
Abstract
Circular RNA (circRNA), a new type of endogenous non-coding RNA, is abundantly present in eukaryotic cells, and characterized as stable high conservation and tissue specific expression. It has been generated increasing attention because of their close association with the progress of diseases. The liver is the vital organ of humans, while it is prone to acute and chronic diseases due to the influence of multiple pathogenic factors. Moreover, hepatocellular carcinoma (HCC) is the one of most common cancer and the leading cause of cancer death worldwide. Overwhelming evidences indicate that some circRNAs are differentially expressed in liver diseases, such as, HCC, chronic hepatitis B, hepatic steatosis and hepatoblastoma tissues, etc. Additionally, these circRNAs are related to proliferation, invasion, migration, angiogenesis, apoptosis, and metastasis of cell in liver diseases and act as oncogenic agents or suppressors, and linked to clinical manifestations. In this review, we briefly summarize the biogenesis, characterization and biological functions, recent detection and identification technologies of circRNA, and regulation network mechanism of circRNA in liver diseases, and discuss their potential values as biomarkers or therapeutic targets for liver diseases, especially on HCC.
Similar content being viewed by others
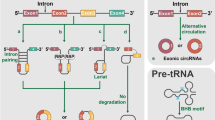


Introduction
CircRNA, as a type of endogenous non-coding RNA (ncRNA), was first found in the virus in 1976. But it was considered as one of the non-function by-products from splicing errors in a long time [1,2,3], thus, it was thought unlikely to play important role in biological processes. With the development of high-throughput sequencing technology and bioinformatics, circRNA has been found to be abundantly present in eukaryotic cells [4]. It is an abnormally stable RNA molecule by covalently closed loop structure, therefore the expression of circRNA is higher than linear mRNA, miRNA, and other types of RNA in whole blood, plasma, and platelets. Furthermore, circRNA exhibits longer half-time and more detectable than that of linear RNA by resisting to RNA exonuclease or RNase R. Hence, it is not surprising that circRNA could be ideal biomarker for disease [5, 6]. Besides, circRNA is involved in various physiological and pathological pathways by regulating gene expression and protein activity [7]. Increasing evidences indicate circRNAs have a strong association with diseases including systemic lupus erythematosus [8], disease of the central nervous system [9, 10], diabetes [11], cancers [12] and liver diseases [13], etc.
Liver disease accounts for significant morbidity, economic burden and costs [14], and leads to approximately 2 million deaths per year worldwide, of which one million deaths were from cirrhosis complications, and the other 1 million deaths were from viral hepatitis and hepatocellular carcinoma (HCC) [15]. Worldwide, liver cancer is the most common hepatic malignancies, ranking fifth in incidence rates and third in cause of cancer-linked deaths [16]. In addition, chronic infections with hepatitis B (HBV) were responsible for 33% of liver cancer death, alcohol for 30%, HCV for 21%, and other causes for 16% deaths in 2015 [17]. Unsatisfactorily, there is a lack of more effective diagnostic and prognostic markers besides α-fetoprotein (AFP) [18, 19]. More than 75% of patients with liver disease are often diagnosed in terminal-stage of HCC and the overall ratio of mortality to incidence is 0.95 [20, 21]. Moreover, although the 5-year overall survival rate is as high as 50%, more than 70% patients have a recurrence [22], which seriously affects the prognosis of HCC. Therefore, it is very urgent to elucidate the pathogenesis of HCC and to find valuable targets for diagnosis and treatment.
Growing evidences indicate that several dysregulated circRNAs are involved in HCC and play a vital role in many biological processes [23]. In this review, we briefly described the biogenesis, characterization, biological functions, detection and validation technologies of circRNAs. In addition, we introduced the network regulation mechanism of circRNAs in liver diseases. It not only shows that circRNAs play critical roles in the progression of liver diseases but also that they may be employed as new diagnostic and prognostic biomarkers.
Biogenesis and characterization of circRNA
Unlike linear RNA, circRNA forms as covalently closed loop structures with neither poly-adenylated tail nor 5′–3′ polarity via back-splicing. It can be simply classified into three categories according to their composition and sequence: exonic circRNA (ecircRNA), circular intronic RNA (ciRNA) and exon–intron circRNA (EIciRNA) (Fig. 1). Though the circularization mechanisms of circRNA have not been clarified. There are four mechanisms are suggested to explain the biogenesis of circRNA: (1) Lariat-driven circularization (exon skipping), during the splicing of pre-mRNA, the 3′ splice acceptor upstream of the exon interconnects with the 5′ splice donor downstream to produce a lasso structure containing the exon and intron, if the intron is removed, ecircRNAs or EIciRNAs are generated [24]. (2) Intron pairing-driven circularization, when there are opposite complementary sequences in the flanking introns of circRNA, upstream introns and downstream introns complement each other based on reverse repeat and complementary sequences, and introns are removed or retained to form ecircRNAs or EIcirRNAs, respectively [25]. (3) RNA-binding protein (RBP)-driven circularization, in this model, RBP can promote the interaction between upstream introns and downstream introns, resulting in the production of ecircRNAs or EIciRNA [26]. (4) Lariat introns-driven circularization, the formation of the lasson introns-driven circularization mainly refers to the combination of the 7-nt GU rich element near the 5 splicing point and the 11-nt C-rich element near the 3′ branching point to escape the degradation of the debranching enzyme and form a stable ciRNA [27].

Biogenesis and function of circRNA. a RBP-driven circularization; b Intron pairing-driven circularization; c Lariat-driven circularization; d Lariat introns-driven circularization; e Gene transcription regulation; f sponging miRNA; g protein interaction; h transcription regulation
CircRNA has the following specific characteristics. (1) In recent years, circRNA has been found to be abundant in almost all species, e. g., human [28], mouse [28], zebrafish [29] and plants [30], etc. It was reported that there are 1950 species of circRNAs in human more than that of nematode and mouse [31]. Additionally, circRNA can be detected in human saliva, blood, urine, and exosomes, which can be employed as novel and potential biomarkers [32]. (2) CircRNA is highly stable in eukaryotes and covalently closed, which can protect them from exonuclease-mediated degradation. (3) The expression of circRNA varied with time, space and tissue type. For instance, some neuro-associated circRNAs were found to increase with age in Drosophila [33]. There are also obvious differences in spatial expression, such as most ecircRNAs are found in the cytoplasm, and ciRNA is present in the nucleus. Besides, some circRNAs originate from human and mouse which are tissue-specific and mostly expressed in the brain [34]. (4) It is highly conserved in different species, Jeck et al. [35] found that there are 69 circRNAs in mouse testes are precisely homologous to human fibroblasts. These characteristics indicate that circRNA plays an important role in the occurrence and development of the disease and serves as potential biomarkers in bodily fluid for disease diagnosis.
Biological functions of circRNA
CircRNAs were found to have many potential biological functions on the basis of its characteristics, such as the following (Fig. 1): (1) Nuclear retained circRNAs can modulate transcription and splicing. Li [36] found that it participated in transcription regulation by interacting with U1 small nuclear ribonucleoproteins (U1snRNPs), the EIciRNA-U1 snRNP complexes bind to Pol II at the promoters of its parental genes to enhance gene expression. CircSEP3, as a nuclear retained circRNA roots in exon 6 of SEPALLATA3 in arabidopsis, was reported to regulate the splicing of its linear counterpart [37]. (2) CircRNA can act as miRNA sponge. The structure of circRNAs displayed numerous miRNA binding sites which assist the interactions with miRNAs [38]. CiRS-7 has been reported to contained 70 conserved binding sites of miR-7, and then it could increase expression level of miR-7 target by sponging miR-7 [39]. (3) circRNAs can perform its function by interacting with proteins, such as serving as protein sponge, protein scaffolding and protein recruitment, etc. [10, 40]. For example, circFOXO3 can promote cardiac senescence act as a scaffold to connect with the anti-senescent protein ID-1 and the transcription factor E2F1 as well as the anti-stress proteins FAK and hypoxia-inducible factor-1α (HIF1A) [41]. Besides, circRNA derived from human antigen R (HuR), an RNA binding protein (RBP) with AU-rich elements. And circHuR could directly interacted with CCHC-type zinc finger nucleic acid binding protein and inhibited its binding to HuR promoter, then reducing the expression level of HuR and restricting the progression of gastric cancer [42]. (4) circRNA can be translatable. Although most of the circRNAs haven’t been associated with ribosomes for translation, a small part of endogenous circRNAs are translatable, such as the human circHO which was found to encode protein and control the myoblast proliferation in human and murine and human [43]. Besides, recent studies have shown that N6-methyladenosine (m6A) modification can regulate the translation and degradation of circRNA, thus affecting the occurrence and development of tumors [44].
Detection and validation methods for circRNA
Technologies and methods to analyze and identify circRNA have greatly and considerably improved with the developments of sequencing technology and biochip technology; however, error and bias that influence accuracy are yet to be completely eliminated [45]. With advances in sequencing technology including RNA sequencing (RNA-Seq), next generation sequencing (NGS), microarrays etc., and bioinformatics tools consisting of circRNA annotations, circRNA identification and network analysis of ceRNA, the types and biological functions of circRNAs are expanding, such as biomarkers for diseases [46]. Common detection and verification methods of circRNA are introduced in this section (Fig. 2).

