version 3.6.1). To identify the different abundances among groups, linear discriminant analysis (LDA) and effect size (LEfSe) analysis were performed under the condition α = 0.05, with an LDA score of at least 2.0 [58]. Story’s FDR was used to correct the multiple tests.
Availability of data and materials
All of the data generated or analyzed during this study are available from the corresponding author on reasonable request.
Abbreviations
- AUC:
-
Area under the curve
- Bp:
-
Base-pairs
- CO2 :
-
Carbon dioxide
- NCBI:
-
National Center for Biotechnology Information
- NR:
-
Non-redundant
- CAZy:
-
Carbohydrate-Active enZYmes database
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- SparCC:
-
Sparse Correlations for Compositional
- LDA:
-
Linear discriminant analysis
- LEfSe:
-
Linear discriminant analysis effect size
References
Mahnic A, Rupnik M. Different host factors are associated with patterns in bacterial and fungal gut microbiota in Slovenian healthy cohort. PLoS ONE. 2018;13: e0209209.
Ramayo-Caldas Y, Mach N, Lepage P, Levenez F, Denis C, Lemonnier G, et al. Phylogenetic network analysis applied to pig gut microbiota identifies an ecosystem structure linked with growth traits. ISME J. 2016;10:2973–7.
Quan J, Cai G, Ye J, Yang M, Ding R, Wang X, et al. A global comparison of the microbiome compositions of three gut locations in commercial pigs with extreme feed conversion ratios. Sci Rep. 2018;8:4536.
Wall R, Ross RP, Shanahan F, O’Mahony L, O’Mahony C, Coakley M, et al. Metabolic activity of the enteric microbiota influences the fatty acid composition of murine and porcine liver and adipose tissues. Am J Clin Nutr. 2009;89:1393–401.
Hawksworth DL, Lucking R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. Microbiol Spectr. 2017;5. http://www.ncbi.nlm.nih.gov/pubmed/28752818.
Nilsson RH, Anslan S, Bahram M, Wurzbacher C, Baldrian P, Tedersoo L. Mycobiome diversity: high-throughput sequencing and identification of fungi. Nat Rev Microbiol. 2019;17:95–109.
Weinstock JV. Autoimmunity: The worm returns. Nature. 2012;491:183–5.
McFarland LV, Bernasconi P. A review of an innovative biotherapeutic agent. Microb Ecol Health Dis. 1993;6:157–71.
Williamson LL, McKenney EA, Holzknecht ZE, Belliveau C, Rawls JF, Poulton S, et al. Got worms? Perinatal exposure to helminths prevents persistent immune sensitization and cognitive dysfunction induced by early-life infection. Brain Behav Immun. 2016;51:14–28.
Iannotti EL, Kafkewitz D, Wolin MJ, Bryant MP. Glucose fermentation products in Ruminococcus albus grown in continuous culture with Vibrio succinogenes: changes caused by interspecies transfer of H 2. J Bacteriol. 1973;114:1231–40.
King J, Pana ZD, Lehrnbecher T, Steinbach WJ, Warris A. Recognition and Clinical Presentation of Invasive Fungal Disease in Neonates and Children. J Pediatric Infect Dis Soc. 2017;6:S12–21.
Summers KL, Frey JF, Ramsay TG, Arfken AM. The piglet mycobiome during the weaning transition: a pilot study1. J Anim Sci. 2019;97:2889–900.
Kandler O, Konig H. Cell wall polymers in Archaea (Archaebacteria). Cell Mol Life Sci. 1998;54:305–8.
Brochier-Armanet C, Boussau B, Gribaldo S, Forterre P. Mesophilic Crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nat Rev Microbiol. 2008;6:245–52.
Brugere JF, Borrel G, Gaci N, Tottey W, O’Toole PW, Malpuech-Brugere C. Archaebiotics: proposed therapeutic use of archaea to prevent trimethylaminuria and cardiovascular disease. Gut Microbes. 2014;5:5–10.
Blais Lecours P, Marsolais D, Cormier Y, Berberi M, Hache C, Bourdages R, et al. Increased prevalence of Methanosphaera stadtmanae in inflammatory bowel diseases. PLoS ONE. 2014;9: e87734.
Moissl-Eichinger C, Pausan M, Taffner J, Berg G, Bang C, Schmitz RA. Archaea Are Interactive Components of Complex Microbiomes. Trends Microbiol. 2018;26:70–85.
Borrel G, Brugere JF, Gribaldo S, Schmitz RA, Moissl-Eichinger C. The host-associated archaeome. Nat Rev Microbiol. 2020;18:622–36.