Copyright© 2021 Dovepress
Methods for assay of circRNA. a direct sequencing of a reverse transcribed circRNA typically employs a primer that is complementary to an exon. The sequencing reaction does not discriminate between linear RNA and circRNA, but bioinformatics analysis allows identification of circular junctions (red arrowhead); b Microarrays make use of probes that straddle unique and defined circularization junctions. Typically random primers are used to generate labeled cRNAs from circRNA templates. Probes span the circular junctions and hybridize to the cRNAs with higher Tms than the partly complementary sequences of linear mature RNA; c Reverse transcriptase (RT) quantitative PCR entails specific amplification of sequences derived from circRNAs by using primers that flank the circular junctions. The configuration of the amplifying primers is such that mature linear mRNA is not amplified. Reprinted with permission from Abdullah et al. [46]
Detection methods for circRNA
At present, RNA-Seq and microarrays are widely applied for circRNA detection. RNA-seq is primarily used for the discovery of novel circRNAs. The head-to-tail junctions are the only direct evidence for detecting circRNAs [47]. Generally, RNaseR is used to treat the total RNA to demonstrate the stability of circRNAs by eliminating linear RNAs, but not circRNAs. Then, microarray probes hybridized to the fluorescently labeled cRNA, which is from circRNA and tarted the head-to-tail junctions. As an extensive and alternative genome-wide assay, circRNA-specific microarray has been demonstrated great specificity and sensitivity compared to NGS; e. g., the related products of Arraystar (https://www.arraystar.com/) and Cofactor Genomics (https://cofactorgenomics.com/) [46]. In addition, circRNA-associated bioinformatics tools including circRNA_finder, find_circ, CIRCexplorer, CIRI and MapSplice, etc., can identify circRNA mainly based on the presence of backsplice junction-spanning sequencing reads. Recently, more and more circRNAs have been discovered in liver diseases through the above-mentioned technologies and have been proved to be differentially expressed by bioinformatics tools [48]. For example, Guan et al. [49] and Yu et al. [50] detected 1245 (756 up-regulation and 489 down-regulation) circRNAs and 257 differedtial expression circRNAs (DECs) (213 up-regulation and 44 down-regulation) from three pairs of HCC tissue and the microarray dataset GSE78520 by using microarray, respectively. RNA-seq can be employed for detection of circRNAs by RNA fragmentation, capture, sequencing, and subsequent computational analysis. Hu et al. [51] detected 72, 277 known circRNAs and identified 220 DECs by RNA-seq in 30 primary HCC tissues, including 15 HCC tissues with pulmonary metastasis after curative resection and 15 normal tissues without pulmonary metastasis. Moreover, further researches for DECs are contributed to finding the diagnostic markers and therapeutic targets for liver diseases. Among all of the 1245 DECs, Guan et al. [52] also reported that hsa_circ_0016788 had a highly potential diagnostic value and might be a promising biomarker of HCC. Qiao et al. [53] detected that the expression of hsa_circ_0003998 which was the most significantly upregulated among the 22 DECs (14 up-regulated and 8 down-regulated) in HCC tissues by RNA-seq. In addition, hsa_circ_0003998 was related to serum AFP level and clinicopathological factors, which suggesting it would be an early diagnostic biomarker of HCC.
Validation methods for circRNA
Based on the development of second-generation sequencing and bioinformatics analysis, it is possible to sequence hundreds of millions of short reads. DECs are required to be experimentally verified in the later stage [54]. Standard PCR-based methods including reverse transcription quantitative PCR (RT-qPCR), droplet digital PCR (dd PCR) and northern blot are considered as most widely employed methods for validation and more detailed quantitation of circRNAs [55,56,57]. Besides, Fluorescence in-situ hybridization is used to locate the distribution and abundance circRNA for further study of subsequent functions [58]. At present, a number of online databases have been established to analyze the information, regulatory network and the role of circRNA in diseases and other physiological processes, such as circBase, CIRCpedia, CircInteractome and Circ2Traits [59,60,61,62]. With increasing and continuous improvement of circRNA identification techniques and databases, the role of circRNAs will be more fully clarified.
CircRNA in liver diseases
Liver is a critical organ of the human body, it not only has many biological functions such as controlling metabolism, maintaining energy balance and detoxifying, etc. While, it has a powerful ability to regenerate and repair after injury [63, 64]. However, various pathogenic factors such as drug, alcohol, and virus damage the liver, resulting in acute or chronic liver disease and even HCC [65]. Some evidences suggest that circRNA is involved in regulating liver homeostasis and disease. The latest report showed [66] that 668 circRNAs are specifically expressed in liver tissue of six adult and fetal normal tissues. Li et al. [67] detected that 2412 circRNAs, including 159 circRNAs deriving from 116 host linear transcripts differentially expressed in priming phase of rat liver regeneration and clarified that circRNA abundance is associated with proliferation. Therefore, it suggests circRNA is highly expressed in liver and closely related to liver diseases.
CircRNA in HCC
Recently, compelling evidences already have proved that dysregulation of circRNA is associated with the development of HCC. CircRNAs is involved in proliferation, migration, invasion and apoptosis biological behaviors in HCC patients. According to the role of circRNAs, they can be divided into oncogenic agents and suppressors. Besides, circRNAs could be detected in tissue and body fluid, they are increasingly being used as potential diagnostic and prognostic markers for HCC. Here, we mainly focus on the regulation network mechanism of circRNA in HCC which illustrated in Fig. 3.

Roles and regulation network mechanism of HCC-related circRNAs. The diagram shows oncogenic agent and suppressor circRNAs in HCC and their influence on the biological processes, including proliferation, EMT, angiogenesis, invasion/migration, tumor growth, cell cycle and apoptosis. Oncogenic agent and suppressor circRNAs indicated in red and green arrow, respectively
CircRNA as oncogenic agent in HCC development
With development of circRNA microarray analysis and RNA-Seq detection, circRNA is highly expressed in biological samples including tissues, cell lines and serum of patients and acts as oncogenic agent in HCC. Previous studies have demonstrated that many miRNAs and target genes act as oncogenic agents in HCC, summarizing the role of circRNA in HCC will help us to better understand the pathogenesis of HCC and explore efficient and therapeutic targets. The following circRNAs can be used as oncogenic agents for HCC and are displayed in Table 1.
Most of circRNAs interact with miRNA as ceRNA and participate in the expression regulation of target genes by blocking the inhibitory effect of miRNA on their target mRNA, thus constructing circRNA-miRNA-mRNA regulation network in pathogenesis. Li et al. [68] reported that circPVT1 could up-regulate SIRT7 by binding with miR-3666, and SIRT7 could further adjust cell proliferation, cell cycle and transcription. Functional tests indicated that circPVT1 downregulation would reduce proliferation and increase cell apoptosis of HCC cell; similarly, hsa_circ_0056836 [69] might accelerate the progression of HCC via miR-766-3p/FOSL2 axis, in which FOSL2 was considered to be associated with photoperiodic regulation, fibrosis and even carcinoma; besides, circHIPK3, which is generated from the second exon of the HIPK3 gene, could promote cell proliferation and migration through AQP3, to transfer water and glycerol by the transmembrane channel, AQP3 played the important role in tumorigenesis and cancer progression by sponging of miR-124 [70, 71]; circ_0015756 [72] was proved highly expressed in HCC tissue, cell and even serum. Moreover, knockdown of circ_0015756 could inhibit tumor metastasis and invasion through miR-610/FGFR1; Yang et al. [73] reported that has_circ_0039053 could accelerate proliferation and invasion via the miR-637/USP21 axis in HCC cells; and Wu et al. [74] found circRASGRF2’s function as an oncogenic agent in HCC, and knockdown of circRASGRF2 suppressed the proliferation and migration of HCC cells by upregulating FAK expression through sponging miR-1224. The reported studies revealed that silence of circ_0091579 could restrain the progress of HCC via regulating miRNA, such as proliferation, migration and invasion in HCC cells [75].
CircRNA is also involved in the development of HCC through many signaling pathways exhibited in Fig. 4. Raf/MEK/ERK pathway plays an important role in cell growth, cell cycle and drug resistance, etc. It was verified that higher expression of circ_CDR1as could regulate MEK/ERK pathway via miR-1287/Raf1 axis, and then played positive roles in proliferation, migration, invasion, and EMT in HCC cells [76]. Similarly, hsa_circ_0003288 was overexpressed in HCC tissues and cells, which promoted the development of HCC by upregulating miR-145/PD-L1 axis via the PI3K/AKT signaling pathway [77]. It was found that circ_0067934 could activate signaling Wnt/β-catenin pathway by sponging of miR-1324 and improving FZD5 expression [78] and then promoting the proliferation, migration and invasion of HCC cells. CircZFR was proved to facilitate the progress of HCC through activating Wnt/β-catenin via regulating miR-3619-5p/CTNNB1 axis and miR-511/AKT1 signaling [79, 80]. It also reported that circZNF609 could accelerate the HCC development by activating the hedgehog pathway through regulation of miR-15a-5p/15b-5p and GLI2 expression. Knockdown of circZNF609 inhibited HCC cell proliferation, metastasis and stemness whereas boosted cell apoptosis [81].

The signaling pathways of circRNA in HCC. The drawing mainly shows that circRNA mediated ceRNA network plays crucial roles in HCC progression through signaling pathways
The change of cell cycle is also a major factor affecting the progression of HCC. For instance, as members of the cell cycle family, CK6, p6 and p27 could influence cell proliferation by regulating the G1/S transition. High expression of circBACH1 [82] and circ-ZEB1.33 [83] could promote the proliferation of HCC cells via HuR/p27 and miR-200a-3p/CDK6, respectively. Besides, circ-ZEB1.33 could also be determined in serum and associated with tumor-node-metastasis (TNM) stages, which is important in the development stage of HCC.
CircRNA might also influence the occurrence and development of HCC through regulating downstream protein directly. Li et al. [84] found that overexpression of hsa_circ_0085616 could promote the proliferation, migration, and invasion of HCC cells by increasing of β-catenin, p-ERK, and p-AKT protein levels.
Epithelial to mesenchymal transition (EMT) refers to a biological process of epithelial cells transforming from differentiated properties to mesenchymal characteristics [85], it is a key part of cancer metastasis [86]. Twist1 was found could regulate expression of vimentin by circCul2, and then promote the EMT, tumor metastasis and malignancy of HCC [87].
Emerging studies showed angiogenesis played an important role in rapid growing and metastasis of tumors [88]. Yang et al. [89] reported that circCRIM1 was upregulated and could promote HCC proliferation and angiogenesis via miR-378a-3p/SKP2 axis. Li et al. [90] found that circ_0001955 regulated HCC proliferation, metastasis, angiogenesis, and apoptosis through targeting the FZD4 via sponging miR-646. It also suggested that CircGFRA1 was to be associated with proliferative, migration and angiogenic activity of HCC by binding miR-149 [91].
Recently, it reported that m6A RNA methylation modulators contributed to the progression and prognosis of HCC. CircKIAA1429 was overexpression in HCC tissues and cell lines, and it could promote HCC migration, invasion, and EMT with a mechanism of m6A-YTHDF3-Zeb1, due to YTHDF3 could improve the stability of Zeb1mRNA [92]. CircMAP2K4 could promote HCC cell proliferation through hsa-miR-139-5p/YTHDF1 axis. Hsa-miR-139-5p and YTHDF1 expression levels were associated with grade, stage and prognosis of HCC [93].
CircRNA is also related to clinical manifestations of HCC patients. For example, circPVT1 [94] and hsa_circ_102559 [95] were closely associated with the overall survival (OS), lymph node metastasis (LNM), and TNM stages. Upregulation of circ-Gprc5a [96], circHOMER1 [97], circFAT1 [98] and cirrc-CSPP1 [99, 100] were positively correlated with the enlargement of tumor size and the higher TNM stage as well as poorer prognosis.
To sum up the above-mentioned, circRNA plays an oncogenic agent role and facilitates development of HCC through different molecular regulatory mechanisms, which will contribute to the discovery of therapeutic targets for HCC in the future.
CircRNA as suppressor in HCC
Compared with the high expression of circRNAs, some of them exhibit low expression which can inhibit the development of HCC and act as suppressors in HCC (Table 2).
CircRNA downregulation participates in proliferation, migration, invasion, and apoptosis by binding of miRNA, which has been proved was suppressor. For example, circMTO1 [101] acts as the sponge of oncogenic miR-9 to upregulate p21 expression and then suppresses progression of HCC. In another study, circADAMTS13 [102] was proved to inhibit HCC progression by sponging oncogenic miR-484. Moreover, downregulation of circSMAD2 [103], circRNA-5692 [104], hsa_circ_0005986 [105], circC3P1 [106] and circSETD3 [107] and circSETD3 were not only impeded the tumorigenesis of HCC but also correlated with clinicopathological characteristics. CircSMAD2 [103]was associated with differentiation of HCC tissues and circRNA-5692 [104] was correlated with AFP level, cirrhosis history, tumor size, and metastasis; hsa_circ_0005986 [105] was linked to chronic hepatitis B family history, tumor size, microvascular invasion (MVI) and BCLC stage; circC3P1 was negatively correlated with TNM stage, tumor size and MVI; circSETD was significantly associated with tumor size and differentiation of HCC tissues. All of the above-mentioned cases indicated the practical value to explore the therapeutic and prognostic targets of HCC.
Some circRNAs inhibit HCC development through some signaling pathways also exhibited in Fig. 4. For instance, hsa_circ_0079929 downregulation could inhibit the proliferation and cell cycle of HCC cells [108]. The functions of hsa_circ_0079929 were relevant to cell cycle regulatory protein CCNB1 and PI3K/AKT/mTOR signaling pathway. Wu et al. [109] found that circ_0004913 was partially inhibited the JAK2/STAT3/AKT signaling pathway through miR-184/HAMP axis, thus restraining the proliferation, metastasis, and glycolysis of HCC cells. Zhu et al. [110] reported that hsa_circ_0004018 suppressed proliferation and migration of HCC cells in-vivo and inhibited wnt/β-catenin signaling pathway through interacting with miR-626/DKK3. Besides, overexpression of these three circRNAs could curb the growth of HCC in-vivo.
Besides, a recent study illustrated that androgen receptor (AR) could inhibit the formation of vasculogenic mimicry of HCC and influence tumor metastasis by AR-circRNA7/miRNA7-5p/VE-Cadherin/Notch4 signaling [111]. Liang et al. also found [112] that circARSP91, as one of the circRNAs suppressed by AR via upregulating ADAR1, could inhibit HCC tumor growth in vitro and vivo, and laid the foundation for exploring new HCC therapies by reactivation of this circRNA.
CircRNA for potential diagnosis in HCC
Although cancer tissues can be detected with the size of less than 1 cm along with the development of imaging technologies such as magnetic resonance imaging and computerized tomography, it still represents a problem of a financial burden [113]. In a spite of the relatively low sensitivity and high rate of AFP (39.0–65.0%) for misdiagnosis of HCC, serumal AFP is still considered as the most commonly indicator for screening in the clinic [114]. Therefore, searching for new and reliable biomarkers is required and many types of circRNAs have been found as potential biomarkers in HCC (Table 3). The powerful multidimensional biomarkers can be efficiently used for prediction of patient response and disease diagnosis.
It was found that the area under the receiver operating characteristic curve (AUC) of circZKSCAN1 [115], has_circ_0005075 [116], hsa_circ_0016788 [52], hsa_circ_0003998 [53], circ-TCF4.85 [117] and circ-EPHB4 [118] reached above 0.800. Among them, has_circ_0005075 was related to tumor size of HCC; high expression of circ_0016788 [119] was correlated to high performance status score and large tumor size, increased Barcelona clinic liver cancer (BCLC) stage and abnormal AFP; the high expression of hsa_circ_0003998 [53] was associated with AFP, tumor size, differentiation and MVI and the lower expression of circ-EPHB4 [118] was positively associated with HBsAg and vascular tumor thrombus. All of them have great significance for diagnosing early HCC. There are also some circRNAs with lower AUC, whereas they are closely associated with the clinicopathological features of HCC. It was reported that hsa_circ_0068669 was associated with MVI and TNM stages and participated in metastasis of HCC, the AUC of curve of hsa_circ_0068699 was 0.64 with the values of sensitivity and specificity were 59.0% and 71.0% [120]. ZKSCAN1 mRNA and circZKSCAN1 are all the post-translational products from ZKSCAN1 gene. It was found that AUC of circZKSCAN1 was 0.834 and exhibited high sensitivity (82.2%) and specificity (72.4%), respectively, which was much higher than that of mRNA of ZKSCAN331 (AUC = 0.474) [115]. Therefore, it suggested that circZKSCAN1 demonstrated better efficiency in the diagnosis of HCC tissues than ZKSCAN1 mRNA.
Many circRNAs have been revealed unique features with the development of HCC, such as: the expression features of hsa_circ_0003570 [121], hsa_circ_0028502 and hsa_circ_0076251 [122] exhibiting stage-specific in HCC tissues, liver cirrhosis (LC) and chronic hepatitis (CH); and the expression levels are gradually increased. Moreover, hsa_circ_0003570 was closely related to tumor size, differentiation, MVI, BCLC stages and TMN stages; hsa_circ_0028502 were related to TNM stages, and the expression levels of hsa_circ_0076251 were related to BCLC stages and the presence of serum HbsAg, respectively, which suggested that these circRNAs also could be used as biomarkers at different stages in the development of HCC.
Serumal circRNA can also be employed as a non-invasive biomarker for the diagnosis of HCC. It showed that the ROC curve analysis for hsa_cirR_00156, hsa_cirR_000224 and hsa_cirR_000520 [123] were employed for discrimination between HCC patients and the healthy group, the AUC were 0.839, 0.974 and 0.943, respectively, which were higher than that of AFP (AUC = 0.726). Moreover, combining these three circRNAs displayed a sensitivity of 100% and a specificity of 83.3%. In another study, the level of circ_104075 [124] in serum was positively related to the stage of HCC. In addition, circ_104075 might be a potential diagnostic biomarker with an AUC of 0.973, the sensitivity and specificity values were 96.0% and 98.3%, respectively, which was also higher than that of AFP for predication of HCC. Moreover, the m6A modification could enhance the interaction between miR-582-3p and YAP3′UTR, and contributed to circRNA_104075 promoted HCC progression by miR-582-3p/YAP. It also displayed that serumal circ-FOXP1 plays an important role in the diagnosis and metastasis of HCC with the AUC of 0.932 and was associated with the tumor size, MVI and TNM stage [125].