Miller TL, Wolin MJ, Conway de Macario E, Macario AJ. Isolation of Methanobrevibacter smithii from human feces. Appl Environ Microbiol. 1982;43:227–32.
Dridi B, Raoult D, Drancourt M. Archaea as emerging organisms in complex human microbiomes. Anaerobe. 2011;17:56–63.
Ferry JG. How to make a living by exhaling methane. Annu Rev Microbiol. 2010;64:453–73.
Hansen EE, Lozupone CA, Rey FE, Wu M, Guruge JL, Narra A, et al. Pan-genome of the dominant human gut-associated archaeon, Methanobrevibacter smithii, studied in twins. Proc Natl Acad Sci U S A. 2011;108(Suppl 1):4599–606.
Lallès J-P, Bosi P, Smidt H, Stokes CR. Weaning — A challenge to gut physiologists. Livest Sci. 2007;108:82–93.
Federici S, Miragoli F, Pisacane V, Rebecchi A, Morelli L, Callegari ML. Archaeal microbiota population in piglet feces shifts in response to weaning: Methanobrevibacter smithii is replaced with Methanobrevibacter boviskoreani. FEMS Microbiol Lett. 2015;362:fnv064.
Arfken AM, Frey JF, Summers KL. Temporal Dynamics of the Gut Bacteriome and Mycobiome in the Weanling Pig. Microorganisms. 2020;8:868.
Arfken AM, Frey JF, Ramsay TG, Summers KL. Yeasts of Burden: Exploring the Mycobiome-Bacteriome of the Piglet GI Tract. Front Microbiol. 2019;10:2286.
Xiong X, Liu X, Wang Z, Xu Q, Xu J, Rao Y. Identifying biomarkers of the gut bacteria, bacteriophages and serum metabolites associated with three weaning periods in piglets. BMC Vet Res. 2022;18:104.
Deng F, Peng Y, Zhang Z, Howe S, Wu Z, Dou J, et al. Weaning Time Affects the Archaeal Community Structure and Functional Potential in Pigs. Front Microbiol. 2022;13: 845621.
Deng F, Li Y, Peng Y, Wei X, Wang X, Howe S, et al. The Diversity, Composition, and Metabolic Pathways of Archaea in Pigs. Animals (Basel). 2021;11:2139.
Scupham AJ, Presley LL, Wei B, Bent E, Griffith N, McPherson M, et al. Abundant and diverse fungal microbiota in the murine intestine. Appl Environ Microbiol. 2006;72:793–801.
Limon JJ, Skalski JH, Underhill DM. Commensal Fungi in Health and Disease. Cell Host Microbe. 2017;22:156–65.
Hibbett DS, Binder M, Bischoff JF, Blackwell M, Cannon PF, Eriksson OE, et al. A higher-level phylogenetic classification of the Fungi. Mycol Res. 2007;111:509–47.
Adl SM, Simpson AG, Lane CE, Lukes J, Bass D, Bowser SS, et al. The revised classification of eukaryotes. J Eukaryot Microbiol. 2012;59:429–93.
Bonfante P, Desiro A. Who lives in a fungus? The diversity, origins and functions of fungal endobacteria living in Mucoromycota. ISME J. 2017;11:1727–35.
Horz HP, Conrads G. The discussion goes on: What is the role of Euryarchaeota in humans? Archaea. 2010;2010: 967271.
Mihajlovski A, Dore J, Levenez F, Alric M, Brugere JF. Molecular evaluation of the human gut methanogenic archaeal microbiota reveals an age-associated increase of the diversity. Environ Microbiol Rep. 2010;2:272–80.
Ni S, Boone DR. Catabolism of Dimethylsulfide and Methane Thiol by Methylotrophic Methanogens. In: Oremland RS, editor. Biogeochemistry of global change. Boston: Springer; 1993. https://doi.org/10.1007/978-1-4615-2812-8_42.
Deng J, Wei CH, Luo HB, Huang ZG, Wan SL. High-throughput sequencing of the archaeal community in cellar pit mud used in luzhou flavor liquor production. Modern Food ence and Technology. 2015;31:205–10.
Schultze K, Schimek C, Wostemeyer J, Burmester A. Sexuality and parasitism share common regulatory pathways in the fungus Parasitella parasitica. Gene. 2005;348:33–44.
Burgeff H. Untersuchungen qber Sexualitat und Parasitismus bei Mucorineen. Bot Abhandl. 1924;4:5–135.
Wo¨stemeyer J, Wo¨stemeyer A, Burmester A, Czempinski K. Gene transfer from the zygomycete Parasitella parasitica to its host: an evolutionary link between sex and parasitism. Eukaryotism and Symbiosis. 1997;145–52.