Zhang et al. [126] found that the expression level of plasma hsa_circ_0001445 in HCC patients, cirrhosis patients and hepatitis B patients were lower than those in healthy group. Moreover, ROC curve was used to distinguish HCC patients from the healthy control (AUC = 0.862), cirrhosis (AUC = 0.672) and the hepatitis B patients (AUC = 0.764) according to the plasma level of hsa_circ_0001445. Besides, the combination of hsa_circ_0001445 and AFP revealed better diagnostic value than any of them. Another study [127] demonstrated that circ-CDYL combining with HDGF and HIF1AN were proved as promising biomarkers for diagnostic of early HCC than that of AFP with the odds ratios (ORs) of 124.58 (95% CI 13.260–1170.560).
CircRNA as potential prognostic biomarker in HCC
HCC is characterized as high degree of malignancy, poor prognosis, low 5-year survival rate and high recurrence rate. At present, AFP and routine liver function tests are often used as monitoring methods to predict the recurrence of HCC. In addition, α‐fetoprotein‐L3 and Golgi protein73 play important role in prediction of HCC [128]. However, they performed lower sensitivity and limited efficiency. It is showed that circRNA is closely associated with the recurrence-free survival (RFS) and OS of HCC by univariate and multivariate Cox regression analysis, dysregulated circRNA might be used as a prognostic factor for the selection of rational treatment regimens for HCC, due to circRNA shows stable conserved and high specificity in different tissues even the stages of disease [129]. For instance, hsa_circ_0128298 [130], circLARP4 [131], circTP63 [132], circBIRC6 [133], hsa_circ_104348 [134] and circZNF566 [135], etc., were overexpressed in HCC tissues and cell lines. Kaplan–Meier survival analysis showed that HCC patients with circRNA high expression and had shorter overall survival than those with low expression. In addition, hsa_circ_0128298 [130], circLARP4 [131] and circBIRC6 [133] were testified to be the independent prognostic factors of OS by multivariate Cox regression analysis. Moreover, studies have found that high expression of hsa_circ_0128298 was associated with vascular cancer embolus, LNM and organ metastasis; overexpression of circTP63 [132] was related to TNM stage, tumor differentiation, and LNM; circLARP4 [131] and circZNF566 [135] were associated with clinicopathological features, including tumor differentiation, tumor size and TNM stage, etc., circ-BIRC6 [133] was associated with TNM stage and MVI in HCC tissues; hsa_circ_104348 [134] expression level was related to tumor size, LNM as well as TNM. Besides, univariate and multivariate Cox regression analysis showed circZKSCAN1 was an independent prognostic factor of OS and RFS by binding FMRP, and the combination of circZKSCAN1 and CCAR1 could improve the prognostic value of HCC [136].
Moreover, cSMARCA5 [137], hsa_circ_0001649 [138] and circTRIM33-12 [139] were downregulated in HCC tissues. It reported that cSMARCA5 was associated with tumor differentiation, tumor stage, tumor size and MVI; and circTRIM33-12 was linked to multiple tumors, tumor size, encapsulation invasion, AFP levels and MVI. Furtherly, hsa_circ_0001649 was related to OS, the lower expressions of cSMARCA5 and circTRIM33-12 in HCC patients exhibited poor OS and RFS by Kaplan–Meier analysis; and multivariate analyses indicated that both them were the independent risk factors of OS and RFS. Therefore, they revealed the critical guiding significance to the prognosis of HCC. Recently, circRNAs act as potential prognosis biomarkers were listed in Table 4.
CircRNA of HBV related HCC
HBV infection is a major risk factor in the high incidence of HCC in the areas of Asia and sub‑Saharan Africa [140]. It reported that high expression of circRNA_100338 in HCC tissues compared with paired pericancerous live tissues samples with hepatitis B, the cumulative survival rate (72.0%) of HCC patients in the circRNA_100338-high group was significantly lower than that of the low expression group (42.9%); circRNA_100338 was highly correlated with TNM stage, invasion and metastasis in HCC via antagonizing miR-141-3p [141]. Hsa_circ_0027089, a plasmatic circRNA, was proved to discriminate HBV-related HCC from HBV-related cirrhosis and healthy participants with AUC values of 0.765 and 0.794 [142]. It might act as a potential biomarker for clinical diagnosis and evaluation in HCC with HBV. Another study identified 24 and 23 circRNAs with up-regulation and down-regulation by microarray in three paired HBV‑related HCC tissues and adjacent non‑tumorous tissues; and the differentially expressed circRNA/miRNA interactions were predicted by miRNA target prediction software [143]. Here it can be concluded that circRNA plays a critical role in distinguishing different types of HCC, e.g., plasmatic circRNA could act as a valuable diagnosis biomarker to distinguish HBV-related HCC and HBV-related cirrhosis.
CircRNA in other liver diseases
Chronic hepatitis B (CHB)
Zhou et al. [144] provided the first evidence for differentially expressed circRNAs in CHB. Interestingly, a bioinformatics method for identification of CHB-associated circRNA and the silico analysis were established for four predicted circRNA-miRNA-mRNA pathways in progression of HBV-associated liver disease. They found 72 up-regulated and 95 down-regulated circRNAs were changed more than twice in CHB tissues compared with the normal tissues. Subsequently, four pathways of circRNA-miRNA-mRNA were also discovered, including: hsa_circ_0005389-miR-4505/miR-6752-5p/miR-5787-IRF7, hsa_circ_0000650-miR-6873-3p-TGFβ2, hsa_circ_0000650-miR-210-5p-HBV and hsa_circ_0000038-miR-370/miR-939-HBV. Besides, it was found that circ_0004812 was upregulation in CHB and promoted FSTL1 expression by binding to miR-1287-5p [145]. Additionally, the enhanced expression mRNA and its related protein of IFN-α and IFN-β in the circ_0004812 knockdown cells, suggests that circ_0004812 could regulate HBV-induced immune suppression by miR-1287-5p/FSTL1 axis. They demonstrated a new perspective on the new pathogenesis of CHB through circRNA.
Hepatic steatosis
Hepatic steatosis is recognized as one of the common chronic liver diseases in western countries and the Asia–Pacific area [146,147,148]. A large proportion of patients with non-alcoholic fatty liver disease (NAFLD) may further develop non-alcoholic steatohepatitis (NASH), fibrosis, cirrhosis, or even HCC [149]. Some circRNAs derived from mitochondria were down-regulated in NASH [150]. Among them, hsa_circ_0089762 was associated with mitochondrial reactive oxygen species (mROS) release and metaflammation in NASH. In terms of the mechanism, hsa_circ_0089762 directly interacts with ATP5B to block the binding of ATP5B to CypD and inhibit the opening of mitochondrial permeability transition pore, thus inhibiting mROS output and fibroblast activation. Therefore, mitochondrial circRNA could be a new therapeutic target for NASH. circRNA_002581 overexpression was found to up-regulate CPEB1 by sponging miR-122 in the NASH mice model [151]. Further study indicated that circRNA_002581-miR-122-CPEB1 was linked to autophagy by PTEN-AMPK-mTOR pathway. Besides, circRNA_0046367 and circRNA_0046366 could act as the antagonists of miR-34a are involved in NAFLD [152, 153]. Therefore, these pathways provide novel approaches for NASH pathogenesis, diagnosis and treatment.
Hepatoblastoma
Hepatoblastoma is the most common malignant liver cancer in infants and toddlers [154]. Circ_0015756 was found significantly up-regulated in human hepatoblastoma tissues and cell lines and circ_0015756 siRNA decreased the viability, proliferation and invasion ability of hepatoblastoma cells in vitro by sponging miR-1250-3p [155]. Li et al. [156] demonstrated that circSETD3 downregulated in Hepatoblastoma tissues and cell lines, and it exerted as a tumor suppressor to inhibit proliferation, migration, EMT process, and induce apoptosis by miR-423-3p/Bim axis.
Conclusions
By these tokens, a large number studies have shown that circRNAs are abnormally expressed in liver diseases and play a vital role in regulating different molecules, signaling pathways, pathophysiological activities, etc. However, the study on the relationship between circRNA and liver disease is still in the stage of infancy. Most of the molecular mechanisms of circRNA in liver diseases are remained unclear, although the development and clarifying researches of circRNA in liver diseases are accelerating. Besides, there are still many deficiencies in the study of circRNA. Firstly, compelling evidences are mainly focused on HCC, while a few are concentrated on other liver diseases. Secondly, the sample sizes in many studies of circRNA as diagnostic biomarker are relatively small, and methodologies are required to be standardized to eliminate possible artifacts. Last but not least, most circRNAs are derived from the pathological tissues of the liver, the role and function of circRNA in serum, exosomes and microvesicles are still seldom reported. We believed that the application of circRNA has a broad prospect in the drug therapeutic target, diagnosis and treatment of liver diseases with the improvement of detection, validation and functional analysis methods of circRNA.
Availability of data and materials
Data will be provided based on reasonable request.
Abbreviations
- AFP:
-
Alpha fetoprotein
- AR:
-
Androgen receptor
- AUC:
-
Area under the receiver operating characteristic curve
- BCLC:
-
Barcelona clinic liver cancer
- CH:
-
Chronic hepatitis
- ciRNA:
-
Circular intronic RNA
- circRNA:
-
Circular RNA
- dd PCR:
-
Droplet digital PCR
- DECs:
-
Differential expression of circRNAs
- EIciRNA:
-
Exon–intron circRNA
- EcircRNA:
-
Exonic circRNAs
- HCC:
-
Hepatocellular carcinoma
- HuR:
-
Human antigen R
- LC:
-
Liver cirrhosis
- LNM:
-
Lymph node metastasis
- m6A:
-
N6-methyladenosine
- mROS:
-
Mitochondrial reactive oxygen species
- MVI:
-
Microvascular invasion
- NASH:
-
Non-alcoholic steatohepatitis
- ncRNA:
-
Non-coding RNA
- NGS:
-
Next generation sequencing
- ORs:
-
Odds ratios
- OS:
-
Overall survival
- RFS:
-
Recurrence-free survival
- RNA-seq:
-
RNA sequencing
- TNM:
-
Tumor-node-metastasis
References
Danan M, Schwartz S, Edelheit S, Sorek R. Transcriptome-wide discovery of circular RNAs in Archaea. Nucleic Acids Res. 2012;40(7):3131–42.
Jeck WR, Sorrentino JA, Wang K, Slevin MK, Burd CE, Liu J, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. 2013;19(2):141–57.
Cocquerelle C, Mascrez B, Hétuin D, Bailleul B. Mis-splicing yields circular RNA molecules. FASEB J. 1993;7(1):155–60.
Patop IL, Wüst S, Kadener S. Past, present, and future of circRNAs. Embo J. 2019;38(16):e100836.
Patop IL, Kadener S. circRNAs in Cancer. Curr Opin Genet Dev. 2018;48:121–7.
Memczak S, Papavasileiou P, Peters O, Rajewsky N. Identification and characterization of circular RNAs as a new class of putative biomarkers in human blood. PLoS ONE. 2015;10(10):e0141214.
Chen LL. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. 2020;21(8):475–90.
Li H, Li K, Lai W, Li X, Wang H, Yang J, et al. Comprehensive circular RNA profiles in plasma reveals that circular RNAs can be used as novel biomarkers for systemic lupus erythematosus. Clin Chimica Acta. 2018;480:17–25.
Li GF, Li L, Yao ZQ, Zhuang SJ. Hsa_circ_0007534/miR-761/ZIC5 regulatory loop modulates the proliferation and migration of glioma cells. Biochem Biophys Res Commun. 2018. https://doi.org/10.1016/j.bbrc.2018.03.219.
Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20(11):675–91.
Sakshi S, Jayasuriya R, Ganesan K, Xu B, Ramkumar KM. Role of circRNA-miRNA-mRNA interaction network in diabetes and its associated complications. Mol Ther Nucleic Acids. 2021;26:1291–302.
Tang X, Ren H, Guo M, Qian J, Yang Y, Gu C. Review on circular RNAs and new insights into their roles in cancer. Comput Struct Biotechnol J. 2021;19:910–28.
Chien Y, Tsai PH, Lai YH, Lu KH, Liu CY, Lin HF, et al. CircularRNA as novel biomarkers in liver diseases. J Chin Med Assoc. 2020;83(1):15–7.
Asrani SK, Devarbhavi H, Eaton J, Kamath PS. Burden of liver diseases in the world. J Hepatol. 2018. https://doi.org/10.1016/j.jhep.2018.09.014
Mokdad AA, Lopez AD, Shahraz S, Lozano R, Mokdad AH, Stanaway J, et al. Liver cirrhosis mortality in 187 countries between 1980 and 2010: a systematic analysis. BMC Med. 2014;12:24.
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021. https://doi.org/10.3322/caac.21660.
Akinyemiju T, Abera S, Ahmed M, Alam N, Alemayohu MA, Allen C, et al. The burden of primary liver cancer and underlying etiologies from 1990 to 2015 at the global, regional, and national level: results from the global burden of disease study 2015. JAMA Oncol. 2017;3(12):1683–91.
Bruix J, Sherman M. Management of hepatocellular carcinoma: an update. Hepatology. 2011;53(3):1020–2.
Galle PR. Treating hepatobiliary cancers: the oncology way. Dig Dis. 2017;35(4):384–6.
Juárez-Hernández E, Motola-Kuba D, Chávez-Tapia NC, Uribe M, Barbero BV. Biomarkers in hepatocellular carcinoma: an overview. Expert Rev Gastroenterol Hepatol. 2017;11(6):549–58.
Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136(5):E359–86.
Sherman M. Recurrence of hepatocellular carcinoma. N Engl J Med. 2008;359(19):2045–7.
Shen H, Liu B, Xu J, Zhang B, Wang Y, Shi L, et al. Circular RNAs: characteristics, biogenesis, mechanisms and functions in liver cancer. J Hematol Oncol. 2021;14(1):134.
Suzuki H, Tsukahara T. a view of pre-mRNA splicing from RNase R resistant RNAs. Int J Mol Sci. 2014;15(6):9331.
Zhang XO, Wang HB, Zhang Y, Lu X, Chen LL, Yang L. Complementary sequence-mediated exon circularization. Cell. 2014;159(1):134–47.
Conn SJ, Pillman KA, Toubia J, Conn VM, Salmanidis M, Phillips CA, et al. The RNA binding protein quaking regulates formation of circRNAs. Cell. 2015;160(6):1125–34.
Tang L, Li P, Jang M, Zhu W. Circular RNAs and cardiovascular regeneration. Front Cardiovasc Med. 2021;8:672600.
Guo JU, Agarwal V, Guo H, Bartel DP. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014;15(7):409.
Shen Y, Guo X, Wang W. Identification and characterization of circular RNAs in zebrafish. FEBS Lett. 2017;591(1):213–20.
Lai X, Bazin J, Webb S, Crespi M, Zubieta C, Conn SJ. CircRNAs in Plants. Adv Exp Med Biol. 2018;1087:329–43.
Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–8.
Wang Y, Mo Y, Gong Z, Yang X, Yang M, Zhang S, et al. Circular RNAs in human cancer. Mol Cancer. 2017;16(1):25.
Westholm JO, Miura P, Olson S, Shenker S, Joseph B, Sanfilippo P, et al. Genome-wide analysis of drosophila circular RNAs reveals their structural and sequence properties and age-dependent neural accumulation. Cell Rep. 2014;9(5):1966–80.
Xia S, Feng J, Lei L, Hu J, Xia L, Wang J, et al. Comprehensive characterization of tissue-specific circular RNAs in the human and mouse genomes. Brief Bioinform. 2017;18(6):984–92.
Jeck WR, Sorrentino JA, Wang K, Slevin MK, Burd CE, Liu J, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. 2013;19(2):141–57.
Conn VM, Hugouvieux V, Nayak A, Conos SA, Capovilla G, Cildir G, et al. A circRNA from SEPALLATA3 regulates splicing of its cognate mRNA through R-loop formation. Nat Plants. 2017;3(5):17053.
Li Z, Huang C, Bao C, Chen L, Lin M, Wang X, et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol. 2015;22(3):256–64.
Kulcheski FR, Christoff AP, Margis R. Circular RNAs are miRNA sponges and can be used as a new class of biomarker. J Biotechnol. 2016;238:42–51.
Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495(7441):384–8.
Huang A, Zheng H, Wu Z, Chen M, Huang Y. Circular RNA-protein interactions: functions, mechanisms, and identification. Theranostics. 2020;10(8):3503–17.
Du WW, Yang W, Chen Y, Wu ZK, Foster FS, Yang Z, et al. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur Heart J. 2016;38(18):1402.
Yang F, Hu A, Li D, Wang J, Guo Y, Liu Y, et al. Circ-HuR suppresses HuR expression and gastric cancer progression by inhibiting CNBP transactivation. Mol Cancer. 2019;18(1):158.
Legnini I, Timoteo GD, Rossi F, Morlando M, Briganti F, Sthandier O, et al. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol Cell. 2017;66(1):22–37.
Zhang L, Hou C, Chen C, Guo Y, Yuan W, Yin D, et al. The role of N(6)-methyladenosine (m(6)A) modification in the regulation of circRNAs. Mol Cancer. 2020;19(1):105.
Szabo L, Salzman J. Detecting circular RNAs: bioinformatic and experimental challenges. Nat Rev Genet. 2016;17(11):679–92.
Ely A, Bloom K, Maepa MB, Arbuthnot P. Recent update on the role of circular RNAs in hepatocellular carcinoma. J Hepatocell Carcinoma. 2021;8:1–17.
Li S, Teng S, Xu J, Su G, Zhang Y, Zhao J, et al. Microarray is an efficient tool for circRNA profiling. Brief Bioinform. 2019;20(4):1420–33.
Chen L, Wang C, Sun H, Wang J, Liang Y, Wang Y, et al. The bioinformatics toolbox for circRNA discovery and analysis. Brief Bioinform. 2021;22(2):1706–28.