Palmer C, Bik EM, DiGiulio DB, Relman DA, Brown PO. Development of the human infant intestinal microbiota. PLoS Biol. 2007;5: e177.
Dridi B, Henry M, El Khechine A, Raoult D, Drancourt M. High prevalence of Methanobrevibacter smithii and Methanosphaera stadtmanae detected in the human gut using an improved DNA detection protocol. PLoS ONE. 2009;4: e7063.
Wampach L, Heintz-Buschart A, Hogan A, Muller EEL, Narayanasamy S, Laczny CC, et al. Colonization and Succession within the Human Gut Microbiome by Archaea, Bacteria, and Microeukaryotes during the First Year of Life. Front Microbiol. 2017;8:738.
Fricke WF, Seedorf H, Henne A, Kruer M, Liesegang H, Hedderich R, et al. The genome sequence of Methanosphaera stadtmanae reveals why this human intestinal archaeon is restricted to methanol and H2 for methane formation and ATP synthesis. J Bacteriol. 2006;188:642–58.
Miller CC, Ziboh VA, Wong T, Fletcher MP. Dietary supplementation with oils rich in (n-3) and (n-6) fatty acids influences in vivo levels of epidermal lipoxygenase products in guinea pigs. J Nutr. 1990;120:36–44.
Anton R, Camacho M, Puig L, Vila L. Hepoxilin B3 and its enzymatically formed derivative trioxilin B3 are incorporated into phospholipids in psoriatic lesions. J Invest Dermatol. 2002;118:139–46.
Frese SA, Parker K, Calvert CC, Mills DA. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome. 2015;3:28.
Holman DB, Gzyl KE, Mou KT, Allen HK. Weaning Age and Its Effect on the Development of the Swine Gut Microbiome and Resistome. mSystems. 2021;6:e0068221.
Zhao W, Wang Y, Liu S, Huang J, Zhai Z, He C, et al. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS ONE. 2015;10: e0117441.
Mach N, Berri M, Estelle J, Levenez F, Lemonnier G, Denis C, et al. Early-life establishment of the swine gut microbiome and impact on host phenotypes. Environ Microbiol Rep. 2015;7:554–69.
Konopka A. What is microbial community ecology? ISME J. 2009;3:1223–30.
Vo N, Tsai TC, Maxwell C, Carbonero F. Early exposure to agricultural soil accelerates the maturation of the early-life pig gut microbiota. Anaerobe. 2017;45:31–9.
Hamer HM, Jonkers D, Venema K, Vanhoutvin S, Troost FJ, Brummer RJ. Review article: the role of butyrate on colonic function. Aliment Pharmacol Ther. 2008;27:104–19.
De Filippis F, Pasolli E, Tett A, Tarallo S, Naccarati A, De Angelis M, et al. Distinct Genetic and Functional Traits of Human Intestinal Prevotella copri Strains Are Associated with Different Habitual Diets. Cell Host Microbe. 2019;25(444–53): e3.
Pedersen HK, Gudmundsdottir V, Nielsen HB, Hyotylainen T, Nielsen T, Jensen BA, et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature. 2016;535:376–81.
Flint HJ, Bayer EA, Rincon MT, Lamed R, White BA. Polysaccharide utilization by gut bacteria: potential for new insights from genomic analysis. Nat Rev Microbiol. 2008;6:121–31.
Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Metagenomic biomarker discovery and explanation. Genome Biol. 2011;12:R60.
Funding
This work was financially supported by the Doctoral Foundation of Nanchang Normal University (NSBSJJ2019009).
Author information
Authors and Affiliations
Contributions
XX conceived and designed the experiments, analyzed data, wrote and revised the manuscript. YR conceived and designed the experiments, and revised the manuscript. XT revised the manuscript. ZW analyzed the data. JG, YY and HW performed the experiments. XL conceived and designed the experiments, and revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The study was carried out in compliance with the ARRIVE guidelines. All animal work was conducted according to the guidelines for the care and use of experimental animals established by the State Council of the People’s Republic of China (Decree No. 2, 1988). This study was also approved by the Animal Care and Use Committee (ACUC) of Nanchang Normal University (No. NCNU2019-009).
Consent for publication
Not applicable.
Competing interests
All authors declare no conflicts of interest, financial or otherwise.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xiong, X., Rao, Y., Tu, X. et al. Gut archaea associated with bacteria colonization and succession during piglet weaning transitions. BMC Vet Res 18, 243 (2022). https://doi.org/10.1186/s12917-022-03330-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-022-03330-4