Guan Z, Tan J, Gao W, Li X, Yang Y, Li X, Li Y, Wang Q. Circular RNA hsa_circ_0016788 regulates hepatocellular carcinoma tumorigenesis through miR-486/CDK4 pathway. J Cell Physiol. 2018. https://doi.org/10.1002/jcp.26612.
Yu X, Sheng P, Sun J, Zhao X, Zhang J, Li Y, et al. The circular RNA circMAST1 promotes hepatocellular carcinoma cell proliferation and migration by sponging miR-1299 and regulating CTNND1 expression. Cell Death Dis. 2020;11(5):340.
Hu ZQ, Zhou SL, Li J, Zhou ZJ, Wang PC, Xin HY, et al. Circular RNA sequencing identifies CircASAP1 as a key regulator in hepatocellular carcinoma metastasis. Hepatology. 2019. https://doi.org/10.1002/hep.31068.
Guan Z, Tan J, Gao W, Li X, Yang Y, Li X, et al. Circular RNA hsa_circ_0016788 regulates hepatocellular carcinoma tumorigenesis through miR-486/CDK4 pathway. J Cell Physiol. 2018;234(1):500–8.
Qiao GL, Chen L, Jiang WH, Yang C, Yang CM, Song LN, et al. Hsa_circ_0003998 may be used as a new biomarker for the diagnosis and prognosis of hepatocellular carcinoma. Onco Targets Ther. 2019;12:5849–60.
Lei M, Zheng G, Ning Q, Zheng J, Dong D. Translation and functional roles of circular RNAs in human cancer. Mol Cancer. 2020;19(1):30.
Jeck WR, Sharpless NE. Detecting and characterizing circular RNAs. Nat Biotechnol. 2014;32(5):453–61.
Hindson CM, Chevillet JR, Briggs HA, Gallichotte EN, Ruf IK, Hindson BJ, et al. Absolute quantification by droplet digital PCR versus analog real-time PCR. Nat Methods. 2013;10(10):1003–5.
Schneider T, Schreiner S, Preußer C, Bindereif A, Rossbach O. Northern blot analysis of circular RNAs. Methods Mol Biol. 2018;1724:119–33.
Zirkel A, Papantonis A. Detecting circular RNAs by RNA fluorescence in situ hybridization. Methods Mol Biol. 2018;1724:69–75.
Glažar P, Papavasileiou P, Rajewsky N. circBase: a database for circular RNAs. RNA. 2014;20(11):1666–70.
Zhang XO, Dong R, Zhang Y, Zhang JL, Luo Z, Zhang J, et al. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016;26(9):1277–87.
Dudekula DB, Panda AC, Grammatikakis I, De S, Abdelmohsen K, Gorospe M. CircInteractome: a web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016;13(1):34–42.
Ghosal S, Das S, Sen R, Basak P, Chakrabarti J. Circ2Traits: a comprehensive database for circular RNA potentially associated with disease and traits. Front Genet. 2013;4:283.
Michalopoulos GK, Bhushan B. Liver regeneration: biological and pathological mechanisms and implications. Nat Rev Gastroenterol Hepatol. 2021;18(1):40–55.
Trefts E, Gannon M, Wasserman DH. The liver. Curr Biol. 2017;27(21):R1147-r1151.
Zeng X, Yuan X, Cai Q, Tang C, Gao J. Circular RNA as an epigenetic regulator in chronic liver diseases. Cells. 2021. https://doi.org/10.3390/cells10081945.
Song M, Xia L, Sun M, Yang C, Wang F. Circular RNA in liver: health and diseases. Adv Exp Med Biol. 2018;1087:245–57.
Li L, Guo J, Chen Y, Chang C, Xu C. Comprehensive circRNA expression profile and selection of key circRNAs during priming phase of rat liver regeneration. BMC Genom. 2017;18(1):80.
Li Y, Shi H, Yuan J, Qiao L, Dong L, Wang Y. Downregulation of circular RNA circPVT1 restricts cell growth of hepatocellular carcinoma through downregulation of Sirtuin 7 via microRNA-3666. Clin Exp Pharmacol Physiol. 2020;47(7):1291–300.
Li Z, Liu Y, Yan J, Zeng Q, Hu Y, Wang H, et al. Circular RNA hsa_circ_0056836 functions an oncogenic gene in hepatocellular carcinoma through modulating miR-766-3p/FOSL2 axis. Aging. 2020;12(3):2485–97.
Chen G, Shi Y, Liu M, Sun J. circHIPK3 regulates cell proliferation and migration by sponging miR-124 and regulating AQP3 expression in hepatocellular carcinoma. Cell Death Dis. 2018;9(2):175.
Jablonski EM, Mattocks MA, Sokolov E, Koniaris LG, Hughes FM Jr, Fausto N, et al. Decreased aquaporin expression leads to increased resistance to apoptosis in hepatocellular carcinoma. Cancer Lett. 2007;250(1):36–46.
Liu L, Yang X, Li NF, Lin L, Luo H. Circ_0015756 promotes proliferation, invasion and migration by microRNA-7-dependent inhibition of FAK in hepatocellular carcinoma. Cell Cycle. 2019;18(21):2939–53.
Yang TB, Yi F, Liu WF, Yang YH, Yang C, Sun J. Identification of hsa_circ_0039053 as an up-regulated and oncogenic circRNA in hepatocellular carcinoma via the miR-637-mediated USP21 activation. J Cancer. 2020;11(23):6950–9.
Wu D, Xia A, Fan T, Li G. circRASGRF2 functions as an oncogenic gene in hepatocellular carcinoma by acting as a miR-1224 sponge. Mol Ther Nucleic Acids. 2021;23:13–26.
Niu WY, Chen L, Zhang P, Zang H, Zhu B, Shao WB. Circ_0091579 promotes proliferative ability and metastasis of liver cancer cells by regulating microRNA-490-3p. Eur Rev Med Pharmacol Sci. 2019;23(23):10264–73.
Zhang B, Li F, Zhu Z, Ding A, Luo J. CircRNA CDR1as/miR-1287/Raf1 axis modulates hepatocellular carcinoma progression through MEK/ERK pathway. Cancer Manag Res. 2020;12:8951–64.
Xu G, Zhang P, Liang H, Xu Y, Shen J, Wang W, et al. Circular RNA hsa_circ_0003288 induces EMT and invasion by regulating hsa_circ_0003288/miR-145/PD-L1 axis in hepatocellular carcinoma. Cancer Cell Int. 2021;21(1):212.
Zhu Q, Lu G, Luo Z, Gui F, Wu J, Zhang D, et al. CircRNA circ_0067934 promotes tumor growth and metastasis in hepatocellular carcinoma through regulation of miR-1324/FZD5/Wnt/β-catenin axis. Biochem Biophys Res Commun. 2018;497(2):626–32.
Tan A, Li Q, Chen L. CircZFR promotes hepatocellular carcinoma progression through regulating miR-3619-5p/CTNNB1 axis and activating Wnt/β-catenin pathway. Arch Biochem Biophys. 2019;661:196–202.
Yang X, Liu L, Zou H, Zheng YW, Wang KP. circZFR promotes cell proliferation and migration by regulating miR-511/AKT1 axis in hepatocellular carcinoma. Dig Liver Dis. 2019;51(10):1446–55.
He Y, Huang H, Jin L, Zhang F, Zeng M, Wei L, et al. CircZNF609 enhances hepatocellular carcinoma cell proliferation, metastasis, and stemness by activating the Hedgehog pathway through the regulation of miR-15a-5p/15b-5p and GLI2 expressions. Cell Death Dis. 2020;11(5):358.
Liu B, Yang G, Wang X, Liu J, Lu Z, Wang Q, et al. CircBACH1 (hsa_circ_0061395) promotes hepatocellular carcinoma growth by regulating p27 repression via HuR. J Cell Physiol. 2020;235(10):6929–41.
Gong Y, Mao J, Wu D, Wang X, Li L, Zhu L, et al. Circ-ZEB1.33 promotes the proliferation of human HCC by sponging miR-200a-3p and upregulating CDK6. Cancer Cell Int. 2018;18:116.
Li W, Zhou X, Wu X, Wei J, Huang Z. The role of circular RNA hsa_circ_0085616 in proliferation and migration of hepatocellular carcinoma cells. Cancer Manag Res. 2019;11:7369–76.
Yang J, Antin P, Berx G, Blanpain C, Brabletz T, Bronner M, et al. Guidelines and definitions for research on epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2020;21(6):341–52.
Dongre A, Weinberg RA. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat Rev Mol Cell Biol. 2019;20(2):69–84.
Meng J, Chen S, Han JX, Qian B, Wang XR, Zhong WL, et al. Twist1 regulates vimentin through Cul2 circular RNA to promote EMT in hepatocellular carcinoma. Cancer Res. 2018;78(15):4150–62.
Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285(21):1182–6.
Ji Y, Yang S, Yan X, Zhu L, Yang W, Yang X, et al. CircCRIM1 promotes hepatocellular carcinoma proliferation and angiogenesis by sponging miR-378a-3p and regulating SKP2 expression. Front Cell Dev Biol. 2021;9:796686.
Li X, Lv J, Hou L, Guo X. Circ_0001955 acts as a miR-646 sponge to promote the proliferation, metastasis and angiogenesis of hepatocellular carcinoma. Dig Dis Sci. 2021. https://doi.org/10.1007/s10620-021-07053-8.
Yu YX, Ge TW, Zhang P. Circular RNA circGFRA1 promotes angiogenesis, cell proliferation and migration of hepatocellular carcinoma by combining with miR-149. Eur Rev Med Pharmacol Sci. 2020;24(21):11058–64.
Wang M, Yang Y, Yang J, Yang J, Han S. circ_KIAA1429 accelerates hepatocellular carcinoma advancement through the mechanism of m(6)A-YTHDF3-Zeb1. Life Sci. 2020;257:118082.
Chi F, Cao Y, Chen Y. Analysis and validation of circRNA-miRNA network in regulating m(6)A RNA methylation modulators reveals CircMAP2K4/miR-139–5p/YTHDF1 axis involving the proliferation of hepatocellular carcinoma. Front Oncol. 2021;11:560506.
Zhu Y, Liu Y, Xiao B, Cai H, Liu M, Ma L, et al. The circular RNA PVT1/miR-203/HOXD3 pathway promotes the progression of human hepatocellular carcinoma. Biol Open. 2019. https://doi.org/10.1242/bio.043687.
Li J, Yu Z, Zhu Q, Tao C, Xu Q. hsa_circ_102559 acts as the sponge of miR-130a-5p to promote hepatocellular carcinoma progression through regulation of ANXA2. Cell Transplant. 2020;29:963689720968748.
Lin Y, Huang G, Jin H, Jian Z. Circular RNA Gprc5a promotes HCC progression by activating YAP1/TEAD1 signalling pathway by sponging miR-1283. Onco Targets Ther. 2020;13:4509–21.
Zhao M, Dong G, Meng Q, Lin S, Li X. Circ-HOMER1 enhances the inhibition of miR-1322 on CXCL6 to regulate the growth and aggressiveness of hepatocellular carcinoma cells. J Cell Biochem. 2020;121(11):4440–9.
Wei H, Yan S, Hui Y, Liu Y, Guo H, Li Q, et al. CircFAT1 promotes hepatocellular carcinoma progression via miR-30a-5p/REEP3 pathway. J Cell Mol Med. 2020. https://doi.org/10.1111/jcmm.16085.
Sun Q, Yu R, Wang C, Yao J, Zhang L. Circular RNA circ-CSPP1 regulates CCNE2 to facilitate hepatocellular carcinoma cell growth via sponging miR-577. Cancer Cell Int. 2020;20:202.
Jia N, Song Z, Chen B, Cheng J, Zhou W. A novel circular RNA circCSPP1 promotes liver cancer progression by sponging miR-1182. Onco Targets Ther. 2021;14:2829–38.
Han D, Li J, Wang H, Su X, Hou J, Gu Y, et al. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology. 2017;66(4):1151–64.
Qiu L, Huang Y, Li Z, Dong X, Chen G, Xu H, et al. Circular RNA profiling identifies circADAMTS13 as a miR-484 sponge which suppresses cell proliferation in hepatocellular carcinoma. Mol Oncol. 2019;13(2):441–55.
Zhang X, Luo P, Jing W, Zhou H, Liang C, Tu J. circSMAD2 inhibits the epithelial-mesenchymal transition by targeting miR-629 in hepatocellular carcinoma. Onco Targets Ther. 2018;11:2853–63.
Liu Z, Yu Y, Huang Z, Kong Y, Hu X, Xiao W, et al. CircRNA-5692 inhibits the progression of hepatocellular carcinoma by sponging miR-328-5p to enhance DAB2IP expression. Cell Death Dis. 2019;10(12):900.
Fu L, Chen Q, Yao T, Li T, Sheng Y, Hu Y, et al. Hsa_circ_0005986 inhibits carcinogenesis by acting as a miR-129-5p sponge and is used as a novel biomarker for hepatocellular carcinoma. Oncotarget. 2017;8(27):43878–88.
Zhong L, Wang Y, Cheng Y, Wang W, Lu B, Zhu L, et al. Circular RNA circC3P1 suppresses hepatocellular carcinoma growth and metastasis through miR-4641/PCK1 pathway. Biochem Biophys Res Commun. 2018;499(4):1044–9.
Xu L, Feng X, Hao X, Wang P, Zhang Y, Zheng X, et al. CircSETD3 (Hsa_circ_0000567) acts as a sponge for microRNA-421 inhibiting hepatocellular carcinoma growth. J Exp Clin Cancer Res. 2019;38(1):98.
Zheng H, Chen T, Li C, Xu C, Ding C, Chen J, et al. A circular RNA hsa_circ_0079929 inhibits tumor growth in hepatocellular carcinoma. Cancer Manag Res. 2019;11:443–54.
Wu M, Sun T, Xing L. Circ_0004913 inhibits cell growth, metastasis, and glycolysis by absorbing miR-184 to regulate HAMP in hepatocellular carcinoma. Cancer Biother Radiopharm. 2020. https://doi.org/10.1089/cbr.2020.3779.
Zhu P, Liang H, Huang X, Zeng Q, Liu Y, Lv J, et al. Circular RNA Hsa_circ_0004018 inhibits Wnt/β-catenin signaling pathway by targeting microRNA-626/DKK3 in hepatocellular carcinoma. Onco Targets Ther. 2020;13:9351–64.
Bao S, Jin S, Wang C, Tu P, Hu K, Lu J. Androgen receptor suppresses vasculogenic mimicry in hepatocellular carcinoma via circRNA7/miRNA7–5p/VE-cadherin/Notch4 signalling. J Cell Mol Med. 2020. https://doi.org/10.1111/jcmm.16022.
Liang S, Yan P, Liang Y, Yin S, Shen J, Zhou S, et al. Circular RNA expression is suppressed by androgen receptor (AR)-regulated adenosine deaminase that acts on RNA (ADAR1) in human hepatocellular carcinoma. Cell Death Dis. 2017;8(11):e3171.
Bruix J, Reig M, Sherman M. Evidence-based diagnosis, staging, and treatment of patients with hepatocellular carcinoma. Gastroenterology. 2016;150(4):835–53.
Tian MM, Fan YC, Zhao J, Gao S, Zhao ZH, Chen LY, et al. Hepatocellular carcinoma suppressor 1 promoter hypermethylation in serum. A diagnostic and prognostic study in hepatitis B. Clin Res Hepatol Gastroenterol. 2017;41(2):171–80.
Yao Z, Luo J, Hu K, Lin J, Huang H, Wang Q, et al. ZKSCAN1 gene and its related circular RNA (circZKSCAN1) both inhibit hepatocellular carcinoma cell growth, migration, and invasion but through different signaling pathways. Mol Oncol. 2017;11(4):422–37.
Shang X, Li G, Liu H, Li T, Liu J, Zhao Q, et al. Comprehensive circular RNA profiling reveals that hsa_circ_0005075, a new circular rna biomarker, is involved in hepatocellular crcinoma development. Medicine. 2016;95(22):e3811.
Gao J, Dai C, Yu X, Yin XB, Zhou F. Circ-TCF4.85 silencing inhibits cancer progression through microRNA-486–5p-targeted inhibition of ABCF2 in hepatocellular carcinoma. Mol Oncol. 2020;14(2):447–61.
Tan Y, Du B, Zhan Y, Wang K, Wang X, Chen B, et al. Antitumor effects of circ-EPHB4 in hepatocellular carcinoma via inhibition of HIF-1α. Mol Carcinog. 2019;58(6):875–86.
Cheng F, Wang L, Zhang J. Circular RNA 0016788 displays as a biomarker for tumor progression and poor prognosis in surgical hepatocellular carcinoma patients. J Clin Lab Anal. 2020;34(7):e23300.
Yao T, Chen Q, Shao Z, Song Z, Fu L, Xiao B. Circular RNA 0068669 as a new biomarker for hepatocellular carcinoma metastasis. J Clin Lab Anal. 2018;32(8):e22572.
Fu L, Wu S, Yao T, Chen Q, Xie Y, Ying S, et al. Decreased expression of hsa_circ_0003570 in hepatocellular carcinoma and its clinical significance. J Clin Lab Anal. 2018. https://doi.org/10.1002/jcla.22239.
Jiang Z, Shen L, Wang S, Wu S, Hu Y, Guo J, et al. Hsa_circ_0028502 and hsa_circ_0076251 are potential novel biomarkers for hepatocellular carcinoma. Cancer Med. 2019;8(17):7278–87.
Matboli M, Shafei AE, Ali MA, Ashry AM, Kamal KM, Agag MA, et al. circRNAs (hsa_circ_00156, hsa_circ _000224, and hsa_circ _000520) are novel potential biomarkers in hepatocellular carcinoma. J Cell Biochem. 2018. https://doi.org/10.1002/jcb.28045.
Zhang X, Xu Y, Qian Z, Zheng W, Wu Q, Chen Y, et al. circRNA_104075 stimulates YAP-dependent tumorigenesis through the regulation of HNF4a and may serve as a diagnostic marker in hepatocellular carcinoma. Cell Death Dis. 2018;9(11):1091.
Wang W, Li Y, Li X, Liu B, Han S, Li X, et al. Circular RNA circ-FOXP1 induced by SOX9 promotes hepatocellular carcinoma progression via sponging miR-875–3p and miR-421. Biomed Pharmacother. 2020;121:109517.
Zhang X, Zhou H, Jing W, Luo P, Qiu S, Liu X, et al. The circular RNA hsa_circ_0001445 regulates the proliferation and migration of hepatocellular carcinoma and may serve as a diagnostic biomarker. Dis Markers. 2018;2018:3073467.
Wei Y, Chen X, Liang C, Ling Y, Yang X, Ye X, et al. A noncoding regulatory RNAs network driven by Circ-CDYL acts specifically in the early stages hepatocellular carcinoma. Hepatology. 2020;71(1):130–47.
Li B, Liu A, Wen Y, Yang G, Zhao J, Li X, et al. The prognostic values of serum markers in hepatocellular carcinoma after invasive therapies based on real-world data. J Clin Lab Anal. 2021;35(9):e23932.
Wang X, Wang X, Li W, Zhang Q, Chen J, Chen T. Up-regulation of hsa_circ_0000517 predicts adverse prognosis of hepatocellular carcinoma. Front Oncol. 2019;9:1105.
Chen D, Zhang C, Lin J, Song X, Wang H. Screening differential circular RNA expression profiles reveal that hsa_circ_0128298 is a biomarker in the diagnosis and prognosis of hepatocellular carcinoma. Cancer Manag Res. 2018;10:1275–83.
Chen Z, Zuo X, Pu L, Zhang Y, Han G, Zhang L, et al. circLARP4 induces cellular senescence through regulating miR-761/RUNX3/p53/p21 signaling in hepatocellular carcinoma. Cancer Sci. 2019;110(2):568–81.
Wang J, Che J. CircTP63 promotes hepatocellular carcinoma progression by sponging miR-155-5p and upregulating ZBTB18. Cancer Cell Int. 2021;21(1):156.
Yang G, Wang X, Liu B, Lu Z, Xu Z, Xiu P, et al. circ-BIRC6, a circular RNA, promotes hepatocellular carcinoma progression by targeting the miR-3918/Bcl2 axis. Cell Cycle. 2019;18(9):976–89.
Huang G, Liang M, Liu H, Huang J, Li P, Wang C, et al. CircRNA hsa_circRNA_104348 promotes hepatocellular carcinoma progression through modulating miR-187-3p/RTKN2 axis and activating Wnt/β-catenin pathway. Cell Death Dis. 2020;11(12):1065.
Li S, Weng J, Song F, Li L, Xiao C, Yang W, et al. Circular RNA circZNF566 promotes hepatocellular carcinoma progression by sponging miR-4738-3p and regulating TDO2 expression. Cell Death Dis. 2020;11(6):452.
Zhu YJ, Zheng B, Luo GJ, Ma XK, Lu XY, Lin XM, et al. Circular RNAs negatively regulate cancer stem cells by physically binding FMRP against CCAR1 complex in hepatocellular carcinoma. Theranostics. 2019;9(12):3526–40.
Yu J, Xu QG, Wang ZG, Yang Y, Zhang L, Ma JZ, et al. Circular RNA cSMARCA5 inhibits growth and metastasis in hepatocellular carcinoma. J Hepatol. 2018;68(6):1214–27.
Zhang X, Qiu S, Luo P, Zhou H, Jing W, Liang C, et al. Down-regulation of hsa_circ_0001649 in hepatocellular carcinoma predicts a poor prognosis. Cancer Biomark. 2018;22(1):135–42.
Zhang PF, Wei CY, Huang XY, Peng R, Yang X, Lu JC, et al. Circular RNA circTRIM33-12 acts as the sponge of microRNA-191 to suppress hepatocellular carcinoma progression. Mol Cancer. 2019;18(1):105.
de Martel C, Maucort-Boulch D, Plummer M, Franceschi S. World-wide relative contribution of hepatitis B and C viruses in hepatocellular carcinoma. Hepatology. 2015;62(4):1190–200.
Huang XY, Huang ZL, Xu YH, Zheng Q, Chen Z, Song W, et al. Comprehensive circular RNA profiling reveals the regulatory role of the circRNA-100338/miR-141-3p pathway in hepatitis B-related hepatocellular carcinoma. Sci Rep. 2017;7(1):5428.
Zhu K, Zhan H, Peng Y, Yang L, Gao Q, Jia H, et al. Plasma hsa_circ_0027089 is a diagnostic biomarker for hepatitis B virus-related hepatocellular carcinoma. Carcinogenesis. 2020;41(3):296–302.
Cui S, Qian Z, Chen Y, Lei L, Peng L, Ding H. Screening of up- and downregulation of circRNAs in HBV-related hepatocellular carcinoma by microarray. Oncol Lett. 2018;15(1):423.
Zhou TC, Li X, Chen LJ, Fan JH, Lai X, Tang Y, et al. Differential expression profile of hepatic circular RNAs in chronic hepatitis B. J Viral Hepat. 2018. https://doi.org/10.1111/jvh.12944.
Zhang L, Wang Z. Circular RNA hsa_circ_0004812 impairs IFN-induced immune response by sponging miR-1287-5p to regulate FSTL1 in chronic hepatitis B. Virol J. 2020;17(1):40.
Fitzmaurice C, Abate D, Abbasi N, Abbastabar H, Abd-Allah F, Abdel-Rahman O, et al. Global, regional, and national cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life-years for 29 cancer groups, 1990 to 2017: a systematic analysis for the global burden of disease study. JAMA Oncol. 2019;5(12):1749–68.
Wang FS, Fan JG, Zhang Z, Gao B, Wang HY. The global burden of liver disease: the major impact of China. Hepatology. 2015;60(6):2099–108.
Goh GBB, Clarence K, Ying LS, Venkatanarasimha Nanda KK, Rafidah AB, Krishnamoorthy TL, et al. Perceptions of non-alcoholic fatty liver disease—an Asian community-based study. Gastroenterol Rep. 2015;4(2):131–5.
Zhang T, Xu J, Ye L, Lin X, Xu Y, Pan X, et al. Age, gender and geographic differences in global health burden of cirrhosis and liver cancer due to nonalcoholic steatohepatitis. J Cancer. 2021;12(10):2855–65.
Zhao Q, Liu J, Deng H, Ma R, Liao JY, Liang H, et al. Targeting mitochondria-located circRNA SCAR alleviates NASH via reducing mROS output. Cell. 2020;183(1):76-93 e22.
Jin X, Gao J, Zheng R, Yu M, Ren Y, Yan T, et al. Antagonizing circRNA_002581-miR-122-CPEB1 axis alleviates NASH through restoring PTEN-AMPK-mTOR pathway regulated autophagy. Cell Death Dis. 2020;11(2):123.
Guo XY, Chen JN, Sun F, Wang YQ, Pan Q, Fan JG. circRNA_0046367 prevents hepatoxicity of lipid peroxidation: an inhibitory role against hepatic steatosis. Oxid Med Cell Longev. 2017;2017:3960197.
Guo XY, Sun F, Chen JN, Wang YQ, Pan Q, Fan JG. circRNA_0046366 inhibits hepatocellular steatosis by normalization of PPAR signaling. World J Gastroenterol. 2018;24(3):323–37.
Kappler R, Eichenmüller M, Trippel F, Strom TM, Schweinitz DV. Abstract 3086: the genetic landscape of the childhood liver cancer hepatoblastoma. Can Res. 2014;74(19 Supplement):3086–3086.
Liu BH, Zhang BB, Liu XQ, Zheng S, Dong KR, Dong R. Expression profiling identifies circular RNA signature in hepatoblastoma. Cell Physiol Biochem. 2018;45(2):706–19.
Li X, Wang H, Liu Z, Abudureyimu A. CircSETD3 (Hsa_circ_0000567) suppresses hepatoblastoma pathogenesis via targeting the miR-423–3p/Bcl-2-interacting mediator of cell death axis. Front Genet. 2021;12:724197.
Guo W, Zhao L, Wei G, Liu P, Zhang Y, Fu L. Circ_0015756 aggravates hepatocellular carcinoma development by regulating FGFR1 via sponging miR-610. Cancer Manag Res. 2020;12:7383–94.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (No. 81872584), Key R&D and Promotion Projects of Henan Province (No. 222102310587), Key Scientific Research Project Plan of Henan Province (No. 21A330001), Natural Science Foundation of Shenzhen (No. JCYJ20210324093211030), Medical Scientific Research Foundation of Guangdong Province (No. A2020490), Science and Technology Development Plan of Kaifeng (No. 2103007) and the Interdisciplinary Research for First-class Discipline Construction Project of Henan University (No. 2019YLXKJC04).
Author information
Authors and Affiliations
Contributions
Study conception and designs: PW, YZ, and ZQ; article screening: PG, LD, LL and PW; drafting of the manuscript: PW, YZ and NL; critical revision: NL, ZQ and ZY. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, P., Zhang, Y., Deng, L. et al. The function and regulation network mechanism of circRNA in liver diseases. Cancer Cell Int 22, 141 (2022). https://doi.org/10.1186/s12935-022-02559-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-022-02559-1